
Citrus Canker Pathogen, Its Mechanism of Infection, Eradication, and Impacts
 , ,
, ,
Abstract
:1. Introduction
2. Historical Perspective
3. Pathogen Profile
Major Types of Pathovars of Xanthomonas citri
4. Types of Citrus Bacterial Canker
Pathogenic Variants of Citri Pathovar
5. Symptoms
5.1. On Leaves
5.2. On Twigs and Fruits
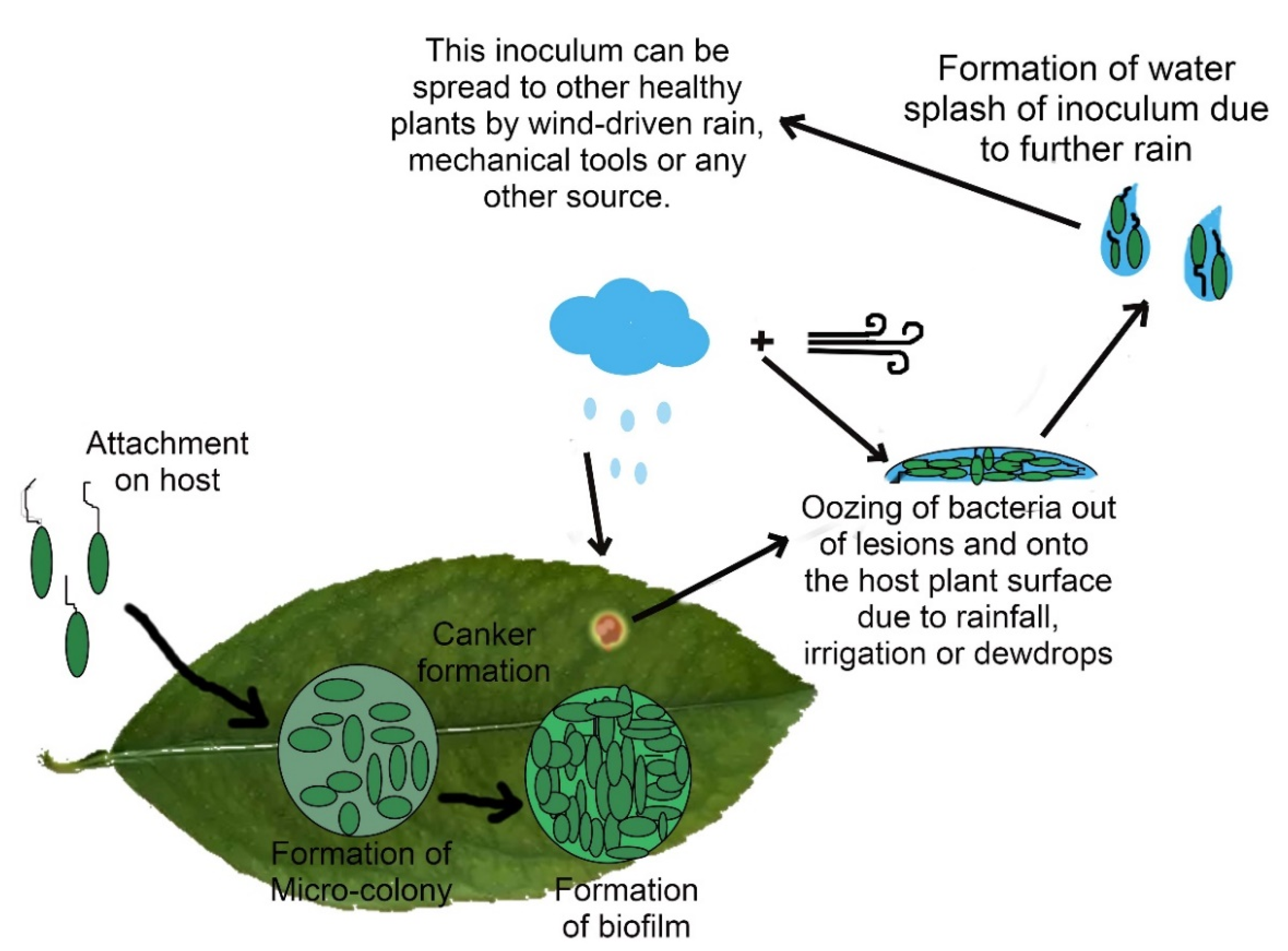
6. Pathogen’s Infection Mechanisms
6.1. Role of Adhesin Proteins in Bacterial Adhesion
6.2. Significance of Type III Protein Secretion System
6.3. Xanthan and Biofilm Formation
6.4. Damage to Host’s Machinery
7. Eradication and Control Measures
7.1. Physical Approaches
7.1.1. Tree Removal
7.1.2. Periodic Inspection of Citrus Orchards
7.1.3. Windbreaks
7.1.4. Planting Resistant Varieties
7.1.5. Pruning or Defoliation
7.1.6. Other Precautionary Measures
7.2. Chemical Approaches
7.2.1. Copper Sprays
7.2.2. Alternative Bactericides
7.2.3. Application of Antibiotics
7.2.4. Post-Harvest Sanitization
7.2.5. Systematic Acquired Resistance (SAR) Inducers
7.2.6. Chemical Control of Citrus Leaf Miner (CLM)
7.3. Biological Control
7.3.1. Genetic Engineering
7.3.2. Biological Control of CLM
7.3.3. Use of Plant Extracts
7.3.4. Endophytic Bacteria
7.3.5. Treatment with Bacteriophages
8. Conclusions and Future Prospects
Author Contributions
Funding
Conflicts of Interest
References
- Ismail, M.; Zhang, J. Post-harvest citrus diseases and their control. Outlooks Pest Manag. 2004, 15, 29. [Google Scholar] [CrossRef]
- Graham, J.H.; Gottwald, T.R.; Cubero, J.; Achor, D.S. Xanthomonas axonopodis pv. citri: Factors affecting successful eradication of citrus canker. Mol. Plant Pathol. 2004, 5, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Thakre, B.; Soni, U.; Gour, C.; Vishwakarma, R.; Jashwani, N. Field identification, eradication and current management of citrus canker caused by xanthomonas campestris pv. citriin satpura platun of madhya pradesh, india. Plant Arch. 2017, 17, 371–374. [Google Scholar]
- Jetter, K.M.; Civerolo, E.L.; Sumner, D.A. Ex ante economics of exotic disease policy: Citrus canker in California. In Exotic Pests and Diseases: Biology and Economics for Biosecurity; Wiley: Hoboken, NJ, USA, 2000; pp. 121–149. [Google Scholar]
- Jenkins, A.E.; Fawcett, H. Records of Citrus scab mainly from herbarium specimens of the genus Citrus in England and the United States. Phytopathology 1933, 23, 475–482. [Google Scholar]
- Raychaudhuri, S.; Verma, J.; Nariani, T.; Sen, B. The history of plant pathology in India. Annu. Rev. Phytopathol. 1972, 10, 21–36. [Google Scholar] [CrossRef]
- Ye, G.; Hong, N.; Zou, L.-F.; Zou, H.-S.; Zakria, M.; Wang, G.-P.; Chen, G.-Y. Tale-based genetic diversity of Chinese isolates of the citrus canker pathogen Xanthomonas citri subsp. citri. Plant Dis. 2013, 97, 1187–1194. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.A. Further data on the susceptibility of rutaceous plants to citrus-canker. J. Agric. Res. 1918, 15, 661–665. [Google Scholar]
- Das, A. Citrus canker-A review. J. Appl. Hortic. 2003, 5, 52–60. [Google Scholar] [CrossRef]
- Rossetti, V. Citrus canker in Latin America: A review. In Proceedings of the International Society of Citriculture, Orlando, FL, USA, 1–8 May 1977; pp. 918–923. [Google Scholar]
- Doidge, E.M. Citrus canker in South Africa. South Afr. Fruit Grow. 1916, 3, 265–268. [Google Scholar]
- Sharma, S.K.; Sharma, R.R. Citrus canker approaching century: A review. Tree For. Sci. Biotechnol. 2009, 2, 54–56. [Google Scholar]
- Schubert, T.S.; Miller, J. Bacterial citrus canker; Fla. Department Agric. & Consumer Services, Division of Plant Industry: Miami, FL, USA, 1996. [Google Scholar]
- Canteros, B.I.; Gochez, A.M.; Moschini, R.C. Management of citrus canker in Argentina, a success story. Plant Pathol. J. 2017, 33, 441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, E.; Kunta, M.; Ancona, V.; da Graça, J.V.; Ayin, C.; Santillana, G.; Mavrodieva, V. The Return of Asiatic Citrus Canker to Texas: Surveys and Eradication Efforts. Plant Health Prog. 2021, 22, 143–148. [Google Scholar] [CrossRef]
- Juhasz, C.; Leduc, A.; Boyer, C.; Guérin, F.; Vernière, C.; Pruvost, O.; Wonni, I.; Ouédraogo, L. First report of Xanthomonas citri pv. citri causing Asiatic citrus canker in Burkina Faso. Plant Dis. 2013, 97, 1653. [Google Scholar] [CrossRef] [PubMed]
- Gottwald, T.R.; Graham, J.H. Citrus diseases with global ramifications including citrus canker and huanglongbing. CABI Rev. 2014, 1–11. [Google Scholar] [CrossRef]
- Jabeen, T.; Arshad, H.M.I.; Saleem, K.; Ali, S.; Ullah, E.; Naureen, S.; Babar, M.M. Morphological and biochemical characterization of Xanthomonas axenopodis pv. citri isolates causing citrus canker disease in Pakistan. PSM Microbiol. 2016, 1, 10–17. [Google Scholar]
- Luthra, J.; Sattar, A. Citrus canker and its control in Punjab. Punjab Fruit J 1942, 6, 179–182. [Google Scholar]
- Bedi, K. Some important observations on the citrus canker in Punjab. Punjab Hort. J 1961, 2, 89–91. [Google Scholar]
- Gade, R.; Lad, R. Biological management of major citrus diseases in Central India-a review. Int. J. Curr. Microbiol. App. Sci 2018, 6, 296–308. [Google Scholar]
- Ference, C.M.; Gochez, A.M.; Behlau, F.; Wang, N.; Graham, J.H.; Jones, J.B. Recent advances in the understanding of Xanthomonas citri ssp. citri pathogenesis and citrus canker disease management. Mol. Plant Pathol. 2018, 19, 1302–1318. [Google Scholar] [CrossRef]
- Gabriel, D.; Kingsley, M.; Hunter, J.; Gottwald, T. Reinstatement of Xanthomonas citri (ex Hasse) and X. phaseoli (ex Smith) to species and reclassification of all X. campestris pv. citri strains. Int. J. Syst. Evol. Microbiol. 1989, 39, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Schaad, N.W.; Postnikova, E.; Lacy, G.; Sechler, A.; Agarkova, I.V.; Stromberg, P.E.; Stromberg, V.K.; Vidaver, A.M. Emended classification of xanthomonad pathogens on citrus. Syst. Appl. Microbiol. 2006, 29, 690–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vauterin, L.; Hoste, B.; Kersters, K.; Swings, J. Reclassification of xanthomonas. Int. J. Syst. Evol. Microbiol. 1995, 45, 472–489. [Google Scholar] [CrossRef] [Green Version]
- Vauterin, L.; Swings, J.; Kersters, K.; Gillis, M.; Mew, T.; Schroth, M.; Palleroni, N.; Hildebrand, D.; Stead, D.; Civerolo, E. Towards an improved taxonomy of Xanthomonas. Int. J. Syst. Evol. Microbiol. 1990, 40, 312–316. [Google Scholar] [CrossRef] [Green Version]
- Bansal, K.; Kumar, S.; Patil, P.B. Taxonomic repositioning of twelve Xanthomonas campestris, seven Xanthomonas axonopodis and one Pseudomonas cissicola reference pathovars to Xanthomonas citri. bioRxiv 2021. [Google Scholar] [CrossRef]
- Hasse, C.H. Pseudomonas citri, the cause of citrus canker. J. Agric. Res. 1915, 4, 97–104. [Google Scholar]
- Peltier, G.L.; Neal, D.C. Overwintering of the citrus-canker organism in the bark tissue of hardy citrus hybrids. J. Agric. Res. 1918, 14, 523–524. [Google Scholar]
- Moreira, L.M.; Almeida, N.F.; Potnis, N.; Digiampietri, L.A.; Adi, S.S.; Bortolossi, J.C.; da Silva, A.C.; da Silva, A.M.; de Moraes, F.E.; de Oliveira, J.C. Novel insights into the genomic basis of citrus canker based on the genome sequences of two strains of Xanthomonas fuscans subsp. aurantifolii. BMC Genom. 2010, 11, 238. [Google Scholar] [CrossRef]
- Fonseca, N.P.; Patané, J.S.L.; Varani, A.M.; Felestrino, É.B.; Caneschi, W.L.; Sanchez, A.B.; Cordeiro, I.F.; Lemes, C.G.d.C.; Assis, R.d.A.B.; Garcia, C.C.M.; et al. Analyses of Seven New Genomes of Xanthomonas citri pv. aurantifolii Strains, Causative Agents of Citrus Canker B and C, Show a Reduced Repertoire of Pathogenicity-Related Genes. Front. Microbiol. 2019, 10, 2361. [Google Scholar] [CrossRef]
- Vauterin, L.; Rademaker, J.; Swings, J. Synopsis on the taxonomy of the genus Xanthomonas. Phytopathology 2000, 90, 677–682. [Google Scholar] [CrossRef] [Green Version]
- Young, J.; Allen, C.; Coutinho, T.; Denny, T.; Elphinstone, J.; Fegan, M.; Gillings, M.; Gottwald, T.; Graham, J.; Iacobellis, N. Plant-pathogenic bacteria as biological weapons–real threats? Phytopathology 2008, 98, 1060–1065. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Stall, R.E.; Jones, J.B.; Cubero, J.; Gottwald, T.R.; Graham, J.H.; Dixon, W.N.; Schubert, T.S.; Chaloux, P.H.; Stromberg, V.K. Detection and characterization of a new strain of citrus canker bacteria from Key/Mexican lime and alemow in South Florida. Plant Dis. 2004, 88, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Vauterin, L.; Yang, P.; Hoste, B.; Vancanneyt, M.; Civerolo, E.; Swings, J.; Kersters, K. Differentiation of Xanthomonas campestris pv. citri strains by sodium dodecyl sulfate-polyacrylamide gel electrophoresis of proteins, fatty acid analysis, and DNA-DNA hybridization. Int. J. Syst. Evol. Microbiol. 1991, 41, 535–542. [Google Scholar] [CrossRef] [Green Version]
- Mendonça, L.; Zambolim, L.; Badel, J. Bacterial citrus diseases: Major threats and recent progress. J. Bacteriol. Mycol. Open Access 2017, 5, 4–2017. [Google Scholar]
- Gottwald, T.R.; Graham, J.H.; Schubert, T.S. Citrus canker: The pathogen and its impact. Plant Health Prog. 2002, 3, 15. [Google Scholar] [CrossRef] [Green Version]
- Jalan, N.; Aritua, V.; Kumar, D.; Yu, F.; Jones, J.B.; Graham, J.H.; Setubal, J.C.; Wang, N. Comparative Genomic Analysis of Xanthomonas axonopodis pv. citrumelo F1, Which Causes Citrus Bacterial Spot Disease, and Related Strains Provides Insights into Virulence and Host Specificit. J. Bacteriol. 2011, 193, 6342–6357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vernière, C.; Hartung, J.; Pruvost, O.; Civerolo, E.; Alvarez, A.M.; Maestri, P.; Luisetti, J. Characterization of phenotypically distinct strains of Xanthomonas axonopodis pv. citri from Southwest Asia. Eur. J. Plant Pathol. 1998, 104, 477–487. [Google Scholar] [CrossRef]
- Urrutia, M. Isolation, pathogenicity, and partial host range of Alternaria limicola, causal agent of mancha foliar de los citricos in Mexico. Plant Dis. 1994, 78, 879. [Google Scholar]
- Egel, D.; Graham, J.; Stall, R. Genomic relatedness of Xanthomonas campestris strains causing diseases of citrus. Appl. Environ. Microbiol. 1991, 57, 2724–2730. [Google Scholar] [CrossRef] [Green Version]
- Rybak, M.; Minsavage, G.V.; Stall, R.E.; Jones, J.B. Identification of Xanthomonas citri ssp. citri host specificity genes in a heterologous expression host. Mol. Plant Pathol. 2009, 10, 249–262. [Google Scholar] [CrossRef]
- Bouzar, H.; Jones, J.; Stall, R.; Hodge, N.; Minsavage, G.; Benedict, A.; Alvarez, A. Physiological, chemical, serological, and pathogenic analyses of a worldwide collection of Xanthomonas campestris pv. vesicatoria strains. Phytopathology 1994, 84, 663–671. [Google Scholar] [CrossRef]
- Alvarez, A.; Benedict, A.; Mizumoto, C.; Pollard, L.; Civerolo, E. Analysis of Xanthomonas campestris pv. citri and X. c. citrumelo with monoclonal antibodies. Phytopathology 1991, 81, 857–865. [Google Scholar] [CrossRef]
- Stall, R. Xanthomonas campestris pv. citri detection and identification by enzyme-linked immunosorbent assay. Plant Dis 1982, 231, 231–236. [Google Scholar]
- Stall, R.E.; Miller, J.; Marco, G.; Canteros de Echenique, B. Pathogenicity of three strains of citrus canker organism on grapefruit. In Proceedings of the Fifth International Conference on Plant Pathogenic Bacteria, Cali, Colombia, 16–23 August 1981. [Google Scholar]
- Pruvost, O.; Hartung, J.; Civerolo, E.; Dubois, C.; Perrier, X. Plasmid DNA fingerprints distinguish pathotypes of Xanthomonas campestris pv. citri, the causal agent of citrus bacterial canker disease. Phytopathology 1992, 82, 485–490. [Google Scholar] [CrossRef]
- Gordon, J.L.; Lefeuvre, P.; Escalon, A.; Barbe, V.; Cruveiller, S.; Gagnevin, L.; Pruvost, O. Comparative genomics of 43 strains of Xanthomonas citri pv. citri reveals the evolutionary events giving rise to pathotypes with different host ranges. BMC Genom. 2015, 16, 1098. [Google Scholar] [CrossRef]
- Civerolo, E. Bacterial canker disease of citrus [Xanthomonas campestris]. J. Rio Gd. Val. Hortic. Soc. 1984, 37, 127–146. [Google Scholar]
- Gottwald, T.R. Citrus canker and citrus huanglongbing, two exotic bacterial diseases threatening the citrus industries of the Western Hemisphere. Outlooks Pest Manag. 2007, 18, 274. [Google Scholar] [CrossRef]
- Jalan, N.; Kumar, D.; Andrade, M.O.; Yu, F.; Jones, J.B.; Graham, J.H.; White, F.F.; Setubal, J.C.; Wang, N. Comparative genomic and transcriptome analyses of pathotypes of Xanthomonas citri subsp. citri provide insights into mechanisms of bacterial virulence and host range. BMC Genom. 2013, 14, 551. [Google Scholar] [CrossRef] [Green Version]
- Swarup, S.; Yang, Y.; Kingsley, M.T.; Gabriel, D.W. An Xanthomonas citri pathogenicity gene, pthA, pleiotropically encodes gratuitous avirulence on nonhosts. Mol. Plant-Microbe Interact 1992, 5, 204–213. [Google Scholar] [CrossRef]
- Cubero, J.; Graham, J. Genetic relationship among worldwide strains of Xanthomonas causing canker in citrus species and design of new primers for their identification by PCR. Appl. Environ. Microbiol. 2002, 68, 1257–1264. [Google Scholar] [CrossRef] [Green Version]
- Schaad, N.W.; Jones, J.B.; Chun, W. Laboratory Guide for the Identification of Plant Pathogenic Bacteria; American Phytopathological Society: Saint Paul, MN, USA, 2001. [Google Scholar]
- Schubert, T.S.; Rizvi, S.A.; Sun, X.; Gottwald, T.R.; Graham, J.H.; Dixon, W.N. Meeting the challenge of eradicating citrus canker in Florida—Again. Plant Dis. 2001, 85, 340–356. [Google Scholar] [CrossRef] [Green Version]
- Bui Thi Ngoc, L.; Vernière, C.; Jarne, P.; Brisse, S.; Guérin, F.; Boutry, S.; Gagnevin, L.; Pruvost, O. From local surveys to global surveillance: Three high-throughput genotyping methods for epidemiological monitoring of Xanthomonas citri pv. citri pathotypes. Appl. Environ. Microbiol. 2009, 75, 1173–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patané, J.S.L.; Martins, J.; Rangel, L.T.; Belasque, J.; Digiampietri, L.A.; Facincani, A.P.; Ferreira, R.M.; Jaciani, F.J.; Zhang, Y.; Varani, A.M.; et al. Origin and diversification of Xanthomonas citri subsp. citri pathotypes revealed by inclusive phylogenomic, dating, and biogeographic analyses. BMC Genom. 2019, 20, 700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Jalan, N.; Zhou, X.; Goss, E.; Jones, J.B.; Setubal, J.C.; Deng, X.; Wang, N. Positive selection is the main driving force for evolution of citrus canker-causing Xanthomonas. ISME J. 2015, 9, 2128–2138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escalon, A.; Javegny, S.; Vernière, C.; Noël, L.D.; Vital, K.; Poussier, S.; Hajri, A.; Boureau, T.; Pruvost, O.; Arlat, M. Variations in type III effector repertoires, pathological phenotypes and host range of Xanthomonas citri pv. citri pathotypes. Mol. Plant Pathol. 2013, 14, 483–496. [Google Scholar] [CrossRef] [Green Version]
- Webster, J.; Bogema, D.; Chapman, T.A. Comparative Genomics of Xanthomonas citri pv. citri A* Pathotype Reveals Three Distinct Clades with Varying Plasmid Distribution. Microorganisms 2020, 8, 1947. [Google Scholar] [CrossRef]
- Bansal, K.; Midha, S.; Kumar, S.; Patil, P.B. Ecological and evolutionary insights into Xanthomonas citri pathovar diversity. Appl. Environ. Microbiol. 2017, 83, e02993-16. [Google Scholar] [CrossRef] [Green Version]
- Koizumi, M. Citrus canker: The world situation. In Citrus Canker: An International Perspective; Timmer, L.W., Ed.; University of Florida: Gainesville, FL, USA, 1985; pp. 2–7. [Google Scholar]
- Loucks, K.W. Citrus Canker and Its Eradication in Florida; Department of Agriculture, Division of Plant Industry, Florida Department of Agriculture and Consumer Services: Gainesville, FL, USA, 1934. [Google Scholar]
- Brunings, A.M.; Gabriel, D.W. Xanthomonas citri: Breaking the surface. Mol. Plant Pathol. 2003, 4, 141–157. [Google Scholar] [CrossRef]
- Van Sluys, M.; Monteiro-Vitorello, C.; Camargo, L.; Menck, C.; Da Silva, A.; Ferro, J.; Oliveira, M.; Setubal, J.; Kitajima, J.; Simpson, A. Comparative genomic analysis of plant-associated bacteria. Annu. Rev. Phytopathol. 2002, 40, 169–189. [Google Scholar] [CrossRef]
- Gottig, N.; Garavaglia, B.; Garofalo, C.; Zimaro, T.; Sgro, G.; Ficarra, F.; Dunger, G.; Daurelio, L.; Thomas, L.; Gehring, C. Mechanisms of infection used by Xanthomonas axonopodis pv. citri in citrus canker disease. Curr. Res. Technol. Educ. Top. Appl. Microbiol. Microb. Biotechnol. 2010, 1, 196–204. [Google Scholar]
- Henderson, I.R.; Navarro-Garcia, F.; Desvaux, M.; Fernandez, R.C.; Ala’Aldeen, D. Type V protein secretion pathway: The autotransporter story. Microbiol. Mol. Biol. Rev. 2004, 68, 692–744. [Google Scholar] [CrossRef] [Green Version]
- Hodak, H.; Clantin, B.; Willery, E.; Villeret, V.; Locht, C.; Jacob-Dubuisson, F. Secretion signal of the filamentous haemagglutinin, a model two-partner secretion substrate. Mol. Microbiol. 2006, 61, 368–382. [Google Scholar] [CrossRef] [PubMed]
- Gottig, N.; Garavaglia, B.S.; Garofalo, C.G.; Orellano, E.G.; Ottado, J. A filamentous hemagglutinin-like protein of Xanthomonas axonopodis pv. citri, the phytopathogen responsible for citrus canker, is involved in bacterial virulence. PLoS ONE 2009, 4, e4358. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gómez, L.; Boller, T. Flagellin perception: A paradigm for innate immunity. Trends Plant Sci. 2002, 7, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Alfano, J.R.; Collmer, A. The type III (Hrp) secretion pathway of plant pathogenic bacteria: Trafficking harpins, Avr proteins, and death. J. Bacteriol. 1997, 179, 5655–5662. [Google Scholar] [CrossRef] [Green Version]
- Büttner, D.; Bonas, U. Getting across—Bacterial type III effector proteins on their way to the plant cell. EMBO J. 2002, 21, 5313–5322. [Google Scholar] [CrossRef]
- Morel, J.-B.; Dangl, J.L. The hypersensitive response and the induction of cell death in plants. Cell Death Differ. 1997, 4, 671–683. [Google Scholar] [CrossRef] [Green Version]
- Tampakaki, A.P.; Fadouloglou, V.E.; Gazi, A.; Panopoulos, N.; Kokkinidis, M. Conserved features of type III secretion. Cell. Microbiol. 2004, 6, 805–816. [Google Scholar] [CrossRef]
- Hajri, A.; Brin, C.; Hunault, G.; Lardeux, F.; Lemaire, C.; Manceau, C.; Boureau, T.; Poussier, S. A «repertoire for repertoire» hypothesis: Repertoires of type three effectors are candidate determinants of host specificity in Xanthomonas. PLoS ONE 2009, 4, e6632. [Google Scholar] [CrossRef]
- Duan, Y.P.; Castaneda, A.; Zhao, G.; Erdos, G.; Gabriel, D. Expression of a single, host-specific, bacterial pathogenicity gene in plant cells elicits division, enlargement, and cell death. Mol. Plant-Microbe Interact. 1999, 12, 556–560. [Google Scholar] [CrossRef] [Green Version]
- Dunger, G.; Arabolaza, A.; Gottig, N.; Orellano, E.; Ottado, J. Participation of Xanthomonas axonopodis pv. citri hrp cluster in citrus canker and nonhost plant responses. Plant Pathol. 2005, 54, 781–788. [Google Scholar] [CrossRef]
- Rigano, L.A.; Siciliano, F.; Enrique, R.; Sendín, L.; Filippone, P.; Torres, P.S.; Qüesta, J.; Dow, J.M.; Castagnaro, A.P.; Vojnov, A.A. Biofilm formation, epiphytic fitness, and canker development in Xanthomonas axonopodis pv. citri. Mol. Plant-Microbe Interact. 2007, 20, 1222–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunger, G.; Relling, V.M.; Tondo, M.L.; Barreras, M.; Ielpi, L.; Orellano, E.G.; Ottado, J. Xanthan is not essential for pathogenicity in citrus canker but contributes to Xanthomonas epiphytic survival. Arch. Microbiol. 2007, 188, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.-P.; Lu, K.-M.; Chen, Y.-H. A novel two-component response regulator links rpf with biofilm formation and virulence of Xanthomonas axonopodis pv. citri. PLoS ONE 2013, 8, e62824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.-L.; Liu, Y.-N.; Barber, C.; Dow, J.; Wootton, J.; Daniels, M. Genetic and molecular analysis of a cluster of rpf genes involved in positive regulation of synthesis of extracellular enzymes and polysaccharide in Xanthomonas campestris pathovar campestris. Mol. Gen. Genet. MGG 1991, 226, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Rigano, L.A.; Marano, M.R.; Castagnaro, A.P.; Do Amaral, A.M.; Vojnov, A.A. Rapid and sensitive detection of citrus bacterial canker by loop-mediated isothermal amplification combined with simple visual evaluation methods. BMC Microbiol. 2010, 10, 176. [Google Scholar] [CrossRef]
- Nembaware, V.; Seoighe, C.; Sayed, M.; Gehring, C. A plant natriuretic peptide-like gene in the bacterial pathogen Xanthomonas axonopodis may induce hyper-hydration in the plant host: A hypothesis of molecular mimicry. BMC Evol. Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Gottig, N.; Garavaglia, B.S.; Daurelio, L.D.; Valentine, A.; Gehring, C.; Orellano, E.G.; Ottado, J. Xanthomonas axonopodis pv. citri uses a plant natriuretic peptide-like protein to modify host homeostasis. Proc. Natl. Acad. Sci. USA 2008, 105, 18631–18636. [Google Scholar] [CrossRef] [Green Version]
- Takei, Y. Does the natriuretic peptide system exist throughout the animal and plant kingdom? Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2001, 129, 559–573. [Google Scholar] [CrossRef]
- Maryani, M.M.; Morse, M.V.; Bradley, G.; Irving, H.R.; Cahill, D.M.; Gehring, C.A. In situ localization associates biologically active plant natriuretic peptide immuno-analogues with conductive tissue and stomata. J. Exp. Bot. 2003, 54, 1553–1564. [Google Scholar] [CrossRef]
- Meier, S.; Bastian, R.; Donaldson, L.; Murray, S.; Bajic, V.; Gehring, C. Co-expression and promoter content analyses assign a role in biotic and abiotic stress responses to plant natriuretic peptides. BMC Plant Biol. 2008, 8, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottig, N.; Garavaglia, B.S.; Daurelio, L.D.; Valentine, A.; Gehring, C.; Orellano, E.G.; Ottado, J. Modulating host homeostasis as a strategy in the plant-pathogen arms race. Commun. Integr. Biol. 2009, 2, 89–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garavaglia, B.S.; Thomas, L.; Gottig, N.; Dunger, G.; Garofalo, C.G.; Daurelio, L.D.; Ndimba, B.; Orellano, E.G.; Gehring, C.; Ottado, J. A eukaryotic-acquired gene by a biotrophic phytopathogen allows prolonged survival on the host by counteracting the shut-down of plant photosynthesis. PLoS ONE 2010, 5, e8950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cernadas, R.A.; Benedetti, C.E. Role of auxin and gibberellin in citrus canker development and in the transcriptional control of cell-wall remodeling genes modulated by Xanthomonas axonopodis pv. citri. Plant Sci. 2009, 177, 190–195. [Google Scholar] [CrossRef]
- Pereira, A.L.; Carazzolle, M.F.; Abe, V.Y.; de Oliveira, M.L.; Domingues, M.N.; Silva, J.C.; Cernadas, R.A.; Benedetti, C.E. Identification of putative TAL effector targets of the citrus canker pathogens shows functional convergence underlying disease development and defense response. BMC Genom. 2014, 15, 157. [Google Scholar] [CrossRef] [Green Version]
- Behlau, F.; Fonseca, A.; Belasque Jr, J. A comprehensive analysis of the Asiatic citrus canker eradication programme in São Paulo state, Brazil, from 1999 to 2009. Plant Pathol. 2016, 65, 1390–1399. [Google Scholar] [CrossRef]
- Leite Jr, R.; Mohan, S. Integrated management of the citrus bacterial canker disease caused by Xanthomonas campestris pv. citri in the State of Paraná, Brazil. Crop Prot. 1990, 9, 3–7. [Google Scholar] [CrossRef]
- Graham, J.H.; Dewdney, M.M.; Myers, M.E. Streptomycin and copper formulations for control of citrus canker on grapefruit. In Proceedings of the Florida State Horticultural Society, Crystal River, FL, USA, 6–8 June 2010; pp. 92–99. [Google Scholar]
- Gottwald, T.R.; Hughes, G.; Graham, J.H.; Sun, X.; Riley, T. The citrus canker epidemic in Florida: The scientific basis of regulatory eradication policy for an invasive species. Phytopathology 2001, 91, 30–34. [Google Scholar] [CrossRef] [Green Version]
- Zansler, M.; Spreen, T.; Muraro, R. Florida’s Citrus Canker Eradication Program (CCEP): Benefit–Costs Analysis, EDIS document FE531; University of Florida: Gainesville, FL, USA, 2005. [Google Scholar]
- Dewdney, M.; Graham, J.; Rogers, M. Citrus canker. In Florida Citrus Pest Management Guide, SP-43; University of Florida: Gainesville, FL, USA, 2016; pp. 93–96. [Google Scholar]
- Gottwald, T.; Timmer, L. The efficacy of windbreaks in reducing the spread of citrus canker caused by Xanthomonas campestris pv. citri. Trop. Agric. 1995, 72, 194–201. [Google Scholar]
- Timmer, L.W.; Graham, J.H.; Chamberlain, H.L. Fundamentals of Citrus Canker Management; EDIS: Gainesville, FL, USA; University of Florida: Gainesville, FL, USA, 2006; pp. 1–5. [Google Scholar]
- Behlau, F.; Belasque Jr, J.; Bergamin Filho, A.; Graham, J.; Leite, R., Jr.; Gottwald, T. Copper sprays and windbreaks for control of citrus canker on young orange trees in southern Brazil. Crop Prot. 2008, 27, 807–813. [Google Scholar] [CrossRef]
- Tinnner, L.W. Evaluation of bactericides for control of citrus canker in Argentina. In Proceedings of the Florida State Horticultural Society, Tampa, FL, USA, 7 June 1988; pp. 6–9. [Google Scholar]
- Patel, M.; Padhya, A. Sodium arsenite-Copper sulphate spray for the control of citrus canker. Curr. Sci. 1964, 33, 87–88. [Google Scholar]
- Graham, J.; Dewdney, M. Research progress on management of Citrus Canker. Citrus Indus. 2012, 93, 15–17. [Google Scholar]
- Bock, C.; Graham, J.; Gottwald, T.; Cook, A.; Parker, P. Wind speed effects on the quantity of Xanthomonas citri subsp. citri dispersed downwind from canopies of grapefruit trees infected with citrus canker. Plant Dis. 2010, 94, 725–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottwald, T.R.; Graham, J.H. A device for precise and nondisruptive stomatal inoculation of leaf tissue with bacterial pathogens. Phytopathology 1992, 82, 930–935. [Google Scholar] [CrossRef]
- Canteros, B. Citrus canker in Argentina-control, eradication and current management. In Proceedings of the International Citrus Canker Research Workshop, Orlando, FL, USA, 7 November 2005; pp. 20–22. [Google Scholar]
- Alva, A.; Graham, J.; Anderson, C. Soil pH and copper effects on young ‘Hamlin’orange trees. Soil Sci. Soc. Am. J. 1995, 59, 481–487. [Google Scholar] [CrossRef]
- da Silva Scapin, M.; Behlau, F.; Scandelai, L.H.M.; Fernandes, R.S.; Junior, G.J.S.; Ramos, H.H. Tree-row-volume-based sprays of copper bactericide for control of citrus canker. Crop Prot. 2015, 77, 119–126. [Google Scholar] [CrossRef]
- Graham, J.; Johnson, E.; Myers, M.; Young, M.; Rajasekaran, P.; Das, S.; Santra, S. Potential of nano-formulated zinc oxide for control of citrus canker on grapefruit trees. Plant Dis. 2016, 100, 2442–2447. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.L.; Campos, M.G.; Ozcan, A.; Mendis, H.C.; Young, M.; Myers, M.E.; Atilola, M.; Doomra, M.; Thwin, Z.; Johnson, E.G. Multifunctional Surface, Subsurface, and Systemic Therapeutic (MS3T) Formulation for the Control of Citrus Canker. J. Agric. Food Chem. 2021, 69, 10807–10818. [Google Scholar] [CrossRef]
- Caccalano, M.; Dilarri, G.; Zamuner, C.; Domingues, D.; Ferreira, H. Hexanoic acid: A new potential substitute for copper-based agrochemicals against citrus canker. J. Appl. Microbiol. 2021, 131, 2488–2499. [Google Scholar] [CrossRef]
- Cavalca, L.B.; Zamuner, C.F.; Saldanha, L.L.; Polaquini, C.R.; Regasini, L.O.; Behlau, F.; Ferreira, H. Hexyl gallate for the control of citrus canker caused by Xanthomonas citri subsp citri. MicrobiologyOpen 2020, 9, e1104. [Google Scholar] [CrossRef]
- Graham, J.H.; Gottwald, T.R.; Leite, R.P. Prospects for Control of Citrus Canker with Novel Chemical Compounds; Florida State Horticultural Society: Orlando, FL, USA, 2006; pp. 82–88. [Google Scholar]
- Behlau, F.; Jones, J.B.; Myers, M.E.; Graham, J.H. Monitoring for resistant populations of Xanthomonas citri subsp. citri and epiphytic bacteria on citrus trees treated with copper or streptomycin using a new semi-selective medium. Eur. J. Plant Pathol. 2012, 132, 259–270. [Google Scholar] [CrossRef]
- Behlau, F.; Paloschi, A.; Marin, T.G.; Santos, T.A.; Ferreira, H.; do Nascimento, L.M. Chlorine dioxide, peroxyacetic acid, and calcium oxychloride for post-harvest decontamination of citrus fruit against Xanthomonas citri subsp. citri, causal agent of citrus canker. Crop Prot. 2021, 146, 105679. [Google Scholar] [CrossRef]
- Gorlach, J.; Volrath, S.; Knauf-Beiter, G.; Hengy, G.; Beckhove, U.; Kogel, K.; Oostendorp, M.; Staub, T.; Ward, E.; Kessmann, H. Benzothiadiazole, a novel class of inducers of systemic acquired resistance, activates gene expression and dis-ease resistance in wheat. Plant Cell 1996, 8, 629–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, C.; Mou, Z. Salicylic acid and its function in plant immunity F. J. Integr. Plant Biol. 2011, 53, 412–428. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, S.; Ding, P.; Wang, D.; Cheng, Y.T.; He, J.; Gao, M.; Xu, F.; Li, Y.; Zhu, Z. Control of salicylic acid synthesis and systemic acquired resistance by two members of a plant-specific family of transcription factors. Proc. Natl. Acad. Sci. USA 2010, 107, 18220–18225. [Google Scholar] [CrossRef] [Green Version]
- Graham, J.H.; Myers, M.E. Evaluation of soil applied systemic acquired resistance inducers integrated with copper bactericide sprays for control of citrus canker on bearing grapefruit trees. Crop Prot. 2016, 90, 157–162. [Google Scholar] [CrossRef]
- Graham, J.H.; Myers, M.E. Integration of soil applied neonicotinoid insecticides and acibenzolar-S-methyl for systemic acquired resistance (SAR) control of citrus canker on young citrus trees. Crop Prot. 2013, 54, 239–243. [Google Scholar] [CrossRef]
- Powell, C.A.; Burton, M.S.; Pelosi, R.; Ritenour, M.A.; Bullock, R.C. Seasonal abundance and insecticidal control of citrus leafminer in a citrus orchard. HortScience 2007, 42, 1636–1638. [Google Scholar] [CrossRef] [Green Version]
- Stein, B.; Ramallo, J.; Foguet, L.; Graham, J.H. Citrus Leafminer Control and Copper Sprays for Management of Citrus Canker on Lemon in Tucuman, Argentina; Florida State Horticultural Society: Palm Beach Gardens, FL, USA, 2007; pp. 127–131. [Google Scholar]
- Stelinski, L.; Miller, J.; Rogers, M. Mating disruption of citrus leafminer mediated by a noncompetitive mechanism at a remarkably low pheromone release rate. J. Chem. Ecol. 2008, 34, 1107–1113. [Google Scholar] [CrossRef]
- Stelinski, L.L.; Czokajlo, D. Suppression of citrus leafminer, Phyllocnistis citrella, with an attract-and-kill formulation. Entomol. Exp. Et Appl. 2010, 134, 69–77. [Google Scholar] [CrossRef]
- Hao, G.; Pitino, M.; Duan, Y.; Stover, E. Reduced susceptibility to Xanthomonas citri in transgenic citrus expressing the FLS2 receptor from Nicotiana benthamiana. Mol. Plant-Microbe Interact. 2016, 29, 132–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Düring, K. Genetic engineering for resistance to bacteria in transgenic plants by introduction of foreign genes. Mol. Breed. 1996, 2, 297–305. [Google Scholar] [CrossRef]
- Castro, M.S.; Fontes, W. Plant defense and antimicrobial peptides. Protein Pept. Lett. 2005, 12, 11–16. [Google Scholar] [CrossRef]
- Pelegrini, P.B.; Franco, O.L. Plant γ-thionins: Novel insights on the mechanism of action of a multi-functional class of defense proteins. Int. J. Biochem. Cell Biol. 2005, 37, 2239–2253. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Rodríguez, J.J.; Ochoa-Zarzosa, A.; López-Gómez, R.; López-Meza, J.E. Plant antimicrobial peptides as potential anticancer agents. BioMed Res. Int. 2015, 2015, 735087. [Google Scholar] [CrossRef] [Green Version]
- Peng, A.; Zhang, J.; Zou, X.; He, Y.; Xu, L.; Lei, T.; Yao, L.; Li, Q.; Chen, S. Pyramiding the antimicrobial PR1aCB and AATCB genes in ‘Tarocco’blood orange (Citrus sinensis Osbeck) to enhance citrus canker resistance. Transgenic Res. 2021, 30, 635–647. [Google Scholar] [CrossRef]
- He, Y.; Chen, S.; Peng, A.; Zou, X.; Xu, L.; Lei, T.; Liu, X.; Yao, L. Production and evaluation of transgenic sweet orange (Citrus sinensis Osbeck) containing bivalent antibacterial peptide genes (Shiva A and Cecropin B) via a novel Agrobacterium-mediated transformation of mature axillary buds. Sci. Hortic. 2011, 128, 99–107. [Google Scholar] [CrossRef]
- Boscariol, R.L.; Monteiro, M.; Takahashi, E.K.; Chabregas, S.M.; Vieira, M.L.C.; Vieira, L.G.; Pereira, L.F.; de AA Mourão Filho, F.; Cardoso, S.C.; Christiano, R.S. Attacin A Gene from Tricloplusia ni Reduces Susceptibility to Xanthomonas axonopodis pv. citri in Transgenic Citrus sinensisHamlin’. J. Am. Soc. Hortic. Sci. 2006, 131, 530–536. [Google Scholar] [CrossRef] [Green Version]
- Reynoird, J.; Mourgues, F.; Norelli, J.; Aldwinckle, H.; Brisset, M.-N.; Chevreau, E. First evidence for improved resistance to fire blight in transgenic pear expressing the attacin E gene from Hyalophora cecropia. Plant Sci. 1999, 149, 23–31. [Google Scholar] [CrossRef]
- Furman, N.; Kobayashi, K.; Zanek, M.C.; Calcagno, J.; Garcia, M.L.; Mentaberry, A. Transgenic sweet orange plants expressing a dermaseptin coding sequence show reduced symptoms of citrus canker disease. J. Biotechnol. 2013, 167, 412–419. [Google Scholar] [CrossRef]
- Zhang, X.; Francis, M.I.; Dawson, W.O.; Graham, J.H.; Orbović, V.; Triplett, E.W.; Mou, Z. Over-expression of the Arabidopsis NPR1 gene in citrus increases resistance to citrus canker. Eur. J. Plant Pathol. 2010, 128, 91–100. [Google Scholar] [CrossRef]
- de Oliveira, M.L.P.; de Lima Silva, C.C.; Abe, V.Y.; Costa, M.G.C.; Cernadas, R.A.; Benedetti, C.E. Increased resistance against citrus canker mediated by a citrus mitogen-activated protein kinase. Mol. Plant-Microbe Interact. 2013, 26, 1190–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes, B.M.J.; Cardoso, S.; Boscariol-Camargo, R.; Cruz, R.; Mourão Filho, F.; Bergamin Filho, A. Reduction in susceptibility to Xanthomonas axonopodis pv. citri in transgenic Citrus sinensis expressing the rice Xa21 gene. Plant Pathol. 2010, 59, 68–75. [Google Scholar] [CrossRef]
- Omar, A.A.; Murata, M.M.; El-Shamy, H.A.; Graham, J.H.; Grosser, J.W. Enhanced resistance to citrus canker in transgenic mandarin expressing Xa21 from rice. Transgenic Res. 2018, 27, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.-Z.; Chen, C.-W.; Wang, Y.; Liu, J.-H.; Moriguchi, T. Ectopic expression of MdSPDS1 in sweet orange (Citrus sinensis Osbeck) reduces canker susceptibility: Involvement of H 2 O 2 production and transcriptional alteration. BMC Plant Biol. 2011, 11, 55. [Google Scholar] [CrossRef] [Green Version]
- Yoda, H.; Fujimura, K.; Takahashi, H.; Munemura, I.; Uchimiya, H.; Sano, H. Polyamines as a common source of hydrogen peroxide in host-and nonhost hypersensitive response during pathogen infection. Plant Mol. Biol. 2009, 70, 103–112. [Google Scholar] [CrossRef]
- Sun, L.; Ke, F.; Nie, Z.; Wang, P.; Xu, J. Citrus genetic engineering for disease resistance: Past, present and future. Int. J. Mol. Sci. 2019, 20, 5256. [Google Scholar] [CrossRef] [Green Version]
- Peng, A.; Chen, S.; Lei, T.; Xu, L.; He, Y.; Wu, L.; Yao, L.; Zou, X. Engineering canker-resistant plants through CRISPR/Cas9-targeted editing of the susceptibility gene Cs LOB 1 promoter in citrus. Plant Biotechnol. J. 2017, 15, 1509–1519. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Zhang, Y.; Orbović, V.; Xu, J.; White, F.F.; Jones, J.B.; Wang, N. Genome editing of the disease susceptibility gene Cs LOB 1 in citrus confers resistance to citrus canker. Plant Biotechnol. J. 2017, 15, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, S.; Peng, A.; Xie, Z.; He, Y.; Zou, X. CRISPR/Cas9-mediated editing of CsWRKY22 reduces susceptibility to Xanthomonas citri subsp. citri in Wanjincheng orange (Citrus sinensis (L.) Osbeck). Plant Biotechnol. Rep. 2019, 13, 501–510. [Google Scholar] [CrossRef]
- Parajuli, S.; Huo, H.; Gmitter, F.G.; Duan, Y.; Luo, F.; Deng, Z. Editing the CsDMR6 gene in citrus results in resistance to the bacterial disease citrus canker. Hortic. Res. 2022, 9, uhac082. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Orbović, V.; Wang, N. CRISPR-LbCas12a-mediated modification of citrus. Plant Biotechnol. J. 2019, 17, 1928–1937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoy, M.A.; Jessey, C. Ageniaspis citricola (Hymenoptera: Encyrtidae) established in Bermuda. Fla. Entomol. 2004, 87, 229–230. [Google Scholar] [CrossRef]
- Johnson, S.J.; Henne, D.C.; Bourgeois, W.J. Biological control of the citrus leafminer with Ageniaspis citricola (Hymenoptera: Encyrtidae) in Louisiana; Florida State Horticultural Society: Lake Beuna, FL, USA, 2003; pp. 224–226. [Google Scholar]
- Abdullahi, A.; Tijjani, A.; Abubakar, A.; Khairulmazmi, A.; Ismail, M. Plant biomolecule antimicrobials: An alternative control measures for food security and safety. In Herbal Biomolecules in Healthcare Applications; Elsevier: Amsterdam, The Netherlands, 2022; pp. 381–406. [Google Scholar]
- Tahir, H.A.; Sahi, S.T.; Habib, A.; Haq, I.U.; Ahmad, A.; Ashraf, W. Evaluation of plant extracts as biocontrol agents against Xanthomonas axonopodis pv citri the cause of citrus canker. Pak. J. Phytopathol. 2016, 28, 35–43. [Google Scholar]
- Leksomboon, C.; Thaveechai, N.; Kositratana, W. Potential of plant extracts for controlling citrus canker of lime. Agric. Nat. Resour. 2001, 35, 392–396. [Google Scholar]
- Quispel, A. A search for signals in endophytic microorganisms. In Molecular Signals in Plant-Microbe Communications; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]
- Sturz, A.; Christie, B.; Nowak, J. Bacterial endophytes: Potential role in developing sustainable systems of crop production. Crit. Rev. Plant Sci. 2000, 19, 1–30. [Google Scholar] [CrossRef]
- Tan, X.; Huang, S.; Ren, J.; Yan, W.; Cen, Z. Characterization of an endophytic bacterium strain Bc51 suppressing citrus canker. Acta Phytopathol. Sin. 2007, 37, 9–17. [Google Scholar]
- Rabbee, M.F.; Ali, M.S.; Baek, K.-H. Endophyte Bacillus velezensis isolated from Citrus spp. Controls streptomycin-resistant Xanthomonas citri subsp. citri that causes citrus bacterial canker. Agronomy 2019, 9, 470. [Google Scholar] [CrossRef] [Green Version]
- Qian, J.; Zhang, T.; Tang, S.; Zhou, L.; Li, K.; Fu, X.; Yu, S. Biocontrol of citrus canker with endophyte Bacillus amyloliquefaciens QC-Y. Plant Prot. Sci. 2020, 57, 1–13. [Google Scholar] [CrossRef]
- Lai, J.; Kuang, W.; Liu, B.; Song, S. Identification of endophytic bacterial strain GN223 and its effectiveness against citrus canker disease in navel orange under field conditions. Biocontrol Sci. Technol. 2022, 32, 14–29. [Google Scholar] [CrossRef]
- Daungfu, O.; Youpensuk, S.; Lumyong, S. Endophytic bacteria isolated from citrus plants for biological control of citrus canker in lime plants. Trop. Life Sci. Res. 2019, 30, 73. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.N.; Ali, M.S.; Choi, S.-J.; Hyun, J.-W.; Baek, K.-H. Biocontrol of citrus canker disease caused by Xanthomonas citri subsp. citri using an endophytic Bacillus thuringiensis. Plant Pathol. J. 2019, 35, 486. [Google Scholar] [CrossRef] [PubMed]
- Balogh, B.; Canteros, B.I.; Stall, R.E.; Jones, J.B. Control of citrus canker and citrus bacterial spot with bacteriophages. Plant Dis. 2008, 92, 1048–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WAKIMOTO, S. Some characteristics of citrus canker bacteria, Xanthomonas citri (Hasse) Dowson, and the related phages isolated from Japan. Jpn. J. Phytopathol. 1967, 33, 301–310. [Google Scholar] [CrossRef]
- Balogh, B. Characterization and Use of Bacteriophages Associated with Citrus Bacterial Pathogens for Disease Control; University of Florida: Gainesville, FL, USA, 2006. [Google Scholar]
- Ahmad, A.A.; Askora, A.; Kawasaki, T.; Fujie, M.; Yamada, T. The filamentous phage XacF1 causes loss of virulence in Xanthomonas axonopodis pv. citri, the causative agent of citrus canker disease. Front. Microbiol. 2014, 5, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obradovic, A.; Jones, J.B.; Momol, M.; Balogh, B.; Olson, S. Management of tomato bacterial spot in the field by foliar applications of bacteriophages and SAR inducers. Plant Dis. 2004, 88, 736–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, Y.E.; Saleh, A.A.; Al-Saleh, M.A. Management of asiatic citrus canker under field conditions in Saudi Arabia using bacteriophages and acibenzolar-S-methyl. Plant Dis. 2017, 101, 761–765. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.A.; Terol, J.; Ibanez, V.; López-García, A.; Pérez-Román, E.; Borredá, C.; Domingo, C.; Tadeo, F.R.; Carbonell-Caballero, J.; Alonso, R. Genomics of the origin and evolution of Citrus. Nature 2018, 554, 311–316. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Orbovic, V.; Jones, J.B.; Wang, N. Modification of the PthA4 effector binding elements in Type I Cs LOB 1 promoter using Cas9/sg RNA to produce transgenic Duncan grapefruit alleviating XccΔpthA4: dCs LOB 1.3 infection. Plant Biotechnol. J. 2016, 14, 1291–1301. [Google Scholar] [CrossRef]
- Dutt, M.; Barthe, G.; Irey, M.; Grosser, J. Transgenic citrus expressing an Arabidopsis NPR1 gene exhibit enhanced resistance against Huanglongbing (HLB.; Citrus Greening). PLoS ONE 2015, 10, e0137134. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Pathotype | References | |||||
|---|---|---|---|---|---|---|
| A | B | C | D | E | ||
| Common disease name | Asiatic canker | cancrosis B/false canker | Mexican lime cancrosis/cancrosis C | Citrus bacteriosis/Mexican bacteriosis(later named as citrus leaf spot) | Citrus Bacterial Spot (CBS)/Florida Nursery strain of CC | [34,35,36] |
| Pathovar | citri | Aurantifolii (Strain-B) | aurantifolii (Strain-C) | aurantifolii (Strain-D) (misunderstood) | citrumelo | |
| Origin | Asia | Argentina | Brazil | Mexico | Florida | |
| Known geographical distribution | Many citrus growing regions especially in Asia, USA, South America, Oceania | Argentina, Uruguay, Paraguay | State of Sao Paulo, Brazil | Mexico | Florida | |
| Discovery | 1830 (controversial) | 1923 on lemon | 1963 on Key/Mexican lime | 1981 on Key/Mexican lime | 1984 on Swingle citrumelo | [6,34,37] |
| Susceptible host | Sweet orange, grapefruit, lemon, pummelo, mandarin, sweet lime, also observed in some other rutaceous plants | Lemons and Mexican lime, also observed in sweet orange, grapefruit, cider, mandarin, Volkamer lemon, sweet lime | Mexican lime, also detected in sour orange and lemon | Mexican lime | Swingle citrumelo, Grapefruit, mandarin, sour orange, sweet orange, lemon, Key/Mexican lime | [30,34,35,36] |
| pthAor its functional homologs | Present | Present | Present | - | Absent | [30,31,38] |
| Pathogenicity | Highest | Low | High | Lower | lowest | |
| Symptoms | Distinctive corky necrotic lesions, sometimes possessing chlorotic or water-soaked haloes | Same as A, but symptoms take longer to appear, and lesions may vary in size from A | Similar to A | Similar to A | Flat water-soaked spots which may be surrounded by necrosis | [30,35] |
| Parts of plant that may be affected | Leaves, twigs, young stems, or fruits | Leaves, twigs, young stems, or fruits | Leaves, twigs, young stems, or fruits | Leaves, twigs, young stems, or fruits | Usually twigs and leaves only | |
| Defoliation and dieback | May occur | May occur | May occur | May occur | Does not occur | |
| No. of Bacterial strains | Many strains | Many strains | Many strains | Only one strain known (Xc 90) | Many strains | [39] |
| Xcc | XauB | XauC | References | ||
|---|---|---|---|---|---|
| Host range | Diverse | Less diverse | Restricted | [30] | |
| Xanthum gum production | Highest | Almost 3 times less than Xcc | Almost 2 times less than Xcc | ||
| Cellular growth | Non-fastidious, similar cellular mass values as XauC | Fastidious | Non-fastidious, similar cellular mass values as Xcc. | ||
| Comparative genetic analysis | |||||
| Genes related to flagellum synthesis | Major 3 clusters of genes (F1, F2, and F3) present. Another 4th cluster is also present, consisting of 2 genes. Unrelated genes are also present in the region between F1 and F2 (XACSR9). | F2 gene cluster is absent. No genes are present in the region between F1 and F2. | All four gene clusters are present. No genes are present in the region between F1 and F2. | ||
| Presence of XacPNP gene | Present | Absent | Absent | ||
| Type 1 Secretion System genes (T1SS) | hlyB and hlyD encoding genes and TolC present | hlyB and hlyD encoding genes absent, TolC present | hlyB and hlyD encoding genes and TolC present | [31] | |
| Type 4 Secretion System genes (T4SS) | Both in plasmid and chromosome | Only in plasmid, lack chromosomal copy | Only in plasmid, lack chromosomal copy | ||
| Genes involved in the regulation and synthesis of Type IV pilus (T4p) | Many genes are present including pilX, pilV, pilA, pilL, and fimT, forming atleast 4 clusters of genes. | Among different clusters of genes, pilX, pilA, pilV.pil anf fimT genes are absent | Among different clusters of genes, pilX, pilA, pilV, pil, and fimT genes are absent | ||
| Genes related to synthesis of Hemagglutinin and Hemolysin | Present in two regions of genome XAC4112-XAC4125 XAC1810-XAC1819 | The genes present in 2nd region (XAC1810-XAC 1819) including fhaB and fhaC are absent | The genes present in 2nd region (XAC1810-XAC 1819) including fhaB and fhaC are absent | ||
| vapBC and tspO gene | Present | Absent | Absent | ||
| Effector XopS | Present (in some cases as pseudogene) | Absent | Absent | ||
| Effector XopK | Present | Found as pseudogene | Found as pseudogene | [30,31,51] | |
| Effector XopF1, xopB, xopE4, xopJ, xopAF, xopAG | Absent | Present (in some cases as pseudogene) | Present (in some cases as pseudogene) | ||
| Effectors xopE2, xopN, xopP, xopAE | Present | Present | Absent | ||
| Basic physiological, biochemical, and serological tests | [39] | ||||
| Reaction with Mab A1 (ELISA test) | Reacts | Does not react | Does not react | ||
| Phage sensitivity | Bacteriophage Cp1 | Variable response | Insensitive | Insensitive | |
| Bacteriophage Cp2 | Variable response | Insensitive | Insensitive | ||
| Bacteriophage Cp3 | Insensitive | Sensitive | Insensitive | ||
| Casein hydrolysis test | Positive | Negative | Positive | ||
| Gelatin hydrolysis test | Positive | Negative | Negative | ||
| Growth in presence of | 3% NaCl | Grows | No growth observed | No growth observed | |
| Maltose | Grows | No growth observed | No growth observed | [54] | |
| Aspartic acid | Grows | No growth observed | No growth observed | ||
| General features of genome | [15,30] | ||||
| Genome | |||||
| Size (bp) | 5,274,174 | 4,877,808 | 5,012,633 | ||
| # of contigs | 3 | 239 | 351 | ||
| %GC | 64.7 | 64.9 | 64.8 | ||
| Protein coding genes | |||||
| Total | 4427 | 3804 | 3921 | ||
| With functional assignment | 2779 | 2694 | 2728 | ||
| Hypothetical | 262 | 117 | 184 | ||
| Conserved hypothetical | 1386 | 993 | 1009 | ||
| RNAs | |||||
| rRNA operons | 2 | 2 | 2 | ||
| tRNAs | 54 | 51 | 51 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahbaz, E.; Ali, M.; Shafiq, M.; Atiq, M.; Hussain, M.; Balal, R.M.; Sarkhosh, A.; Alferez, F.; Sadiq, S.; Shahid, M.A. Citrus Canker Pathogen, Its Mechanism of Infection, Eradication, and Impacts. Plants 2023, 12, 123. https://doi.org/10.3390/plants12010123
Shahbaz E, Ali M, Shafiq M, Atiq M, Hussain M, Balal RM, Sarkhosh A, Alferez F, Sadiq S, Shahid MA. Citrus Canker Pathogen, Its Mechanism of Infection, Eradication, and Impacts. Plants. 2023; 12(1):123. https://doi.org/10.3390/plants12010123
Chicago/Turabian StyleShahbaz, Esha, Mobeen Ali, Muhammad Shafiq, Muhammad Atiq, Mujahid Hussain, Rashad Mukhtar Balal, Ali Sarkhosh, Fernando Alferez, Saleha Sadiq, and Muhammad Adnan Shahid. 2023. "Citrus Canker Pathogen, Its Mechanism of Infection, Eradication, and Impacts" Plants 12, no. 1: 123. https://doi.org/10.3390/plants12010123