
Coalescent-Based Species Delimitation in Herbaceous Bamboos (Bambusoideae, Olyreae) from Eastern Brazil: Implications for Taxonomy and Conservation in a Group with Weak Morphological Divergence Coupled with Low Genetic Diversity
 , and
, and
Abstract
:1. Introduction
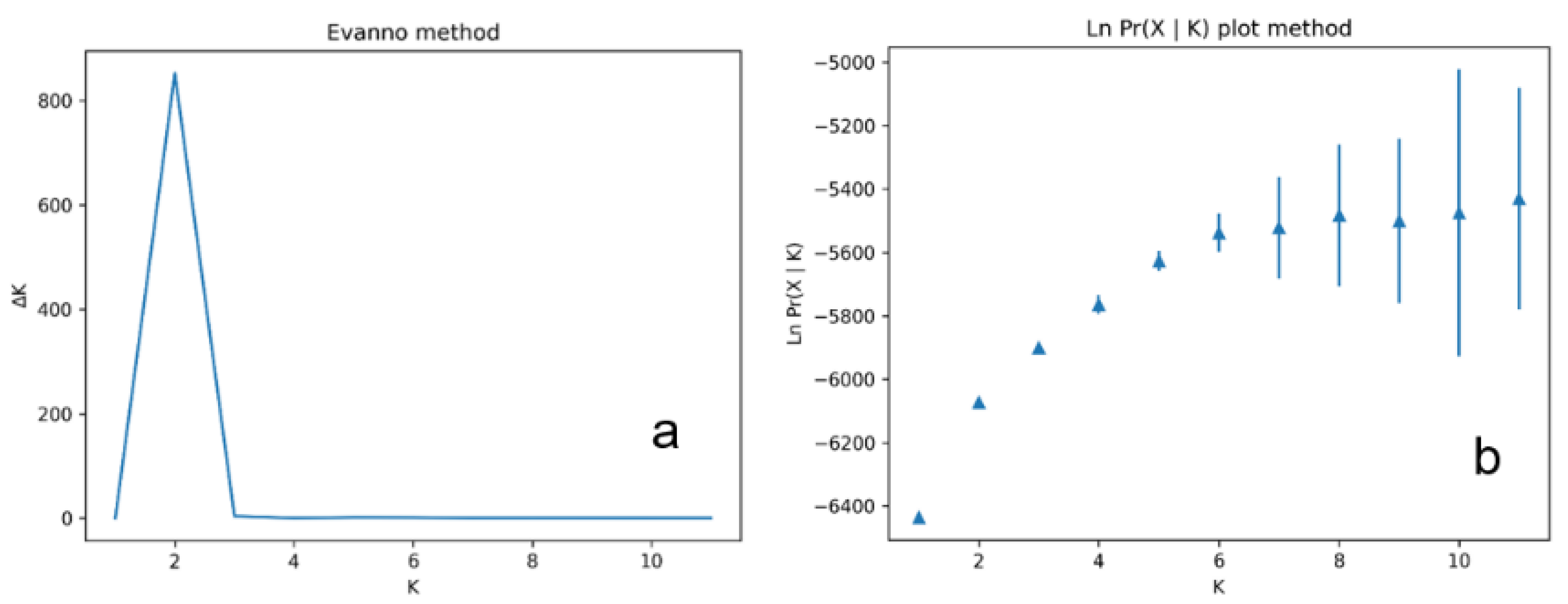
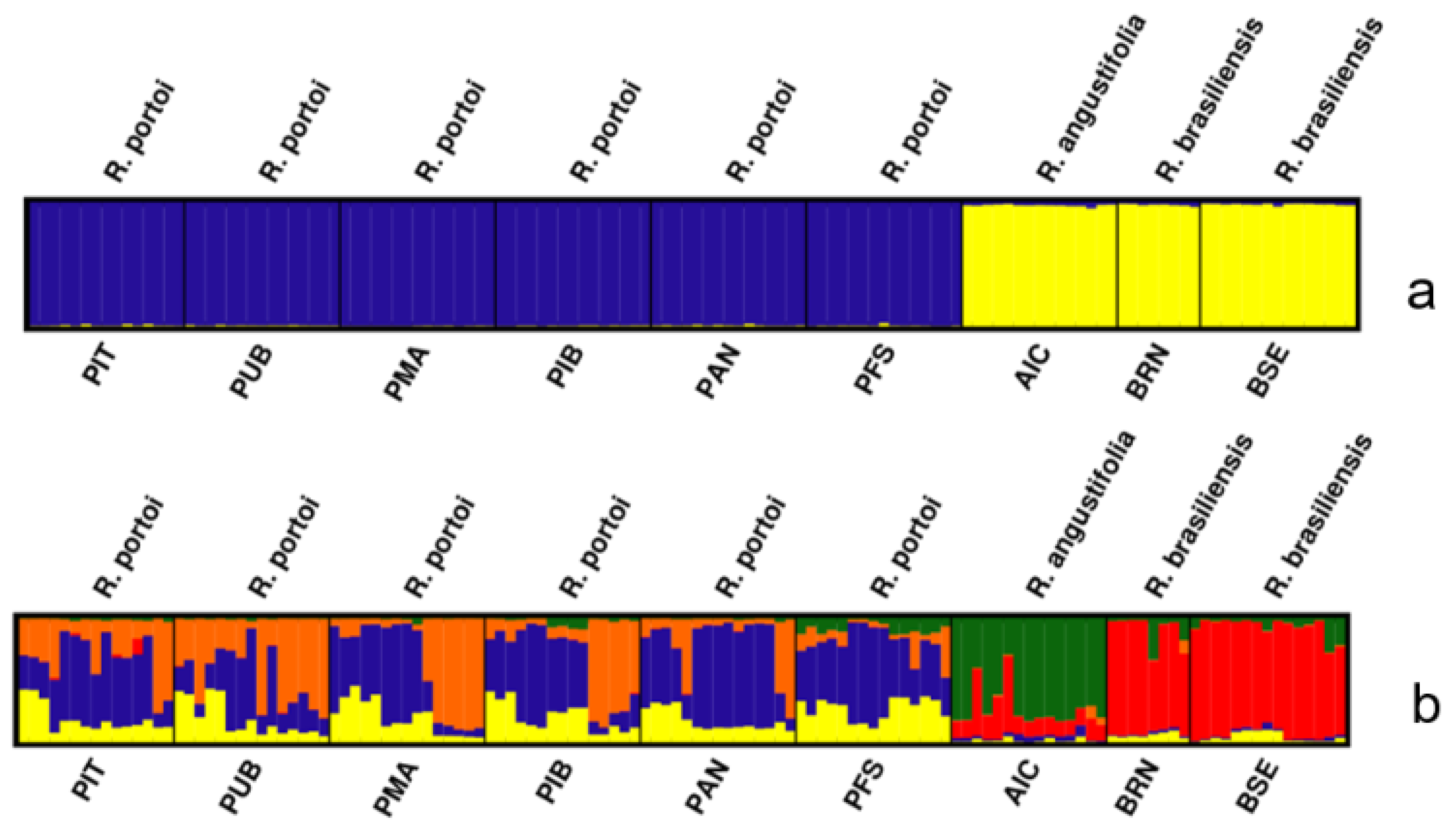
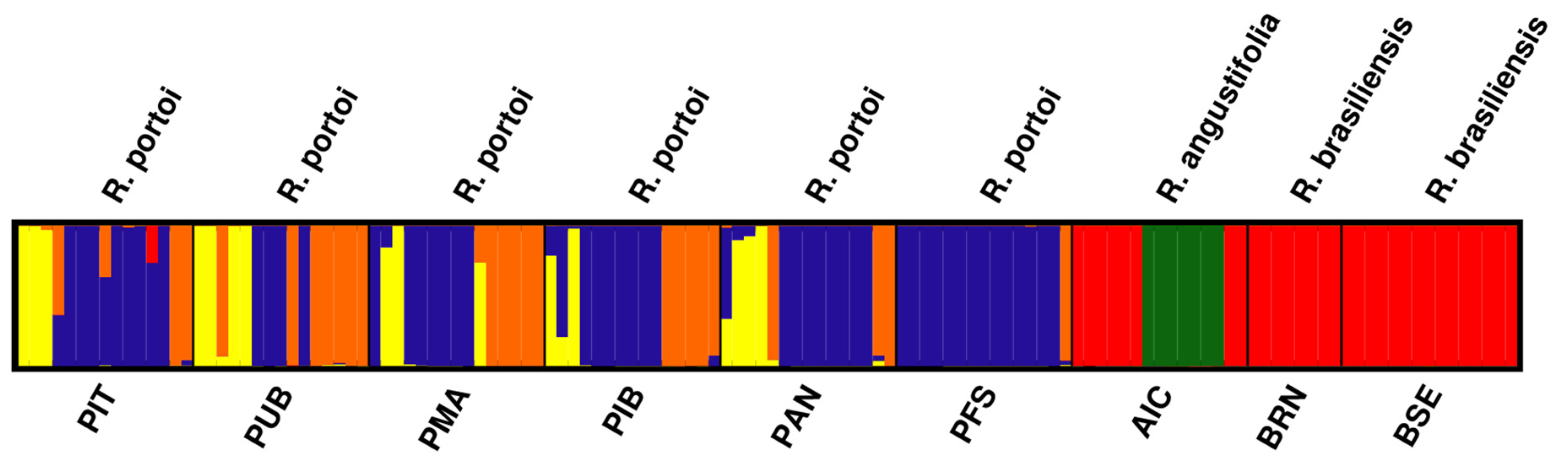
2. Results
3. Materials and Methods
3.1. Sampling
3.2. DNA Isolation, PCR Amplification, and Sequencing
3.3. Sequence Alignment and Bayesian Species Delimitation
3.4. ISSR-PCR Amplification and Polymorphism Test
3.5. Species Delimitation Using Cluster Algorithms
4. Discussion
Species Boundaries in Raddia portoi and R. angustifolia and Implications for Their Conservation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wiens, J.J. Species Delimitation: New Approaches for Discovering Diversity. Syst. Biol. 2007, 56, 875–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, R.R.; Coyne, J.A. Mathematical Consequences of the Genealogical Species Concept. Evolution 2002, 56, 1557–1565. [Google Scholar] [PubMed]
- Knowles, L.L.; Carstens, B.C. Delimiting Species without Monophyletic Gene Trees. Syst. Biol. 2007, 56, 887–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z. Molecular Evolution: A Statistical Approach, 1st ed.; Oxford University Press: Oxford, UK, 2014; ISBN 978-0-19-960260-5. [Google Scholar]
- Yang, Z.; Rannala, B. Bayesian Species Delimitation Using Multilocus Sequence Data. Proc. Natl. Acad. Sci. USA 2010, 107, 9264–9269. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Rannala, B. Unguided Species Delimitation Using DNA Sequence Data from Multiple Loci. Mol. Biol. Evol. 2014, 31, 3125–3135. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Hausdorf, B.; Hennig, C. Species Delimitation Using Dominant and Codominant Multilocus Markers. Syst. Biol. 2010, 59, 491–503. [Google Scholar] [CrossRef] [Green Version]
- Carstens, B.C.; Pelletier, T.A.; Reid, N.M.; Satler, J.D. How to Fail at Species Delimitation. Mol. Ecol. 2013, 22, 4369–4383. [Google Scholar] [CrossRef]
- Leaché, A.D.; Fujita, M.K. Bayesian Species Delimitation in West African Forest Geckos (Hemidactylus Fasciatus). Proc. R. Soc. B Biol. Sci. 2010, 277, 3071–3077. [Google Scholar] [CrossRef] [Green Version]
- Duminil, J.; Di Michele, M. Plant Species Delimitation: A Comparison of Morphological and Molecular Markers. Plant Biosyst. 2009, 143, 528–542. [Google Scholar] [CrossRef]
- Shaffer, H.B.; Thomson, R.C. Delimiting Species in Recent Radiations. Syst. Biol. 2007, 56, 896–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, R.P.; Borba, E.L.; Longhi-Wagner, H.M.; Pereira, A.C.S.; Lambert, S.M. Genetic and Morphological Variability in the Raddia brasiliensis Complex (Poaceae: Bambusoideae). Plant Syst. Evol. 2008, 274, 25–35. [Google Scholar] [CrossRef]
- Oliveira, R.P.; Borba, E.L.; Longhi-Wagner, H.M. Morphometrics of Herbaceous Bamboos of the Raddia brasiliensis Complex (Poaceae-Bambusoideae): Implications for the Taxonomy of the Genus and New Species from Brazil. Plant Syst. Evol. 2008, 270, 159–182. [Google Scholar] [CrossRef]
- de Carvalho, M.L.S.; de Jesus, I.S.D.; da Silva, R.M.; Leite, K.R.B.; Schnadelbach, A.S.; Clark, L.G.; de Oliveira, R.P. Cryptic Speciation in the Herbaceous Bamboo Genus Piresia (Poaceae, Olyreae). Bot. J. Linn. Soc. 2020, 192, 82–96. [Google Scholar] [CrossRef]
- Ferreira, F.M.; Oliveira, R.P.; Welker, C.A.D.; da Costa Dórea, M.; de Carvalho Lima, A.L.; Oliveira, I.L.C.; dos Santos, F.; van den Berg, C.; Clark, L.G. Phylogenetic Relationships within Parianinae (Poaceae: Bambusoideae: Olyreae) with Emphasis on Eremitis: Evidence from Nuclear and Plastid DNA Sequences, Macromorphology, and Pollen Ectexine Patterns. Mol. Phylogenet. Evol. 2019, 139, 106541. [Google Scholar] [CrossRef]
- Ferreira, F.M.; Silva, C.; Welker, C.A.D.; Dórea, M.C.; Leite, K.R.B.; Clark, L.G.; Oliveira, R.P. Eremitis berbertii and E. fluminensis (Poaceae, Bambusoideae): New Species from the Brazilian Atlantic Forest and Updates on Leaf Microcharacters in the Genus. Novon 2020, 28, 240–252. [Google Scholar] [CrossRef]
- Ferreira, F.M.; Welker, C.A.D.; Clark, L.G.; Oliveira, R.P. Eremitis jardimii (Poaceae, Bambusoideae), a New Species from Bahia, Brazil. Kew Bull. 2020, 75, 52. [Google Scholar] [CrossRef]
- Oliveira, R.P.; Clark, L.G.; Schnadelbach, A.S.; Monteiro, S.H.N.; Borba, E.L.; Longhi-Wagner, H.M.; van den Berg, C. A Molecular Phylogeny of Raddia and Its Allies within the Tribe Olyreae (Poaceae, Bambusoideae) Based on Noncoding Plastid and Nuclear Spacers. Mol. Phylogenet. Evol. 2014, 78, 105–117. [Google Scholar] [CrossRef]
- Soderstrom, T.R.; Zuloaga, F.O. New Species of Grasses in Arberella, Cryptochloa, and Raddia (Poaceae: Bambusoideae: Olyreae). Brittonia 1985, 37, 22–35. [Google Scholar] [CrossRef]
- Oliveira, R.P. Estudos Taxonômicos, Filogenéticos e Biossistemáticos Em Raddia (Poaceae: Bambusoideae). Ph.D. Dissertation, Universidade Estadual de Feira de Santana, Feira de Santana, Brazil, 2006. [Google Scholar]
- Oliveira, R.P.; Longhi-Wagner, H.M.; Leite, K.R.B. A Contribuição Da Anatomia Foliar Para a Taxonomia de Raddia Bertol. (Poaceae: Bambusoideae). Acta Bot. Bras. 2008, 22, 1–19. [Google Scholar] [CrossRef]
- de Oliveira, R.P.; Shirasuna, R.T. Raddia in Lista de Espécies Da Flora Do Brasil. Available online: http://floradobrasil2015.jbrj.gov.br/FB13550 (accessed on 4 November 2022).
- Filgueiras, T.; Oliveira, R.P.; Sfair, J.C.; Monteiro, N.P.; Borges, R.A.X. Poaceae. In Livro Vermelho da Flora do Brasil; Martinelli, G., Moraes, M.A., Eds.; Instituto de Pesquisas Jardim Botânico do Rio de Janeiro: Rio de Janeiro, Brasil, 2013; pp. 858–881. [Google Scholar]
- Ministério do Meio Ambiente Portaria n. 148, de 7 de Junho de 2022. In Diário Of. Da União; Imprensa Nacional: Brasília, Brazil, 2022; Seção 1; pp. 74–103.
- Secretaria Estadual do Meio Ambiente do Estado da Bahia Portaria n. 40, de 21 de Agosto de 2017. In Diário Of. do Estado da Bahia; Empresa Gráfica da Bahia: Salvador, Brazil, 2017; pp. 22–27.
- Burke, S.V.; Lin, C.-S.; Wysocki, W.P.; Clark, L.G.; Duvall, M.R. Phylogenomics and Plastome Evolution of Tropical Forest Grasses (Leptaspis, Streptochaeta: Poaceae). Front. Plant Sci. 2016, 7, 1993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Sanchez, E.; Sosa, V.; Ortiz-Rodriguez, A.E.; Davidse, G. Historical Biogeography of the Herbaceous Bamboo Tribe Olyreae (Bambusoideae: Poaceae). Folia Geobot. 2019, 54, 177–189. [Google Scholar] [CrossRef]
- Wysocki, W.P.; Clark, L.G.; Attigala, L.; Ruiz-Sanchez, E.; Duvall, M.R. Evolution of the Bamboos (Bambusoideae; Poaceae): A Full Plastome Phylogenomic Analysis. BMC Evol. Biol. 2015, 15, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, R.P.; Silva, C.; Welker, C.A.D.; Da Dórea, M.C.; Oliveira, I.L.C.; Vieira, J.P.S.; Leite, K.R.B.; Clark, L.G. Reinterpreting the Phylogenetic Position, Systematics and Distribution of the Raddia-Sucrea Lineage (Poaceae, Olyrinae), with a New Monotypic and Endangered Herbaceous Bamboo Genus from Brazil. Bot. J. Linn. Soc. 2020, 192, 34–60. [Google Scholar] [CrossRef]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 17 November 2022).
- Doyle, J.J.; Doyle, J.L. A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Sun, Y.; Skinner, D.Z.; Liang, G.H.; Hulbert, S.H. Phylogenetic Analysis of Sorghum and Related Taxa Using ITS. Theor. Appl. Genet. 1994, 89, 26–32. [Google Scholar] [CrossRef]
- Demesure, B.; Sodzi, N.; Petit, R.J. A Set of Universal Primers for Amplification of Polymorphic Non-Coding Regions of Mitochondrial and Chloroplast DNA in Plants. Mol. Ecol. 1995, 4, 129–131. [Google Scholar] [CrossRef]
- Doyle, J.J.; Davis, J.I.; Soreng, R.J.; Garvin, D.; Anderson, M.J. Chloroplast DNA Inversions and the Origin of the Grass Family (Poaceae). Proc. Natl. Acad. Sci. USA 1992, 89, 7722–7726. [Google Scholar] [CrossRef] [Green Version]
- Dun, I.S.; Blattner, F.R. Charons 36 to 40: Multi Enzyone, High Capacity, Recombination Deficient Replacement Vectors with Polylinkers and Ploystuffers. Nucleic Acids Res. 1987, 15, 2677–2698. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols, a Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Desfeux, C.; Lejeune, B. Systematics of Euromediterranean Silene (Caryophyllaceae): Evidence from a Phylogenetic Analysis Using ITS Sequences. C. R. Acad. Sci. III 1996, 319, 351–358. [Google Scholar]
- Staden, R.; Beal, K.F.; Bonfield, J.K. The Staden Package, 1998. In Bioinformatics Methods and Protocols; Humana Press: Totowa, NJ, USA, 2000; pp. 115–130. [Google Scholar]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A Novel Method for Rapid Multiple Sequence Alignment Based on Fast Fourier Transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sela, I.; Ashkenazy, H.; Katoh, K.; Pupko, T. GUIDANCE2: Accurate Detection of Unreliable Alignment Regions Accounting for the Uncertainty of Multiple Parameters. Nucleic Acids Res. 2015, 43, W7–W14. [Google Scholar] [CrossRef] [PubMed]
- Flouri, T.; Jiao, X.; Rannala, B.; Yang, Z. Species Tree Inference with BPP Using Genomic Sequences and the Multispecies Coalescent. Mol. Biol. Evol. 2018, 35, 2585–2593. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z. The BPP Program for Species Tree Estimation and Species Delimitation. Curr. Zool. 2015, 61, 854–865. [Google Scholar] [CrossRef]
- Wolfe, A.D.; Xiang, Q.-Y.; Kephart, S.R. Assessing Hybridization in Natural Populations of Penstemon (Scrophulariaceae) Using Hypervariable Intersimple Sequence Repeat (ISSR) Bands. Mol. Ecol. 1998, 7, 1107–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falush, D.; Stephens, M.; Pritchard, J.K.; Smith, J.W. Inference of Population Structure Using Multilocus Genotype Data: Linked Loci and Correlated Allele Frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef] [PubMed]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of Population Structure Using Multilocus Genotype Data: Dominant Markers and Null Alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the Number of Clusters of Individuals Using the Software STRUCTURE: A Simulation Study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Wen, W.; Falush, D. Documentation for STRUCTURE Software: Version 2; University of Chicago: Chicago, IL, USA, 2010. [Google Scholar]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A Website and Program for Visualizing STRUCTURE Output and Implementing the Evanno Method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Hunter, J.D. Matplotlib: A 2D Graphics Environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A Cluster Matching and Permutation Program for Dealing with Label Switching and Multimodality in Analysis of Population Structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, N.A. DISTRUCT: A Program for the Graphical Display of Population Structure. Mol. Ecol. Resour. 2004, 4, 137–138. [Google Scholar] [CrossRef]
- Reeves, P.A.; Richards, C.M.; Paterson, A. Distinguishing Terminal Monophyletic Groups from Reticulate Taxa: Performance of Phenetic, Tree-Based, and Network Procedures. Syst. Biol. 2007, 56, 302–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeves, P.A.; Richards, C.M. Accurate Inference of Subtle Population Structure (and Other Genetic Discontinuities) Using Principal Coordinates. PLoS One 2009, 4, e4269. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing 2021; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Gower, J.C. Some Distance Properties of Latent Root and Vector Methods Used in Multivariate Analysis. Biometrika 1966, 53, 325–338. [Google Scholar] [CrossRef]
- Paradis, E.; Schliep, K. Ape 5.0: An Environment for Modern Phylogenetics and Evolutionary Analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Jaccard, P. Étude Comparative de La Distribution Florale Dans Une Portion Des Alpes et Des Jura. Bull. Soc. Vaud. Sci. Nat. 1901, 37, 547–579. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package 2019. Available online: vegandevs.github.io/vegan (accessed on 15 August 2022).
- Scrucca, L.; Fop, M.; Murphy, T.B.; Raftery, A.E. Mclust 5: Clustering, Classification and Density Estimation Using Gaussian Finite Mixture Models. R J. 2016, 8, 289–317. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of Molecular Variance Inferred from Metric Distances among DNA Haplotypes: Application to Human Mitochondrial DNA Restriction Data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research—An Update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Manni, F.; Guerard, E.; Heyer, E. Geographic Patterns of (Genetic, Morphologic, Linguistic) Variation: How Barriers Can Be Detected by Using Monmonier’s Algorithm. Hum. Biol. 2004, 76, 173–190. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Genetic Distance between Populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Vekemans, X.; Beauwens, T.; Lemaire, M.; Roldán-Ruiz, I. Data from Amplified Fragment Length Polymorphism (AFLP) Markers Show Indication of Size Homoplasy and of a Relationship between Degree of Homoplasy and Fragment Size. Mol. Ecol. 2002, 11, 139–151. [Google Scholar] [CrossRef]
- Zhivotovsky, L.A. Estimating Population Structure in Diploids with Multilocus Dominant DNA Markers. Mol. Ecol. 1999, 8, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Edwards, D.L.; Knowles, L.L. Species Detection and Individual Assignment in Species Delimitation: Can Integrative Data Increase Efficacy? Proc. R. Soc. B Biol. Sci. 2014, 281, 20132765. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, T.A.; Crisafulli, C.; Wagner, S.; Zellmer, A.J.; Carstens, B.C. Historical Species Distribution Models Predict Species Limits in Western Plethodon Salamanders. Syst. Biol. 2015, 64, 909–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Sanchez, E. Parametric and Non-Parametric Species Delimitation Methods Result in the Recognition of Two New Neotropical Woody Bamboo Species. Mol. Phylogenet. Evol. 2015, 93, 261–273. [Google Scholar] [CrossRef]
- Ruiz-Sanchez, E.; Castro-Castro, A.; Zamora-Tavares, M.D.P. Molecular and Morphological Data Support the Recognition of a New Species of Otatea (Poaceae: Bambusoideae: Guaduinae) from Durango, Mexico. Taxon 2021, 70, 747–760. [Google Scholar] [CrossRef]
- Giarla, T.C.; Voss, R.S.; Jansa, S.A. Hidden Diversity in the Andes: Comparison of Species Delimitation Methods in Montane Marsupials. Mol. Phylogenet. Evol. 2014, 70, 137–151. [Google Scholar] [CrossRef]
- Jacobs, S.J.; Kristofferson, C.; Uribe-Convers, S.; Latvis, M.; Tank, D.C. Incongruence in Molecular Species Delimitation Schemes: What to Do When Adding More Data Is Difficult. Mol. Ecol. 2018, 27, 2397–2413. [Google Scholar] [CrossRef]
- Luo, A.; Ling, C.; Ho, S.Y.W.; Zhu, C.D. Comparison of Methods for Molecular Species Delimitation across a Range of Speciation Scenarios. Syst. Biol. 2018, 67, 830–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janes, J.K.; Miller, J.M.; Dupuis, J.R.; Malenfant, R.M.; Gorrell, J.C.; Cullingham, C.I.; Andrew, R.L. The K = 2 Conundrum. Mol. Ecol. 2017, 26, 3594–3602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, L.S.; Allendorf, F.W. The One-Migrant-per-Generation Rule in Conservation and Management. Conserv. Biol. 1996, 10, 1509–1518. [Google Scholar] [CrossRef]
- Waples, R.S.; Gaggiotti, O. What Is a Population? An Empirical Evaluation of Some Genetic Methods for Identifying the Number of Gene Pools and Their Degree of Connectivity. Mol. Ecol. 2006, 15, 1419–1439. [Google Scholar] [CrossRef]
- Moritz, C. Defining ‘Evolutionarily Significant Units’ for Conservation. Trends Ecol. Evol. 1994, 9, 373–375. [Google Scholar] [CrossRef]
- Bruschi, D.P.; Peres, E.A.; Lourenço, L.B.; de Macedo Bartoleti, L.F.; Sobral-Souza, T.; Recco-Pimentel, S.M. Signature of the Paleo-Course Changes in the São Francisco River as Source of Genetic Structure in Neotropical Pithecopus nordestinus (Phyllomedusinae, Anura) Treefrog. Front. Genet. 2019, 10, 728. [Google Scholar] [CrossRef] [Green Version]
- Coutinho-Abreu, I.V.; Sonoda, I.V.; Fonseca, J.A.; Melo, M.A.; Balbino, V.Q.; Ramalho-Ortigão, M. Lutzomyia longipalpis Sl. in Brazil and the Impact of the Sao Francisco River in the Speciation of This Sand Fly Vector. Parasit. Vectors 2008, 1, 16. [Google Scholar] [CrossRef] [Green Version]
- Malukiewicz, J.; Boere, V.; Fuzessy, L.F.; Grativol, A.D.; de Oliveira e Silva, I.; Pereira, L.C.M.; Ruiz-Miranda, C.R.; Valença, Y.M.; Stone, A.C. Natural and Anthropogenic Hybridization in Two Species of Eastern Brazilian Marmosets (Callithrix jacchus and C. penicillata). PLoS One 2015, 10, e0127268. [Google Scholar] [CrossRef] [Green Version]
- de Abreu Moreira, P.; Brandão, M.M.; de Araujo, N.H.; de Oliveira, D.A.; Fernandes, G.W. Genetic Diversity and Structure of the Tree Enterolobium contortisiliquum (Fabaceae) Associated with Remnants of a Seasonally Dry Tropical Forest. Flora—Morphol. Distrib. Funct. Ecol. Plants 2015, 210, 40–46. [Google Scholar] [CrossRef] [Green Version]
- do Nascimento, F.F.; Pereira, L.G.; Geise, L.; Bezerra, A.M.R.; D’Andrea, P.S.; Bonvicino, C.R. Colonization Process of the Brazilian Common Vesper Mouse, Calomys expulsus (Cricetidae, Sigmodontinae): A Biogeographic Hypothesis. J. Hered. 2011, 102, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Passoni, J.C.; Benozzati, M.L.; Rodrigues, M.T. Phylogeny, Species Limits, and Biogeography of the Brazilian Lizards of the Genus Eurolophosaurus (Squamata: Tropiduridae) as Inferred from Mitochondrial DNA Sequences. Mol. Phylogenet. Evol. 2008, 46, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.T. Herpetofauna da Caatinga. In Ecologia e Conservação da Caatinga; Leal, I.R., Tabarelli, M., Silva, J.M.C., Eds.; Universidade Federal de Pernambuco: Recife, Brazil, 2003; Volume 1, pp. 181–236. [Google Scholar]
- Flora e Funga do Brasil Poaceae in Flora e Funga Do Brasil. Available online: https://floradobrasil.jbrj.gov.br/FB102232 (accessed on 22 October 2022).
- Oliveira, I.L.C.; Matos, A.O.; Silva, C.; Carvalho, M.L.S.; Tyrrell, C.D.; Clark, L.G.; Oliveira, R.P. Delving Deeper into the Phylogenetics of the Herbaceous Bamboos (Poaceae, Bambusoideae, Olyreae): Evaluation of Generic Boundaries within the Parodiolyra/Raddiella Clade Uncovers a New Genus. Bot. J. Linn. Soc. 2020, 192, 61–81. [Google Scholar] [CrossRef]
- Oliveira, R.P. A Tribo Olyreae (Poaceae—Bambusoideae) No Estado Da Bahia, Brasil. Master’s Thesis, Universidade Estadual de Feira de Santana, Feira de Santatana, Brazil, 2001. [Google Scholar]
- Soderstrom, T.R.; Judziewicz, E.J.; Clark, L.G. Distribution Patterns of Neotropical Bamboos. In Proceedings of the a Workshop on Neotropical Distribution Patterns; Heyer, W.R., Vanzolini, P.E., Eds.; Academia Brasileira de Ciências: Rio de Janeiro, Brazil, 1988; pp. 121–157. [Google Scholar]
- Oliveira, R.P.; Longhi-Wagner, H.M.; Filgueiras, T.; Mota, A.C.; Viana, P.L. Poaceae. In Plantas Raras do Brasil; Conservação Internacional and Universidade Estadual de Feira de Santana: Belo Horizonte, Brazil, 2009; pp. 326–340. [Google Scholar]
- Galindo-Leal, C.; Câmara, I.G. Atlantic Forest hotspot status: An overview. In The Atlantic Forest of South America: Biodiversity Status, Threats, and Outlook; Galindo-Leal, C., Câmara, I.G., Eds.; Island Press: Washington, DC, USA, 2003; pp. 3–11. [Google Scholar]
- Leal, I.R.; Tabarelli, M.; Silva, J.M.C. Ecologia e Conservação Da Caatinga; Editora Universitária UFPE: Recife, Brazil, 2003. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scheme | Gamma Distribution for Prior | Best Species Tree and PP | Best Model for Species Number |
|---|---|---|---|
| 1 | θ ~ IG(3, 0.0002) and τ ~ IG(3, 0.0003) | ((A, B), P) 0.61 | P[3] = 0.99 |
| 2 | θ ~ IG(3, 0.0002) and τ ~ IG(3, 0.003) | ((A, B), P) 0.61 | P[3] = 1 |
| 3 | θ ~ IG(3, 0.0002) and τ ~ IG(3, 0.03) | ((A, B), P) 0.66 | P[3] = 1 |
| 4 | θ ~ IG(3, 0.002) and τ ~ IG(3, 0.003) | ((A, B), P) 0.54 | P[3] = 0.99 |
| 5 | θ ~ IG(3, 0.002) and τ ~ IG(3, 0.0003) | ((A, B), P) 0.48 | P[3] = 0.99 |
| 6 | θ ~ IG(3, 0.002) and τ ~ IG(3, 0.03) | ((A, B), P) 0.65 | P[3] = 0.99 |
| 7 | θ ~ IG(3, 0.00002) and τ ~ IG(3, 0.003) | ((A, B), P) 0.59 | P[3] = 1 |
| 8 | θ ~ IG(3, 0.00002) and τ ~ IG(3, 0.0003) | ((A, B), P) 0.6 | P[3] = 1 |
| 9 | θ ~ IG(3, 0.00002) and τ ~ IG(3, 0.003) | ((A, B), P) 0.64 | P[3] = 1 |
| Source of Variation | Degrees of Freedom | Sum of Squares | Variance Component | Variance (%) |
|---|---|---|---|---|
| Among Regions | 1 | 160.902 | 2.136 | 10% |
| Among Pops | 7 | 331.091 | 2.133 | 10% |
| Within Pops | 119 | 2020.992 | 16.983 | 80% |
| Total | 127 | 2512.984 | 21.252 | 100% |
| PIT | PUB | PMA | PIB | PAN | PFS | AIC | BRN | BSE | |
|---|---|---|---|---|---|---|---|---|---|
| 0.000 | 0.001 | 0.020 | 0.095 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | PIT |
| 0.082 | 0.000 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | PUB |
| 0.043 | 0.079 | 0.000 | 0.013 | 0.002 | 0.001 | 0.001 | 0.001 | 0.001 | PMA |
| 0.018 | 0.070 | 0.049 | 0.000 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | PIB |
| 0.069 | 0.069 | 0.074 | 0.073 | 0.000 | 0.001 | 0.001 | 0.001 | 0.001 | PAN |
| 0.121 | 0.179 | 0.167 | 0.095 | 0.149 | 0.000 | 0.001 | 0.001 | 0.001 | PFS |
| 0.183 | 0.153 | 0.201 | 0.173 | 0.151 | 0.224 | 0.000 | 0.001 | 0.001 | AIC |
| 0.191 | 0.205 | 0.263 | 0.236 | 0.217 | 0.278 | 0.204 | 0.000 | 0.002 | BRN |
| 0.197 | 0.212 | 0.266 | 0.209 | 0.190 | 0.286 | 0.180 | 0.104 | 0.000 | BSE |
| Taxon | Voucher | Location | Code | Coordinates | Sample Size | GenBank Accession Numbers |
|---|---|---|---|---|---|---|
| Raddia angustifolia Soderstr. & Zuloaga | RPO 1077 | Itaju do Colônia—BA | AIC | 15°08′33″ S 39°43′28″ W | ISSR: 15; ITS: 8; trnD-trnT: 6 | ITS: OP919392-OP919400; trnD-trnT: OP946258-OP946263 |
| R. portoi Kuhlm | RPO 1450 | Itaeté—BA | PIT | 12°56′40″ S 41°6′20″ W | ISSR: 15; ITS: 6; trnD-trnT: 6 | ITS: OP919419-OP919424; trnD-trnT: OP946281-OP946286 |
| RPO 1501 | Ubaíra—BA | PUB | 13°15′34″ S 39°50′9″ W | ISSR: 15; ITS: 10; trnD-trnT: 8 | ITS: OP919452-OP919461; trnD-trnT: OP946306-OP946313 | |
| MCD 72 | Maracás—BA | PMA | 13°24′24″ S 40°23′52″ W | ISSR: 15; ITS: 11; trnD-trnT: 7 | ITS: OP919425-OP919436 trnD-trnT: OP946287-OP946293 | |
| RPO 1439 | Itaberaba—BA | PIB | 12°18′11.0″ S 40°31′13.0″ W | ISSR: 15; ITS: 4; trnD-trnT: 5 | ITS: OP919414-OP919418; trnD-trnT: OP946276-OP946280 | |
| RPO 1548 | Anguera—BA | PAN | 12°9′49″ S 39°11′12″ W | ISSR: 15; ITS: 7; trnD-trnT: 6 | ITS: OP919407-OP919413; trnD-trnT: OP946270-OP946275 | |
| CS 314 | Feira de Santana—BA | PFS | 12° 6″ S 39° 2″ W | ISSR: 15; ITS: 6; trnD-trnT: 6 | ITS: OP919401-OP919406 trnD-trnT: OP946264-OP946269 | |
| RPO 2143 | Vitória da Conquista | - | 14°52′46.0″ S 40°47′28.0″ W | ITS: 3; trnD-trnT: 4 | ITS: OP919462-OP919464 trnD-trnT: OP946314- OP946317 | |
| MCD 126 | Sanharó—PE | - | 8°17′43.0″ S 36°30′46.0″ W | ITS: 12; trnD-trnT: 50 | ITS: OP919440-OP919451 trnD-trnT: OP946296-946305 | |
| R. brasiliensis Bertol | DC 2674 | Natal—RN | BRN | 5° 47′40″ S 35° 12′39″ W | ISSR: 8; ITS: 3; trnD-trnT: 2 | ITS: OP919437-OP919439 trnD-trnT: OP946294-OP946295 |
| A.P. Prata 2633 | Tobias Barreto—SE | BSE | 10°52′52″ S 39°59′11″ W | ISSR: 15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vieira, J.P.S.; Selbach-Schnadelbach, A.; Braz, M.; Ribeiro, P.L.; van den Berg, C.; Oliveira, R.P. Coalescent-Based Species Delimitation in Herbaceous Bamboos (Bambusoideae, Olyreae) from Eastern Brazil: Implications for Taxonomy and Conservation in a Group with Weak Morphological Divergence Coupled with Low Genetic Diversity. Plants 2023, 12, 107. https://doi.org/10.3390/plants12010107
Vieira JPS, Selbach-Schnadelbach A, Braz M, Ribeiro PL, van den Berg C, Oliveira RP. Coalescent-Based Species Delimitation in Herbaceous Bamboos (Bambusoideae, Olyreae) from Eastern Brazil: Implications for Taxonomy and Conservation in a Group with Weak Morphological Divergence Coupled with Low Genetic Diversity. Plants. 2023; 12(1):107. https://doi.org/10.3390/plants12010107
Chicago/Turabian StyleVieira, João Paulo S., Alessandra Selbach-Schnadelbach, Marcos Braz, Patrícia L. Ribeiro, Cássio van den Berg, and Reyjane P. Oliveira. 2023. "Coalescent-Based Species Delimitation in Herbaceous Bamboos (Bambusoideae, Olyreae) from Eastern Brazil: Implications for Taxonomy and Conservation in a Group with Weak Morphological Divergence Coupled with Low Genetic Diversity" Plants 12, no. 1: 107. https://doi.org/10.3390/plants12010107