
The Biodiversity of Grapevine Bacterial Endophytes of Vitis amurensis Rupr.



Abstract
:1. Introduction
2. Results
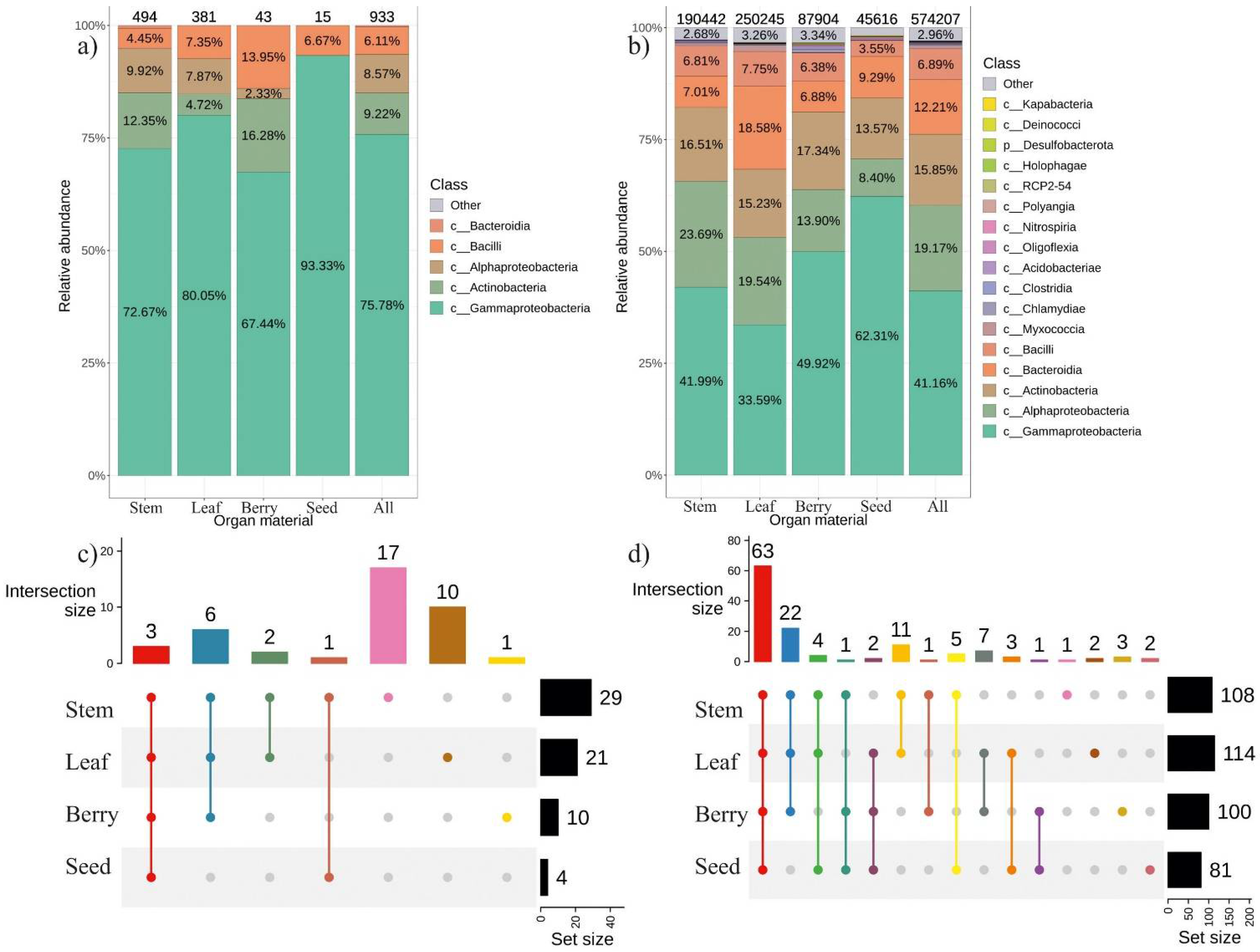
2.1. The General Composition of Endophytic Bacterial Community in Different Organ of V. amurensis
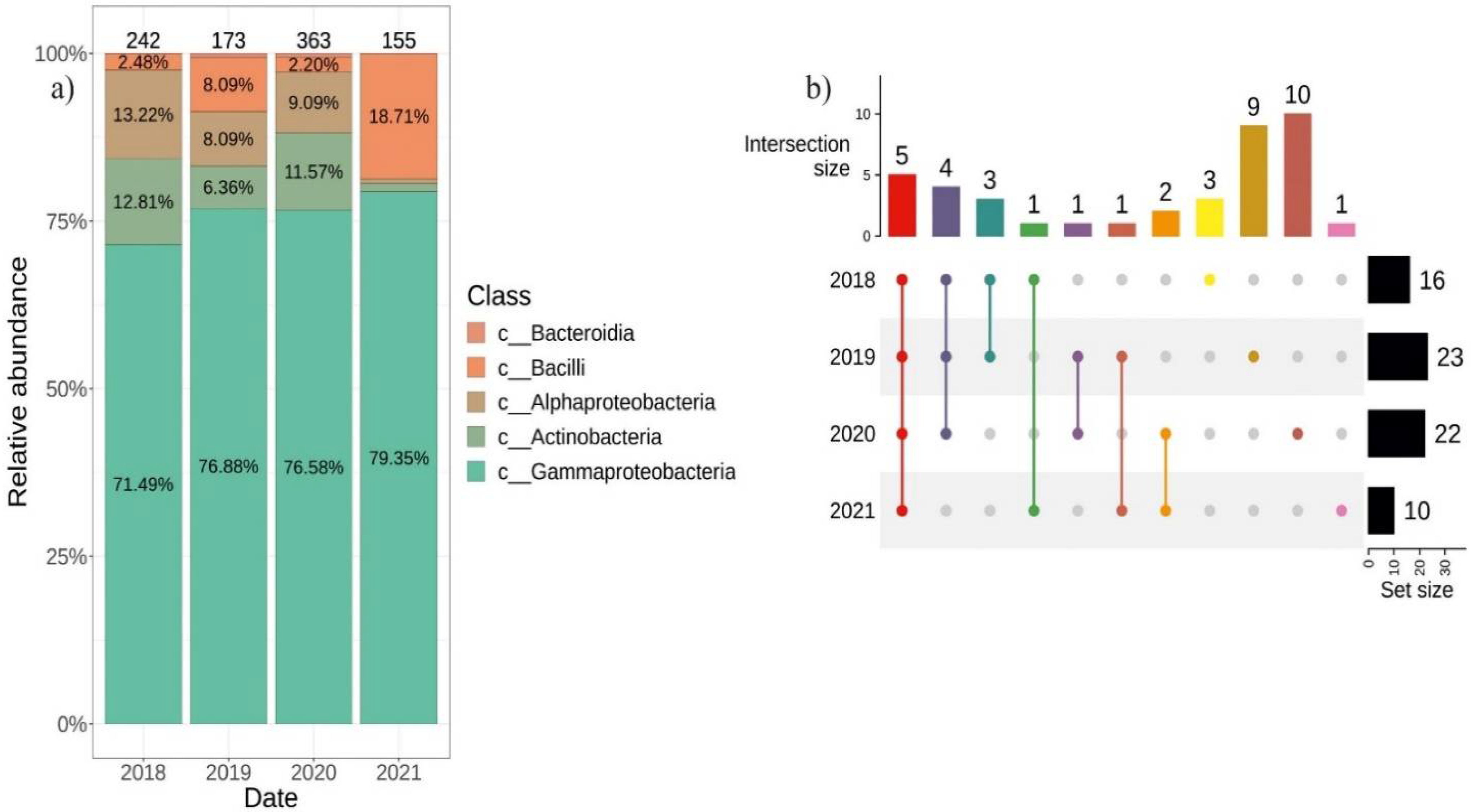
2.2. Differences in the Composition of Endophytic Bacterial Community in V. amurensis Depending on the Year of Tissue Collection
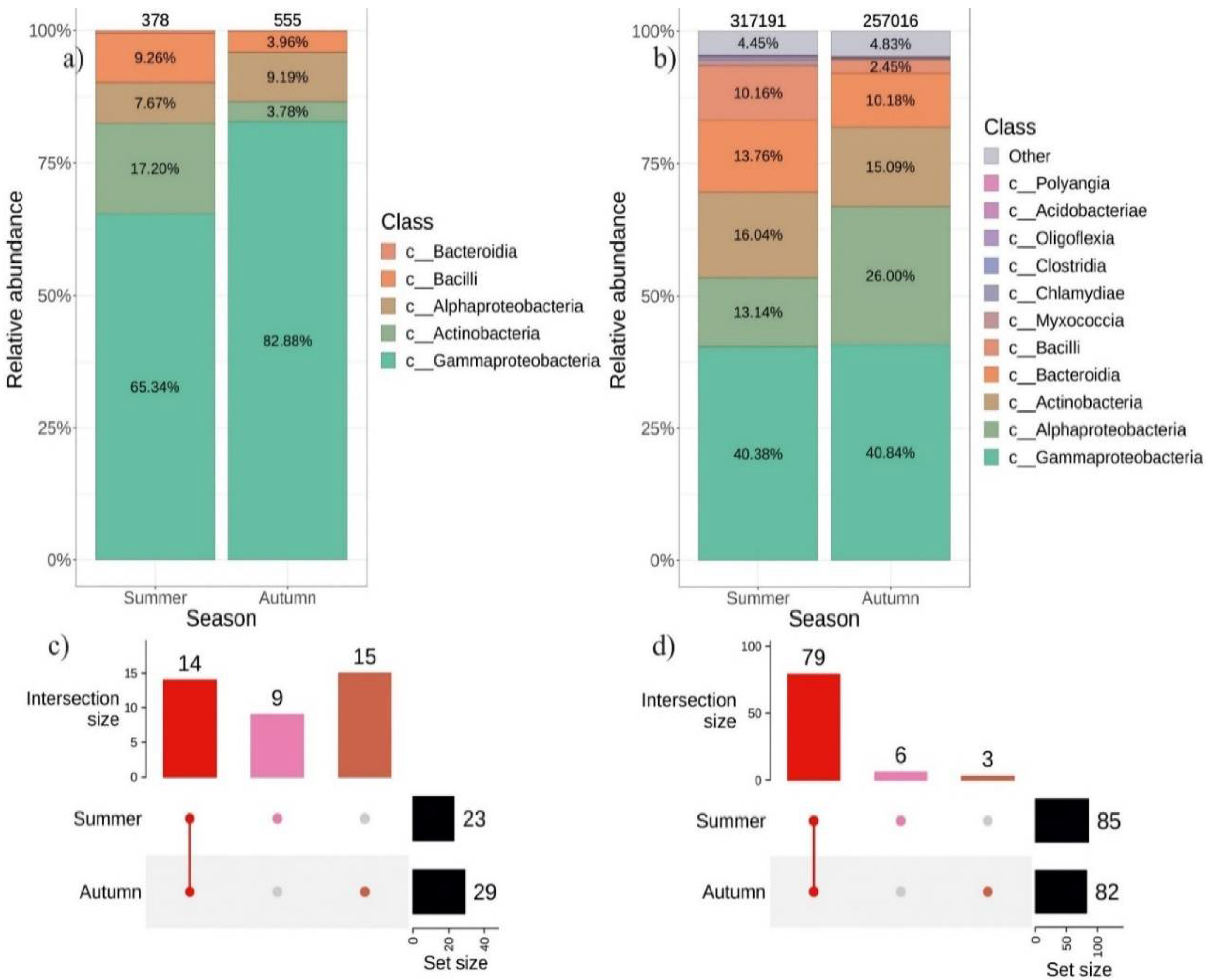
2.3. Seasonal Variations in the Composition of Endophytic Bacteriome in V. amurensis
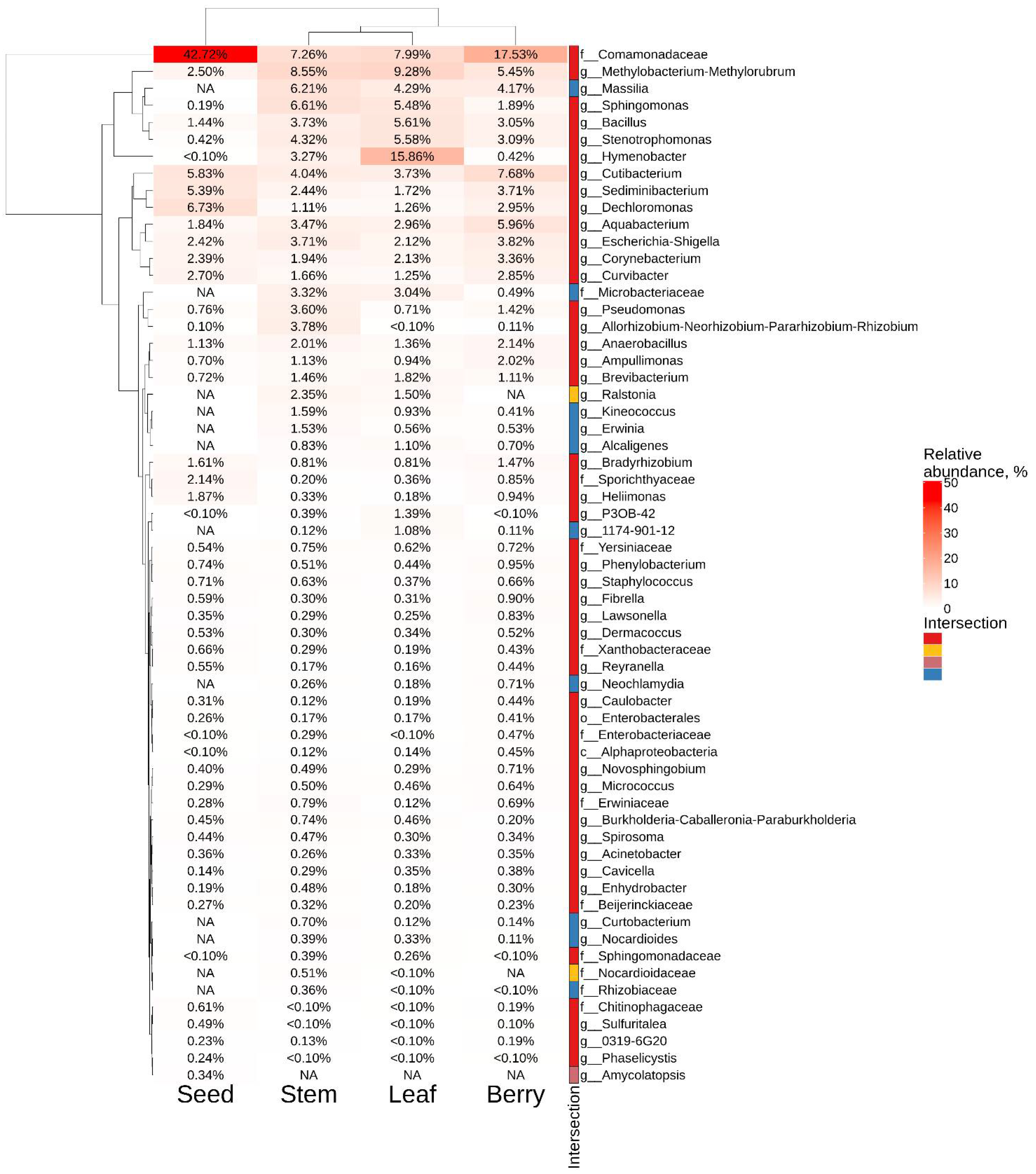
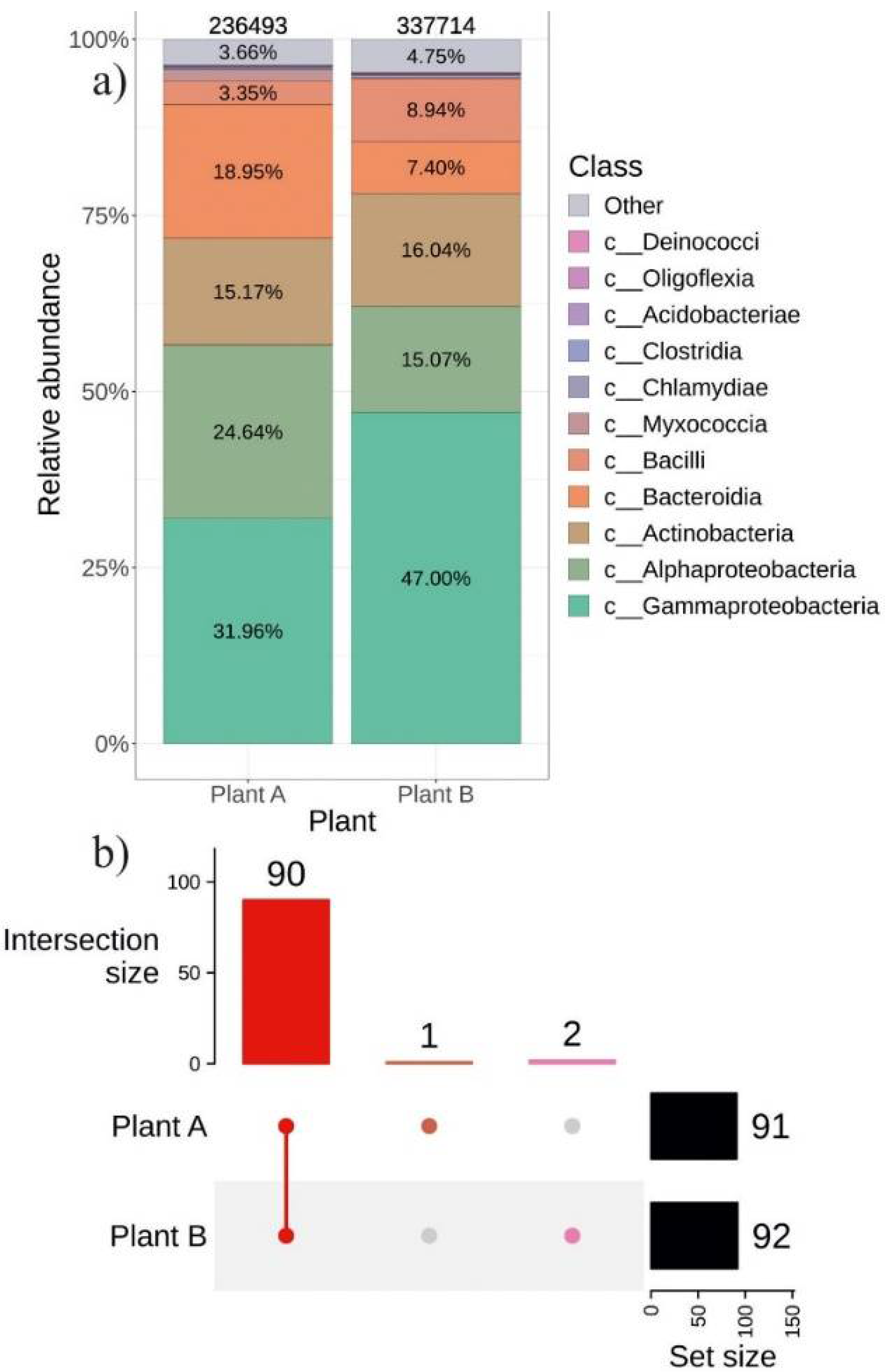
2.4. The Composition of Endophytic Bacterial Community in Different Representatives of V. amurensis Grapevine
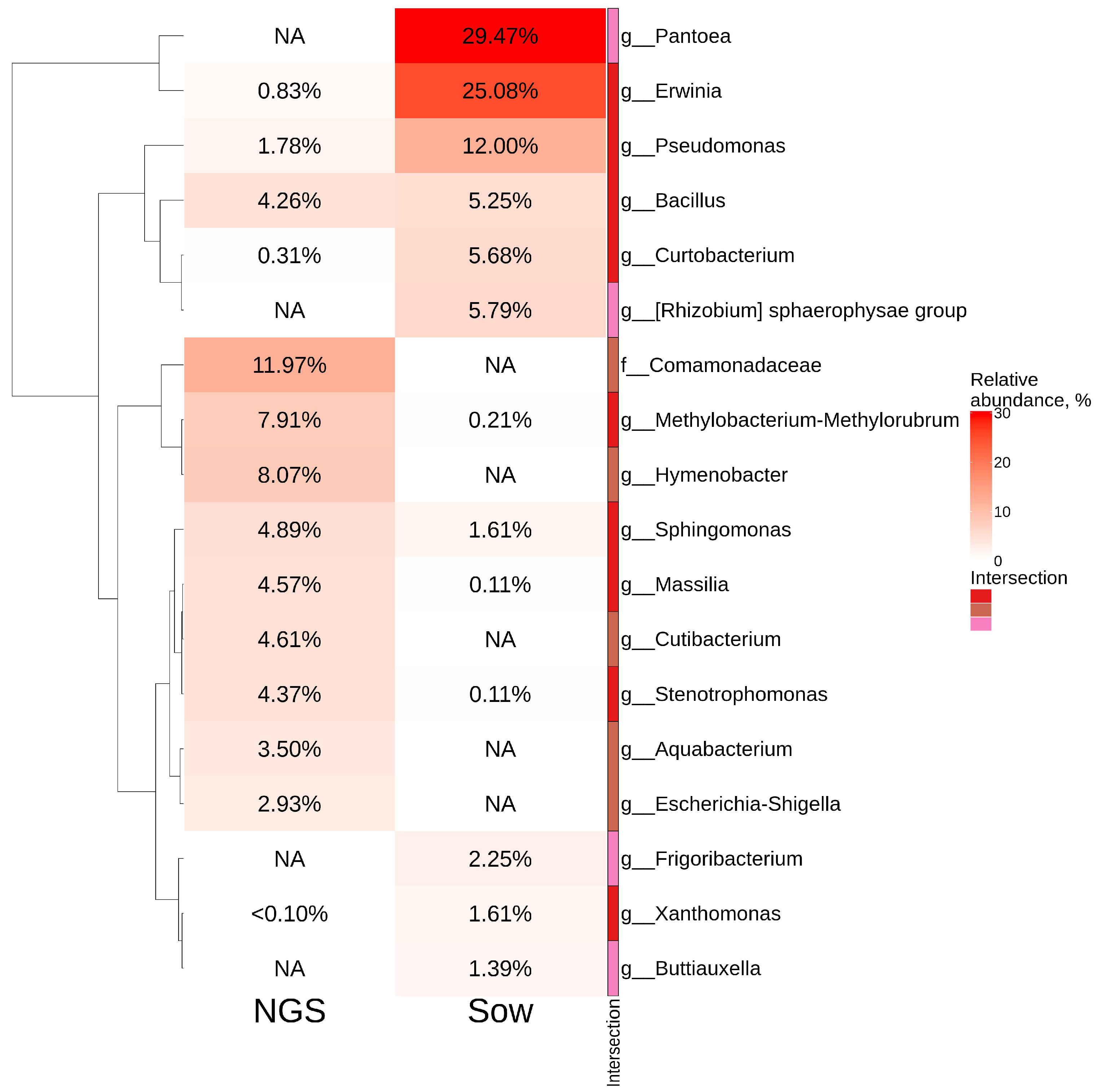
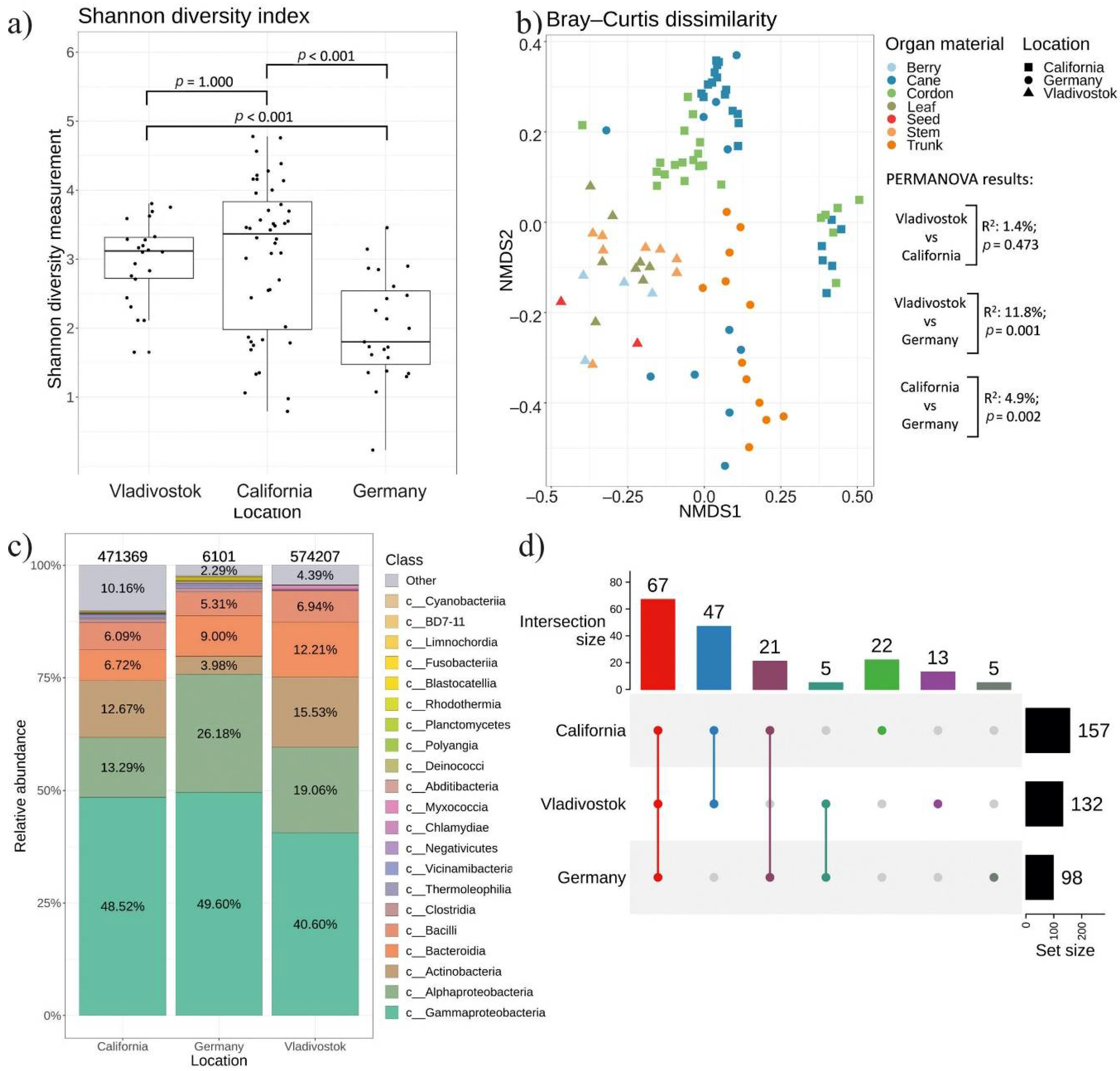
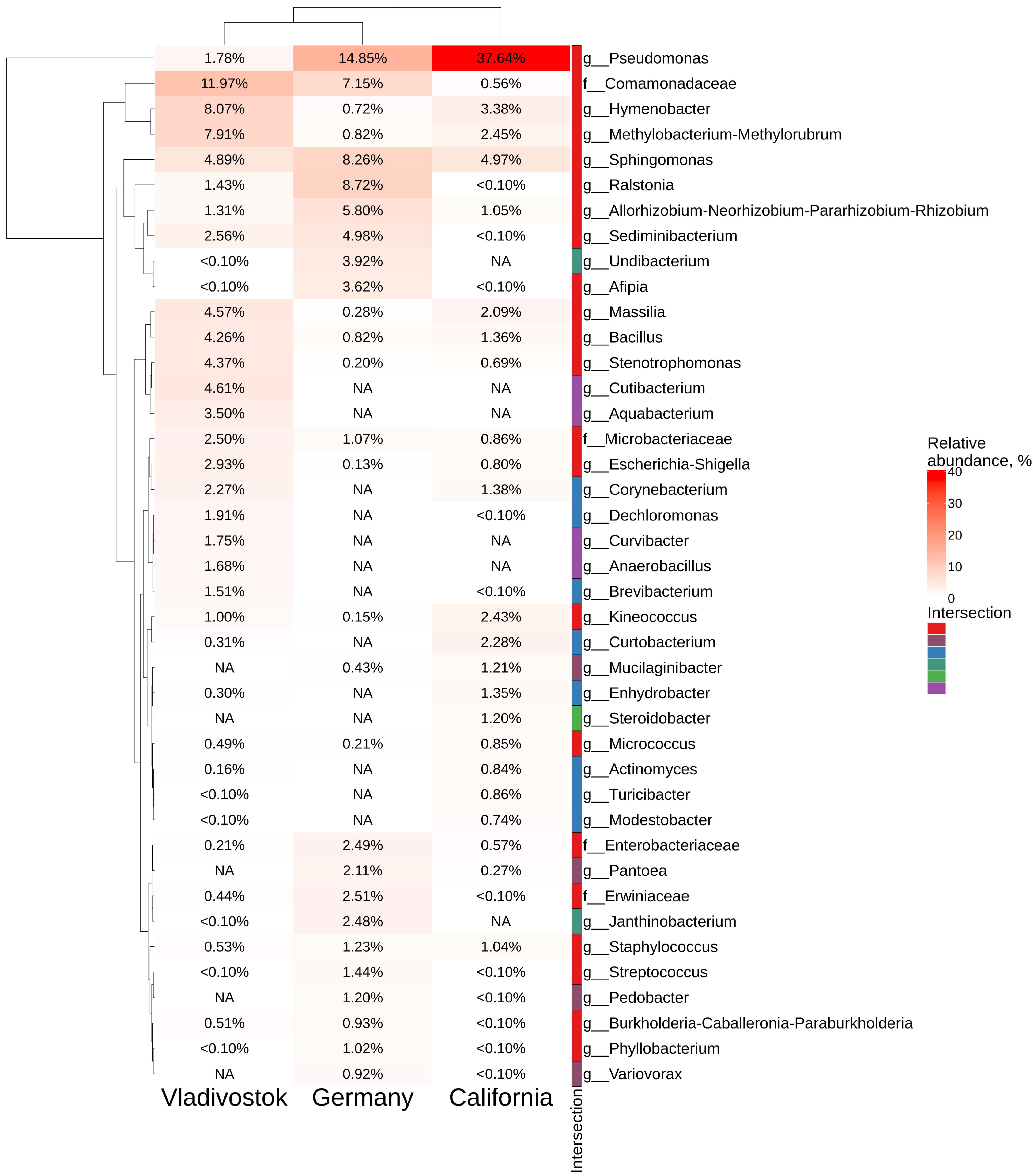
2.5. Comparative Analysis of Endophytic Bacteria Biodiversity in V. amurensis and V. vinifera
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Isolation and Identification of the Endophytic Bacteria
4.3. DNA Extraction, PCR Condition, Library Preparation, and Sequencing
4.4. Computational Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wen, J.; Nie, Z.-L.; Soejima, A.; Meng, Y. Phylogeny of Vitaceae based on the nuclear GAI1 gene sequences. Can. J. Bot. 2007, 85, 731–745. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-Q.; Ickert-Bond, S.M.; Nie, Z.-L.; Zhou, Z.; Chen, L.-Q.; Wen, J. Phylogeny of the Ampelocissus–Vitis clade in Vitaceae supports the New World origin of the grape genus. Mol. Phylogenetics Evol. 2016, 95, 217–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stranska, M.; Lovecka, P.; Vrchotova, B.; Uttl, L.; Bechynska, K.; Behner, A.; Hajslova, J. Bacterial Endophytes from Vitis vinifera L.—Metabolomics Characterization of Plant-Endophyte Crosstalk. Chem. Biodivers. 2021, 18, e2100516. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [Green Version]
- Kandel, S.L.; Joubert, P.M.; Doty, S.L. Bacterial Endophyte Colonization and Distribution within Plants. Microorganisms 2017, 5, 77. [Google Scholar] [CrossRef] [Green Version]
- West, E.R.; Cother, E.J.; Steel, C.C.; Ash, G.J. Thse characterization and diversity of bacterial endophytes of grapevine. Can. J. Microbiol. 2010, 56, 209–216. [Google Scholar] [CrossRef]
- Compant, S.; Mitter, B.; Colli-Mull, J.G.; Gangl, H.; Sessitsch, A. Endophytes of Grapevine Flowers, Berries, and Seeds: Identification of Cultivable Bacteria, Comparison with Other Plant Parts, and Visualization of Niches of Colonization. Microb. Ecol. 2011, 62, 188–197. [Google Scholar] [CrossRef]
- Salvetti, E.; Campanaro, S.; Campedelli, I.; Fracchetti, F.; Gobbi, A.; Tornielli, G.B.; Torriani, S.; Felis, G.E. Whole-Metagenome-Sequencing-Based Community Profiles of Vitis vinifera L. cv. Corvina Berries Withered in Two Post-harvest Conditions. Front. Microbiol. 2016, 7, 973. [Google Scholar] [CrossRef] [Green Version]
- Andreolli, M.; Lampis, S.; Zapparoli, G.; Angelini, E.; Vallini, G. Diversity of bacterial endophytes in 3 and 15 year-old grapevines of Vitis vinifera cv. Corvina and their potential for plant growth promotion and phytopathogen control. Microbiol. Res. 2016, 183, 42–52. [Google Scholar] [CrossRef]
- Gamalero, E.; Bona, E.; Novello, G.; Boatti, L.; Mignone, F.; Massa, N.; Cesaro, P.; Berta, G.; Lingua, G. Discovering the bacteriome of Vitis vinifera cv. Pinot Noir in a conventionally managed vineyard. Sci Rep 2020, 10, 6453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldan, E.; Nigris, S.; Populin, F.; Zottini, M.; Squartini, A.; Baldan, B. Identification of culturable bacterial endophyte community isolated from tissues of Vitisvinifera “Glera”. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2014, 148, 508–516. [Google Scholar] [CrossRef]
- Deyett, E.; Rolshausen, P.E. Endophytic microbial assemblage in grapevine. FEMS Microbiol. Ecol. 2020, 96, fiaa053. [Google Scholar] [CrossRef] [PubMed]
- Deyett, E.; Rolshausen, P.E. Temporal Dynamics of the Sap Microbiome of Grapevine Under High Pierce’s Disease Pressure. Front. Plant Sci. 2019, 10, 1246. [Google Scholar] [CrossRef] [PubMed]
- Campisano, A.; Pancher, M.; Puopolo, G.; Puddu, A.; Lòpez-Fernàndez, S.; Biagini, B.; Yousaf, S.; Pertot, I. Diversity in Endophyte Populations Reveals Functional and Taxonomic Diversity between Wild and Domesticated Grapevines. Am. J. Enol. Vitic. 2015, 66, 12–21. [Google Scholar] [CrossRef]
- Chen, Q.; Diao, L.; Song, H.; Zhu, X. Vitis amurensis Rupr: A review of chemistry and pharmacology. Phytomedicine 2018, 49, 111–122. [Google Scholar] [CrossRef]
- Liu, L.; Li, H. Review: Research progress in amur grape, Vitis amurensis Rupr. Can. J. Plant Sci. 2013, 93, 565–575. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Zhang, Y.; Zhang, H.; Huang, H.; Folta, K.M.; Lu, J. Whole genome wide expression profiles of Vitis amurensisgrape responding to downy mildew by using Solexa sequencing technology. BMC Plant Biol. 2010, 10, 234. [Google Scholar] [CrossRef] [Green Version]
- Dubrovina, A.S.; Kiselev, K.V. Regulation of stilbene biosynthesis in plants. Planta 2017, 246, 597–623. [Google Scholar] [CrossRef]
- Jenifer Selvarani, A.; Nishanthini, P.; Raji, P.; Sree Samanvitha, K.; Paulraj, P.; Iyappan, P.; Chandramohan, M.; Samrot, A.V. Antioxidant and Quorum Quenching Activity against Pseudomonas aeruginosa SU-18 of some Edible Fruit Juices. J. Pure Appl. Microbiol. 2019, 13, 1863–1876. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.A.; Mazumdar, R.M.; Islam, S.; Alam, M.J.; Urmee, S.A. Isolation, Identification and In Vitro Antibiotic Sensitivity Pattern of Citrus Canker Causing Organism Xanthomonas axonopodis. Adv. Life Sci. 2014, 1, 215–222. [Google Scholar]
- Yim, N.; Ha, D.T.; Trung, T.N.; Kim, J.P.; Lee, S.; Na, M.; Jung, H.; Kim, H.S.; Kim, Y.H.; Bae, K. The antimicrobial activity of compounds from the leaf and stem of Vitis amurensis against two oral pathogens. Bioorganic Med. Chem. Lett. 2010, 20, 1165–1168. [Google Scholar] [CrossRef] [PubMed]
- Mousa, W.K.; Shearer, C.R.; Limay-Rios, V.; Zhou, T.; Raizada, M.N. Bacterial endophytes from wild maize suppress Fusarium graminearum in modern maize and inhibit mycotoxin accumulation. Front. Plant Sci. 2015, 6, 805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shehata, H.R.; Lyons, E.M.; Jordan, K.S.; Raizada, M.N. Bacterial endophytes from wild and ancient maize are able to suppress the fungal pathogen Sclerotinia homoeocarpa. J. Appl. Microbiol. 2016, 120, 756–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Liu, T.; Zhang, X.; Xie, J.; Wang, Y.; Yan, R.; Jiang, Y.; Zhu, D. Cultivable Endophytic Bacteria in Seeds of Dongxiang Wild Rice and Their Role in Plant-Growth Promotion. Diversity 2021, 13, 665. [Google Scholar] [CrossRef]
- Liu, L.-H.; Yuan, T.; Zhang, J.-Y.; Tang, G.-X.; Lü, H.; Zhao, H.-M.; Li, H.; Li, Y.-W.; Mo, C.-H.; Tan, Z.-Y.; et al. Diversity of endophytic bacteria in wild rice (Oryza meridionalis) and potential for promoting plant growth and degrading phthalates. Sci. Total Environ. 2022, 806, 150310. [Google Scholar] [CrossRef]
- Faist, H.; Keller, A.; Hentschel, U.; Deeken, R. Grapevine (Vitis vinifera) Crown Galls Host Distinct Microbiota. Appl. Environ. Microbiol. 2016, 82, 5542–5552. [Google Scholar] [CrossRef] [Green Version]
- Campisano, A.; Antonielli, L.; Pancher, M.; Yousaf, S.; Pindo, M.; Pertot, I. Bacterial Endophytic Communities in the Grapevine Depend on Pest Management. PLoS ONE 2014, 9, e112763. [Google Scholar] [CrossRef]
- Greule, A.; Charkoudian, L.K.; Cryle, M.J. Chapter Six—Studying trans-acting enzymes that target carrier protein-bound amino acids during nonribosomal peptide synthesis. In Methods in Enzymology; Schmidt-Dannert, C., Quin, M.B., Eds.; Metabolons and Supramolecular Enzyme Assemblies; Academic Press: Cambridge, MA, USA, 2019; Volume 617, pp. 113–154. [Google Scholar]
- Holmström, C.; Kjelleberg, S. Marine Pseudoalteromonas species are associated with higher organisms and produce biologically active extracellular agents. FEMS Microbiol. Ecol. 1999, 30, 285–293. [Google Scholar] [CrossRef]
- Bell, C.R.; Dickie, G.A.; Harvey, W.L.G.; Chan, J.W.Y.F. Endophytic bacteria in grapevine. Can. J. Microbiol. 1995, 41, 46–53. [Google Scholar] [CrossRef]
- Salomon, M.V.; Bottini, R.; de Souza Filho, G.A.; Cohen, A.C.; Moreno, D.; Gil, M.; Piccoli, P. Bacteria isolated from roots and rhizosphere of Vitis vinifera retard water losses, induce abscisic acid accumulation and synthesis of defense-related terpenes in in vitro cultured grapevine. Physiol. Plant. 2014, 151, 359–374. [Google Scholar] [CrossRef]
- Cohen, A.C.; Dichiara, E.; Jofré, V.; Antoniolli, A.; Bottini, R.; Piccoli, P. Carotenoid profile produced by Bacillus licheniformis Rt4M10 isolated from grapevines grown in high altitude and their antioxidant activity. Int. J. Food Sci. Technol. 2018, 53, 2697–2705. [Google Scholar] [CrossRef]
- Torres, N.; Goicoechea, N.; Zamarreño, A.M.; Carmen Antolín, M. Mycorrhizal symbiosis affects ABA metabolism during berry ripening in Vitis vinifera L. cv. Tempranillo grown under climate change scenarios. Plant Sci. 2018, 274, 383–393. [Google Scholar] [CrossRef]
- Verhagen, B.; Trotel-Aziz, P.; Jeandet, P.; Baillieul, F.; Aziz, A. Improved Resistance Against Botrytis cinerea by Grapevine-Associated Bacteria that Induce a Prime Oxidative Burst and Phytoalexin Production. Phytopathology 2011, 101, 768–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aleynova, O.A.; Nityagovsky, N.N.; Suprun, A.R.; Kiselev, K.V. Enhancement of Stilbene Biosynthesis in Grape-Cell Cultures by Natural Products Based on Endophytes of the Wild Grape Species Vitis amurensis RUPR. Appl. Biochem. Microbiol. 2022, 58, 45–56. [Google Scholar] [CrossRef]
- Aleynova, O.A.; Suprun, A.R.; Nityagovsky, N.N.; Dubrovina, A.S.; Kiselev, K.V. The Influence of the Grapevine Bacterial and Fungal Endophytes on Biomass Accumulation and Stilbene Production by the In Vitro Cultivated Cells of Vitis amurensis Rupr. Plants 2021, 10, 1276. [Google Scholar] [CrossRef] [PubMed]
- Reasoner, D.J.; Geldreich, E.E. A new medium for the enumeration and subculture of bacteria from potable water. Appl. Environ. Microbiol. 1985, 49, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kiselev, K.V.; Dubrovina, A.S.; Tyunin, A.P. The methylation status of plant genomic DNA influences PCR efficiency. J. Plant Physiol. 2015, 175, 59–67. [Google Scholar] [CrossRef]
- Lane, D. 1 16S/23S rRNA sequencing. Nucleic Acid Tech. Bact. Syst. 1991, 115–175. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package, R Package Version 2.5-7. 2020. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 28 November 2020).
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Summer | Average t, °C | Precipitation, mm | Autumn | Average t, °C | Precipitation, mm |
|---|---|---|---|---|---|
| The norm | 18.1 | 159 | The norm | 16 | 103 |
| July 2018 | 18.5 | 124 | September 2018 | 16.4 | 163 |
| July 2019 | 17.1 | 131 | September 2019 | 17.3 | 44 |
| July 2020 | 14.7 | 281 | September 2020 | 16.3 | 138 |
| July 2021 | 21.3 | 24 | September 2021 | 17.7 | 120 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aleynova, O.A.; Nityagovsky, N.N.; Dubrovina, A.S.; Kiselev, K.V. The Biodiversity of Grapevine Bacterial Endophytes of Vitis amurensis Rupr. Plants 2022, 11, 1128. https://doi.org/10.3390/plants11091128
Aleynova OA, Nityagovsky NN, Dubrovina AS, Kiselev KV. The Biodiversity of Grapevine Bacterial Endophytes of Vitis amurensis Rupr. Plants. 2022; 11(9):1128. https://doi.org/10.3390/plants11091128
Chicago/Turabian StyleAleynova, Olga A., Nikolay N. Nityagovsky, Alexandra S. Dubrovina, and Konstantin V. Kiselev. 2022. "The Biodiversity of Grapevine Bacterial Endophytes of Vitis amurensis Rupr." Plants 11, no. 9: 1128. https://doi.org/10.3390/plants11091128