
Changes in Diversity and Community Composition of Root Endophytic Fungi Associated with Aristolochia chilensis along an Aridity Gradient in the Atacama Desert

 and
and
Abstract
:1. Introduction
2. Results
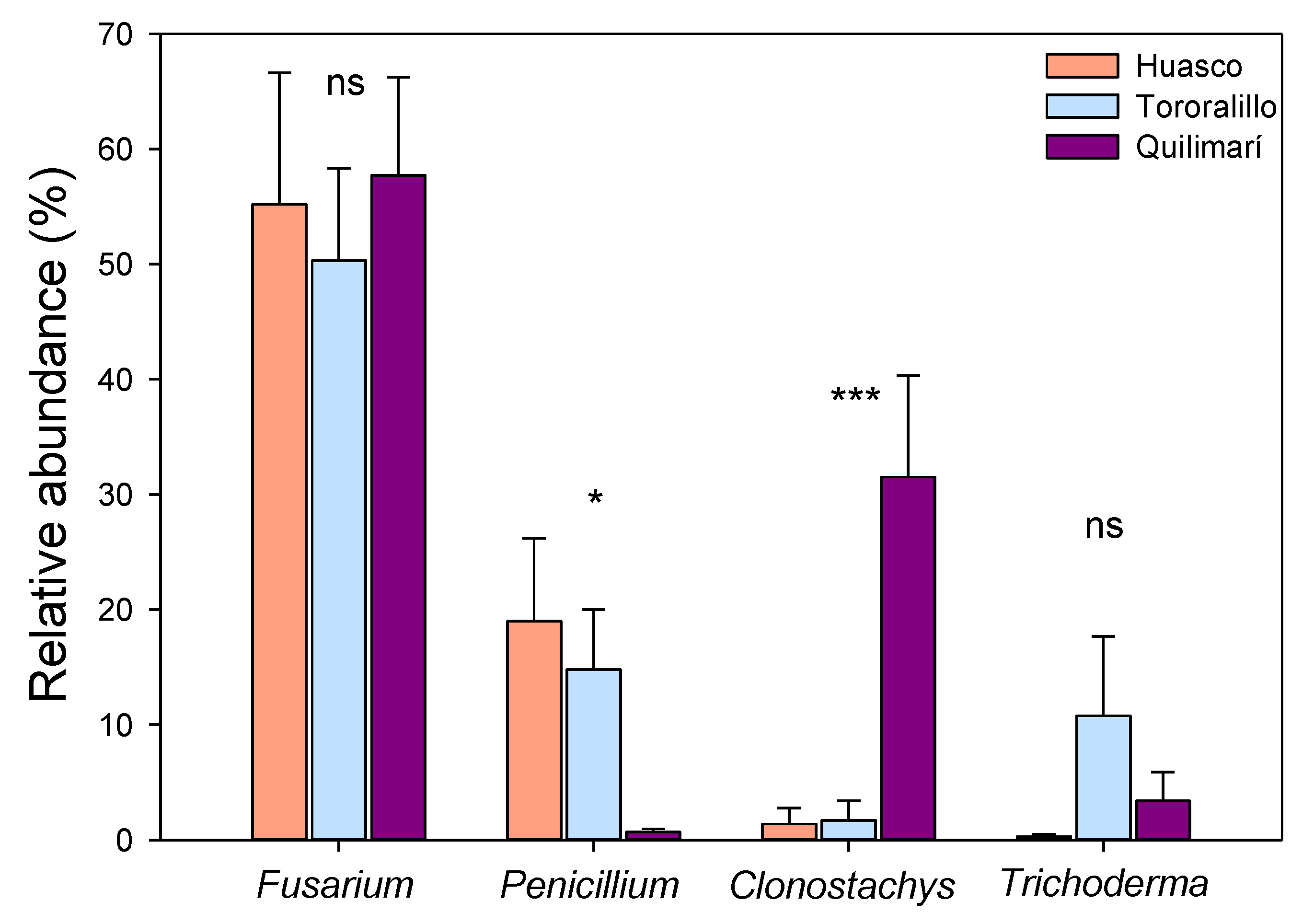
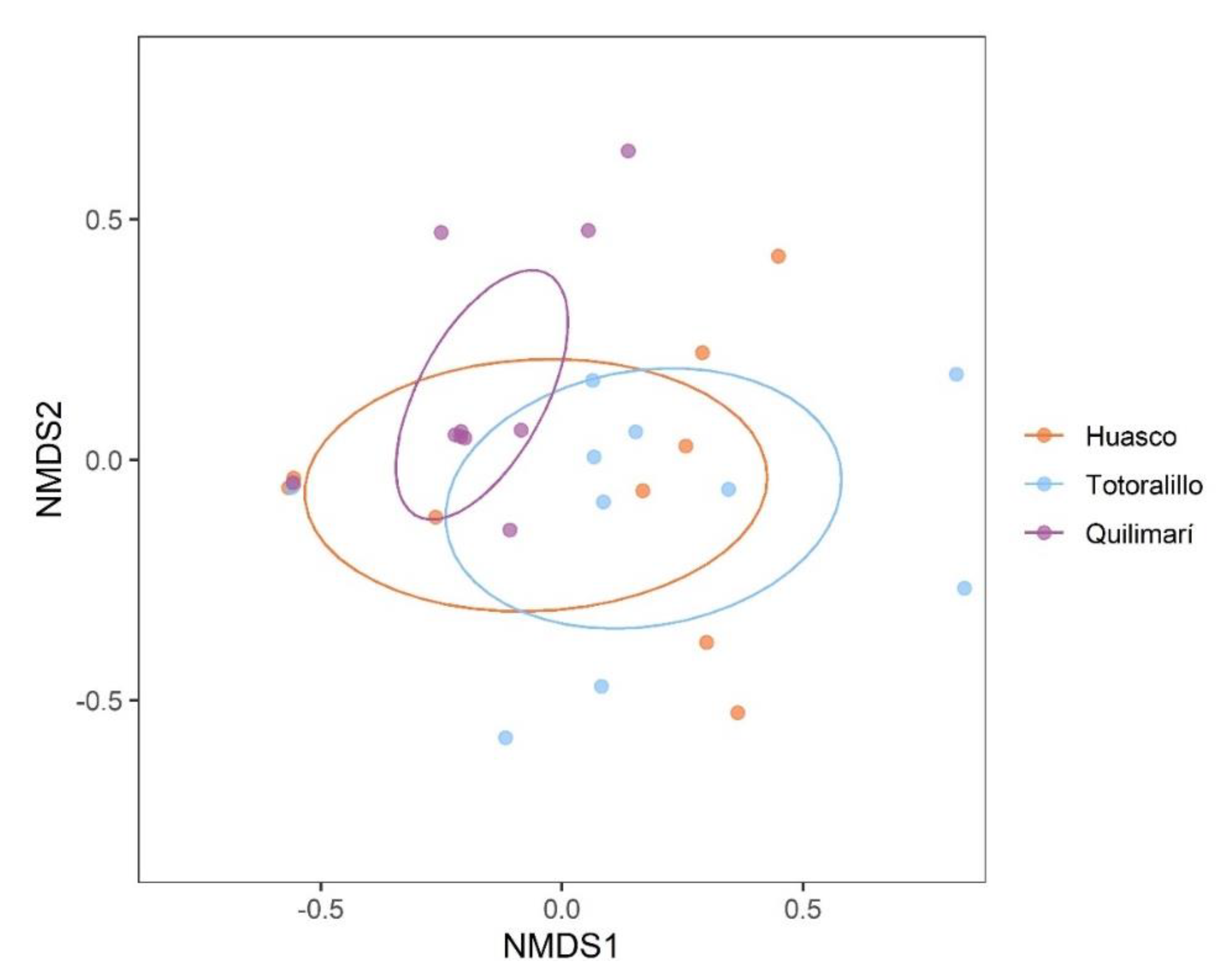
2.1. Diversity and Community Composition of Root Endophytic Fungi
2.2. Edaphic Factors and Location Differences
2.3. Field Experiment
3. Discussion
4. Materials and Methods
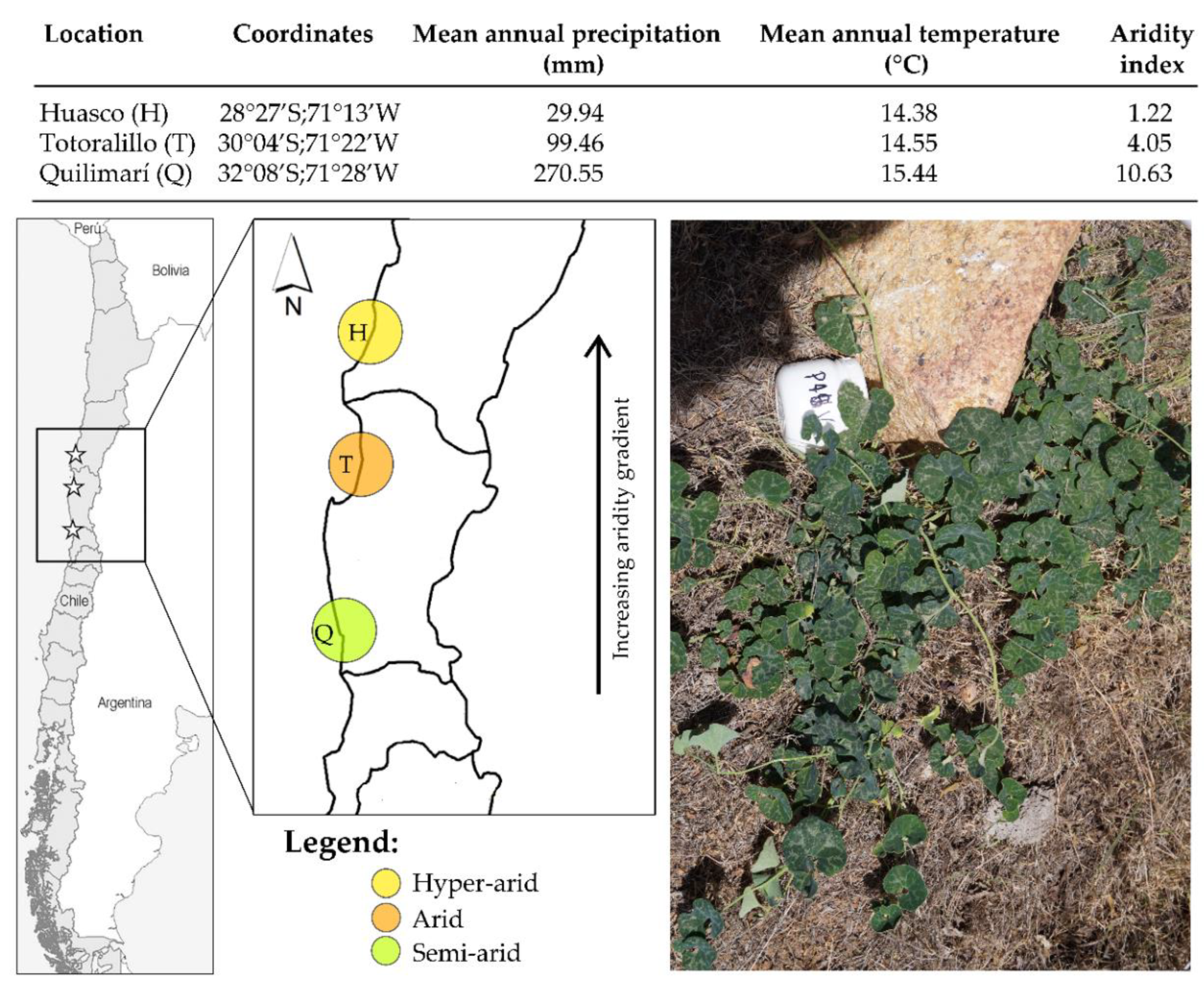
4.1. Study Species and Study Sites
4.2. Isolation of FE
4.3. DNA Extraction and Molecular Characterization of FE
4.4. Edaphic Factors
4.5. Field Experiment
4.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodriguez, R.J.; White, J.F., Jr.; Arnold, A.; Redman, R.S. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Lodge, D.J.; Fisher, P.J.; Sutton, B.C. Endophytic Fungi of Manilkara bidentata Leaves in Puerto Rico. Mycologia 1996, 88, 733–738. [Google Scholar] [CrossRef]
- Arnold, A.; Maynard, Z.; Gilbert, G.; Coley, P.; Kursar, T. Are tropical fungal endophytes hyperdiverse? Ecol. Lett. 2000, 3, 267–274. [Google Scholar] [CrossRef]
- Higgins, K.L.; Arnold, A.E.; Miadlikowska, J.; Sarvate, S.D.; Lutzoni, F. Phylogenetic relationships, host affinity, and geographic structure of boreal and arctic endophytes from three major plant lineages. Mol. Phylogenet. Evol. 2007, 42, 543–555. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, M.T.; Arnold, A.E. Diverse bacteria inhabit living hyphae of phylogenetically diverse fungal endophytes. App. Environ. Microbiol. 2010, 76, 4063–4075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmerman, N.B.; Vitousek, P.M. Fungal endophyte communities reflect environmental structuring across a Hawaiian landscape. Proc. Natl. Acad. Sci. USA 2012, 109, 13022–13027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, J.B.; Weiblen, G.D.; May, G. Host associations and beta diversity of fungal endophyte communities in New Guinea rainforest trees. Mol. Ecol. 2016, 25, 825–841. [Google Scholar] [CrossRef]
- González-Teuber, M.; Vilo, C.; Guevara-Araya, M.J.; Salgado-Luarte, C.; Gianoli, E. Leaf resistance traits influence endophytic fungi colonization and community composition in a South American temperate rainforest. J. Ecol. 2020, 108, 1019–1029. [Google Scholar] [CrossRef]
- Arnold, A.E.; Lutzoni, F. Diversity and host range of foliar fungal endophytes: Are tropical leaves biodiversity hotspots? Ecology 2017, 88, 541–549. [Google Scholar] [CrossRef]
- U’Ren, J.M.; Lutzoni, F.; Miadlikowska, J.; Laetsch, A.D.; Arnold, A. Host and geographic structure of endophytic and endolichenic fungi at a continental scale. Am. J. Bot. 2012, 99, 898–914. [Google Scholar] [CrossRef] [Green Version]
- Faeth, S.H.; Hammon, K.E. Fungal endophytes in oak trees: Long-term patterns of abundance and associations with leafminers. Ecology 1997, 78, 810–819. [Google Scholar] [CrossRef]
- Suwannarach, N.; Bussaban, B.; Nuangmek, W.; McKenzie, E.H.; Hyde, K.D.; Lumyong, S. Diversity of endophytic fungi associated with Cinnamomum bejolghota (Lauraceae) in Northern Thailand. Chiang Mai J. Sci. 2012, 39, 389–398. [Google Scholar]
- Pieterse, Z.; Aveling, T.A.; Jacobs, A.; Cowan, D.A. Seasonal variability in fungal endophytes from Aizoaceae plants in the Succulent Karoo biodiversity hotspot, South Africa. J. Arid Environ. 2018, 156, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Suryanarayanan, T.S.; Kumaresan, V.; Johnson, J.A. Foliar fungal endophytes from two species of the mangrove Rhizophora. Can. J. Microbiol. 1998, 44, 1003–1006. [Google Scholar] [CrossRef]
- Suryanarayanan, T.S.; Murali, T.S.; Venkatesan, G. Occurrence and distribution of fungal endophytes in tropical forests across a rainfall gradient. Can. J. Bot. 2002, 80, 818–826. [Google Scholar] [CrossRef]
- Oita, S.; Ibáñez, A.; Lutzoni, F.; Miadlikowska, J.; Geml, J.; Lewis, L.A.; Hom, L.A.; Carbone, I.; U’Ren, J.M.; Arnold, A.E. Climate and seasonality drive the richness and composition of tropical fungal endophytes at a landscape scale. Commun. Biol. 2021, 4, 313. [Google Scholar] [CrossRef]
- Giauque, H.; Hawkes, C.V. Climate affects symbiotic fungal endophyte diversity and performance. Am. J. Bot. 2013, 100, 1435–1444. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Raghavan, S.; Garcia, M.; Sinsabaugh, R.L.; Natvig, D.O.; Lowrey, T.K. Endophytic fungal symbionts associated with gypsophilous plants. Botany 2014, 92, 295–301. [Google Scholar] [CrossRef]
- Massimo, N.C.; Devan, M.N.; Arendt, K.R.; Wilch, M.H.; Riddle, J.M.; Furr, S.H.; Steen, C.; U’Ren, J.M.; Sandberg, D.C.; Arnold, A.E. Fungal endophytes in aboveground tissues of desert plants: Infrequent in culture, but highly diverse and distinctive symbionts. Microb. Ecol. 2015, 70, 61–76. [Google Scholar] [CrossRef]
- Noy-Meir, I. Desert ecosystems: Environment and producers. Annu. Rev. Ecol. Evol. Syst. 1973, 4, 25–51. [Google Scholar] [CrossRef]
- Maestre, F.T.; Delgado-Baquerizo, M.; Jeffries, T.C.; Eldridge, D.J.; Ochoa, V.; Gozalo, B.; Querod, J.; García-Gómez, M.; Gallardof, A.; Ulrichg, W.; et al. Increasing aridity reduces soil microbial diversity and abundance in global drylands. Proc. Natl. Acad. Sci. USA 2015, 112, 15684–15689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, S.; Liu, Z.; Lin, Y.; Yang, J.; Chen, W.; Wei, G. Bacterial communities in oil contaminated soils: Biogeography and co-occurrence patterns. Soil Biol. Biochem. 2016, 98, 64–73. [Google Scholar] [CrossRef] [Green Version]
- Kivlin, S.N.; Winston, G.C.; Goulden, M.L.; Treseder, K.K. Environmental filtering affects soil fungal community composition more than dispersal limitation at regional scales. Fungal Ecol. 2014, 12, 14–25. [Google Scholar] [CrossRef] [Green Version]
- Lau, M.K.; Arnold, A.E.; Johnson, N.C. Factors influencing communities of foliar fungal endophytes in riparian woody plants. Fungal Ecol. 2013, 6, 365–378. [Google Scholar] [CrossRef]
- Gargouri, M.; Karray, F.; Chebaane, A.; Mhiri, N.; Partida-Martínez, L.P.; Sayadi, S.; Mliki, A. Increasing aridity shapes beta diversity and the network dynamics of the belowground fungal microbiome associated with Opuntia ficus-indica. Sci. Total Environ. 2021, 773, 145008. [Google Scholar] [CrossRef]
- Información Oficial Hidrometeorológica y de Calidad de Aguas en Línea. Available online: http://snia.dga.cl/BNAConsultas/reportes (accessed on 10 December 2015).
- De Martonne, E. Aridité et indices d´aridité. Académie des Sciences. Comptes Rendus 1923, 182, 1935–1938. [Google Scholar]
- Jackson, C.R.; Randolph, K.C.; Osborn, S.L.; Tyler, H.L. Culture dependent and independent analysis of bacterial communities associated with commercial salad leaf vegetables. BMC Microbiol. 2013, 13, 274. [Google Scholar] [CrossRef] [Green Version]
- Bodenhausen, N.; Horton, M.W.; Bergelson, J. Bacterial communities associated with the leaves and roots of Arabidopis thaliana. PLoS ONE 2013, 8, e56329. [Google Scholar] [CrossRef]
- Bui, A.; Orr, D.; Lepori-Bui, M.; Konicek, K.; Young, H.S.; Moeller, H.V. Soil fungal community composition and functional similarity shift across distinct climatic conditions. FEMS Microbiol. Ecol. 2020, 96, fiaa193. [Google Scholar] [CrossRef]
- Penner, S.; Sapir, Y. Foliar Endophytic Fungi Inhabiting an Annual Grass Along an Aridity Gradient. Curr. Microbiol. 2021, 78, 2080–2090. [Google Scholar] [CrossRef]
- Herrera, J.; Poudel, R.; Nebel, K.A.; Collins, S.L. Precipitation increases the abundance of some groups of root-associated fungal endophytes in a semiarid grassland. Ecosphere 2011, 2, 1–14. [Google Scholar] [CrossRef]
- Brockett, B.F.; Prescott, C.E.; Grayston, S.J. Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar] [CrossRef]
- Kaisermann, A.; Maron, P.A.; Beaumelle, L.; Lata, J.C. Fungal communities are more sensitive indicators to non-extreme soil moisture variations than bacterial communities. Appl. Soil Ecol. 2015, 86, 158–164. [Google Scholar] [CrossRef]
- Schwinning, S.; Sala, O.E. Hierarchy of responses to resource pulses in arid and semi-arid ecosystems. Oecologia 2004, 141, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [Green Version]
- Andrew, D.R.; Fitak, R.R.; Munguia-Vega, A.; Racolta, A.; Martinson, V.G.; Dontsova, K. Abiotic factors shape microbial diversity in Sonoran Desert soils. Appl. Environ. Microbiol. 2012, 78, 7527–7537. [Google Scholar] [CrossRef] [Green Version]
- Loro, M.; Valero-Jiménez, C.A.; Nozawa, S.; Márquez, L.M. Diversity and composition of fungal endophytes in semiarid Northwest Venezuela. J. Arid Environ. 2012, 85, 46–55. [Google Scholar] [CrossRef]
- González-Teuber, M.; Vilo, C.; Bascuñán-Godoy, L. Molecular characterization of endophytic fungi associated with the roots of Chenopodium quinoa inhabiting the Atacama Desert, Chile. Genom. Data 2017, 11, 109–112. [Google Scholar] [CrossRef]
- González-Teuber, M.; Urzúa, A.; Plaza, P.; Bascuñán-Godoy, L. Effects of root endophytic fungi on response of Chenopodium quinoa to drought stress. Plant Ecol. 2018, 219, 231–240. [Google Scholar] [CrossRef]
- González-Teuber, M.; Contreras, R.A.; Zuñiga, G.E.; Barrera, D.; Bascunan-Godoy, L. Synergistic association with root endophytic fungi improves physiological and biochemical responses of Chenopodium quinoa to salt stress. Front. Ecol. Evol. 2022, 9, 963. [Google Scholar] [CrossRef]
- Zhang, T.; Yao, Y.F. Endophytic fungal communities associated with vascular plants in the high arctic zone are highly diverse and host-plant specific. PLoS ONE 2015, 10, e0130051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Frenne, P.; Graae, B.J.; Rodríguez-Sánchez, F.; Kolb, A.; Chabrerie, O.; Decocq, G.; De Kort, H.; De Schrijver, A.D.; Diekmann, M.; Eriksson, O.; et al. Latitudinal gradients as natural laboratories to infer species’ responses to temperature. J. Ecol. 2013, 101, 784–795. [Google Scholar] [CrossRef]
- Caddy-Retalic, S.; Andersen, A.N.; Aspinwall, M.J.; Breed, M.F.; Byrne, M.; Christmas, M.J.; Bradley, N.D.; Fordham, D.A.; Guerin, G.R.; Hoffmann, A.A.; et al. Bioclimatic transect networks: Powerful observatories of ecological change. Ecol. Evol. 2017, 7, 4607–4619. [Google Scholar] [CrossRef] [PubMed]
- Bull, A.T.; Andrews, B.A.; Dorador, C.; Goodfellow, M. Introducing the Atacama Desert. Antonie Leeuwenhoek 2018, 111, 1269–1272. [Google Scholar] [CrossRef] [Green Version]
- Valdivia, C.E.; Niemeyer, H.M. Noncorrelated evolution between herbivore and pollinator-linked features in Aristolochia chilensis (Aristolochiaceae). Biol. J. Linn. Soc. 2007, 91, 239–245. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, A. El Árbol Urbano en Chile, 3rd ed.; Fundación Claudio Gay: Santiago, Chile, 1998; p. 79. [Google Scholar]
- Verbist, K.; Robertson, A.W.; Cornelis, W.M.; Gabriels, D. Seasonal predictability of daily rainfall characteristics in central northern Chile for dry-land management. JAMC 2010, 49, 1938–1955. [Google Scholar] [CrossRef] [Green Version]
- Dirección Meteorológica de Chile. Available online: http://www.meteochile.cl/PortalDMC-web/index.xhtml (accessed on 20 December 2016).
- Rodriguez, R.; Marticorena, C.; Alarcón, D.; Baeza, C.; Cavieres, L.; Finot, V.L.; Fuentes, N.; Kiessling, A.; Mihoc, M.; Pauchard, A.; et al. Catálogo de las plantas vasculares de Chile. Gayana Bot. 2018, 75, 1–430. [Google Scholar] [CrossRef] [Green Version]
- Arnold, A.E.; Maynard, Z.; Gilbert, G.S. Fungal endophytes in dicotyledonous neotropical trees: Patterns of abundance and diversity. Mycol. Res. 2001, 105, 1502–1507. [Google Scholar] [CrossRef] [Green Version]
- Arnold, A.E.; Mejia, L.C.; Kyllo, D.; Rojas, E.I.; Maynard, Z.; Robbins, N.; Herre, E.A. Fungal endophytes limit pathogen damage in a tropical tree. Proc. Natl. Acad. Sci. USA 2003, 100, 15649–15654. [Google Scholar] [CrossRef] [Green Version]
- Gamboa, M.A.; Laureano, S.; Bayman, P. Measuring diversity of endophytic fungi in leaf fragments: Does size matter? Mycopathologia 2002, 156, 41–45. [Google Scholar] [CrossRef]
- Nicholson, T.P.; Rudd, B.A.; Dawson, M.; Lazarus, C.M.; Simpson, T.J.; Cox, R.J. Design and utility of oligonucleotide gene probes for fungal polyketide synthases. Chem. Biol. 2001, 8, 157–178. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- Sadzawka, A.; Carrasco, M.; Grez, R.; Mora, M.; Flores, H.; Neaman, A. Métodos de análisis recomendados para los suelos de Chile. In Serie Actas–Instituto de Investigaciones Agropecuarias; Instituto de Investigaciones Agropecuarias, Centro Regional de Investigación La Platina: Santiago, Chile, 2006; p. 164. [Google Scholar]
- Arnold, A.E.; Henk, D.A.; Eells, R.L.; Lutzoni, F.; Vilgalys, R. Diversity and phylogenetic affinities of foliar fungal endophytes in loblolly pine inferred by culturing and environmental PCR. Mycologia 2007, 99, 185–206. [Google Scholar] [CrossRef] [PubMed]
- McArdle, B.H.; Anderson, M.J. Fitting multivariate models to community data: A comment on distance-based redundancy analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Warton, D.I.; Wright, S.T.; Wang, Y. Distance-based multivariate analyses confound location and dispersion effects. Methods Ecol. Evol. 2012, 3, 89–101. [Google Scholar] [CrossRef]
- Paliy, O.; Shankar, V. Application of multivariate statistical techniques in microbial ecology. Mol. Ecol. 2016, 25, 1032–1057. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: http://www.R-project.org/ (accessed on 1 October 2016).




{kind=link}
{kind=link}
{kind=link}
| Location | Description | Identity (%) | No. of Isolates | Accession Number |
|---|---|---|---|---|
| Huasco | Aspergillus ustus | 99.8% | 2 | HQ607918.1 |
| Aspergillus versicolor | 100% | 1 | MT560203.1 | |
| Clonostachys sp. | 99.8% | 3 | MH864436.1 | |
| Fusarium oxysporum | 100% | 41 | MT560381.1 | |
| Fusarium sp. | 100% | 19 | MT447523.1 | |
| Hypocrea lixii | 99.5% | 1 | EF191308.1 | |
| Penicillium gladioli | 99.5% | 3 | DQ339568.1 | |
| Penicillium murcianum | 99.5% | 3 | KP016843.1 | |
| Penicillium polonicum | 100% | 6 | MT529240.1 | |
| Totoralillo | Cladosporium ramotenellum | 100% | 1 | MT441591.1 |
| Clonostachys rosea | 100% | 14 | MT441592.1 | |
| Diaporthe columnaris | 100% | 2 | MT441604.1 | |
| Epicoccum nigrum | 100% | 1 | MT441593.1 | |
| Fungal sp. | 100% | 11 | MT441613.1 | |
| Fusarium decemcellulare | 100% | 3 | MT441594.1 | |
| Fusarium oxysporum 1 | 100% | 3 | MT441590.1 | |
| Fusarium oxysporum 2 | 100% | 39 | MT530243.1 | |
| Fusarium sp. | 100% | 115 | MT441596.1 | |
| Fusarium sambucinum | 100% | 8 | MT441588.1 | |
| Hypocrea viridescens | 100% | 2 | MT441598.1 | |
| Meyerozyma caribbica | 100% | 1 | MT441599.1 | |
| Meyerozyma guilliermondii | 100% | 14 | MN592978.1 | |
| Penicillium crustosum | 100% | 1 | MT441603.1 | |
| Penicillium glabrum | 100% | 5 | MT441616.1 | |
| Penicillium murcianum | 100% | 2 | MT441602.1 | |
| Penicillium sp. | 100% | 1 | MT441612.1 | |
| Phialemoniopsis cornearis | 100% | 9 | MT441611.1 | |
| Phomopsis sp. | 100% | 17 | MT441605.1 | |
| Preussia australis | 100% | 1 | MT441614.1 | |
| Sarocladium spinificis | 100% | 1 | MT441606.1 | |
| Talaromyces amestolkiae | 100% | 9 | MT441607.1 | |
| Talaromyces minioluteus | 100% | 1 | MT441589.1 | |
| Talaromyces minioluteus | 100% | 15 | MT441589.1 | |
| Trichoderma atroviride | 100% | 1 | MT441609.1 | |
| Trichoderma breve | 97% | 1 | MT441608.1 | |
| Trichoderma lixii | 100% | 1 | MT441597.1 | |
| Trichoderma sp. | 100% | 1 | MT441610.1 | |
| Trichoderma virilente | 99.8% | 8 | NR_138447.1 | |
| Quilimarí | Aspergillus flavipes | 99.8 | 1 | EU645664.1 |
| Fusarium oxysporum 1 | 100 | 73 | MT560381.1 | |
| Fusarium oxysporum 2 | 99.4 | 117 | ON081646.1 | |
| Fusarium oxysporum 3 | 99.6 | 1 | MH454072.1 | |
| Fusarium sp. | 99.5 | 2 | HQ130713.1 | |
| Meyerozyma guilliermondii | 95.7 | 1 | MH545918.1 | |
| Meyerozyma guilliermondii | 97.8 | 2 | LC317638.1 | |
| Trichoderma sp. | 99.8 | 2 | KX459438.1 | |
| Uncultured Clonostachys | 96.5 | 2 | MK407059.1 | |
| Uncultured fungus clone | 99.7 | 2 | KF800414.1 |
| Indexes | H | T | Q |
|---|---|---|---|
| Chao | 32.62 ± 4.62 a | 54.43 ± 5.54 b | 76.38 ± 1.63 c |
| Bootstrap | 20.65 ± 3.02 a | 54.26 ± 10.45 b | 59.92 ± 1.16 c |
| FE frequency | 7.90 ± 1.79 a | 28.81 ± 6.03 b | 20.20 ± 4.64 b |
| Shannon | 0.88 ± 0.16 a | 1.47 ± 0.19 b | 0.91 ± 0.14 ab |
| Simpson | 0.47 ± 0.09 a | 0.58 ± 0.07 a | 0.45 ± 0.06 a |
| df | SS | MS | Pseudo-F | R2 | p | |
|---|---|---|---|---|---|---|
| Locations | 2 | 934.7 | 467.3 | 2.63 | 0.16 | 0.008 |
| Residuals | 27 | 4790.1 | 177.4 | 0.83 | ||
| Total | 29 | 5724.8 | 1 | |||
| H vs. T | 1 | 100.5 | 0.74 | 0.03 | 0.740 | |
| H vs. Q | 1 | 542.7 | 3.42 | 0.15 | 0.036 | |
| T vs. Q | 1 | 758.9 | 3.17 | 0.15 | 0.004 |
| Edaphic Factors | AIC | F | p |
|---|---|---|---|
| Soil water potential | 82.04 | 1.63 | 0.03 * |
| Soil moisture | 82.03 | 1.51 | 0.04 * |
| pH | 81.42 | 1.27 | 0.15 ns |
| N | 81.16 | 1.22 | 0.16 ns |
| P | 81.18 | 0.99 | 0.46 ns |
| K | 81.39 | 0.84 | 0.74 ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guevara-Araya, M.J.; Escobedo, V.M.; Palma-Onetto, V.; González-Teuber, M. Changes in Diversity and Community Composition of Root Endophytic Fungi Associated with Aristolochia chilensis along an Aridity Gradient in the Atacama Desert. Plants 2022, 11, 1511. https://doi.org/10.3390/plants11111511
Guevara-Araya MJ, Escobedo VM, Palma-Onetto V, González-Teuber M. Changes in Diversity and Community Composition of Root Endophytic Fungi Associated with Aristolochia chilensis along an Aridity Gradient in the Atacama Desert. Plants. 2022; 11(11):1511. https://doi.org/10.3390/plants11111511
Chicago/Turabian StyleGuevara-Araya, María José, Víctor M. Escobedo, Valeria Palma-Onetto, and Marcia González-Teuber. 2022. "Changes in Diversity and Community Composition of Root Endophytic Fungi Associated with Aristolochia chilensis along an Aridity Gradient in the Atacama Desert" Plants 11, no. 11: 1511. https://doi.org/10.3390/plants11111511