
Endophytic Fungal and Bacterial Microbiota Shift in Rice and Barnyardgrass Grown under Co-Culture Condition

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growth
2.2. Sample Collection
2.3. DNA Extraction and PCR Amplification
2.4. Sequence Analysis
2.5. Statistical Analysis
3. Results
3.1. Soil Physicochemical Analysis
3.2. Sequencing Results
3.3. Alpha Diversity and Community Composition in Soil Microbiota
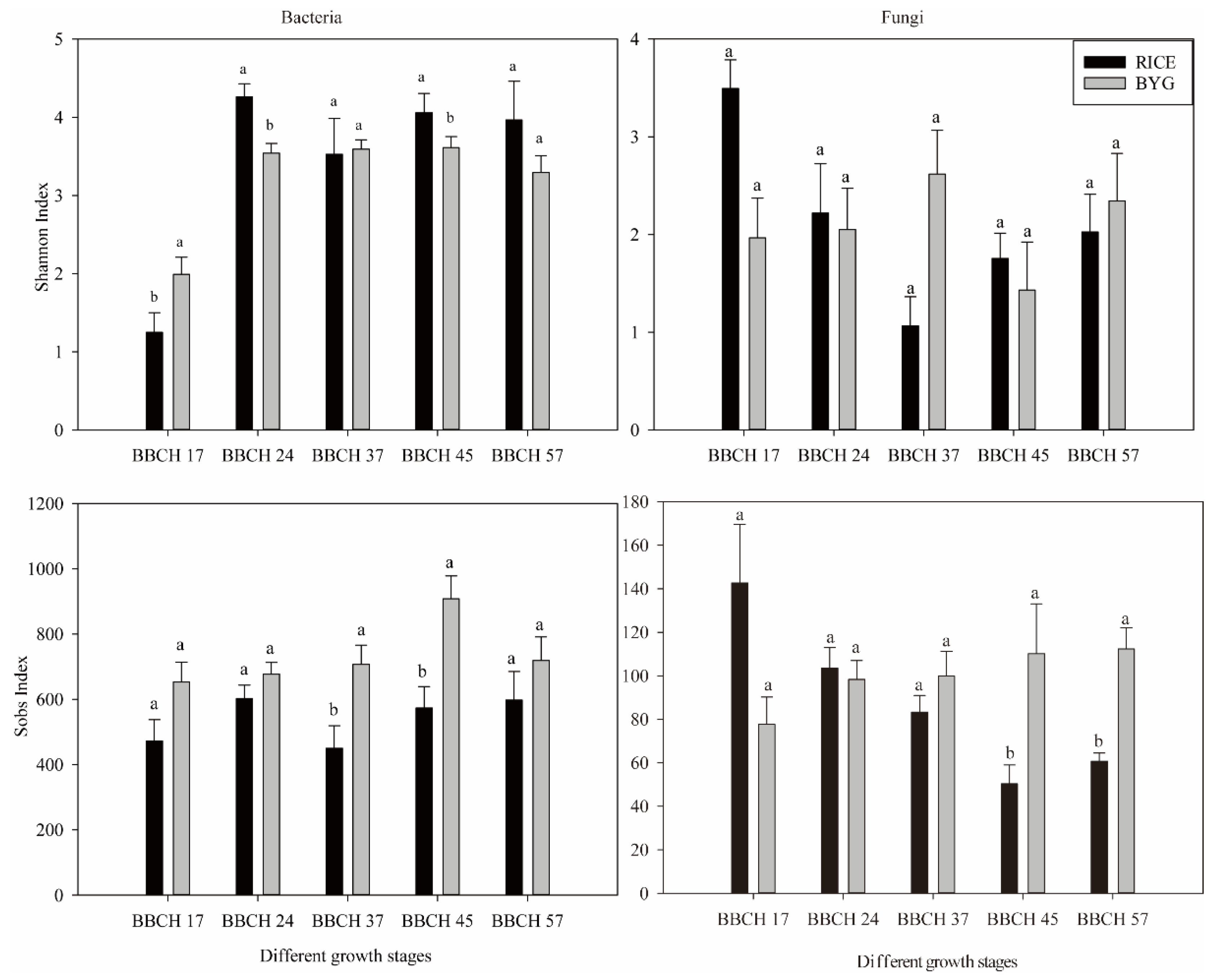
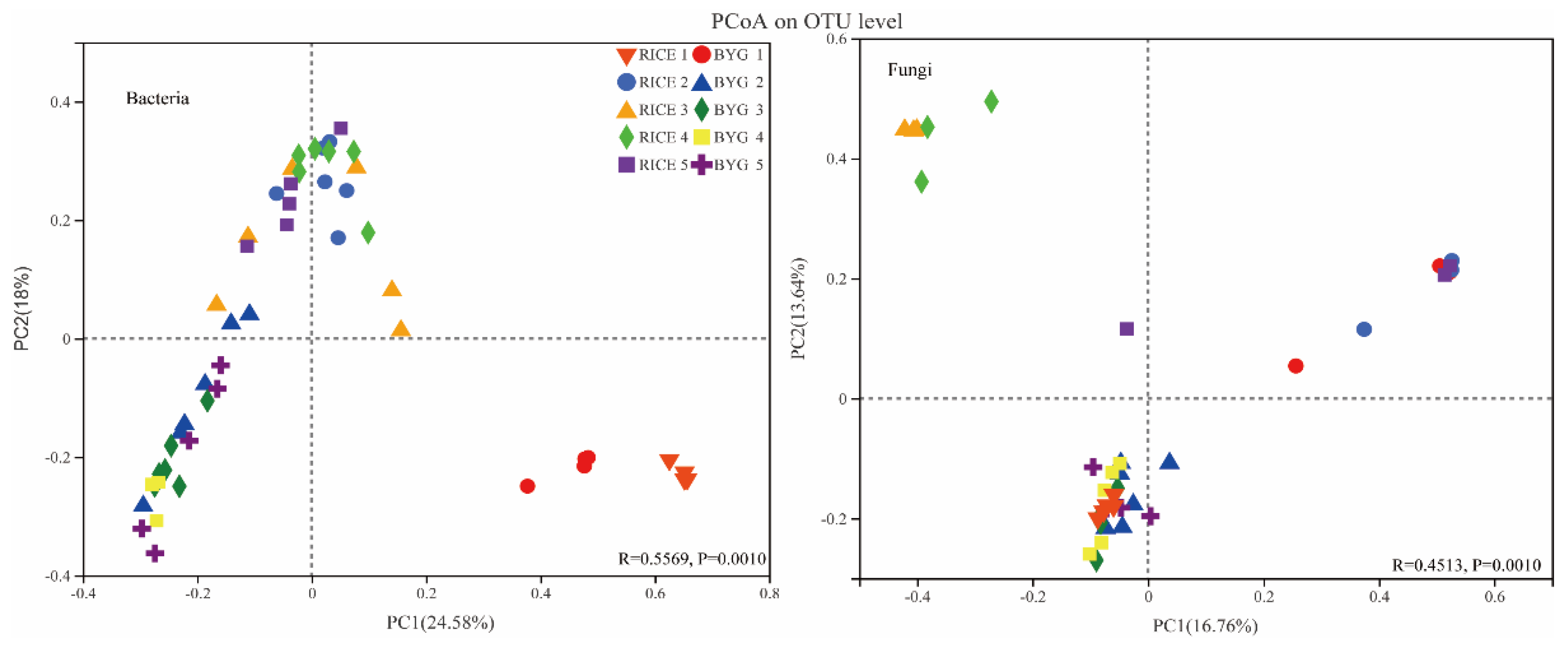
3.4. Alpha- and Beta-Diversity in Rice Roots Endophytic Communities
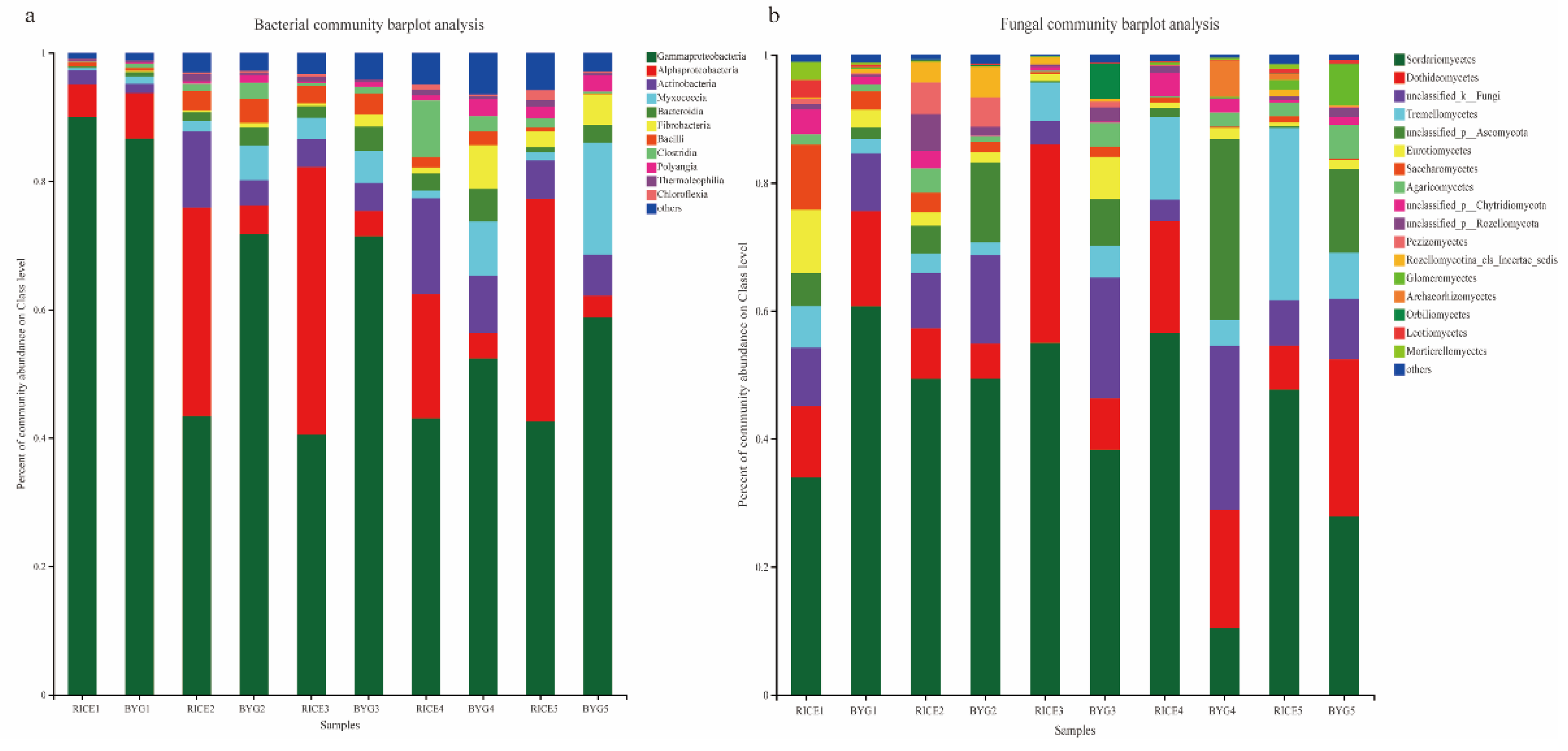
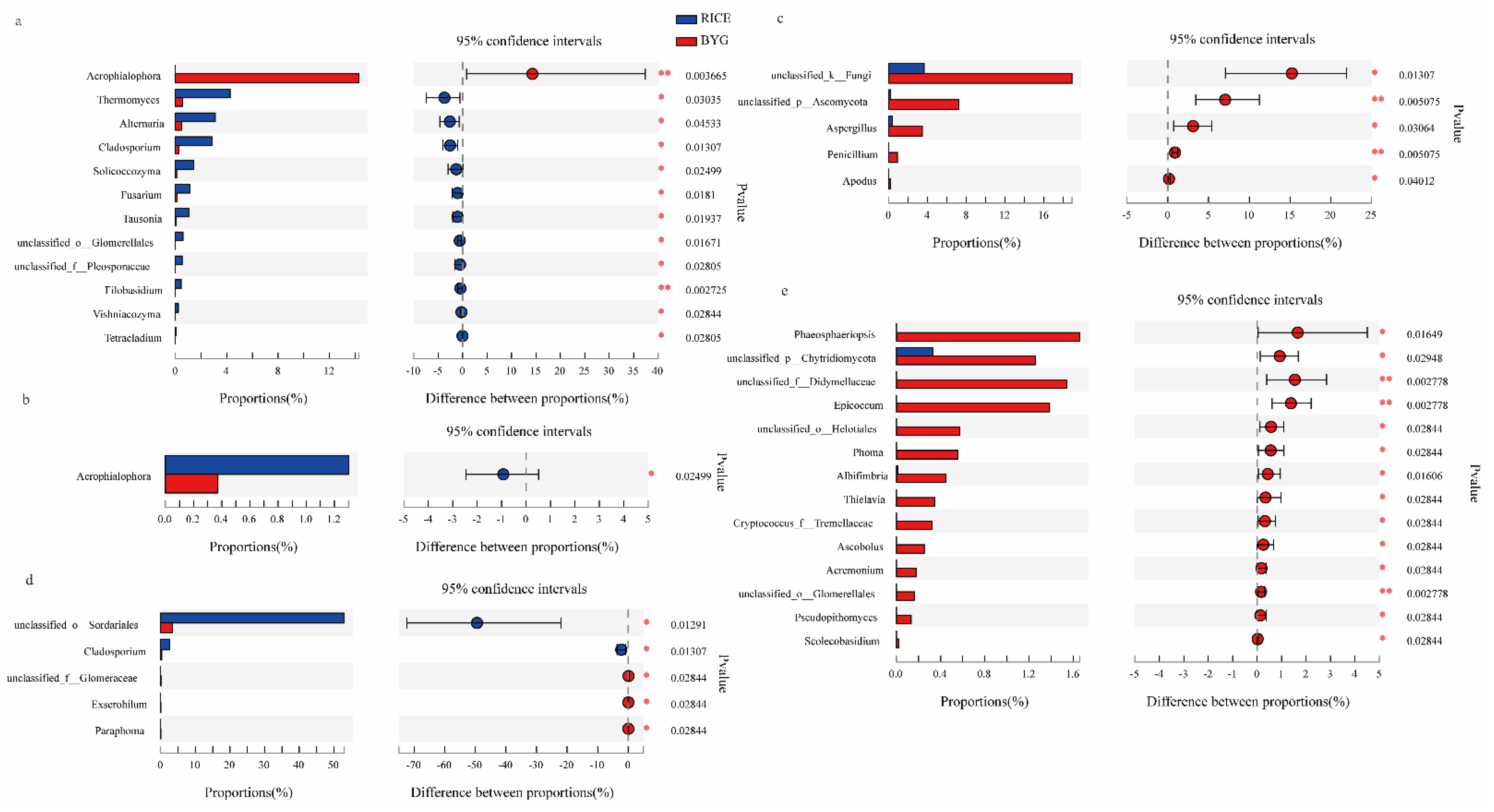
3.5. Endophytic Bacterial and Fungal Species Composition
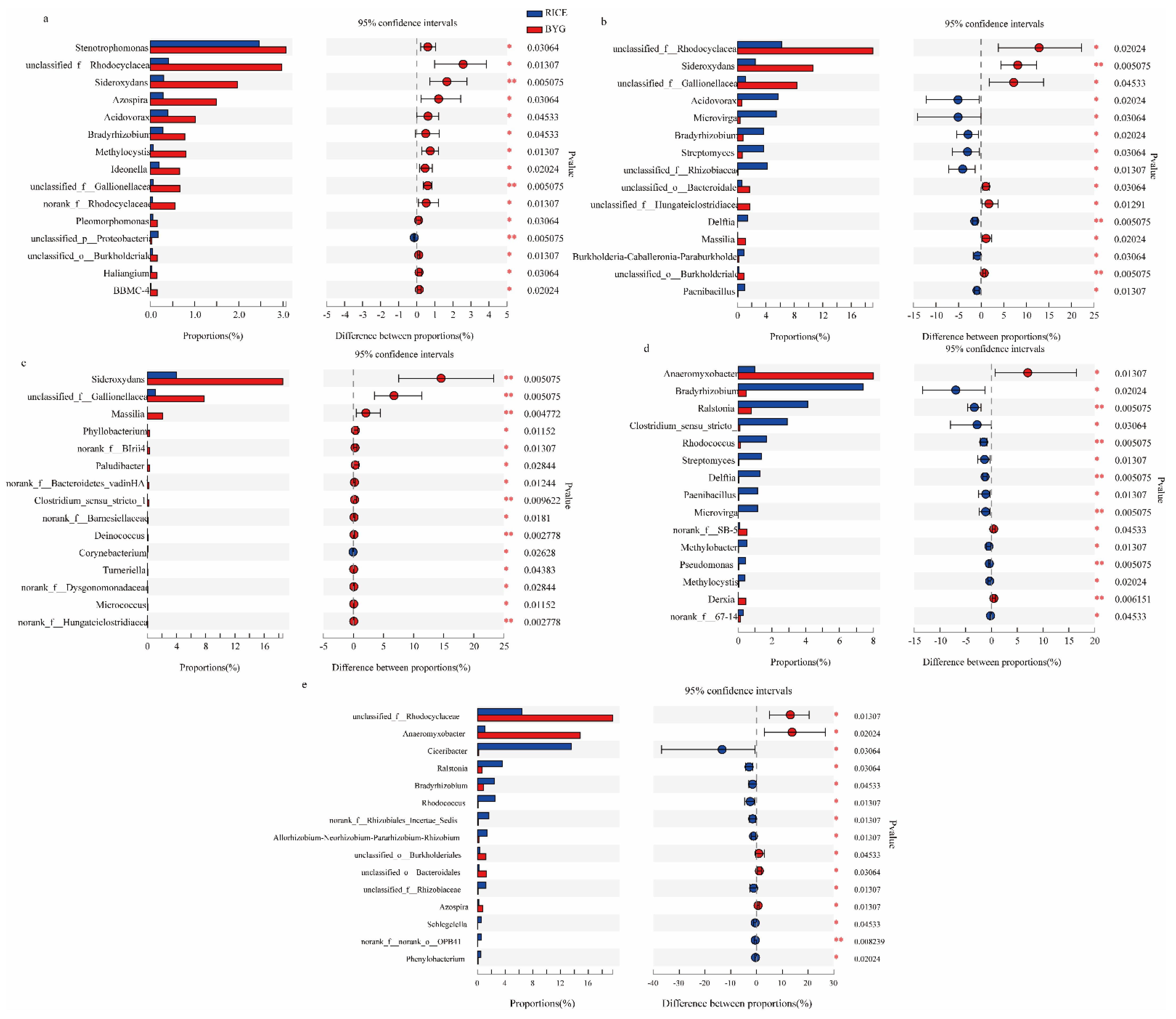
3.5.1. Bacterial Community Composition
3.5.2. Fungal Community Composition
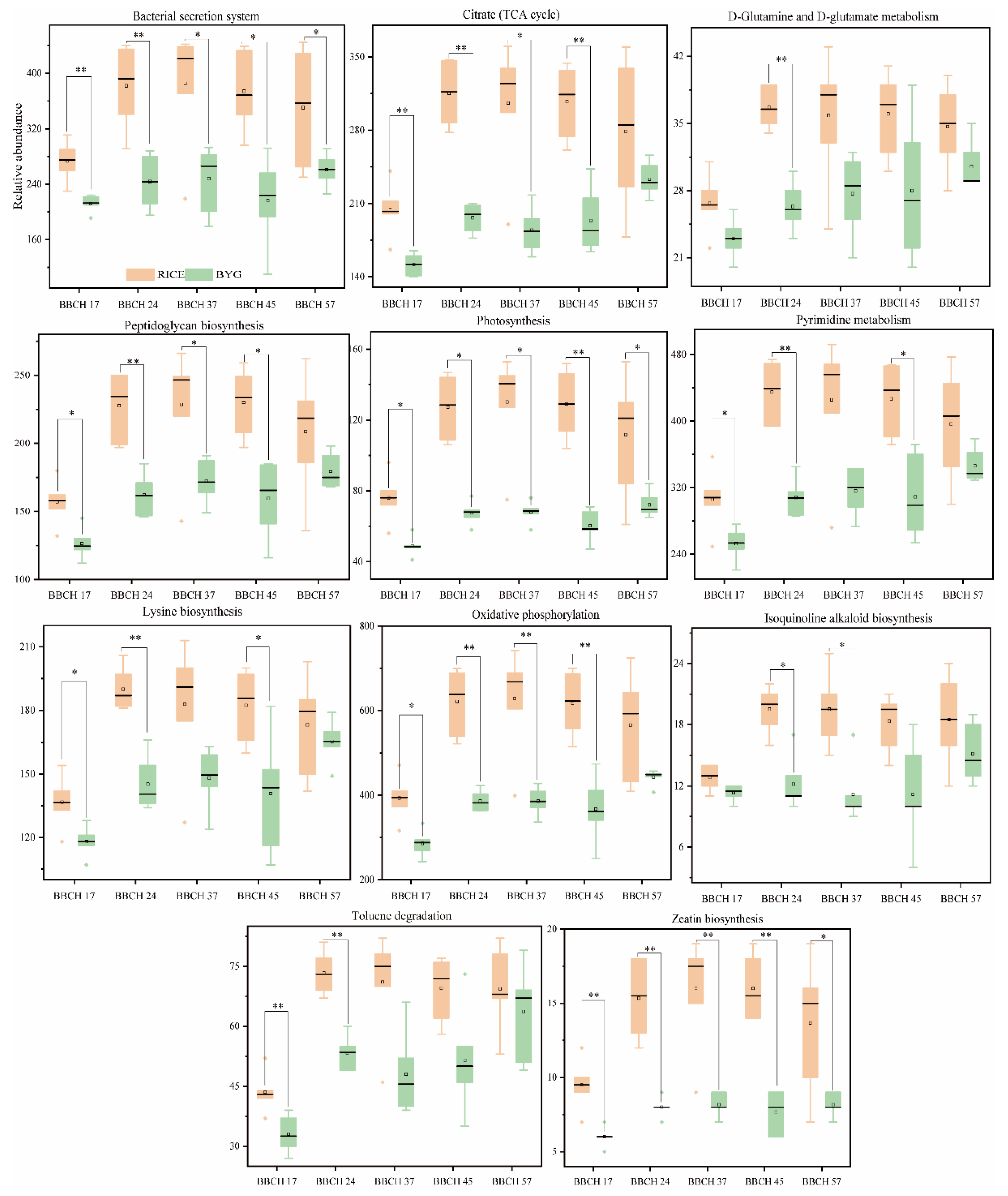
3.6. Bacterial and Fungal Metabolic Pathways differentially Represented in Rice and Barnyardgrass
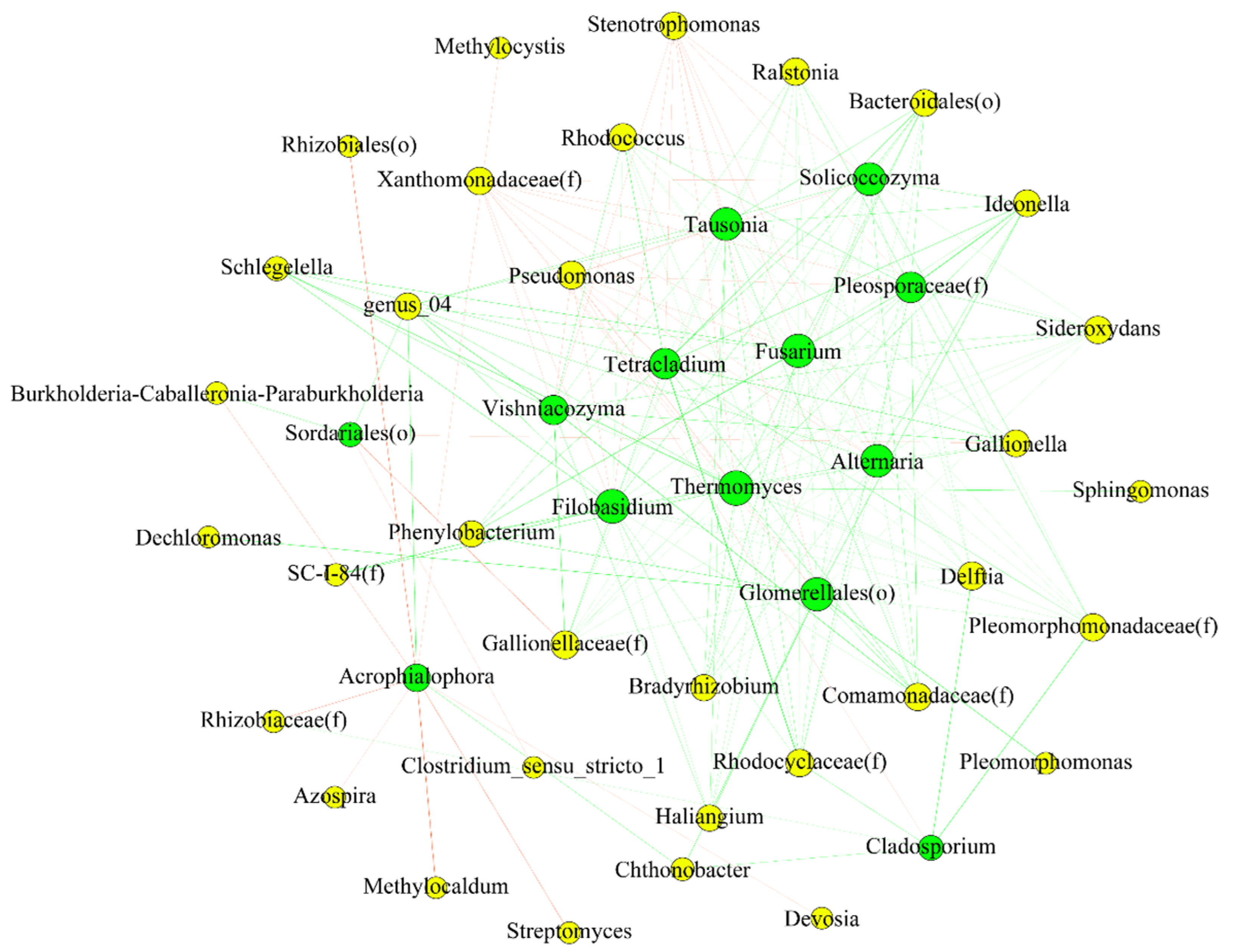
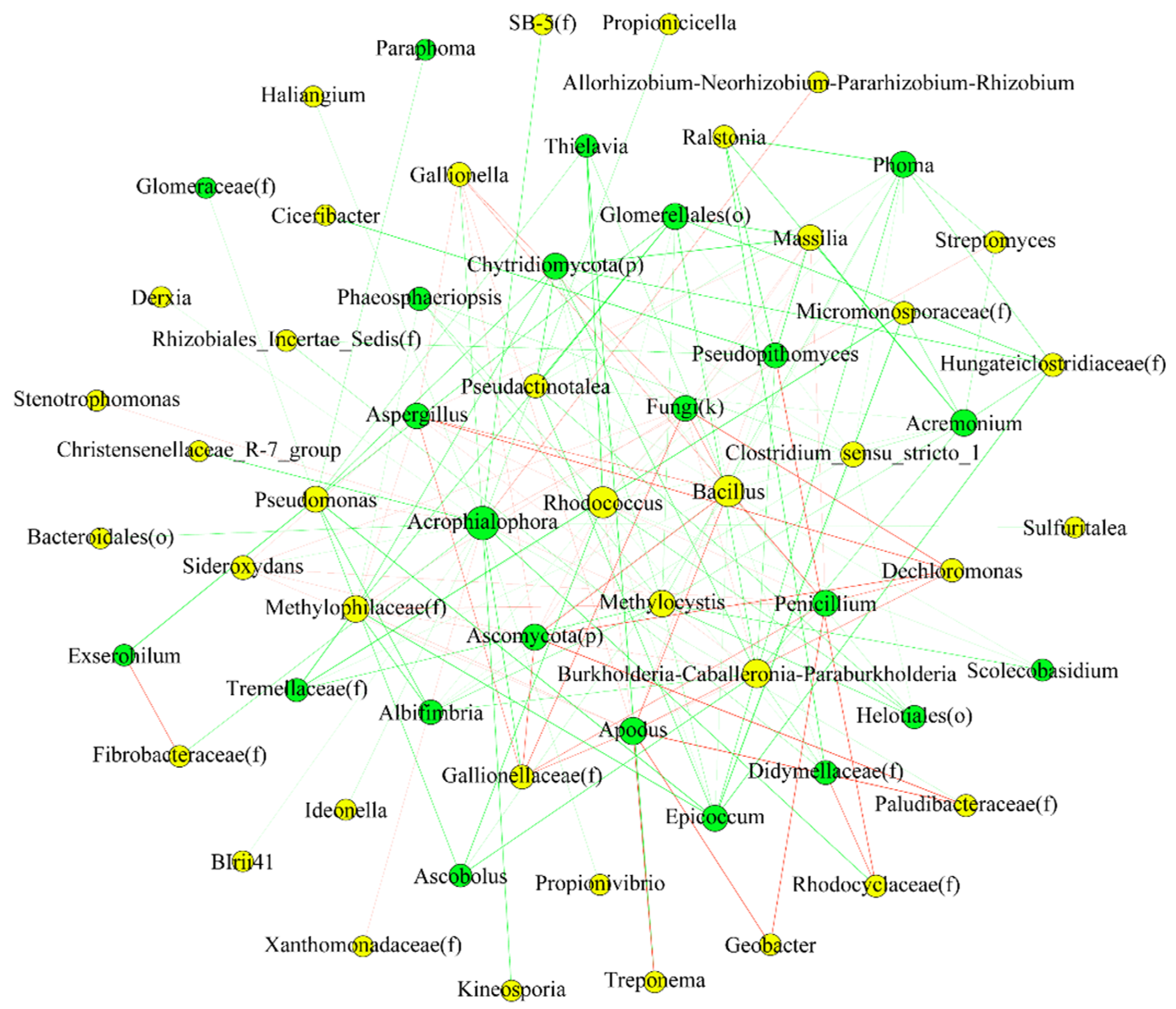
3.7. Interactions between Endophytic Bacterial and Fungal Communities
4. Discussion
4.1. Barnyardgrass mainly Affected the Endophytic Bacteria in Rice
4.2. Rice Relied on Fungi at Vegetative Stage and Bacteria at Reproductive Stage against Barnyardgrass Stress
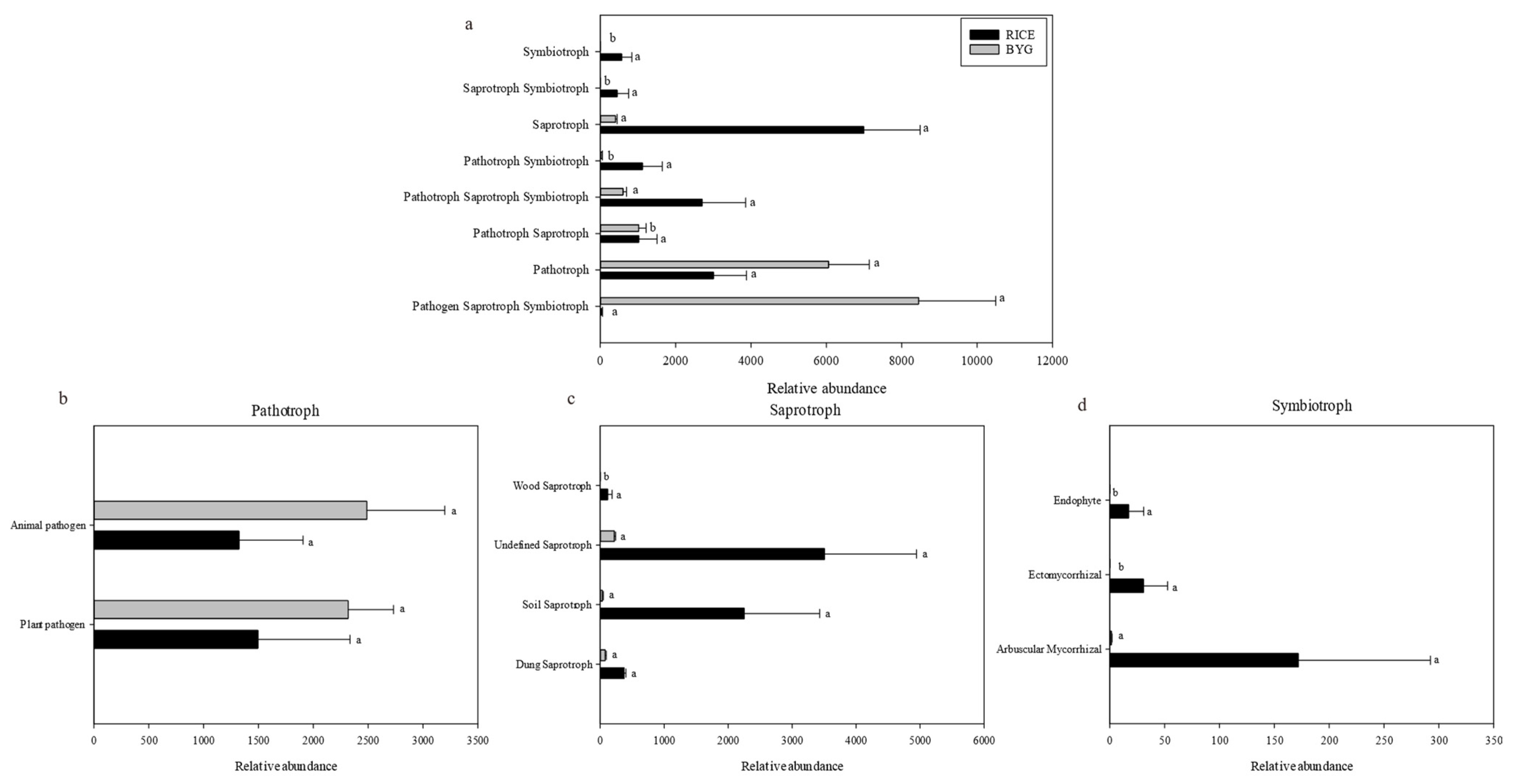
4.3. Rice Utilized Metabolism Pathways and Different Trophic Modes to Improve Its Defensive System
4.4. The Fungal Community May Be Predicted Based on the Bacterial One
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Phukan, J.; Kalita, S.; Bora, P. Weed Management in Direct Seeded Rice: A Review. J. Pharmacogn. Phytochem. 2021, 10, 742–748. [Google Scholar]
- Yang, X.F.; Kong, C.H.; Yang, X.; Li, Y.F. Interference of Allelopathic Rice with Penoxsulam-Resistant Barnyardgrass. Pest Manag. Sci. 2017, 73, 2310–2317. [Google Scholar] [CrossRef] [PubMed]
- Cui, H. Challenges and Approaches to Crop Improvement Through C3-to-C4 Engineering. Front. Plant Sci. 2021, 12, 715391. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.C.; Gu, T.; Zhao, B.; Yang, X.; Peng, Q.; Li, Y.; Bai, L.Y. Effects of Common Echinochloa Varieties on Grain Yield and Grain Quality of Rice. Field Crops Res. 2017, 203, 163–172. [Google Scholar] [CrossRef]
- Kong, C.H.; Zhang, S.Z.; Li, Y.H.; Xia, Z.C.; Yang, X.F.; Meiners, S.J.; Wang, P. Plant Neighbor Detection and Allelochemical Response Are Driven by Root-Secreted Signaling Chemicals. Nat. Commun. 2018, 9, 3867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, A.; Schmid, M.; van Tuinen, D.; Berg, G. Plant-Driven Selection of Microbes. Plant Soil 2009, 321, 235–257. [Google Scholar] [CrossRef]
- Nandy, S.; Das, T.; Tudu, C.K.; Pandey, D.K.; Dey, A.; Ray, P. Fungal Endophytes: Futuristic Tool in Recent Research Area of Phytoremediation. S. Afr. J. Bot. 2020, 134, 285–295. [Google Scholar] [CrossRef]
- Mishra, S.; Bhardwaj, P.; Sharma, S. Metabolomic Insights Into Endophyte-Derived Bioactive Compounds. Front. Microbiol. 2022, 13, 835931. [Google Scholar] [CrossRef]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant Beneficial Endophytic Bacteria: Mechanisms, Diversity, Host Range and Genetic Determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef]
- Jana, S.K.; Islam, M.M.; Mandal, S. Endophytic Microbiota of Rice and Their Collective Impact on Host Fitness. Curr. Microbiol. 2022, 79, 37. [Google Scholar] [CrossRef]
- Ganie, S.A.; Bhat, J.A.; Devoto, A. The Influence of Endophytes on Rice Fitness under Environmental Stresses. Plant Mol. Biol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Waqas, M.; Khan, A.L.; Shahzad, R.; Ullah, I.; Khan, A.R.; Lee, I.J. Mutualistic Fungal Endophytes Produce Phytohormones and Organic Acids That Promote Japonica Rice Plant Growth under Prolonged Heat Stress. J. Zhejiang Univ. Sci. B 2015, 16, 1011–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Singh, S.; Mukherjee, A.; Rastogi, R.P.; Verma, J.P. Salt-Tolerant Plant Growth-Promoting Bacillus pumilus Strain JPVS11 to Enhance Plant Growth Attributes of Rice and Improve Soil Health under Salinity Stress. Microbiol. Res. 2021, 242, 126616. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Cao, L.; Zeng, J.; Franco, C.M.M.; Yang, Y.; Hu, X.; Liu, Y.; Wang, X.; Gao, Y.; Bu, Z.; et al. The Antifungal Action Mode of the Rice Endophyte Streptomyces hygroscopicus OsiSh-2 as a Potential Biocontrol Agent against the Rice Blast Pathogen. Pestic. Biochem. Physiol. 2019, 160, 58–69. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, X.; Kong, C.H. The Response of Allelopathic Rice Growth and Microbial Feedback to Barnyardgrass Infestation in a Paddy Field Experiment. Eur. J. Soil Biol. 2013, 56, 26–32. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants: BBCH Monograph; Federal Biological Research Centre for Agriculture and Forestry: Berlin/Braunschweig, Germany, 2018; ISBN 978-3-95547-071-5. [Google Scholar]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Goebel, B.M. Taxonomic Note: A Place for DNA-DNA Reassociation and 16S RRNA Sequence Analysis in the Present Species Definition in Bacteriology. Int. J. Syst. Evol. Microbiol. 1994, 44, 846–849. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly Accurate OTU Sequences from Microbial Amplicon Reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian Classifier for Rapid Assignment of RRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Kong, C.H. Allelopathic Potential of Barnyard Grass on Rice and Soil Microbes in Paddy. Allelopath. J. 2008, 21, 389–395. [Google Scholar]
- Furtado, B.U.; Gołębiewski, M.; Skorupa, M.; Hulisz, P.; Hrynkiewicz, K. Bacterial and Fungal Endophytic Microbiomes of Salicornia Europaea. Appl. Environ. Microbiol. 2019, 85, e00305-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.Y.; Zhang, N.; Liu, Y.X.; Zhang, X.N.; Hu, B.; Qin, Y.; Xu, H.R.; Wang, H.; Guo, X.; Qian, J.; et al. Root Microbiota Shift in Rice Correlates with Resident Time in the Field and Developmental Stage. Sci. China Life Sci. 2018, 61, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Dyksma, S.; Bischof, K.; Fuchs, B.M.; Hoffmann, K.; Meier, D.; Meyerdierks, A.; Pjevac, P.; Probandt, D.; Richter, M.; Stepanauskas, R.; et al. Ubiquitous Gammaproteobacteria Dominate Dark Carbon Fixation in Coastal Sediments. ISME J 2016, 10, 1939–1953. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Shankhdhar, D.; Sharma, A.; Shankhdhar, S.C. Growth Promotion of the Rice Genotypes by Pgprs Isolated from Rice Rhizosphere. J. Soil Sci. Plant Nutr. 2014, 14, 505–517. [Google Scholar] [CrossRef] [Green Version]
- Gholamalizadeh, R.; Khodakaramian, G.; Ebadi, A.A. Assessment of Rice Associated Bacterial Ability to Enhance Rice Seed Germination and Rice Growth Promotion. Braz. Arch. Biol. Technol. 2017, 60, e17160410. [Google Scholar] [CrossRef] [Green Version]
- Reid, S.J.; Abratt, V.R. Sucrose Utilisation in Bacteria: Genetic Organisation and Regulation. Appl. Microbiol. Biotechnol. 2005, 67, 312–321. [Google Scholar] [CrossRef]
- Gealy, D.; Wailes, E.; Estorninos, L.W., Jr.; Chavez, R. Rice Cultivar Differences in Suppression of Barnyardgrass (Echinochloa crus-galli) and Economics of Reduced Propanil Rates. Weed Sci. 2009, 51, 601–609. [Google Scholar] [CrossRef]
- Ji, S.H.; Gururani, M.A.; Chun, S.C. Isolation and Characterization of Plant Growth Promoting Endophytic Diazotrophic Bacteria from Korean Rice Cultivars. Microbiol. Res. 2014, 169, 83–98. [Google Scholar] [CrossRef]
- Braga, R.M.; Padilla, G.; Araújo, W.L. The Biotechnological Potential of Epicoccum spp.: Diversity of Secondary Metabolites. Crit. Rev. Microbiol. 2018, 44, 759–778. [Google Scholar] [CrossRef]
- Zhang, Z.C.; Cao, J.J.; Gu, T.; Yang, X.; Li, Y.F. Co-Planted Barnyardgrass Reduces Rice Yield by Inhibiting Plant above-and Belowground-Growth during Post-Heading Stages. Crop J. 2021, 9, 1198–1207. [Google Scholar] [CrossRef]
- Lengeler, J.W.; Titgemeyer, F.; Vogler, A.P.; Wöhrl, B.M. Structures and Homologies of Carbohydrate: Phosphotransferase System (PTS) Proteins. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1990, 326, 489–504. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Mhamdi, A.; Chaouch, S.; Han, Y.; Neukermans, J.; Marquez-Garcia, B.; Queval, G.; Foyer, C.H. Glutathione in Plants: An Integrated Overview. Plant Cell Environ. 2012, 35, 454–484. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, F.X.; Pogson, B.; Sun, Z.; McDonald, K.A.; DellaPenna, D.; Gantt, E. Functional Analysis of the Beta and Epsilon Lycopene Cyclase Enzymes of Arabidopsis Reveals a Mechanism for Control of Cyclic Carotenoid Formation. Plant Cell 1996, 8, 1613–1626. [Google Scholar] [CrossRef] [Green Version]
- Maoka, T. Carotenoids as Natural Functional Pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Lavín, J.L.; Marcet-Houben, M.; Gutiérrez-Vázquez, R.L.; Ramírez, L.; Pisabarro, A.G.; Gabaldón, T.; Oguiza, J.A. FUNGALOXPHOS: An Integrated Database for Oxidative Phosphorylation in Fungi. Mitochondrion 2013, 13, 357–359. [Google Scholar] [CrossRef]
- Akram, M. Citric Acid Cycle and Role of Its Intermediates in Metabolism. Cell Biochem. Biophys. 2014, 68, 475–478. [Google Scholar] [CrossRef]
- Gu, Z.C.; Wang, M.; Wang, Y.; Zhu, L.X.; Mur, L.A.J.; Hu, J.; Guo, S. Nitrate Stabilizes the Rhizospheric Fungal Community to Suppress Fusarium Wilt Disease in Cucumber. Mol. Plant Microbe Interact. 2020, 33, 590–599. [Google Scholar] [CrossRef] [Green Version]
- Crowther, T.W.; Boddy, L.; Hefin Jones, T. Functional and Ecological Consequences of Saprotrophic Fungus–Grazer Interactions. ISME J. 2012, 6, 1992–2001. [Google Scholar] [CrossRef] [Green Version]
- Barna, B.; Fodor, J.; Harrach, B.D.; Pogány, M.; Király, Z. The Janus Face of Reactive Oxygen Species in Resistance and Susceptibility of Plants to Necrotrophic and Biotrophic Pathogens. Plant Physiol. Biochem. 2012, 59, 37–43. [Google Scholar] [CrossRef]
- Doehlemann, G.; Ökmen, B.; Zhu, W.; Sharon, A. Plant Pathogenic Fungi. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- de Boer, W.; Folman, L.B.; Summerbell, R.C.; Boddy, L. Living in a Fungal World: Impact of Fungi on Soil Bacterial Niche Development. FEMS Microbiol. Rev. 2005, 29, 795–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendrickson, O. Abundance and Activity of N2-Fixing Bacteria in Decaying Wood. Can. J. For. Res. 2011, 21, 1299–1304. [Google Scholar] [CrossRef]
- Whipps, J.M. Microbial Interactions and Biocontrol in the Rhizosphere. J. Exp. Bot. 2001, 52, 487–511. [Google Scholar] [CrossRef]
- Weller, D.M.; Raaijmakers, J.M.; Gardener, B.B.M.; Thomashow, L.S. Microbial Populations Responsible for Specific Soil Suppressiveness to Plant Pathogens. Annu. Rev. Phytopathol. 2002, 40, 309–348. [Google Scholar] [CrossRef] [Green Version]
- Wheatley, R.E. The Consequences of Volatile Organic Compound Mediated Bacterial and Fungal Interactions. Antonie Leeuw Int. J. G 2002, 81, 357–364. [Google Scholar] [CrossRef]
- de Boer, W.; de Ridder-Duine, A.S.; Klein Gunnewiek, P.J.A.; Smant, W.; Van Veen, J.A. Rhizosphere Bacteria from Sites with Higher Fungal Densities Exhibit Greater Levels of Potential Antifungal Properties. Soil Biol Biochem. 2008, 40, 1542–1544. [Google Scholar] [CrossRef]
- Leveau, J.H.J.; Preston, G.M. Bacterial Mycophagy: Definition and Diagnosis of a Unique Bacterial-Fungal Interaction. New Phytol. 2008, 177, 859–876. [Google Scholar] [CrossRef]
- Rediers, H.; Bonnecarrère, V.; Rainey, P.B.; Hamonts, K.; Vanderleyden, J.; De Mot, R. Development and Application of a DapB-Based in Vivo Expression Technology System to Study Colonization of Rice by the Endophytic Nitrogen-Fixing Bacterium Pseudomonas Stutzeri A15. Appl. Environ. Microbiol. 2003, 69, 6864–6874. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Shannon Index | Chao Index | Coverage Index | |
|---|---|---|---|---|
| Bacteria | soil 1 | 2.97 ± 0.07 a | 144.60 ± 62.58 a | 0.18 ± 0.10 a |
| soil 2 | 2.95 ± 0.01 a | 131.00 ± 34.64 a | 0.16 ± 0.01 a | |
| soil 3 | 2.92 ± 0.01 a | 120.00 ± 40.99 a | 0.20 ± 0.04 a | |
| Fungi | soil 1 | 3.13 ± 0.22 a | 515.43 ± 44.59 a | 0.99 ± 0.00024 a |
| soil 2 | 3.37 ± 0.04 a | 542.99 ± 6.77 a | 1.00 ± 0.00004 a | |
| soil 3 | 3.36 ± 0.02 a | 528.37 ± 7.63 a | 1.00 ± 0.00013 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Yan, Q.; Wang, J.; Peng, Q. Endophytic Fungal and Bacterial Microbiota Shift in Rice and Barnyardgrass Grown under Co-Culture Condition. Plants 2022, 11, 1592. https://doi.org/10.3390/plants11121592
Li S, Yan Q, Wang J, Peng Q. Endophytic Fungal and Bacterial Microbiota Shift in Rice and Barnyardgrass Grown under Co-Culture Condition. Plants. 2022; 11(12):1592. https://doi.org/10.3390/plants11121592
Chicago/Turabian StyleLi, Shuyan, Qiling Yan, Jieyu Wang, and Qiong Peng. 2022. "Endophytic Fungal and Bacterial Microbiota Shift in Rice and Barnyardgrass Grown under Co-Culture Condition" Plants 11, no. 12: 1592. https://doi.org/10.3390/plants11121592