
Distribution Patterns of Essential Oil Terpenes in Native and Invasive Solidago Species and Their Comparative Assessment

 , , , and
, , , and 
Abstract
:1. Introduction
2. Results
2.1. Essential Oil Content of Solidago Species
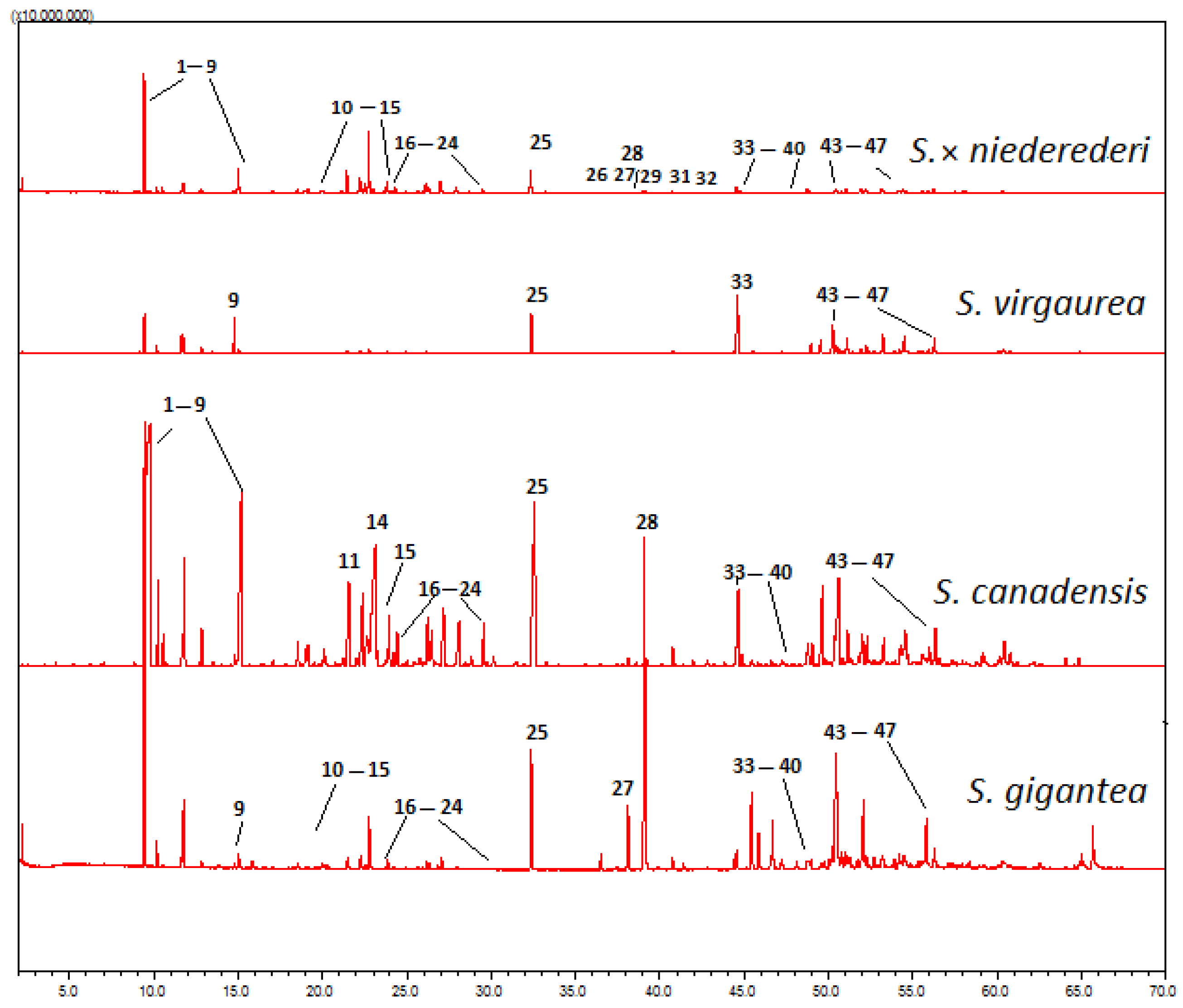
2.2. Chemical Profiles of Essential Oils
2.2.1. Solidago gigantea
2.2.2. Solidago canadensis
2.2.3. Solidago × niederederi
2.2.4. Solidago virgaurea
2.3. Interspecific Differences
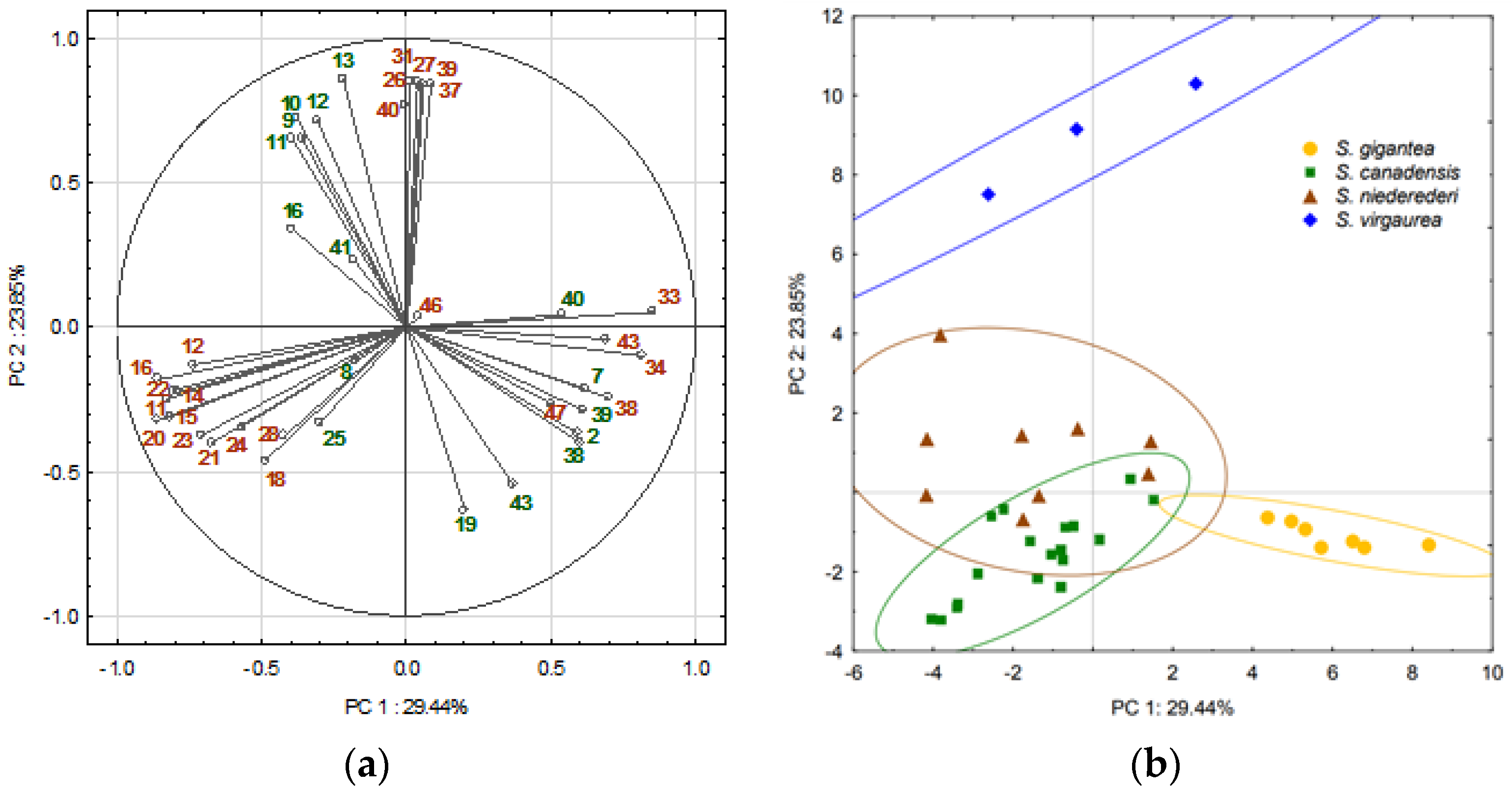
2.4. Principal Component Analysis (PCA)
3. Discussion
4. Material and Methods
4.1. Plant Material
4.2. Isolation of Essential Oils
4.3. Analysis of Essential Oils
4.4. Identification and Quantification of Components
4.5. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Franz, C.M. Essential oil research: Past, present and future. Flavour. Fragr. J. 2010, 25, 112–113. [Google Scholar] [CrossRef]
- Weber, E. Current and potential ranges of three exotic goldenrods (Solidago) in Europe. Conserv. Biol. 2001, 15, 122–128. [Google Scholar] [CrossRef]
- Karpavičienė, B.; Radušienė, J. Morphological and anatomical characterization of Solidago × niederederi and other sympatric Solidago species. Weed Sci. 2016, 64, 61–70. [Google Scholar] [CrossRef]
- Chen, G.Q.; Zhang, C.-B.; Ma, L.; Qiang, S.; Silander, J.A.; Li, L.Q. Biotic homogenization caused by the invasion of Solidago canadensis in China. J. Integr. Agric. 2013, 12, 835–845. [Google Scholar] [CrossRef]
- Khek, E. Floristisches aus Ober-Oesterreich. Allg. Bot. Z Syst. 1905, 11, 21–23. [Google Scholar]
- Gudžinskas, Z.; Petrulaitis, L. New alien plant species recorded in the southern regions of Latvia. Bot. Lith. 2016, 22, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Skokanová, K.; Šingliarová, B.; Španiel, S.; Hodálová, I.; Mereďa, P., Jr. Tracking the expanding distribution of Solidago × niederederi (Asteraceae) in Europe and first records from three countries within the Carpathian region. BioInvasions Rec. 2020, 9, 670–684. [Google Scholar] [CrossRef]
- Pal, R.W.; Chen, S.; Nagy, D.U.; Callaway, R.M. Impacts of Solidago gigantea on other species at home and away. Biol. Invasions 2015, 17, 3317–3325. [Google Scholar] [CrossRef]
- Yuan, Y.; Wang, B.; Zhang, S.; Tang, J.; Tu, C.; Hu, S.; Yong, J.W.H.; Chen, X. Enhanced allelopathy and competitive ability of invasive plant Solidago canadensis in its introduced range. J. Plant Ecol. 2013, 6, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Świerszcz, S.; Szymura, M.; Wolski, K.; Szymuret, T.H. Comparison of methods for restoring meadows invaded by Solidago species. Pol. J. Environ. Stud. 2017, 26, 1251–1258. [Google Scholar] [CrossRef]
- Zihare, L.; Blumberga, D. Insight into bioeconomy. Solidago canadensis as a valid resource. Brief review. Energy Procedia 2017, 128, 275–280. [Google Scholar] [CrossRef]
- Ciesielczuk, T.; Poluszyńska, J.; Rosik-Dulewska, C.; Sporek, M.; Lenkiewicz, M. Uses of weeds as an economical alternative to processed wood biomass and fossil fuels. Ecol. Eng. 2016, 95, 485–491. [Google Scholar] [CrossRef]
- Staab, M.; Pereira-Peixoto, M.H.; Klein, A.-M. Exotic garden plants partly substitute for native plants as resources for pollinators when native plants become seasonally scarce. Oecologia 2020, 194, 465–480. [Google Scholar] [CrossRef] [PubMed]
- ESCOP Monographs. Solidaginis Virgaureae Herba (European Goldenrod). 2018. Available online: https://escop.com/downloads/solidaginis-virgaureae-herba-european-goldenrod (accessed on 10 February 2022).
- Final Community Herbal Monograph on Solidago virgaurea L., herba. (Doc. Ref. EMEA/HMPC/285758/2007) 2008. Available online: https://www.ema.europa.eu/en/documents/herbal-monograph/final-community-herbal-monograph-solidago-virgaurea-l-herba_en.pdf (accessed on 15 April 2022).
- Council of Europe. European Pharmacopoeia. Strasbourg: Council of Europe. 2001. Available online: https://pheur.edqm.eu/app/10-8/search/ (accessed on 15 April 2022).
- Demir, H.; Acik, L.; Bali, E.B.; Koc, Y.; Kaynak, G. Antioxidant and antimicrobial activities of Solidago virgaurea extracts. Afr. J. Biotechnol. 2009, 8, 274–279. [Google Scholar]
- Marksa, M.; Zymone, K.; Ivanauskas, L.; Radušienė, J.; Pukalskas, A.; Raudone, L. Antioxidant profiles of leaves and inflorescences of native, invasive and hybrid Solidago species. Ind. Crops Prod. 2020, 145, 112123. [Google Scholar] [CrossRef]
- Shelepova, O.; Vinogradova, Y.; Zaitchik, B.; Ruzhitsky, A.; Grygorieva, O.; Brindza, J. Constituents of the essential oil in Solidago canadensis L. from Eurasia. Potravin. Slovak J. Food Sci. 2018, 12, 20–25. [Google Scholar] [CrossRef] [Green Version]
- Radušiene, J.; Marksa, M.; Karpavičiene, B. Assessment of Solidago × niederederi origin based on the accumulation of phenolic compounds in plant raw materials. Weed Sci. 2018, 66, 324–330. [Google Scholar] [CrossRef]
- Liu, S.; Shao, X.; Wei, Y.; Li, Y.; Xu, F.; Wang, H. Solidago canadensis L. essential oil vapor effectively inhibits Botrytis cinerea growth and preserves postharvest quality of strawberry as a food model system. Front Microbiol. 2016, 7, 1179. [Google Scholar] [CrossRef]
- Werrie, P.-Y.; Durenne, B.; Delaplace, P.; Fauconnier, M.-L. Phytotoxicity of EOs: Opportunities and constraints for the development of biopesticides. A Review. Foods 2020, 9, 1291. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Grul’ová, D.; Baranová, B.; Caputo, L.; De Martino, L.; Sedlák, V.; Camele, I.; De Feo, V. Antimicrobial activity and chemical composition of essential oil extracted from Solidago canadensis L. growing wild in Slovakia. Molecules 2019, 24, 1206. [Google Scholar] [CrossRef] [Green Version]
- Sarić-Krsmanović, M.; Gajić Umiljendić, J.; Radivojević, L.; Šantrić, L.; Potočnik, I.; Đurović-Pejčev, R. Bio-herbicidal effects of five EOs on germination and early seedling growth of velvetleaf (Abutilon theophrasti Medik.). J. Environ. Sci. Heal. Part B 2019, 54, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Li, D.Q.; Pan, S.H.; Zhu, X.W.; Tan, L.; Cao, Y.F. Anticancer activity and chemical composition of leaf essential oil from Solidago canadensis L. in China. Adv. Mat. Res. 2012, 347, 1584–1589. [Google Scholar] [CrossRef]
- Wandjou, J.G.N.; Quassinti, L.; Gudžinskas, Z.; Nagy, D.U.; Cianfaglione, K.; Bramucci, M.; Maggi, F. Chemical composition and antiproliferative effect of EOs of four Solidago species (S. canadensis, S. gigantea, S. virgaurea and S. × niederederi). Chem. Biodivers. 2020, 17, e2000685. [Google Scholar] [CrossRef]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. EOs’ chemical characterization and investigation of some biological activities: A critical review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perricone, M.; Arace, E.; Corbo, M.R.; Sinigaglia, M.; Bevilacqua, A. Bioactivity of EOs: A review on their interaction with food components. Front. Microbiol. 2015, 6, 76. [Google Scholar] [CrossRef] [Green Version]
- Benelli, G.; Pavela, R.; Cianfaglione, K.; Nagy, D.U.; Canale, A.; Maggi, F. Evaluation of two invasive plant invaders in Europe (Solidago canadensis and Solidago gigantea) as possible sources of botanical insecticides. J. Pest Sci. 2019, 92, 805–821. [Google Scholar] [CrossRef]
- Kalemba, D. Constituents of the essential oil of Solidago virgaurea L. Flavour Fragr. J. 1998, 13, 373–376. [Google Scholar] [CrossRef]
- Kalemba, D.; Marschall, H.; Bradesi, P. Constituents of the essential oil of Solidago gigantea Ait. (giant goldenrod). Flavour Fragr. J. 2001, 16, 19–26. [Google Scholar] [CrossRef]
- Kalemba, D.; Thiem, B. Constituents of the EOs of four micropropagated Solidago species. Flavour Fragr. J. 2004, 19, 40–43. [Google Scholar] [CrossRef]
- Kołodziej, B.; Kowalski, R.; Kędzia, B. Antibacterial and antimutagenic activity of extracts above ground parts of three Solidago species: Solidago virgaurea L. Solidago canadensis L. and Solidago gigantea Ait. J. Med. Plant Res. 2011, 5, 6770–6779. [Google Scholar] [CrossRef]
- Gruľová, D.; Baranová, B.; Ivanova, V.; De Martino, L.; Mancini, E.; De Feo, V. Composition and bio activity of EOs of Solidago spp. and their impact on radish and garden cress. Allelopath. J. 2016, 39, 129–142. [Google Scholar]
- Huang, B.K.; Lei, Y.L.; Qin, L.P.; Liu, J. Chemical composition and cytotoxic activities of the essential oil from inflorescences of Solidago canadensis L., an invasive weed in Southeastern China. J. Essent. Oil-Bear. Plants 2012, 15, 667–671. [Google Scholar] [CrossRef]
- Lawson, S.K.; Sharp, L.G.; Powers, C.N.; McFeeters, R.L.; Satyal, P.; Setzer, W.N. Volatile compositions and antifungal activities of native American medicinal plants: Focus on the Asteraceae. Plants 2020, 9, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Sherei, M.; Khaleel, A.; Motaal, A.A.; Abd-Elbaki, P. Effect of seasonal variation on the composition of the essential oil of Solidago canadensis cultivated in Egypt. J. Essent. Oil-BearPlants 2014, 17, 891–898. [Google Scholar] [CrossRef]
- Semple, J.C. An intuitive phylogeny and summary of chromosome number variation in the goldenrod genus Solidago (Asteraceae: Astereae). Phytoneuron 2016, 32, 1–9. [Google Scholar]
- Macel, M.; de Vos, R.C.H.; Jeroen, J.J.; van der Putten, W.H.; van Dam, N.M. Novel chemistry of invasive plants: Exotic species have more unique metabolomic profiles than native congeners. Ecol. Evol. 2014, 4, 2777–2786. [Google Scholar] [CrossRef]
- Synowiec, A.; Kalemba, D.; Drozdek, E.; Bocianowski, J. Phytotoxic potential of EOs from temperate climate plants against the germination of selected weeds and crops. J. Pest Sci. 2017, 90, 407–419. [Google Scholar] [CrossRef]
- Baser, K.H.C. Biological and pharmacological activities of carvacrol and carvacrol bearing essential oils. Curr. Pharm. Des. 2008, 14, 3106–3119. [Google Scholar] [CrossRef]
- Mastelić, J.; Jerković, I.; Blažević, I.; Poljak-Blaži, M.; Borović, S.; Ivanćić-Baće, I.; Smrećki, V.; Žarković, N.; Brćić-Kostic, K.; Vikić-Topić, D.; et al. Comparative study on the antioxidant and biological activities of carvacrol, thymol, and eugenol derivatives. J. Agric. Food Chem. 2008, 56, 3989–3996. [Google Scholar] [CrossRef]
- Orians, C.M. The effects of hybridization in plants on secondary chemistry: Implications for the ecology and evolution of plant—Herbivore interactions. Am. J. Bot. 2001, 87, 1749–1756. [Google Scholar] [CrossRef] [Green Version]
- Schierenbeck, K.A.; Ellstrand, N.C. Hybridization and the evolution of invasiveness in plants and other organisms. Biol. Invasions 2009, 11, 1093–1105. [Google Scholar] [CrossRef]
- López-Caamal, A.; Tovar-Sánchez, E. Genetic, morphological, and chemical patterns of plant hybridization. Rev. Chil. Hist. Nat. 2014, 87, 16. [Google Scholar] [CrossRef] [Green Version]
- Chizzola, R. Regular monoterpenes and sesquiterpenes (EOs). In Natural Products; Ramawat, K., Mérillon, J.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 2973–3008. [Google Scholar] [CrossRef]
- Verdeguer, M.; Sánchez-Moreiras, A.M.; Araniti, F. Phytotoxic effects and mechanism of action of EOs and terpenoids. Plants 2020, 9, 1571. [Google Scholar] [CrossRef] [PubMed]
- Rolli, E.; Marieschi, M.; Maietti, S.; Sacchetti, G.; Bruni, R. Comparative phytotoxicity of 25 EOs on pre-and postemergence development of Solanum lycopersicum L.: A multivariate approach. Ind. Crops Prod. 2014, 60, 280–290. [Google Scholar] [CrossRef]
- Azizan, K.A.; Ghani, N.H.A.; Nawawi, M.F. Discrimination and prediction of the chemical composition and the phytotoxic activity of Wedelia trilobata essential oil (EO) using metabolomics and chemometrics. Plant Biosyst. 2020, 1–16. [Google Scholar] [CrossRef]
- Mutlu, S.; Atici, Ö.; Esim, N. Bioherbicidal effects of essential oils of Nepeta meyeri Benth. on weed spp. Allelopath. J. 2010, 26, 291–300. [Google Scholar]
- Verdeguer, M.; Blázquez, M.A.; Boira, H. Phytotoxic effects of Lantana camara, Eucalyptus camaldulensis and Eriocephalus africanus essential oils in weeds of Mediterranean summer crops. Biochem. Syst. Ecol. 2009, 37, 362–369. [Google Scholar] [CrossRef]
- Vasilakoglou, I.; Dhima, K.; Paschalidis, K.; Ritzoulis, C. Herbicidal potential on Lolium rigidum of nineteen major essential oil components and their synergy. J. Essent. Oil Res. 2013, 25, 1–10. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Drenaggi, E.; Desneux, N.; Maggi, F. Phytol, (E)-nerolidol and spathulenol from Stevia rebaudiana leaf essential oil as effective and eco-friendly botanical insecticides against Metopolophium dirhodum. Ind. Crops Prod. 2020, 155, 12844. [Google Scholar] [CrossRef]
- Landi, M.; Misra, B.B.; Muto, A.; Bruno, L.; Araniti, F. Phytotoxicity, morphological, and metabolic effects of the sesquiterpenoid nerolidol on Arabidopsis thaliana seedling roots. Plants 2020, 9, 1347. [Google Scholar] [CrossRef]
- Sen, A.; Kurkcuoglu, M.; Yildirim, A.; Senkardes, I.; Bitis, L.; Baser, K.H.C. Chemical composition, antiradical, and enzyme inhibitory potential of essential oil obtained from aerial part of Centaurea pterocaula Trautv. J. Ess. Oil. Res. 2021, 33, 44–52. [Google Scholar] [CrossRef]
- Albouchi, F.; Ghazouani, N.; Souissi, R.; Abderrabba, M.; Boukhris-Bouhachem, S. Aphidicidal activities of Melaleuca styphelioides Sm. essential oils on three citrus aphids: Aphis gossypii Glover; Aphis spiraecola Patch and Myzus persicae (Sulzer). S. Afr. J. Bot. 2018, 117, 149–154. [Google Scholar] [CrossRef]
- Boulogne, I.; Petit, P.; Ozier-Lafontaine, H.; Desfontaines, L.; Loranger-Merciris, G. Insecticidal and antifungal chemicals produced by plants: A review. Environ. Chem. Lett. 2012, 10, 325–347. [Google Scholar] [CrossRef] [Green Version]
- Nararak, J.; Sathantriphop, S.; Kongmee, M.; Mahiou-Leddet, V.; Ollivier, E.; Manguin, S.; Chareonviriyaphap, T. Excito-repellent activity of β-caryophyllene oxide against Aedes aegypti and Anopheles minimus. Acta Trop. 2019, 197, 105030. [Google Scholar] [CrossRef] [PubMed]
- Abd-ElGawad, A.M.; El Gendy, A.E.-N.G.; Assaeed, A.M.; Al-Rowaily, S.L.; Alharthi, A.S.; Mohamed, T.A.; Nassar, M.I.; Dewir, Y.H.; Elshamy, A.I. Phytotoxic effects of plant essential oils: A systematic review and structure-activity relationship based on chemometric analyses. Plants 2021, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Zidorn, C. Guidelines for consistent characterisation and documentation of plant source materials for studies in phytochemistry and phytopharmacology. Phytochemistry 2017, 139, 56–59. [Google Scholar] [CrossRef]
- Barra, A. Factors affecting chemical variability of essential oils: A review of recent developments. Nat. Prod. Commun. 2009, 4, 1147–1154. [Google Scholar] [CrossRef] [Green Version]
- Nowak, M.; Kleinwächter, M.; Manderscheid, R.; Weige, H.J.; Selmar, D. Drought stress increases the accumulation of monoterpenes in sage (Salvia officinalis), an effect that is compensated by elevated carbon dioxide concentration. J. Appl. Bot. Food Qual. 2010, 83, 133–136. Available online: https://ojs.openagrar.de/index.php/JABFQ/article/view/2138 (accessed on 15 April 2022).
- Caser, M.; Chitarra, W.; D’Angiolillo, F.; Perrone, I.; Demasi, S.; Lovisolo, C.; Pistelli, L.; Pistelli, L.; Scariot, V. Drought stress adaptation modulates plant secondary metabolite production in Salvia dolomitica Codd. Ind. Crops Prod. 2019, 129, 85–96. [Google Scholar] [CrossRef]
- Paulsen, J.; Selmar, D. Case study: The difficulty of correct reference values when evaluating the effects of drought stress: A case study with Thymus vulgaris. J. Appl. Bot. Food Qual. 2016, 89, 287–289. [Google Scholar]
- De Almeida, L.F.R.; Portella, R.O.; Facanali, R.R.; Marques, M.O.M.; Frei, F. Dry and wet seasons set the phytochemical profile of the Copaifera langsdorffii Desf. EOs. J. Essent. Oil. Res. 2016, 26, 292–300. [Google Scholar] [CrossRef]
- Ribeiro, A.F.; Andrade, E.H.A.; Salimena, F.R.G.; Maia, J.G.S. Circadian and seasonal study of the cinnamate chemotype from Lippia origanoides Kunth. Biochem. Syst. Ecol. 2014, 55, 249–259. [Google Scholar] [CrossRef]
- Tsusaka, T.; Makino, B.; Ohsawa, R.; Ezura, H. Genetic and environmental factors influencing the contents of essential oil compounds in Atractylodes lancea. PLoS ONE 2019, 14, e0217522. [Google Scholar] [CrossRef] [PubMed]
- Tardugno, R.; Serio, A.; Purgatorio, C.; Savini, V.; Paparella, A.; Benvenuti, S. Thymus vulgaris L. essential oils from Emilia Romagna Apennines (Italy): Phytochemical composition and antimicrobial activity on food-borne pathogens. Nat. Prod. Res. 2022, 36, 837–842. [Google Scholar] [CrossRef] [PubMed]
- Padilla-González, G.F.; Frey, M.; Gómez-Zeledón, J.; Da Costa, F.B.; Spring, O. Metabolomic and gene expression approaches reveal the developmental and environmental regulation of the secondary metabolism of yacón (Smallanthus sonchifolius, Asteraceae). Sci. Rep. 2019, 9, 13178. [Google Scholar] [CrossRef] [PubMed]
- Moshrefi Araghi, A.; Nemati, H.; Azizi, M.; Moshtaghi, N.; Shoor, M.; Hadian, J. Assessment of phytochemical and agro-morphological variability among different wild accessions of Mentha longifolia L. cultivated in field condition. Ind. Crop. Prod. 2019, 140, 111698. [Google Scholar] [CrossRef]
- Zhao, S.Y.; Sun, S.G.; Dai, C.; Gituru, R.W.; Chen, J.M.; Wang, Q.F. Genetic variation and structure in native and invasive Solidago canadensis populations. Weed Res. 2014, 55, 163–172. [Google Scholar] [CrossRef]
- Wei, X.; Koo, I.; Kim, S.; Zhang, X. Compound identification in GC–MS by simultaneously evaluating the mass spectrum and retention index. Analyst 2014, 139, 2507–2514. [Google Scholar] [CrossRef] [Green Version]
- Babushok, V.I.; Linstrom, P.J.; Zenkevich, I.G. Retention indices for frequently reported compounds of plant essential oils. J. Phys. Chem. Ref. 2011, 40, 043101. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| No | Compounds | LRI Exp. | LRI Ref. | S. gigantea (n = 7) | S. canadensis (n = 18) | S. × niederederi (n = 9) | S. virgaurea (n = 3) | p | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | M | SD | F | M | SD | F | M | SD | F | M | SD | |||||
| 1 | α-pinene | 930 | 930 | 100 | 10.7 | 8.31 | 100 | 18.1 | 13.84 | 100 | 17.4 | 12.84 | 100 | 28.6 | 8.96 | 0.196 |
| 2 | camphene | 945 | 948 | 71.4 | 1.8 | 0.77 | 72.2 | 1.7 | 0.69 | 77.8 | 1.2 | 0.34 | 100 | 1.0 | 0.35 | 0.713 |
| 3 | thuja-2,4(10)-diene | 950 | 957 | 28.6 | 0.6 | 0.35 | 66.7 | 0.5 | 0.16 | 44.4 | 0.7 | 0.32 | 66.7 | 0.6 | 0.13 | 0.504 |
| 4 | sabinene | 971 | 976 | 71.4 | 2.5 | 2.07 | 66.7 | 1.1 | 0.79 | 66.7 | 1.3 | 1.79 | 33.3 | 0.6 | – | 0.258 |
| 5 | β-pinene | 974 | 980 | 71.4 | 3.0 | 0.90 | 83.3 | 2.3 | 1.40 | 88.9 | 2.2 | 1.71 | 100 | 3.6 | 1.51 | 0.536 |
| 6 | β-myrcene | 992 | 991 | 42.9 | 1.6 | 0.34 | 55.6 | 0.8 | 0.60 | 33.3 | 0.6 | 0.15 | 66.7 | 1.5 | 0.23 | 0.564 |
| 7 | p-cymene | 1018 | 1014 | 42.9 | 2.5 | 1.42 | 5.6 | 0.5 | – | 0 | 0 | – | 0 | 0 | – | 0.021 |
| 8 | o-cymene | 1022 | 1009–1076 | 42.9 | 10.9 | 6.57 | 55.6 | 0.5 | 0.42 | 33.3 | 0.5 | 0.30 | 0 | 0 | – | 0.338 |
| 9 | limonene | 1023 | 10270 | 71.4 | 1.1 | 0.19 | 100 | 5.2 | 4.48 | 88.9 | 5.8 | 5.05 | 100 | 1.1 | 0.57 | 0.032 |
| 10 | linalool | 1090 | 1098 | 0 | 0 | – | 16.7 | 0.1 | 0.07 | 44.4 | 0.5 | 0.10 | 33.3 | 0.8 | – | 0.105 |
| 11 | 1α-campholenal | 1109 | 1105 | 85.7 | 1.3 a | 0.58 | 100 | 4.1 b | 1.45 | 100 | 3.6 b | 1.61 | 66.7 | 3.1 | 1.35 | 0.004 |
| 12 | trans-pinocarveol | 1135 | 1139 | 100 | 0.8 a | 0.20 | 88.9 | 2.9 b | 1.18 | 100 | 3.0 b | 1.28 | 100 | 1.6 | 0.93 | 0.005 |
| 13 | trans-sabinol | 1139 | 1140 | 42.9 | 0.3 | 0.16 | 0 | 0 | – | 0 | 0 | – | 0 | 0 | – | 0.004 |
| 14 | trans-verbenol | 1143 | 1144 | 100 | 2.9 a | 1.12 | 100 | 13.1 b | 5.39 | 100 | 13.6 b | 5.79 | 100 | 6.6 | 3.43 | 0.001 |
| 15 | cis-verbenol | 1145 | 1142 | 100 | 0.5 a | 0.27 | 100 | 1.9 b | 0.88 | 100 | 1.8 b | 0.88 | 66.7 | 1.2 | 0.23 | 0.001 |
| 16 | pinocarvone | 1158 | 1160 | 85.7 | 0.4 a | 0.04 | 100 | 1.6 b | 0.50 | 88.9 | 1.8 b | 0.62 | 66.7 | 1.4 | 0.34 | 0.003 |
| 17 | p-mentha-1,5-dien-8-ol | 1164 | 1166 | 0 | 0 | – | 5.6 | 0.4 | 0.31 | 44.4 | 0.7 | 0.39 | 0 | 0 | – | 0.016 |
| 18 | α-phellandrene-8-ol | 1165 | 1166 | 0 | 0 a | – | 77.8 | 0.9 b | 0.32 | 0 | 0 a | – | 0 | 0 | – | <0.001 |
| 19 | terpinen-4-ol | 1174 | 1175 | 42.9 | 1.0 | 0.51 | 16.7 | 0.3 | 0.03 | 22.2 | 0.4 | 0.06 | 0 | 0 | – | 0.241 |
| 20 | myrtenal | 1192 | 1193 | 85.7 | 0.5 a | 0.13 | 100 | 1.9 b | 0.63 | 88.9 | 1.7 b | 0.57 | 66.7 | 1.2 | 0.24 | 0.001 |
| 21 | myrtenol | 1193 | 1194 | 85.7 | 0.7 a | 0.35 | 100 | 1.5 b | 0.63 | 88.9 | 1.3 | 0.54 | 33.3 | 1.1 | – | 0.014 |
| 22 | verbenone | 1205 | 1205 | 42.9 | 0.4 a | 0.29 | 100 | 2.7 b | 1.19 | 100 | 2.7 b | 1.38 | 100 | 1.2 | 0.67 | <0.001 |
| 23 | trans-carveol | 1219 | 1215 | 14.3 | 0.6 a | – | 94.4 | 1.9 b | 0.69 | 88.9 | 1.6 | 0.60 | 33.3 | 0.8 | – | <0.003 |
| 24 | carvone | 1241 | 1242 | 0 | 0 a | – | 94.4 | 1.3 b | 0.18 | 77.8 | 1.1 | 0.52 | 0 | 0 | – | <0.001 |
| 25 | bornyl acetate | 1287 | 1285 | 100 | 10.4 | 4.89 | 100 | 11.9 | 4.74 | 100 | 9.3 | 5.73 | 100 | 4.1 | 2.81 | 0.098 |
| 26 | α-cubebene | 1347 | 1345 | 0 | 0 | – | 0 | 0 a | – | 0 | 0 | – | 100 | 1.0 b | 0.40 | <0.001 |
| 27 | α-copaene | 1372 | 1376 | 0 | 0 | – | 11.1 | 0.2 a | 0.01 | 0 | 0 | – | 100 | 2.6 b | 1.34 | <0.001 |
| 28 | β-cubebene | 1380 | 1389 | 0 | 0 a | – | 77.8 | 4.9 b | 3.21 | 11.1 | 6.0 a | 0.04 | 0 | 0 | – | <0.001 |
| 29 | β-elemene | 1389 | 1391 | 42.9 | 0.3 | 0.08 | 55.6 | 0.8 | 0.46 | 55.6 | 0.7 | 0.28 | 0 | 0 | – | 0.243 |
| 30 | (E)-β-caryophyllene | 1414 | 1419 | 71.4 | 0.9 | 0.31 | 88.9 | 0.6 | 0.16 | 44.4 | 0.8 | 0.44 | 66.7 | 0.7 | 0.04 | 0.425 |
| 31 | β-copaene | 1424 | 1432 | 14.3 | 0.4 | – | 22.2 | 0.2 a | 0.13 | 33.3 | 3.6 | 3.80 | 100 | 12.2 | 8.15 | 0.006 |
| 32 | α-humulene | 1448 | 1452 | 42.9 | 0.4 | 0.04 | 38.9 | 0.2 | 0.21 | 22.2 | 1.3 | 1.34 | 0 | 0 | – | 0.588 |
| 33 | γ-muurolene | 1475 | 1477 | 100 | 0.8 a | 0.23 | 0 | 0 b | – | 0 | 0 b | – | 33.3 | 0.7 | – | <0.001 |
| 34 | germacrene D | 1480 | 1480 | 100 | 11.2 a | 4.80 | 22.2 | 8.1 b | 4.21 | 88.9 | 2.5 | 1.65 | 100 | 1.5 | 0.20 | <0.001 |
| 35 | β-selinene | 1481 | 1486 | 57.1 | 0.4 | 0.01 | 16.7 | 0.3 | 0.08 | 22.2 | 0.7 | 0.01 | 0 | 0 | – | 0.155 |
| 36 | epi-cubebol | 1489 | 1493 | 14.3 | 0.7 | – | 0 | 0 | – | 11.1 | 1.0 | – | 66.7 | 1.6 | 1.24 | 0.007 |
| 37 | α-muurolene | 1499 | 1499 | 28.6 | 0.4 | 0.03 | 0 | 0 a | – | 22.2 | 1.1 | 0.16 | 100 | 1.4 b | 0.78 | <0.001 |
| 38 | γ-cadinene | 1507 | 1513 | 85.7 | 0.7 a | 0.26 | 16.7 | 0.4 b | 0.09 | 0 | 0 b | – | 0 | 0 | – | <0.001 |
| 39 | cubebol | 1514 | 1515 | 0 | 0 | – | 0 | 0 a | – | 22.2 | 1.5 | 0.04 | 100 | 2.8 b | 2.08 | <0.001 |
| 40 | δ-cadinene | 1520 | 1524 | 14.3 | 0.5 | – | 0 | 0 a | – | 22.2 | 1.1 | 0.16 | 100 | 1.3 b | 0.90 | <0.001 |
| 41 | epoxyazulene | 1561 | 1554 | 71.4 | 6.4 | 2.98 | 55.6 | 1.9 | 1.22 | 88.9 | 1.6 | 1.35 | 0 | 0 | – | 0.054 |
| 42 | (E)-nerolidol | 1565 | 1564 | 71.4 | 0.7 | 0.28 | 11.1 | 0.4 | 0.13 | 11.1 | 0.4 | – | 0 | 0 | – | 0.002 |
| 43 | spathulenol | 1575 | 1576 | 100 | 5.0 a | 2.97 | 38.9 | 0.9 b | 0.74 | 66.7 | 2.8 | 3.96 | 100 | 1.3 | 0.20 | <0.001 |
| 44 | caryophyllene oxide | 1578 | 1581 | 100 | 3.6 | 2.44 | 100 | 3.6 | 2.17 | 100 | 8.4 | 8.26 | 100 | 8.5 | 2.48 | 0.083 |
| 45 | viridiflorol | 1586 | 1590 | 0 | 0 | – | 0 | 0 | – | 0 | 0 | – | 33.3 | 1.2 | – | 0.010 |
| 46 | humulene epoxide II | 1607 | 1606 | 0 | 0 | – | 0 | 0 a | – | 77.8 | 4.5 b | 8.60 | 0 | 0 | – | <0.001 |
| 47 | isospathulenol | 1627 | 1630 | 100 | 6.1 a | 6.47 | 83.3 | 2.3 | 1.52 | 66.7 | 2.1 | 0.73 | 33.3 | 1.3 b | – | 0.018 |
| Monoterpene hydrocarbons | 100 | 25.4 | 16.52 | 100 | 28.0 | 20.88 | 100 | 26.4 | 18.09 | 100 | 35.9 | 11.86 | 0.608 | |||
| Oxygenated monoterpenes | 100 | 18.5 a | 6.04 | 100 | 46.6 b | 15.42 | 100 | 41.5 b | 16.35 | 100 | 19.2 | 7.60 | <0.001 | |||
| Sesquiterpene hydrocarbons | 100 | 15.3 b | 6.63 | 100 | 7.9 | 6.00 | 100 | 6.0 | 4.85 | 100 | 20.7 a | 10.64 | 0.007 | |||
| Oxygenated sesquiterpenes | 100 | 19.7 a | 8.85 | 100 | 7.1 b | 3.48 | 100 | 17.0 | 15.54 | 100 | 14.7 | 5.25 | <0.001 | |||
| Oxygenated diterpenes | 14.3 | 0.1 | – | 11.1 | 0.9 | 0.19 | 22.2 | 1.0 | 0.51 | 0 | 0 | – | 0.479 | |||
| No | Compounds | a LRI Exp. | b LRI Ref. | S. gigantea (n = 7) | S. canadensis (n = 18) | S. × niederederi (n = 9) | S. virgaurea (n = 3) | p | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | M | SD | F | M | SD | F | M | SD | F | M | SD | |||||
| 1 | α-pinene | 930 | 930 | 100 | 7.8 | 2.18 | 100 | 8.6 | 5.69 | 100 | 11.9 | 10.37 | 100 | 16.0 | 11.77 | 0.856 |
| 2 | 1 camphene | 945 | 948 | 100 | 2.8 a | 0.76 | 100 | 1.2 b | 0.40 | 77.8 | 1.1 b | 0.45 | 66.7 | 0.5 b | 0.07 | <0.001 |
| 3 | thuja-2,4(10)-diene | 950 | 957 | 0 | 0 | – | 11.1 | 0.2 | 0.13 | 33.3 | 0.6 | 0.13 | 33.3 | 0.3 | – | 0.187 |
| 4 | sabinene | 971 | 976 | 42.9 | 1.1 | 0.61 | 44.4 | 0.6 | 0.42 | 33.3 | 1.1 | 1.21 | 66.7 | 0.3 | 0.01 | 0.926 |
| 5 | β-pinene | 974 | 980 | 100 | 2.2 | 0.88 | 94.4 | 1.4 | 0.61 | 88.9 | 1.9 | 1.16 | 66.7 | 3.5 | 0.64 | 0.193 |
| 6 | β-myrcene | 992 | 988 | 28.6 | 1.2 | 0.12 | 5.6 | 1.0 | – | 22.2 | 0.8 | 0.49 | 33.3 | 0.4 | – | 0.372 |
| 7 | o-cymene | 1022 | 1009–1076 | 100 | 2.1 a | 1.40 | 50.0 | 0.5 b | 0.46 | 22.2 | 0.4 b | 0.06 | 0 | 0 b | – | <0.001 |
| 8 | limonene | 1023 | 1027 | 100 | 1.1 a | 0.16 | 100 | 2.5 b | 2.17 | 100 | 2.2 | 1.56 | 66.7 | 1.1 | 0.35 | 0.006 |
| 9 | α-campholenal | 1109 | 1105 | 14.3 | 0.6 a | – | 77.8 | 0.8 | 0.72 | 77.8 | 2.6 | 1.68 | 100 | 2.8 b | 0.52 | 0.007 |
| 10 | trans-pinocarveol | 1135 | 1139 | 42.9 | 0.3 a | 0.16 | 66.7 | 0.9 a | 0.50 | 77.8 | 1.6 | 0.75 | 100 | 3.6 b | 1.83 | 0.003 |
| 11 | trans-verbenol | 1143 | 1144 | 42.9 | 0.5 a | 0.23 | 100 | 2.7 b | 1.81 | 100 | 8.9 b | 6.85 | 100 | 24.6 b | 21.21 | <0.001 |
| 12 | cis-verbenol | 1145 | 1142 | 0 | 0 a | – | 27.8 | 0.4 | 0.27 | 44.4 | 1.6 | 0.39 | 100 | 1.5 b | 0.82 | 0.004 |
| 13 | pinocarvone | 1158 | 1160 | 0 | 0 a | – | 22.2 | 0.6 a | 0.31 | 66.7 | 1.1 | 0.51 | 100 | 1.8 b | 0.04 | <0.001 |
| 14 | myrtenal | 1192 | 1193 | 0 | 0 | – | 72.2 | 0.6 | 0.29 | 55.6 | 1.6 | 0.48 | 66.7 | 1.6 | 0.02 | 0.024 |
| 15 | myrtenol | 1193 | 1194 | 0 | 0 | – | 55.6 | 0.3 | 0.25 | 55.6 | 1.0 | 0.33 | 66.7 | 1.2 | 0.03 | 0.038 |
| 16 | verbenone | 1205 | 1206 | 42.9 | 0.3 a | 0.16 | 88.9 | 1.5 | 1.35 | 77.8 | 5.2 b | 6.77 | 100 | 7.3 b | 8.37 | 0.008 |
| 17 | cis-carveol | 1208 | 1206 | 0 | 0 | – | 5.6 | 1.1 | – | 0 | 0 | – | 33.3 | 0.7 | – | 0.168 |
| 18 | trans-carveol | 1219 | 1215 | 0 | 0 | – | 16.7 | 0.2 | 0.07 | 44.4 | 0.9 | 0.21 | 33.3 | 0.8 | – | 0.066 |
| 19 | bornyl acetate | 1287 | 1285 | 100 | 22.4 a | 5.37 | 100 | 18.7 a | 7.55 | 100 | 14.0 | 5.53 | 100 | 2.6 b | 0.46 | 0.004 |
| 20 | thymol | 1295 | 1297 | 0 | 0 | – | 5.6 | 0.5 | – | 11.1 | 9.5 | – | 33.3 | 3.8 | – | 0.330 |
| 21 | carvacrol | 1306 | 1308 | 0 | 0 | – | 11.1 | 23.3 | 0.63 | 11.1 | 6.4 | – | 66.7 | 3.2 | 0.42 | 0.079 |
| 22 | α-copaene | 1372 | 1376 | 0 | 0 | – | 22.2 | 0.3 | 0.09 | 0 | 0 | – | 66.7 | 0.5 | 0.03 | 0.059 |
| 23 | β-bourbonene | 1378 | 1385 | 85.7 | 0.9 | 0.45 | 50.0 | 0.5 | 0.46 | 22.2 | 2.0 | 2.27 | 33.3 | 1.8 | – | 0.056 |
| 24 | β-cubebene | 1380 | 1389 | 57.1 | 14.1 | 8.41 | 38.9 | 21.3 | 10.25 | 22.2 | 20.1 | 5.25 | 0 | 0 | – | 0.467 |
| 25 | β-elemene | 1389 | 1391 | 28.6 | 0.8 a | 0.04 | 83.3 | 1.7 b | 1.45 | 66.7 | 0.8 | 0.57 | 33.3 | 0.4 | – | 0.005 |
| 26 | (E)-β-caryophyllene | 1414 | 1419 | 85.7 | 1.0 | 0.27 | 61.1 | 2.5 | 2.14 | 66.7 | 2.0 | 1.67 | 66.7 | 1.5 | 1.42 | 0.985 |
| 27 | β-copaene | 1424 | 1432 | 14.3 | 0.5 | – | 50.0 | 0.6 | 0.19 | 22.2 | 3.8 | 4.56 | 66.7 | 3.2 | 0.79 | 0.161 |
| 28 | α-humulene | 1448 | 1452 | 42.9 | 0.5 | 0.08 | 61.1 | 0.8 | 0.54 | 33.3 | 1.9 | 1.89 | 33.3 | 0.4 | – | 0.380 |
| 29 | γ-muurolene | 1475 | 1477 | 100 | 0.8 a | 0.23 | 0 | 0 b | – | 0 | 0 b | – | 33.3 | 0.7 | – | <0.001 |
| 30 | germacrene D | 1480 | 1480 | 42.9 | 5.9 | 5.95 | 50.0 | 19.6 | 15.22 | 66.7 | 8.8 | 5.22 | 66.7 | 5.2 | 6.46 | 0.751 |
| 31 | β-selinene | 1481 | 1486 | 14.3 | 0.6 | – | 38.9 | 0.5 | 0.47 | 0 | 0 | – | 0 | 0 | – | 0.103 |
| 32 | epi-cubebol | 1489 | 1493 | 0 | 0 | – | 0 | 0 | – | 0 | 0 | – | 66.7 | 1.4 | 0.31 | 0.012 |
| 33 | α-muurolene | 1499 | 1499 | 0 | 0 | – | 0 | 0 | – | 0 | 0 | – | 33.3 | 0.3 | – | 0.222 |
| 34 | β-bisabolene | 1505 | 1509 | 0 | 0 | – | 11.1 | 5.0 | 1.05 | 11.1 | 4.7 | – | 33.3 | 1.8 | – | 0.569 |
| 35 | γ-cadinene | 1507 | 1513 | 100 | 1.0 | 0.32 | 5.6 | 0.7 | – | 0 | 0 | – | 0 | 0 | – | <0.001 |
| 36 | cubebol | 1514 | 1515 | 0 | 0 | – | 0 | 0 | – | 0 | 0 | – | 66.7 | 0.7 | 0.10 | 0.014 |
| 37 | δ-cadinene | 1520 | 1524 | 0 | 0 | – | 5.6 | 0.7 | – | 11.1 | 0.9 | – | 33.3 | 0.4 | – | 0.384 |
| 38 | epoxyazulene | 1561 | 1554 | 100 | 6.1 a | 3.22 | 100 | 2.7 | 1.43 | 66.7 | 2.9 b | 2.49 | 33.3 | 0.3 b | – | 0.001 |
| 39 | (E)-nerolidol | 1565 | 1564 | 85.7 | 0.9 a | 0.20 | 38.9 | 0.6 | 0.20 | 0 | 0 b | – | 0 | 0 | – | 0.001 |
| 40 | spathulenol | 1575 | 1576 | 100 | 6.7 a | 6.25 | 88.9 | 1.2 b | 0.98 | 66.7 | 2.1 b | 1.94 | 100 | 3.1 | 1.30 | 0.003 |
| 41 | caryophyllene oxide | 1578 | 1581 | 100 | 3.1 a | 0.77 | 77.8 | 3.9 a | 2.24 | 100 | 10.4 b | 11.18 | 100 | 9.0 | 6.36 | 0.004 |
| 42 | humulene epoxide II | 1607 | 1606 | 57.1 | 2.4 | 1.26 | 38.9 | 2.7 | 1.04 | 22.2 | 3.2 | 0.92 | 66.7 | 5.0 | 1.44 | 0.276 |
| 43 | isospathulenol | 1627 | 1630 | 100 | 6.3 a | 3.11 | 100 | 4.8 a | 2.54 | 66.7 | 3.7 | 2.26 | 0 | 0 b | – | 0.004 |
| Monoterpene hydrocarbons | 100 | 16.6 | 4.08 | 100 | 14.4 | 9.42 | 100 | 17.7 | 10.92 | 100 | 20.1 | 15.28 | 0.634 | |||
| Oxygenated monoterpenes | 100 | 23.1 | 5.64 | 100 | 27.9 | 11.2 | 100 | 35.7 | 13.07 | 100 | 49.0 | 27.81 | 0.072 | |||
| Sesquiterpene hydrocarbons | 100 | 14.9 | 7.85 | 94.4 | 23.3 | 15.3 | 100 | 15.0 | 11.32 | 66.7 | 8.6 | 10.93 | 0.252 | |||
| Oxygenated sesquiterpenes | 100 | 23.3 | 10.03 | 100 | 13.3 | 6.07 | 100 | 16.9 | 12.56 | 100 | 16.9 | 7.63 | 0.181 | |||
| Oxygenated diterpenes | 0 | 0 | – | 5.6 | 0.7 | 0.26 | 11.1 | 0.1 | 0.33 | 0 | 0 | – | 0.375 | |||
| Collection Site | Altitude, m | Latitude N | Longitude E | Number of Accessions | |||
|---|---|---|---|---|---|---|---|
| SG | SC | SN | SV | ||||
| Pavilnys, Vilnius distr. | 215 | 54°40′35″ | 25°23′01″ | 3 | 3 | 1 | – |
| Didieji Pupojai, Vilnius | 206 | 54°42′38″ | 25°23′29″ | 1 | 3 | 3 | 2 |
| Rokantiškės, Vilnius | 203 | 54°40′03″ | 25°22′58″ | 3 | 2 | 1 | – |
| Raudondvaris, Vilnius distr. | 149 | 54°52′32″ | 25°31′08″ | – | 4 | – | – |
| Dvariškės, Vilnius distr. | 149 | 54°49′17″ | 25°16′23″ | – | 3 | 3 | 1 |
| Karklinė, Vilnius distr. | 164 | 54°54′26″ | 25°33′40″ | – | 3 | 1 | – |
| Analytes | Repeatability (Run-to-Run) | Intermediate Precision (Day-to-Day) | ||||||
|---|---|---|---|---|---|---|---|---|
| tR, min | RSD, % | A, % | RSD, % | tR, min | RSD, % | A, % | RSD, % | |
| α-pinene | 11.85 | 0.23 | 24.57 | 0.45 | 11.92 | 0.82 | 24.83 | 1.51 |
| limonene | 15.24 | 0.29 | 9.07 | 0.33 | 15.31 | 0.73 | 9.17 | 1.42 |
| trans-verbenol | 23.20 | 0.22 | 10.36 | 0.41 | 23.36 | 0.92 | 10.54 | 2.21 |
| bornyl acetate | 32.70 | 0.32 | 10.21 | 0.43 | 32.77 | 0.54 | 10.26 | 0.98 |
| β-cubebene | 44.59 | 0.26 | 2.82 | 0.38 | 44.74 | 0.60 | 2.84 | 1.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radušienė, J.; Karpavičienė, B.; Marksa, M.; Ivanauskas, L.; Raudonė, L. Distribution Patterns of Essential Oil Terpenes in Native and Invasive Solidago Species and Their Comparative Assessment. Plants 2022, 11, 1159. https://doi.org/10.3390/plants11091159
Radušienė J, Karpavičienė B, Marksa M, Ivanauskas L, Raudonė L. Distribution Patterns of Essential Oil Terpenes in Native and Invasive Solidago Species and Their Comparative Assessment. Plants. 2022; 11(9):1159. https://doi.org/10.3390/plants11091159
Chicago/Turabian StyleRadušienė, Jolita, Birutė Karpavičienė, Mindaugas Marksa, Liudas Ivanauskas, and Lina Raudonė. 2022. "Distribution Patterns of Essential Oil Terpenes in Native and Invasive Solidago Species and Their Comparative Assessment" Plants 11, no. 9: 1159. https://doi.org/10.3390/plants11091159