
Phytochemical Profiling, Antimicrobial and α-Glucosidase Inhibitory Potential of Phenolic-Enriched Extracts of the Aerial Parts from Echium humile Desf.: In Vitro Combined with In Silico Approach

Abstract
:1. Introduction
2. Results
2.1. Biological Properties
2.1.1. Antimicrobial Activity
2.1.2. Antidiabetic Activity
2.2. HPLC–MS Analysis
2.3. Computational Study
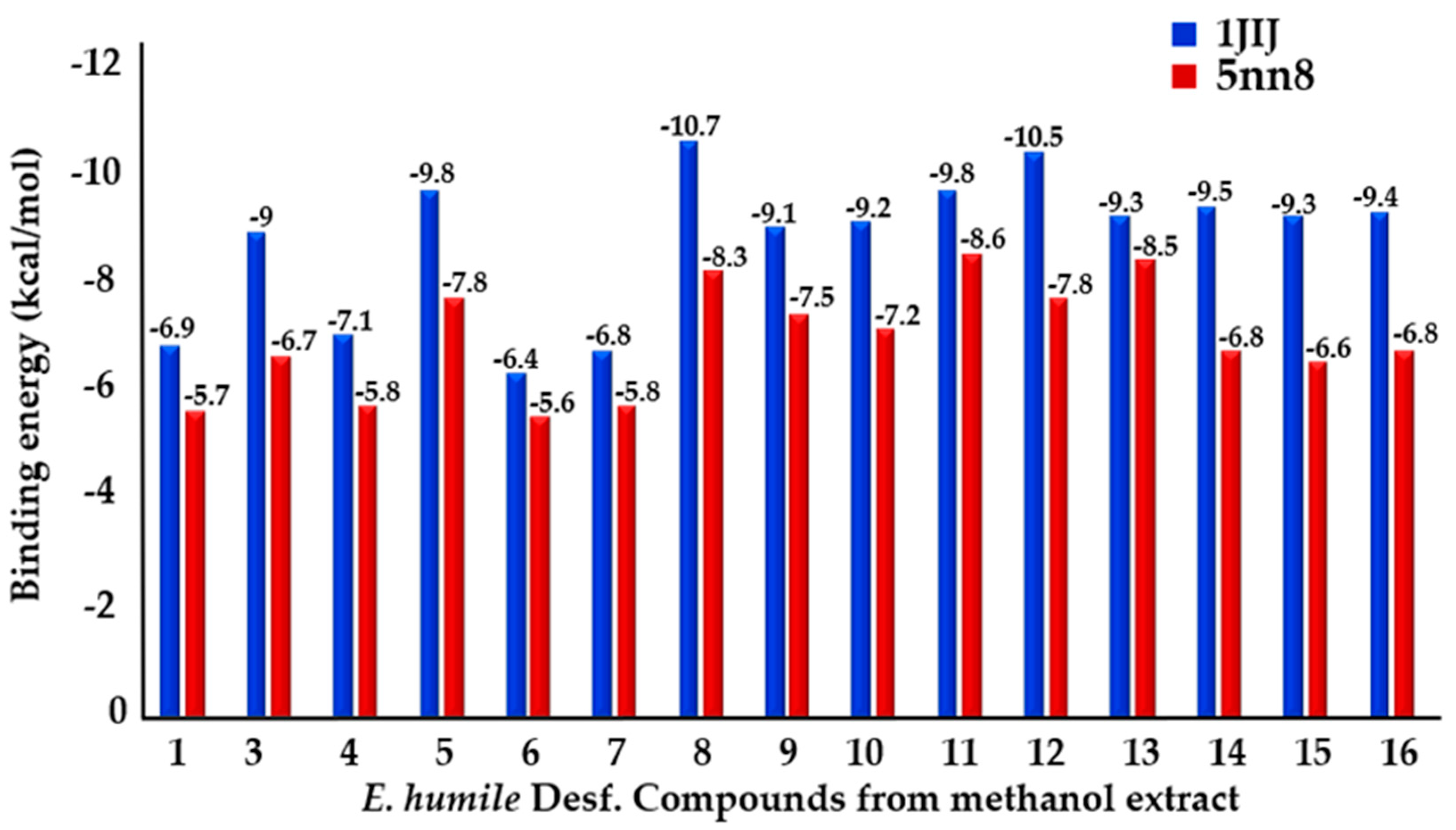
2.3.1. Binding Energies
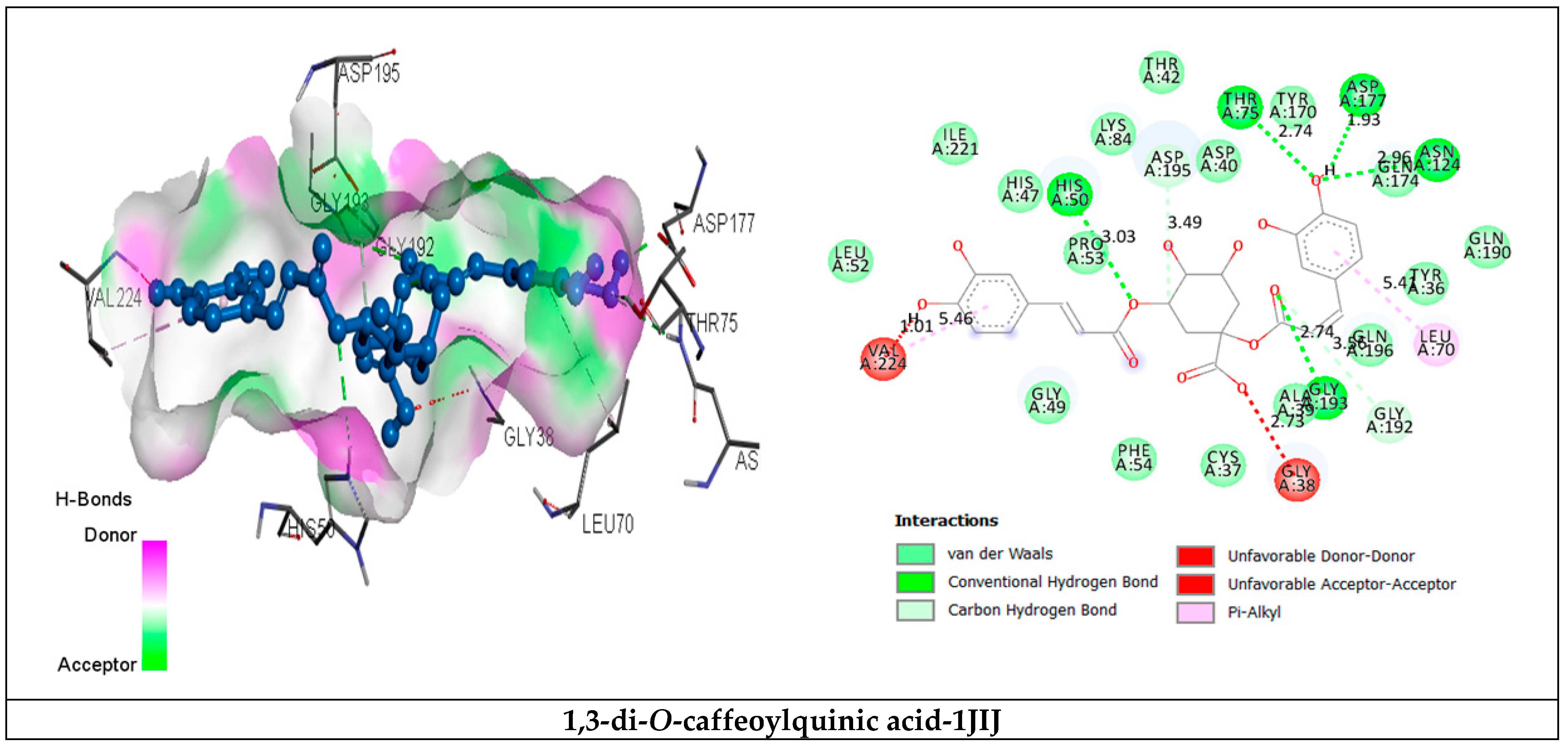
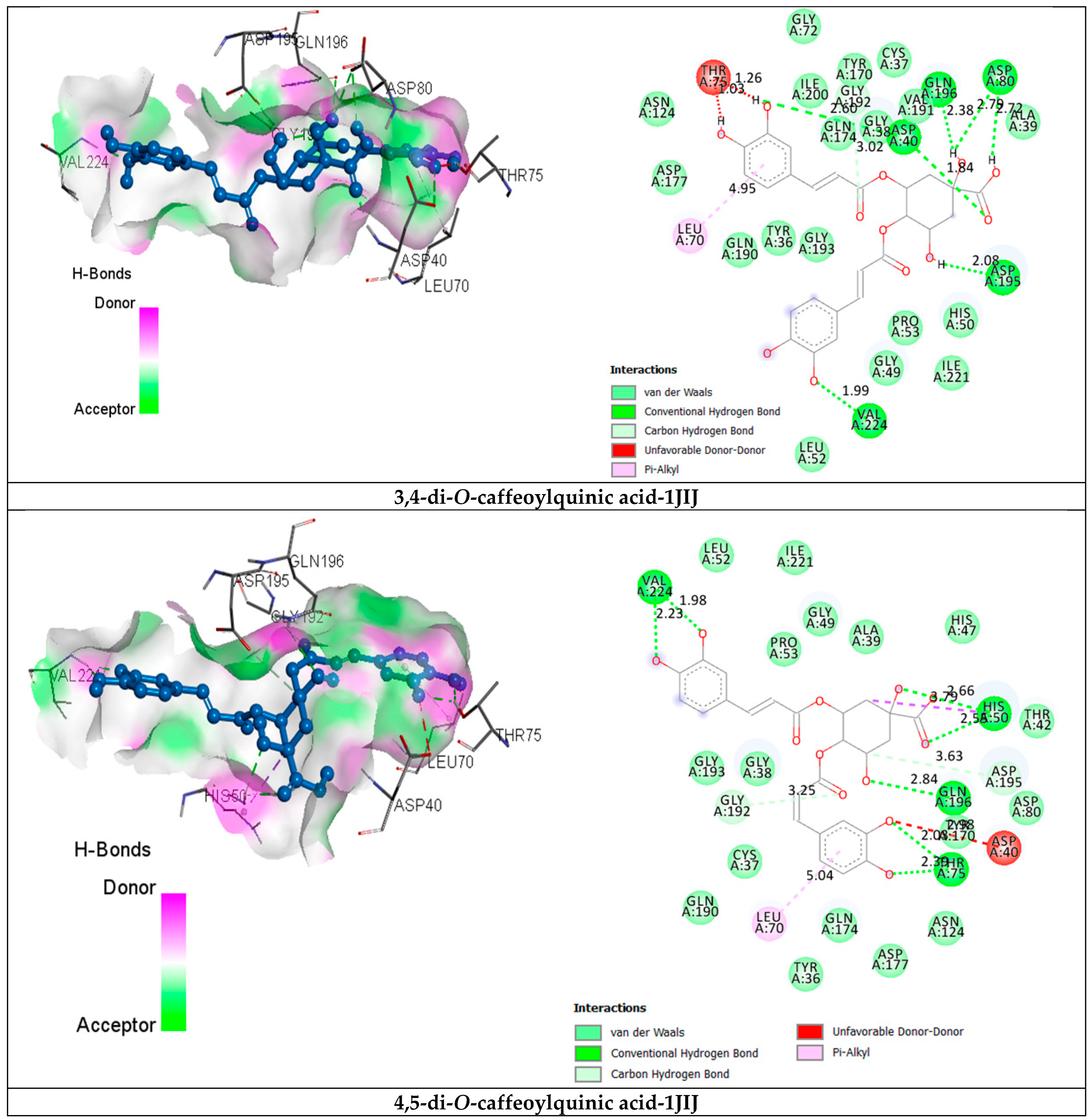
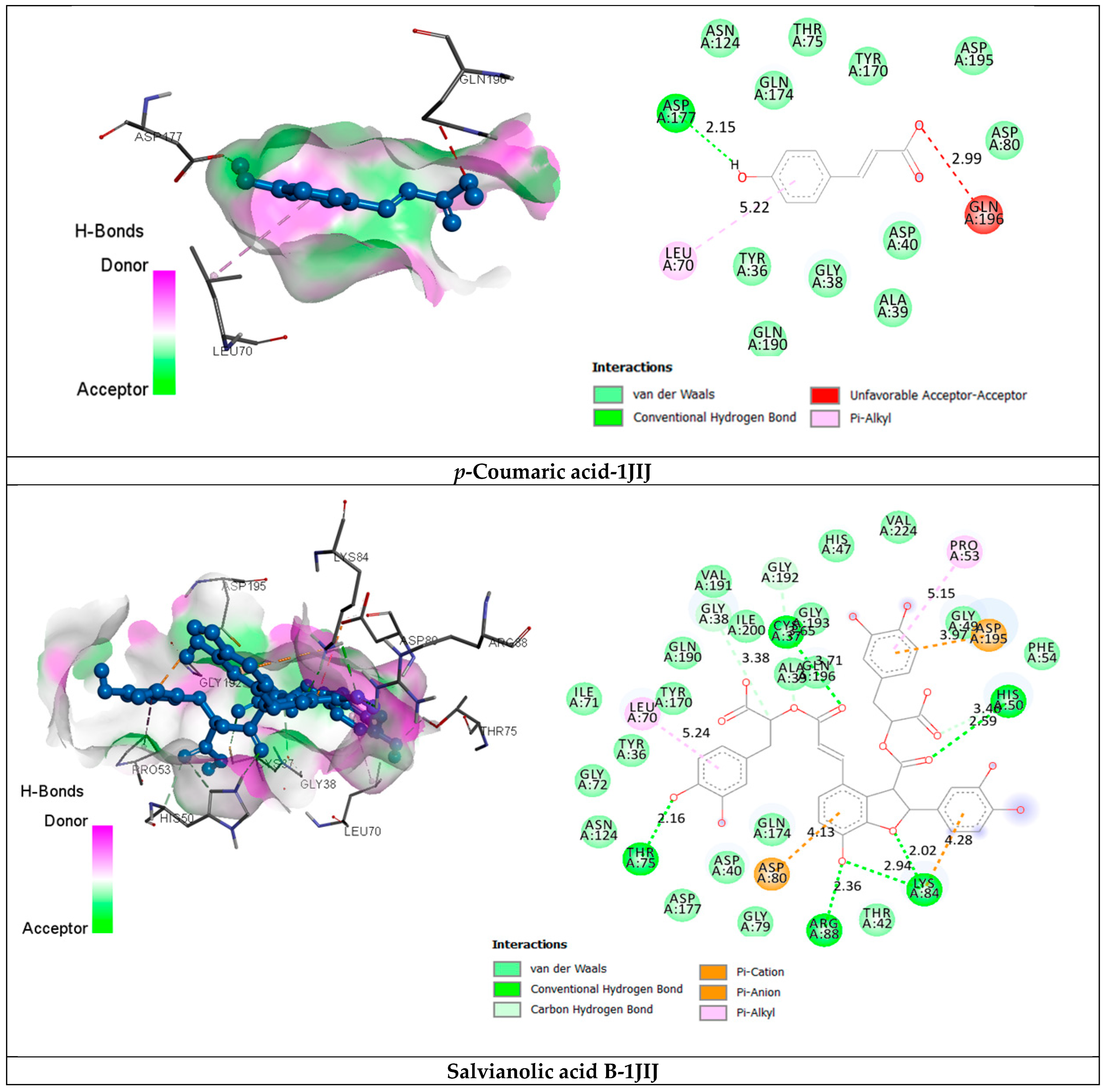
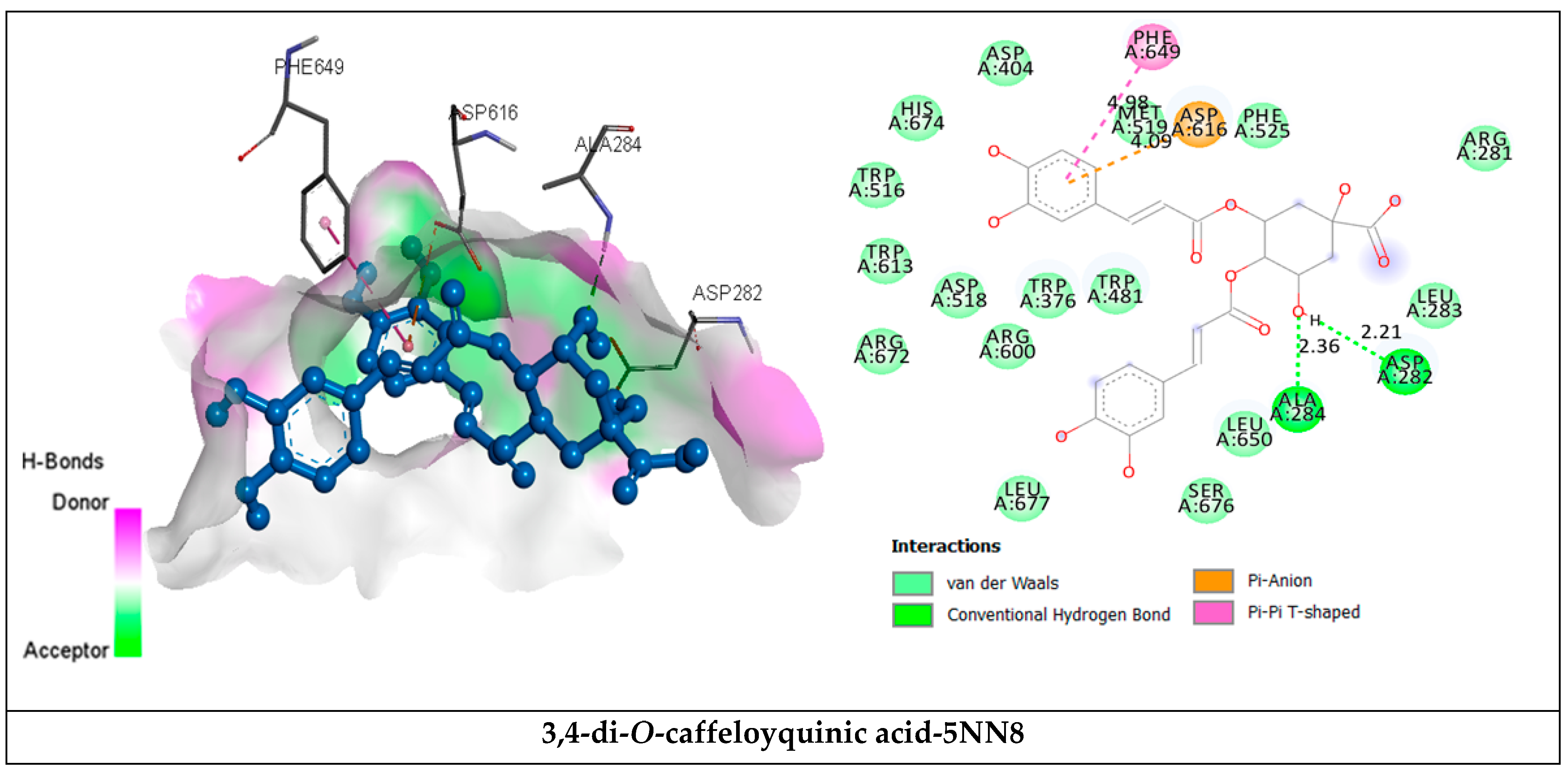
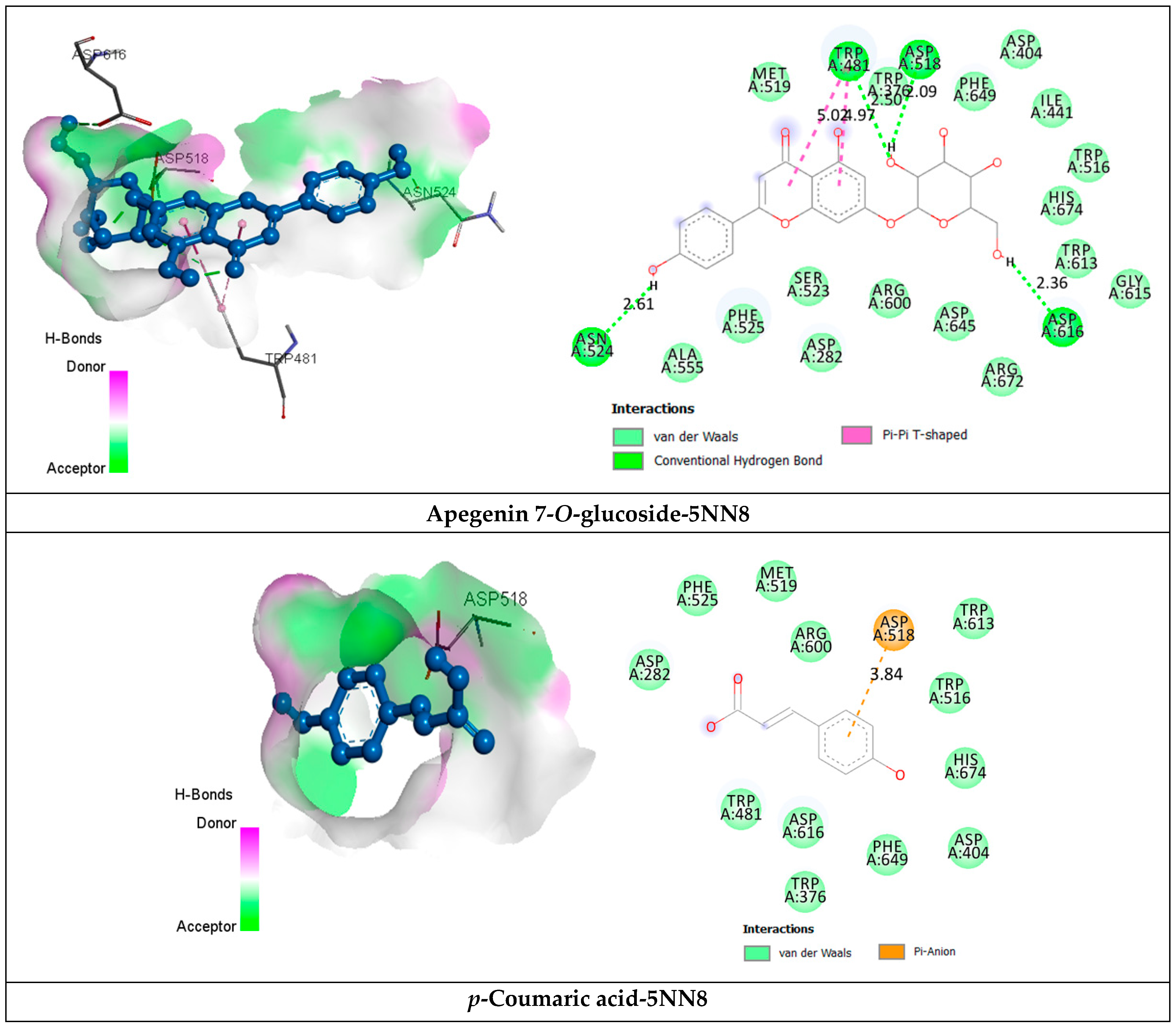
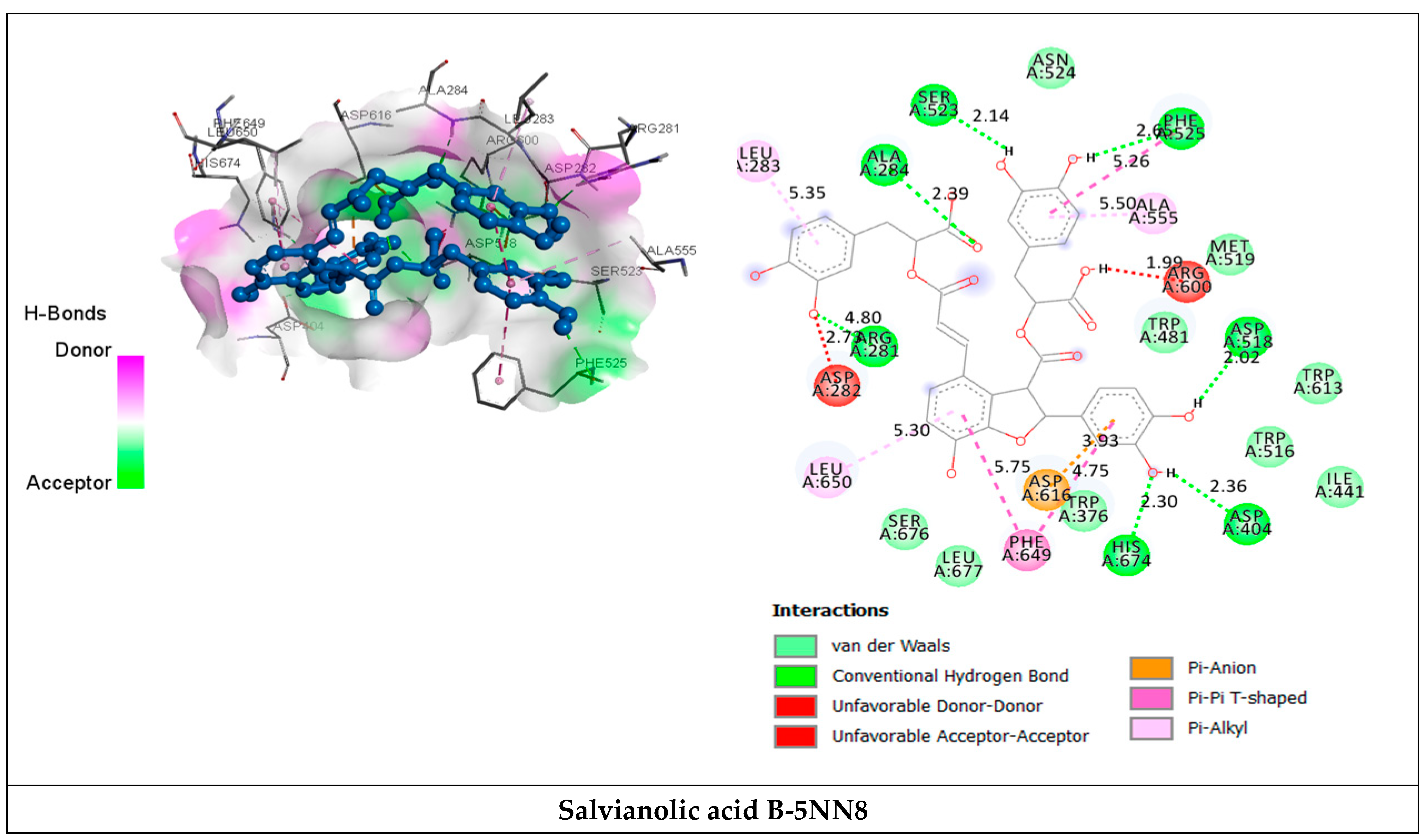
2.3.2. Receptor–Ligand Interaction Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material and Extraction
4.2. Antimicrobial Activity
4.3. α-Glucosidase Inhibitory Assay
4.4. HPLC–MS Analysis of Phenolic Compounds
4.5. Molecular Docking Approach
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zaky, A.S.; Kandeil, M.; Abdel-Gabbar, M.; Fahmy, E.M.; Almehmadi, M.M.; Ali, T.M.; Ahmed, O.M. The Antidiabetic Effects and Modes of Action of the Balanites aegyptiaca Fruit and Seed Aqueous Extracts in NA/STZ-Induced Diabetic Rats. Pharmaceutics 2022, 14, 263. [Google Scholar] [CrossRef] [PubMed]
- Pottathil, S.; Nain, P.; Morsy, M.A.; Kaur, J.; Al-Dhubiab, B.E.; Jaiswal, S.; Nair, A.B. Mechanisms of Antidiabetic Activity of Methanolic Extract of Punica granatum Leaves in Nicotinamide/Streptozotocin-Induced Type 2 Diabetes in Rats. Plants 2020, 9, 1609. [Google Scholar] [CrossRef] [PubMed]
- Asirvatham, R.A.; Mohamed, D.A. Cardiovascular Disease among Patients with Diabetes: The Current Scenario in Saudi Arabia. Curr. Diabetes Rev. 2021, 17, 180. [Google Scholar]
- Huang, C.H.; Chiu, C.H.; Chen, I.W.; Hung, S.Y.; Lin, C.W.; Hsu, B.R.S.; Huang, Y.Y. Antimicrobial resistance and outcomes of community-onset bacterial bloodstream infections in patients with type 2 diabetes. J. Glob. Antimicrob. Resist. 2018, 15, 271. [Google Scholar] [CrossRef]
- Akash, M.S.H.; Rehman, K.; Fiayyaz, F.; Sabir, S.; Kurshid, M. Diabetes-associated infections: Development of antimicrobial resistance and possible treatment strategies. Arch. Microbiol. 2020, 202, 953. [Google Scholar] [CrossRef]
- Xiong, Y.; Ng, K.; Zhang, P.; Warner, R.D.; Shen, S.; Tang, H.-Y.; Liang, Z.; Fang, Z. In Vitro α-Glucosidase and α-Amylase Inhibitory Activities of Free and Bound Phenolic Extracts from the Bran and Kernel Fractions of Five Sorghum Grain Genotypes. Foods 2020, 9, 1301. [Google Scholar] [CrossRef]
- Akinyede, K.A.; Oyewusi, H.A.; Hughes, G.D.; Ekpo, O.E.; Oguntibeju, O.O. In Vitro Evaluation of the Anti-Diabetic Potential of Aqueous Acetone Helichrysum petiolare Extract (AAHPE) with Molecular Docking Relevance in Diabetes Mellitus. Molecules 2022, 27, 155. [Google Scholar] [CrossRef]
- Saddique, F.A.; Ahmad, M.; Ashfaq, U.A.; Muddassar, M.; Sultan, S.; Zaki, M.E.A. Identification of Cyclic Sulfonamides with an N-Arylacetamide Group as α-Glucosidase and α-Amylase Inhibitors: Biological Evaluation and Molecular Modeling. Pharmaceuticals 2022, 15, 106. [Google Scholar] [CrossRef]
- Yuan, S.; Ming-Wei, L.; Qi-Qiang, H.; Larsson, S.C. Egg, cholesterol and protein intake and incident type 2 diabetes mellitus: Results of repeated measurements from a prospective cohort study. Clin. Nutr. 2021, 40, 4180. [Google Scholar] [CrossRef]
- Raoufi, S.; Baluchnejadmojarad, T.; Roghani, M.; Ghazanfari, T.; Khojasteh, F.; Mansouri, M. Antidiabetic potential of salvianolic acid B in multiple low-dose streptozotocin-induced diabetes, Pharm. Biol. 2015, 53, 1803. [Google Scholar]
- Hajlaoui, H.; Arraouadi, S.; Mighri, H.; Ghannay, S.; Aouadi, K.; Adnan, M.; Elasbali, A.M.; Noumi, E.; Snoussi, M.; Kadri, A. HPLC-MS Profiling, Antioxidant, Antimicrobial, Antidiabetic, and Cytotoxicity Activities of Arthrocnemum indicum (Willd.) Moq. Extracts. Plants 2022, 11, 232. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, A.; Singh, B.; Arora, R.; Arora, S. In vitro evaluation of the α-glucosidase inhibitory potential of methanolic extracts of traditionally used antidiabetic plants. BMC Complement. Altern Med. 2019, 19, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajlaoui, H.; Arraouadi, S.; Noumi, E.; Aouadi, K.; Adnan, M.; Khan, M.A.; Kadri, A.; Snoussi, M. Antimicrobial, antioxidant, anti-acetylcholinesterase, antidiabetic, and pharmacokinetic properties of Carum carvi L. and Coriandrum sativum L. essential oils alone and in combination. Molecules 2021, 26, 3625. [Google Scholar] [CrossRef] [PubMed]
- Mseddi, K.; Alimi, F.; Noumi, E.; Veettil, V.N.; Deshpande, S.; Adnan, M.; Hamdi, A.; Elkahoui, S.; Alghamdi, A.; Kadri, A.; et al. Thymus musilii Velen. as a promising source of potent bioactive compounds with its pharmacological properties: In vitro and in silico analysis. Arab. J. Chem. 2020, 13, 6782–6801. [Google Scholar] [CrossRef]
- Ben Mefteh, F.; Daoud, A.; Chenari Bouket, A.; Thissera, B.; Kadri, Y.; Cherif-Silini, H.; Eshelli, M.; Alenezi, F.N.; Vallat, A.; Oszako, T.; et al. Date palm trees root-derived endophytes as fungal cell factories for diverse bioactive metabolites. Int. J. Mol. Sci. 2018, 19, 1986. [Google Scholar] [CrossRef] [Green Version]
- Daoud, A.; Ben Mefteh, F.; Mnafgui, K.; Turki, M.; Jmal, S.; Ben Amar, R.; Ayadi, F.; ElFeki, A.; Abid, L.; Rateb, M.E.; et al. Cardiopreventive effect of ethanolic extract of date palm pollen against isoproterenol induced myocardial infarction in rats through the inhibition of the angiotensin-converting enzyme. Exp. Toxicol. Pathol. 2017, 69, 656–665. [Google Scholar] [CrossRef] [Green Version]
- Hajlaoui, H.; Mighri, H.; Aouni, M.; Gharsallah, N.; Kadri, A. Chemical composition and in vitro evaluation of antioxidant antimicrobial cytotoxicity and anti-acetylcholinesterase properties of Tunisian Origanum majorana L. essential oil. Microb. Pathog. 2016, 95, 86–94. [Google Scholar] [CrossRef]
- Kadri, A.; Zarai, Z.; Chobba, I.B.; Gharsallah, N.; Damak, M.; Békir, A. Chemical composition and in vitro antioxidant activities of Thymelaea hirsuta L: Essential oil from Tunisia. Afr. J. Biotechnol. 2011, 10, 2930–2935. [Google Scholar]
- Felhi, S.; Saoudi, M.; Daoud, A.; Hajlaoui, H.; Ncir, M.; Chaabane, R.; El Feki, A.; Gharsallah, N.; Kadri, A. Investigation of phytochemical contents, in vitro antioxidant and antibacterial behavior and in vivo anti-inflammatory potential of Ecballium elaterium methanol fruits extract. Food Sci. Technol. 2017, 37, 558–563. [Google Scholar] [CrossRef] [Green Version]
- Bakari, S.; Daoud, A.; Felhi, S.; Smaoui, S.; Gharsallah, N.; Kadri, A. Proximate analysis, mineral composition, phytochemical contents antioxidant and antimicrobial activities and GC-MS investigation of various solvent extracts of Cactus cladode. Food Sci Technol. 2017, 27, 286–293. [Google Scholar] [CrossRef] [Green Version]
- Bakari, S.; Hajlaoui, H.; Daoud, A.; Mighri, H.; Ross-Garcia, J.M.; Gharsallah, N.; Kadri, A. Phytochemicals, antioxidant and antimicrobial potentials and LC-MS analysis of hydroalcoholic extracts of leaves and flowers of Erodium glaucophyllum collected from Tunisian Sahara. Food Sci. Technol. 2018, 38, 310–317. [Google Scholar] [CrossRef] [Green Version]
- Felhi, S.; Hajlaoui, H.; Ncir, M.; Bakari, S.; Ktari, N.; Saoudi, M.; Gharsallah, N.; Kadri, A. Nutritional, phytochemical and antioxidant evaluation and FT-IR analysis of freeze-dried extracts of Ecballium elaterium fruit juice from three localities. Food Sci. Technol. 2016, 36, 646–655. [Google Scholar] [CrossRef] [Green Version]
- Bakari, S.; Ncir, M.; Felhi, S.; Hajlaoui, H.; Saoudi, M.; Gharsallah, N.; Kadri, A. Chemical composition and in vitro evaluation of total phenolic, flavonoid, and antioxidant properties of essential oil and solvent extract from the aerial parts of Teucrium polium grown in Tunisia. Food Sci Biotechnol. 2015, 24, 1943–1949. [Google Scholar] [CrossRef]
- Alminderej, F.; Bakari, S.; Almundarij, T.I.; Snoussi, M.; Aouadi, K.; Kadri, A. Antioxidant activities of a new chemotype of Piper cubeba L. fruit essential oil (Methyleugenol/Eugenol): In Silico molecular docking and ADMET studies. Plants 2020, 9, 1534. [Google Scholar] [CrossRef]
- Noumi, E.; Snoussi, M.; Anouar, E.H.; Alreshidi, M.; Veettil, V.N.; Elkahoui, S.; Adnan, M.; Patel, M.; Kadri, A.; Aouadi, K.; et al. HR-LCMS-Based Metabolite Profiling, Antioxidant, and Anticancer Properties of Teucrium polium L. Methanolic Extract: Computational and In Vitro Study. Antioxidants 2020, 9, 1089. [Google Scholar] [CrossRef]
- Aouadi, K.; Hajlaoui, H.; Arraouadi, S.; Ghannay, S.; Snoussi, M.; Kadri, A. HPLC/MS phytochemical profiling with antioxidant activities of Echium humile Desf. extracts: ADMET prediction and computational study targeting human peroxiredoxin 5 receptor. Agronomy 2021, 11, 2165. [Google Scholar] [CrossRef]
- Alminderej, F.; Bakari, S.; Almundarij, T.I.; Snoussi, M.; Aouadi, K.; Kadri, A. Antimicrobial and wound healing potential of a new chemotype from Piper cubeba L. essential oil and in silico study on S. aureus tyrosyl-tRNA synthetase protein. Plants 2021, 10, 205. [Google Scholar] [CrossRef]
- Othman, I.M.M.; Gad-Elkareem, M.A.M.; Anouar, E.H.; Aouadi, K.; Kadri, A.; Snoussi, M. Design, synthesis ADMET and molecular docking of new imidazo[4,5-b]pyridine-5-thione derivatives as potential tyrosyl-tRNA synthetase inhibitors. Bioorg. Chem. 2020, 102, 104105. [Google Scholar] [CrossRef]
- Othman, I.M.M.; Gad-Elkareem, M.A.M.; Anouar, E.H.; Aouadi, K.; Snoussi, M.; Kadri, A. New substituted pyrazolones and dipyrazolotriazines as promising tyrosyl-tRNA synthetase and peroxiredoxin-5 inhibitors: Design, synthesis, molecular docking and structure-activity relationship (SAR) analysis. Bioorg. Chem. 2021, 109, 104704. [Google Scholar] [CrossRef]
- Othman, I.M.M.; Gad-Elkareem, M.A.M.; Anouar, E.H.; Snoussi, M.; Aouadi, K.; Kadri, A. Novel fused pyridine derivatives containing pyrimidine moiety as prospective tyrosyl-tRNA synthetase inhibitors: Design, synthesis, pharmacokinetics and molecular docking studies. J. Mol. Struct. 2020, 1219, 128651. [Google Scholar] [CrossRef]
- Wong, J.X.; Ramli, S. Antimicrobial activity of different types of Centella asiatica extracts against foodborne pathogens and food spoilage microorganisms. LWT 2021, 142, 111026. [Google Scholar] [CrossRef]
- Kalita, D.; Saikia, J. Ethnomedicinal, antibacterial and antifungal potentiality of Centella asiatica, Nerium indicum and Cuscuta reflexa widely used in Tiwa Tribe of Morigaon district of Assam, India. Int. J. Phytomed. 2012, 4, 380. [Google Scholar]
- Rattanakom, S.; Yasurin, P. Chemical profiling of Centella asiatica under different extraction solvents and its antibacterial activity, antioxidant activity. Orient. J. Chem. 2015, 31, 2453. [Google Scholar] [CrossRef]
- Werner, G.; Coque, T.M.; Franz, C.M.A.P.; Grohmann, E.; Hegstad, K.; Jensen, L.; Schaik, W.; Weaver, K. Antibiotic resistant enterococci—Tales of a drug resistance gene trafficker. Int. J. Med. Microbiol. 2013, 303, 360. [Google Scholar] [CrossRef] [PubMed]
- Lou, Z.; Wang, H.; Rao, S.; Sun, J.; Ma, C.; Li, J. p-Coumaric acid kills bacteria through dual damage mechanisms. Food Control 2012, 25, 550. [Google Scholar] [CrossRef]
- Roychoudhury, S.; Sinha, B.; Choudhury, B.P.; Jha, N.K.; Palit, P.; Kundu, S.; Mandal, S.C.; Kolesarova, A.; Yousef, M.I.; Ruokolainen, J.; et al. Scavenging Properties of Plant-Derived Natural Biomolecule Para-Coumaric Acid in the Prevention of Oxidative Stress-Induced Diseases. Antioxidants 2021, 10, 1205. [Google Scholar] [CrossRef]
- Amalan, V.; Vijayakumar, N.; Indumathi, D.; Ramakrishnan, A. Antidiabetic and antihyperlipidemic activity of p-coumaric acid in diabetic rats, role of pancreatic GLUT 2: In vivo approach. Biomed. Pharmacother. 2016, 84, 230. [Google Scholar] [CrossRef]
- Shen, Y.; Song, X.; Li, L.; Sun, J.; Jaiswal, Y.; Huang, J.; Liu, C.; Yang, W.; Williams, L.; Zhang, H.; et al. Protective effects of p-coumaric acid against oxidant and hyperlipidemia-an in vitro and in vivo evaluation. Biomed. Pharmacother. 2019, 111, 579. [Google Scholar] [CrossRef]
- Georgiev, L.; Chochkova, M.; Ivanova, G.; Najdenski, H.; Ninova, M.; Milkova, T. Radical scavenging and antimicrobial activities of cinnamoyl amides of biogenic monoamines. Riv. Ital. Sost. Grasse 2012, 89, 91–102. [Google Scholar]
- Song, Y.; Wen, L.; Sun, J.; Bai, W.; Jiao, R.; Hu, Y.; Peng, X.; He, Y.; Ou, S. Cytoprotective mechanism of ferulic acid against high glucose-induced oxidative stress in cardiomyocytes and hepatocytes. Food Nut. Res. 2016, 60, 30323. [Google Scholar] [CrossRef] [Green Version]
- Senaphan, K.; Kukongviriyapan, U.; Sangartit, W.; Pakdeechote, P.; Pannangpetch, P.; Prachaney, P.; Greenwald, S.E.; Kukongviriyapan, V. Ferulic Acid Alleviates Changes in a Rat Model of Metabolic Syndrome Induced by High-Carbohydrate, High-Fat Diet. Nutrients 2015, 7, 6446–6464. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wu, T.; Yang, Q.; Chen, X.; Wang, M.; Wang, Y.; Peng, X.; Ou, S. Ferulic acid alleviates the symptoms of diabetes in obese rats. J. Funct. Foods. 2014, 9, 141. [Google Scholar] [CrossRef]
- Ma, L.; Tang, L.; Yi, Q. Salvianolic Acids: Potential Source of Natural Drugs for the Treatment of Fibrosis Disease and Cancer. Front. Pharmacol. 2019, 10, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekambaram, S.P.; Perumal, S.S.; Balakrishnan, A.; Marappan, N.; Gajendran, S.S.; Viswanathan, V. Antibacterial synergy between rosmarinic acid and antibiotics against methicillin-resistant Staphylococcus aureus. J. Intercult. Ethnopharmacol. 2016, 5, 358–363. [Google Scholar] [CrossRef]
- Amoah, S.K.; Sandjo, L.P.; Kratz, J.M.; Biavatti, M.W. Rosmarinic Acid—Pharmaceutical and Clinical Aspects. Planta Med. 2016, 82, 388–406. [Google Scholar] [CrossRef] [Green Version]
- Mushtaq, N.; Schmatz, R.; Ahmed, M.; Pereira, L.B.; da Costa, P.; Reichert, K.P.; Dalenogare, D.; Pelinson, L.P.; Vieira, J.M.; Stefanello, N.; et al. Protective e_ect of rosmarinic acid against oxidative stress biomarkers in liver and kidney of strepotozotocin-induced diabetic rats. J. Physiol. Biochem. 2015, 71, 743–751. [Google Scholar] [CrossRef]
- Sotnikova, R.; Kaprinay, B.; Navarova, J. Rosmarinic acid mitigates signs of systemic oxidative stress in streptozotocin-induced diabetes in rats. Gen. Physiol. Biophys. 2015, 34, 449–452. [Google Scholar]
- Kuete, V.; More, G.; Lall, N.; Hussein, A.; Tshikalange, T.E. Antimicrobial Constituents of Artemisia afra Jacq. ex Willd. against Periodontal Pathogens. Evid Based Complement. Alternat. Med. 2012, 2012, 252758. [Google Scholar]
- Komape, N.P.M.; Aderogba, M.; Bagla, V.P.; Masoko, P.; Eloff, J.N. Antibacterial and antioxidant activities of leaf extracts of Combretum vendae (combretecacea) and the isolation of an antibacterial compound. Afr. J. Tradit Complement. Altern. Med. 2014, 11, 73. [Google Scholar] [CrossRef] [Green Version]
- Juárez-Reyes, K.; Brindis, F.; Medina-Campos, O.N.; Pedraza-Chaverri, J.; Bye, R.; Linares, E.; Mata, R. Hypoglycemic, antihyperglycemic, and antioxidant effects of the edible plant Anoda cristata. J. Ethnopharmacol. 2015, 161, 36. [Google Scholar] [CrossRef]
- Semwal, R.B.; Semwal, D.K.; Combrinck, S.; Trill, J.; Gibbons, S.; Viljoen, A. Acacetin—A simple flavone exhibiting diverse pharmacological activities. Phytochem. Lett. 2019, 32, 56. [Google Scholar] [CrossRef]
- Brown, A.; Anderson, D.; Racicot, K.; Pilkenton, S.J.; Apostolidis, E. Evaluation of phenolic phytochemical enriched commercial plant extracts on the in vitro inhibition of α-glucosidase. Front. Nutr. 2017, 4, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jesus, F.; Gonçalves, A.C.; Alves, G.; Silva, L.R. Exploring the phenolic profile, antioxidant, antidiabetic and anti-hemolytic potential of Prunus avium vegetal parts. Food Res. Int. 2019, 116, 600–610. [Google Scholar] [CrossRef] [PubMed]
- El Shibani, F.A.E.S. A Pharmacognostical Study of Arbutus pavarii Pampan. Family Ericaceae and Sarcopoterium spinosum L. Family Rosaceae Growing in Libya. Ph.D. Thesis, Cairo University, Cairo, Egypt, 2017. [Google Scholar]
- Kadri, A.; Aouadi, K. In vitro antimicrobial and α-glucosidase inhibitory potential of enantiopure cycloalkylglycine derivatives: Insights into their in silico pharmacokinetic, druglikeness, and medicinal chemistry properties. J. App. Pharm. Sci. 2020, 10, 107–115. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | IZD (mm) | ||||||
|---|---|---|---|---|---|---|---|
| Extracts | Hexane | Dichloromethane | Ethyl Acetate | Methanol | Aqueous | Chloramphenicol | |
| Gram-positive bacteria | |||||||
| Staphylococcus aureus | – | 11.50 ± 1.00 cdC | 14.00 ± 2.00 abB | 10.00 ± 0.00 dCD | 9.00 ± 0.00 cD | 16.50 ± 1.00 dA | |
| Enterococcus faecalis | 10.50 ± 1.00 cC | 13.00 ± 1.00 bcB | 12.50 ± 1.00 bcBC | 15.50 ± 1.00 aA | 10.50 ± 1.00 bC | 12.00 ± 1.00 eBC | |
| Bacillus cereus | 14.00 ± 1.00 bB | 14.00 ± 1.00 bB | 12.00±1.00 cC | 14.00±1.00 bB | 11.50 ±1.00 bC | 26.00 ± 1.00 aA | |
| Bacillus subtilis | 11.50 ± 1.00 cC | 14.50 ± 1.00 bB | 15.00 ± 1.00 aB | 12.50 ± 1.00 cC | – | 24.00 ± 0.00 bA | |
| Micrococcus luteus | 18.50 ± 0.86 aA | _ | 14.5 ± 1.00 aB | 14.00 ± 0.00 bB | 15.00 ± 0.00 aB | 20.00 ± 2.00 cA | |
| Gram-negative bacteria | |||||||
| Escherichia coli | 10.33 ± 0.76 cC | 18.00 ± 1.00 aB | 8.00 ± 0.00 eD | – | – | 23.50 ± 1.50 bA | |
| Klebsiella pneumoniae | 11.00 ± 0.00 c | 11.00 ± 0.00 dB | 11.50 ± 1.50 cdB | – | 10.50±0.50 bB | 22.00 ± 1.00 bA | |
| Salmonella Enteritidis | 8.00 ± 0.00 dE | 11.50 ± 0.50 cdB | 10.00 ± 0.00 dC | 11.50 ± 0.50 cB | 9.00 ± 0.00 cD | 16.00 ± 0.00 dA | |
| Fungal strains | Cycloheximide | ||||||
| Fusarium sp. | – | – | 11.5 ± 0.71 bC | 14.5 ± 0.71 bB | – | 18.00 ± 1.50 abA | |
| Pythium catenulatum | – | – | 9.00 ± 0.00 cB | 10.50 ± 0.71 cB | – | 17.50 ± 1.50 bA | |
| Fusarium oxysporum | – | – | 13.00 ± 1.00 aB | 20.00 ± 0.00 aA | – | 20.00 ± 2.00 aA | |
| Strains | Hexane | Dichloromethane | Ethyl Acetate | Methanol | Aqueous |
|---|---|---|---|---|---|
| Gram-positive bacteria | |||||
| Staphylococcus aureus | – | 12.5 (25) {2} | 1.56 (12.5) {8} | 3.12 (6.25) {2} | 3.12 (6.25) {2} |
| Enterococcus faecalis | 6.25 (12.5) {2} | 12.5 (50) {4} | 1.56 (12.5) {8} | 0.19 (3.12) {16} | 0.19 (6.25) {32} |
| Bacillus cereus | 0.39 (6.25) {16} | 6.25 (25) {4} | 12.5 (100) {8} | 6.25 (12.5) {2} | – |
| Bacillus subtilis | 6.25 (25) {4} | 50 (100) {2} | 1.56 (12.5) {8} | 0.39 (6.25) {16} | 0.19 (3.12) {16} |
| Micrococcus luteus | 12.5 (50) {4} | – | 1.56 (6.25) {4} | 6.25 (25) {4} | 3.12 (6.25) {2} |
| Gram-negative bacteria | |||||
| Escherichia coli | 3.12 (12.5) {4} | 1.56 (6.25) {4} | 1.56 (12.5) {8} | – | – |
| Klebsiella pneumoniae | 50 (100) {2} | 25 (50) {2} | 12.5 (25) {2} | – | 3.12 (6.25) {2} |
| Salmonella Enteritidis | 1.56 (3.12) {2} | 3.12 (12.5) {4} | 1.56 (3.12) {2} | 0.39 (3.12) {8} | 1.56 (6.25) {4} |
| Fungal strains | |||||
| Fusarium sp. | – | – | 0.39 (3.12) {8} | 0.39 (3.12) {8} | – |
| Pythium catenulatum | _ | _ | 1.56 (6.25) {4} | 0.195 (0.78) {4} | – |
| Fusarium oxysporum | – | – | 0.39 (1.56) {4} | 3.12 (6.25) {2} | – |
| Extracts | IC50 (mg/mL) |
|---|---|
| Hexane | – |
| Dichloromethane | – |
| Ethyl acetate | 11.17 ± 0.62 a |
| Methanol | 0.06 ± 0.29 c |
| Aqueous | 0.80 ± 1.81 b |
| Acarbose | 0.70 ± 0.67 b |
| Peak | Retention Time (min) | MS [M-H]-m/z | Compounds | Concentration (µg/g) |
|---|---|---|---|---|
| 1 | 7.385 | 153.00 | Protocatechuic acid | 4.204 ± 0.06 |
| 2 | 9.189 | 289.00 | (+)-Catechin | 39.286 ± 2.46 |
| 3 | 13.795 | 289.00 | Epicatechin | 26.868 ± 0.82 |
| 4 | 12.993 | 179.00 | Caffeic acid | 9.673 ± 0.1 |
| 5 | 14.960 | 515.00 | 1,3-di-O-caffeoylquinic acid | 22.367 ± 0.15 |
| 6 | 17.087 | 163.00 | p-Coumaric acid | 1335.48 ± 5.22 |
| 7 | 18.744 | 193.00 | Trans-Ferulic acid | 125.522 ± 10.3 |
| 8 | 21.779 | 515.00 | 3,4-di-O-caffeoylquinic acid | 6.7645 ± 0.41 |
| 9 | 22.209 | 359.00 | Rosmarinic acid | 41.154 ± 0.12 |
| 10 | 22.910 | 463.00 | Hyperoside | 2.992 ± 0.01 |
| 11 | 23.754 | 717.00 | Salvianolic acid B | 47.832 ± 1.51 |
| 12 | 23.754 | 515.00 | 4,5-di-O-caffeoylquinic acid | 319.373 ± 0.41 |
| 13 | 24.302 | 431.00 | Apegenin 7-O-glucoside | 2.481 ± 0.48 |
| 14 | 23.451 | 329.00 | Cirsiliol | 41.623 ± 0.19 |
| 15 | 31.852 | 269.00 | Apegenin | 0.973 ± 0.25 |
| 16 | 37.061 | 283.00 | Acacetin | 107.462 ± 17.4 |
| Compounds | Interactions Type | Interacting Residues (1JIJ) | Binding Energy (kcal/mol) |
|---|---|---|---|
| 1,3-di-O-caffeoylquinic acid | van der Waals H bond C-H bond Unfavorable Donor-Donor/ Acceptor-Acceptor Pi-Alkyl | Tyr36, Cys37, Ala39, Thr42, His47, Gly49, Leu52, Pro53, Phe54, Lys84, Tyr170, Gln174, Gln190, Gln196, Ile221. His50 (3.03), Thr75 (2.74), Asp177 (1.93), Asn124 (2.96), Gly193 (2.74). Gly192 (3.56), Asp195 (3.38). Gly38 (2.77), Val224 (2.01). Leu70 (5.41), Val224 (5.46). | −9.8 |
| 3,4-di-O-caffeoylquinic acid | van der Waals H bond C-H bond Unfavorable Donor-Donor Pi-Alkyl | Tyr36, Cys37, Gly38, Ala39, Gly49, His50, Leu52, Pro53, Gly72, Asn124, Tyr170, Gln174, Asp177, Gln190, Val191, Gly193, Ile200, Ile221. Asp40 (1.84) (2.60), Asp80 (2.72) (2.79), Asp195 (2.08), Gln196 (2.38), Val224 (1.99). Gly192 (3.02). Thr75 (1.03) (1.26). Leu70 (4.95). | −10.7 |
| 4,5-di-O-caffeoylquinic acid | van der Waals H bond C-H bond Unfavorable Donor-Donor Pi-Alkyl | Tyr36, Cys37, Gly38, Ala39, Thr42, His47, Gly49, Leu52, Pro53, Asp80, Asn124, Tyr170, Gln174, Asp177, Gln190, Gly193, Ile221. His50 (2.55) (2.66), Thr75 (2.03) (2.39), Gln196 (2.84), Val224 (1.98) (2.23). Asp195 (3.36) Asp40 (2.98) Leu70 (5.04) | −10.5 |
| p-Coumaric acid | van der Waals C-H bond Pi-Cation Pi-Alkyl | Tyr36, Gly38, Ala39, Asp40, Thr75, Asp80, Asn124, Tyr170, Gln174, Gln190, Asp195. Asp177 (2.15) Gln196 (2.99) Leu70 (5.22) | −6.4 |
| Salvianolic Acid B | van der Waals H bond C-H bond Pi-Cation/Anion Pi-Alkyl | Tyr36, Ala39, Asp40, Thr42, His47, Gly49, Phe54, Ile71, Gly72, Gly79, Asn124, Tyr170, Gln174, Asp177, Gln190, Val191, Gly193, Gln196, Ile200, Val224. Cys37 (3.71), His50 (2.59), Thr75 (2.16), Lys84 (2.02) (2.94), Arg88 (2.36). Gly38 (3.38), His50 (3.46), Gly192 (3.65). Asp80 (4.13), Lys84 (4.28), Asp195 (3.97) Pro53 (5.15), Leu70 (5.24) | −9.8 |
| Compounds | Interactions Type | Interacting Residues (5NN8) (Å) | Binding Energy (kcal/mol) |
|---|---|---|---|
| 3,4-di-O-caffeoylquinic acid | van der Waals H bond Pi-Anion Pi-Pi T shaped | Arg281, Leu283, Trp376, Asp404, Trp481, Asp518, Met519, Phe525, Trp516, Arg600, Trp613, Arg672, leu650, His674, Ser676, Leu677. Asp282(2.21), Ala284 (2.36). Asp616 (4.09) Phe649 (4.98) | −8.3 |
| p-Coumaric acid | van der Waals Pi-Anion | Asp282, Trp376, Asp404, Trp481, Trp516, Met519, Phe525, Arg600, Trp613, Asp616, Phe649, His674. Asp518 (3.84). | −5.6 |
| Salvianolic acid B | van der Waals H bond Unfavorable Donor-Donor/ Acceptor-Acceptor Pi-Anion Pi-Pi T shaped | Trp376, Ile441, Trp481, Trp516, Met519, Asn524, Trp613, Ser676, Leu677. Arg281 (4.80), Ala284 (2.39), Asp404 (2.36), Asp518 (2.02), Ser523 (2.14), Phe525 (2.6), His674 (2.30). Asp282 (2.73), Arg600 (1.99). Asp616 (3.93). Phe649 (4.75) (5.75). | −8.6 |
| Apegenin 7-O-glucoside | van der Waals H bond Pi-Pi T shaped Pi-Alkyl | Asp282, Trp376, Ile441, Trp516, Met519, Ser523, Phe525, Ala555, Arg600, Trp613, Gly615, Asp645, Phe649, Arg672, His674. Trp481 (2.50), Asp518 (2.09), Asn524 (2.61), Asp616 (2.36). Trp481 (4.97) (5.02). Leu283 (5.35), Ala555 (5.50), Leu650 (5.30). | −8.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aouadi, K.; Hajlaoui, H.; Arraouadi, S.; Ghannay, S.; Snoussi, M.; Kadri, A. Phytochemical Profiling, Antimicrobial and α-Glucosidase Inhibitory Potential of Phenolic-Enriched Extracts of the Aerial Parts from Echium humile Desf.: In Vitro Combined with In Silico Approach. Plants 2022, 11, 1131. https://doi.org/10.3390/plants11091131
Aouadi K, Hajlaoui H, Arraouadi S, Ghannay S, Snoussi M, Kadri A. Phytochemical Profiling, Antimicrobial and α-Glucosidase Inhibitory Potential of Phenolic-Enriched Extracts of the Aerial Parts from Echium humile Desf.: In Vitro Combined with In Silico Approach. Plants. 2022; 11(9):1131. https://doi.org/10.3390/plants11091131
Chicago/Turabian StyleAouadi, Kaïss, Hafedh Hajlaoui, Soumaya Arraouadi, Siwar Ghannay, Mejdi Snoussi, and Adel Kadri. 2022. "Phytochemical Profiling, Antimicrobial and α-Glucosidase Inhibitory Potential of Phenolic-Enriched Extracts of the Aerial Parts from Echium humile Desf.: In Vitro Combined with In Silico Approach" Plants 11, no. 9: 1131. https://doi.org/10.3390/plants11091131