
Adnate Leaf-Base and the Origin of Ribs in Succulent Stems of Euphorbia L.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
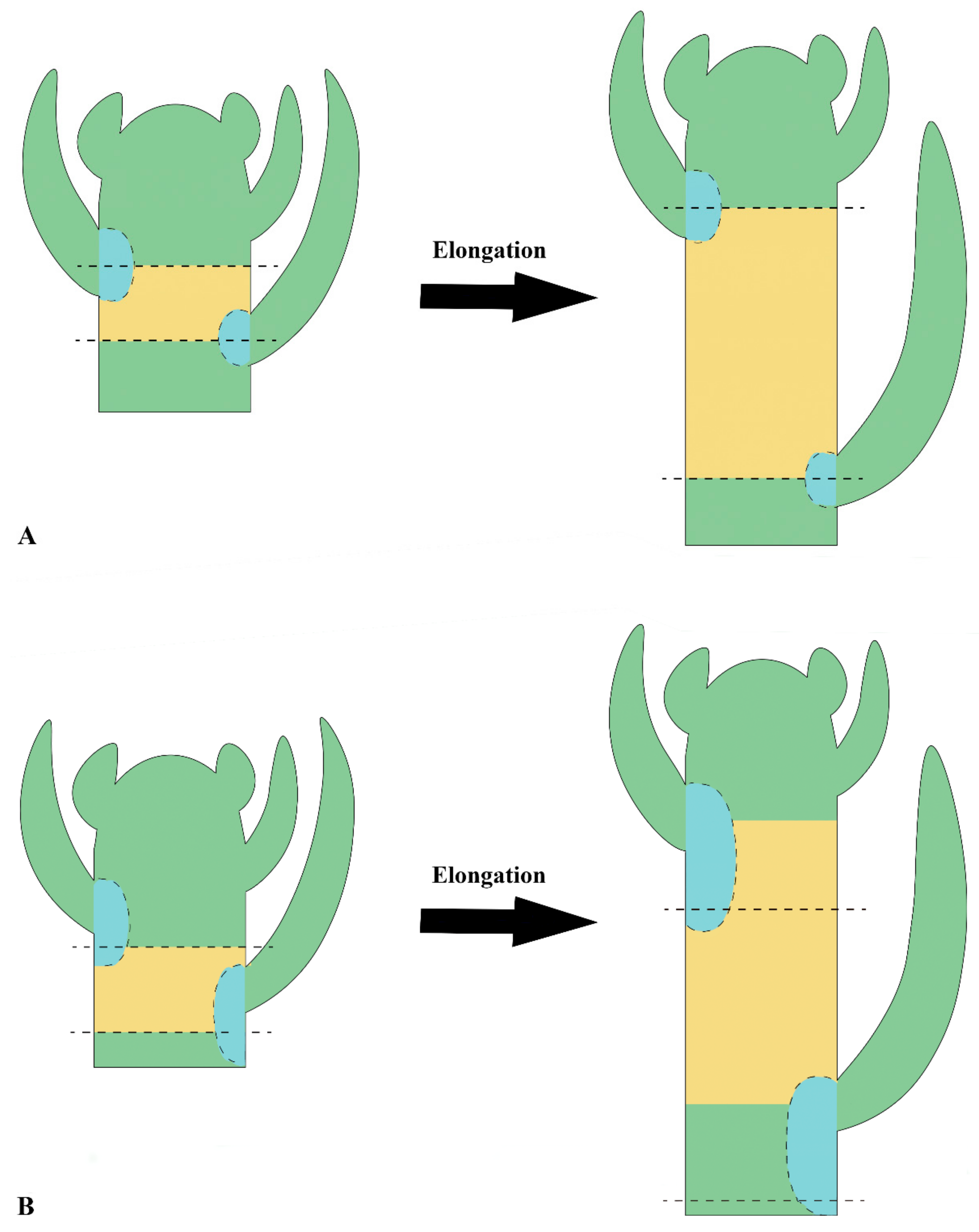{kind=link}
Abstract
:1. Introduction
2. Results
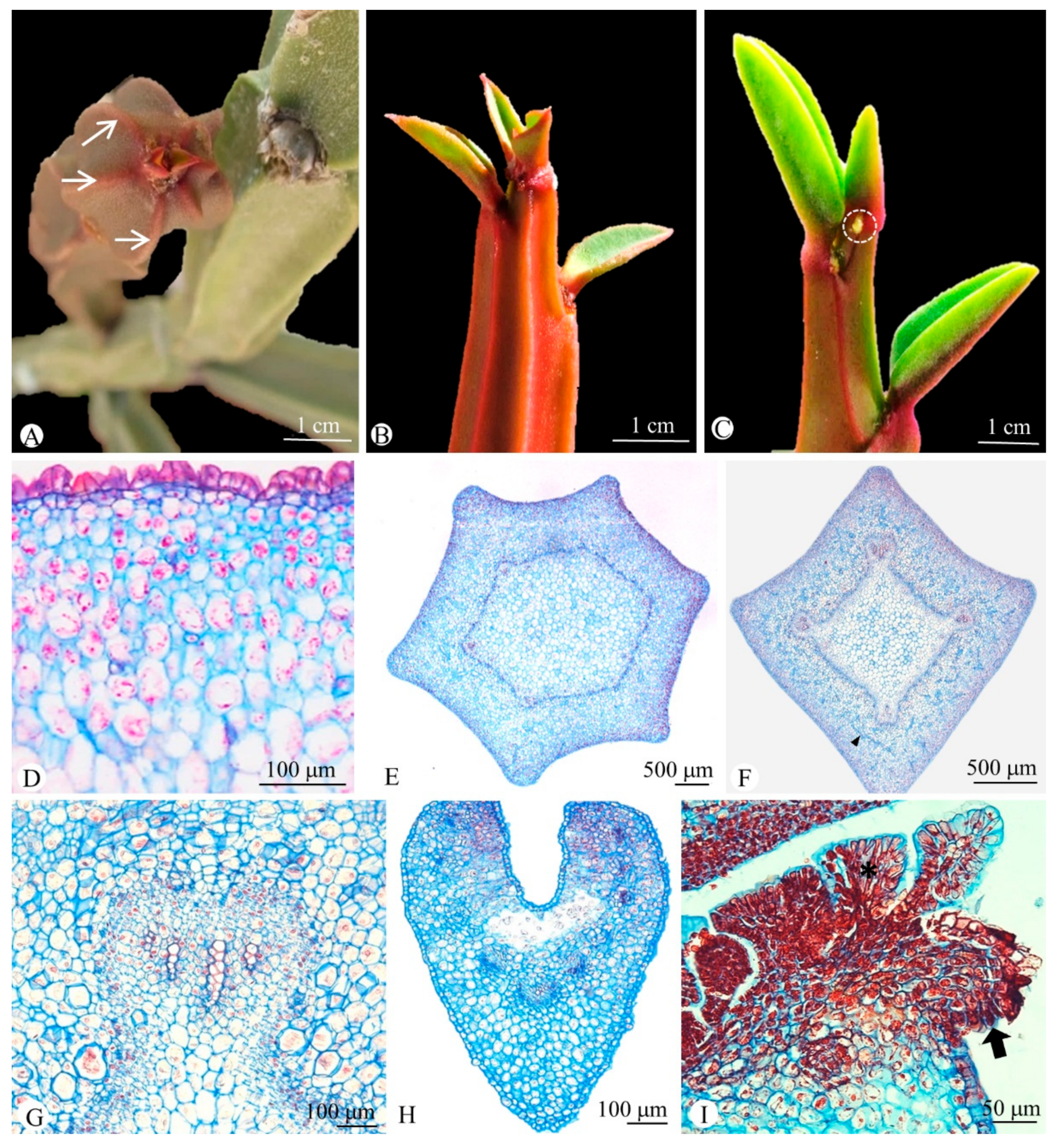
2.1. Anatomy
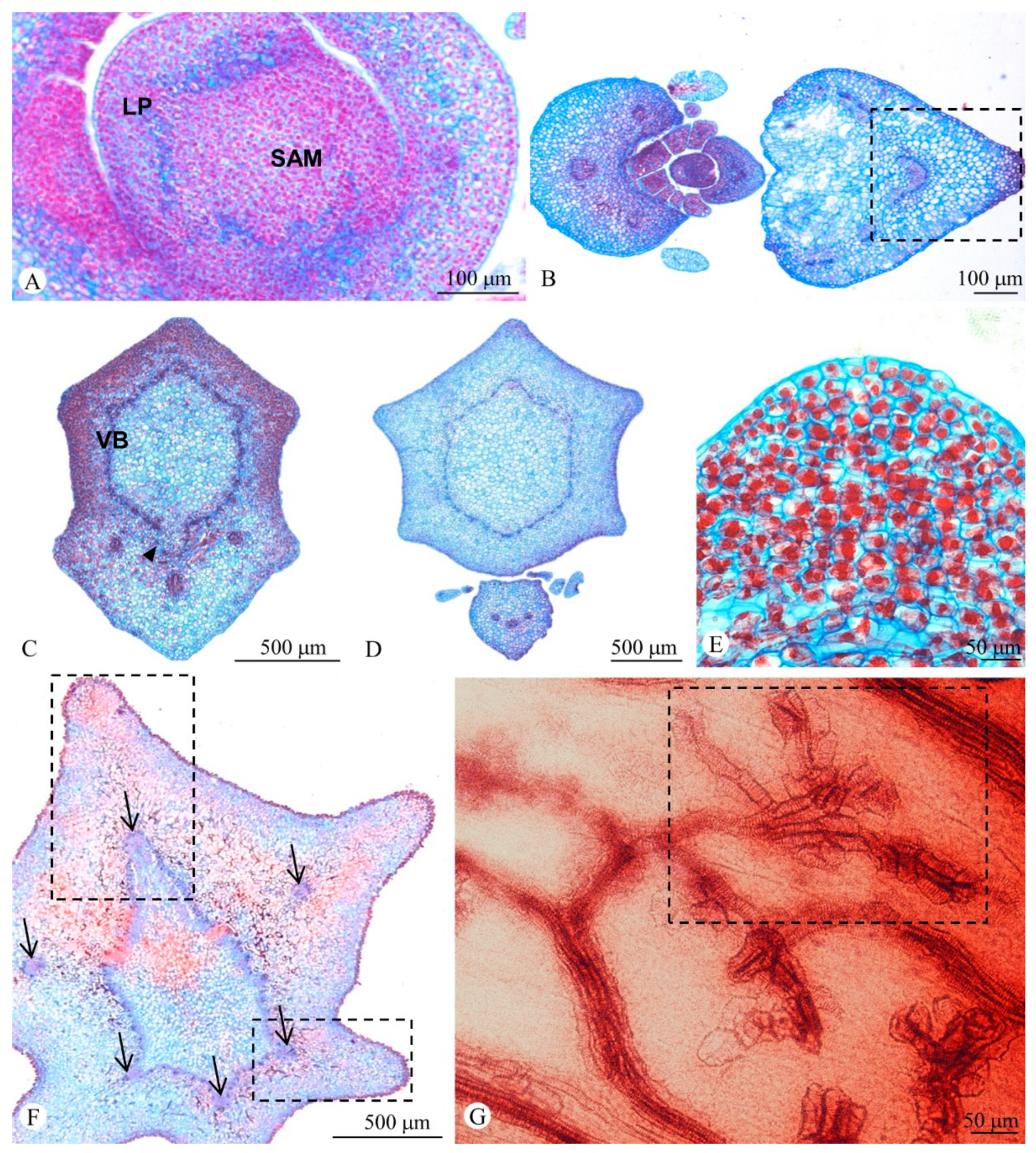
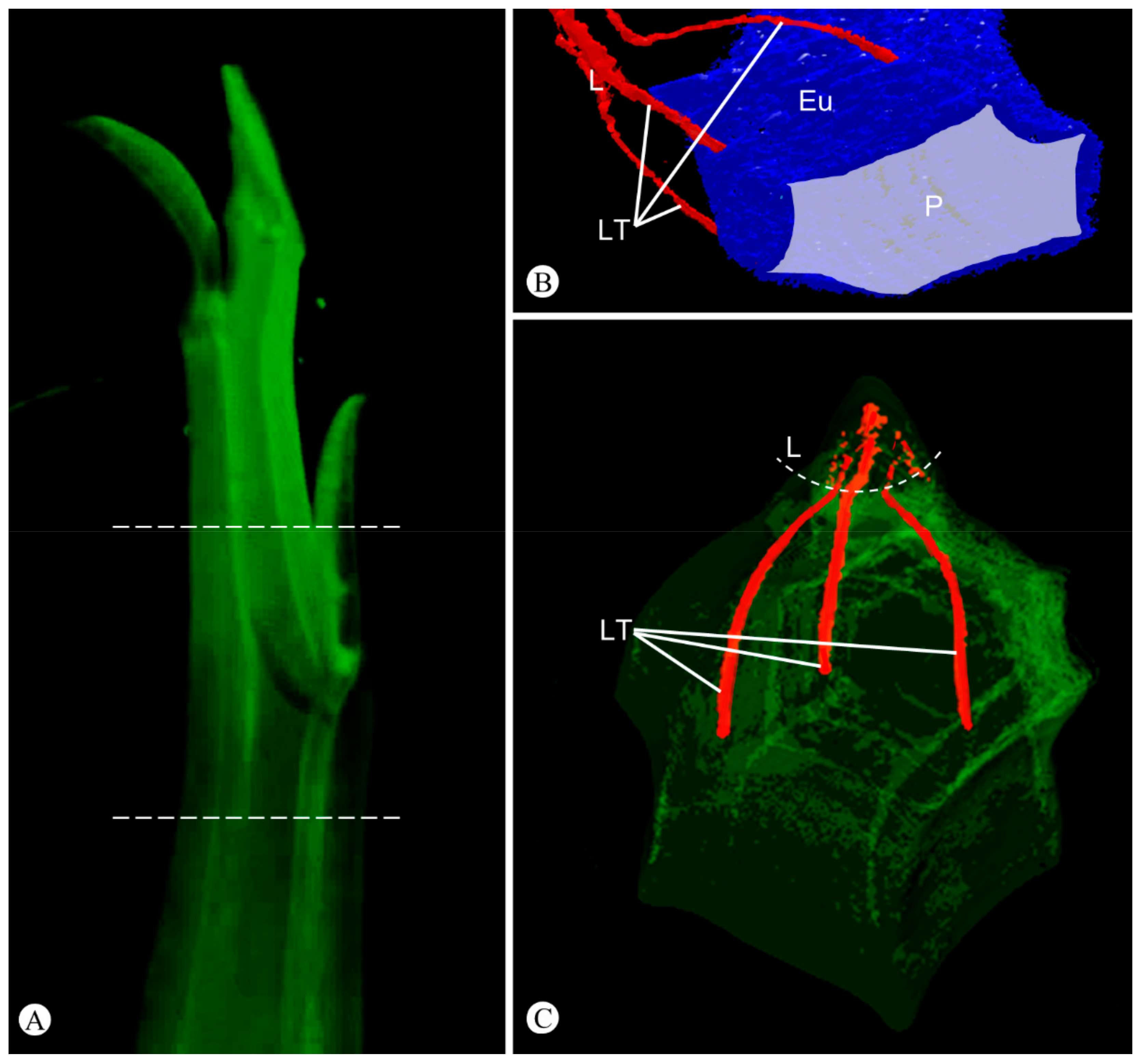
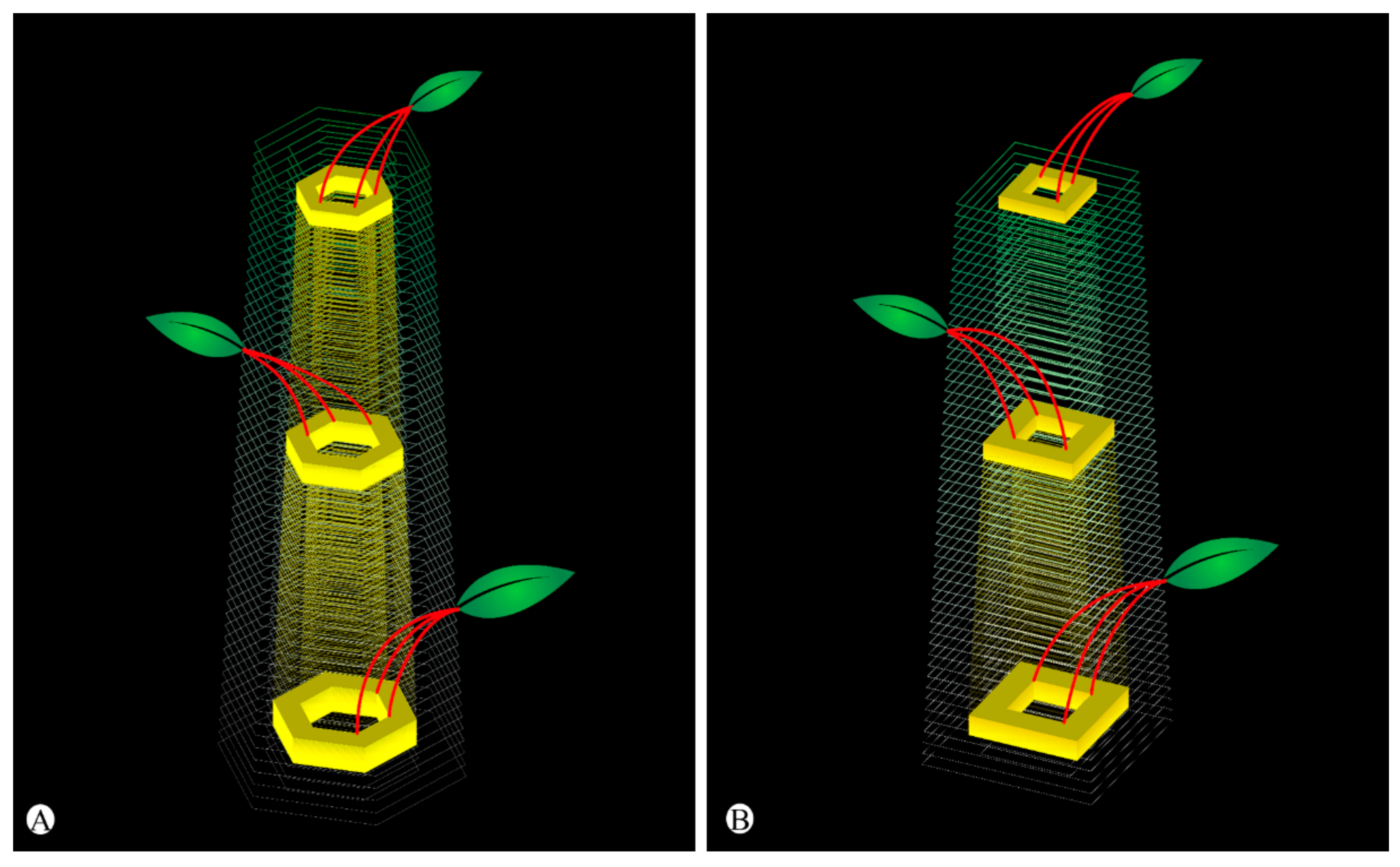
2.2. Shoot Ontogeny and Vasculature
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jacobsen, H. A Handbook of Succulent Plants; Blandford Press: London, UK, 1960. [Google Scholar]
- Albers, F.; Meve, U. Illustrated Handbook of Succulent Plants: Asclepiadaceae; Springer: Berlin, Germany, 2002. [Google Scholar]
- Eggli, U. Illustrated Handbook of Succulent Plants: Monocotyledons; Springer: Berlin, Germany, 2001. [Google Scholar]
- Eggli, U. Illustrated Handbook of Succulent Plants: Dicotyledons; Springer: Berlin, Germany, 2002. [Google Scholar]
- Eggli, U. Illustrated Handbook of Succulent Plants: Crassulaceae; Springer: Berlin, Germany, 2003. [Google Scholar]
- von Willert, D.B.; Eller, M.; Werger, M.J.A.; Brinckmann, E.; Ihlenfeldt, H.D. Life Strategies of Succulents in Deserts with Special Reference to the Namib Desert; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Gibson, A.C. The Anatomy of Succulence. In Crassulacean Acid Metabolism: Proceedings of the Fifth Annual Symposium in Botany, Los Angeles, CA, USA, 14–16 January 1982; Ting, I.P., Gibbs, M., Eds.; American Society of Plant Physiologists: Rockville, MD, USA, 1982; pp. 1–15. [Google Scholar]
- Gibson, A.C.; Nobel, P.S. The Cactus Primer; Harvard University Press: Cambridge, MA, USA, 1986. [Google Scholar]
- Mauseth, J.D. The structure of photosynthetic succulent stems in plants other than cacti. Int. J. Plant Sci. 2004, 165, 1–9. [Google Scholar] [CrossRef]
- Raven, P.H.; Evert, R.F.; Eichhorn, S.E. Biology of Plants, 4th ed.; Worth Publishers: New York, NY, USA, 1986. [Google Scholar]
- Futuyma, D.J. Evolutionary Biology, 3rd ed.; Sinauer Associates: Sunderland, MA, USA, 1997. [Google Scholar]
- Niklas, K. The Evolutionary Biology of Plants; The University of Chicago Press: Chicago, IL, USA, 1997. [Google Scholar]
- Stearn, S.C.; Hoekstra, R.F. Evolution: An Introduction, 2nd ed.; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Hearn, D.J. Developmental patterns in anatomy are shared among separate evolutionary origins of stem succulent and storage root-bearing growth habits in Adenia (Passifloraceae). Am. J. Bot. 2009, 96, 1941–1956. [Google Scholar] [CrossRef] [PubMed]
- Ihlenfeldt, H.D. Lebensformen und Überlebensstrategien bei Sukkulenten. Ber. Deut. Bot. Ges. 1985, 98, 409–423. [Google Scholar]
- Sajeva, M.; Costanzo, M. Succulents: The Illustrated Dictionary; Timber Press: Portland, OR, USA, 1994. [Google Scholar]
- Sajeva, M.; Costanzo, M. Succulents II: The New Illustrated Dictionary; Le Lettere: Florence, Italy, 2000. [Google Scholar]
- Melo-de-Pinna, G.F.A.; Ogura, A.S.; Arruda, E.C.P.; Klak, C. Repeated evolution of endoscopic peripheral vascular bundles in succulent leaves of Aizoaceae (Caryophyllales). Taxon 2014, 63, 1037–1052. [Google Scholar] [CrossRef]
- Melo-de-Pinna, G.F.A.; Hernandes-Lopes, J.; Ogura, A.S.; Santos, L.K.; Silva, D.C.; Haevermans, T. Growth patterns and different arrangements of vascular tissues in succulent leaves. Int. J. Plant. Sci. 2016, 177, 643–660. [Google Scholar] [CrossRef]
- Ogura, A.S.; Hernandes-Lopes, J.; Melo-de-Pinna, G.F.A. A new anatomical interpretation for abaxialization in unifacial leaf blade of stone plants (Aizoaceae, Caryophyllales). Braz. J. Bot. 2018, 41, 751–764. [Google Scholar] [CrossRef]
- Mauseth, J.D.; Kiesling, R. Comparative anatomy of Neoraimondia roseiflora and Neocardenasia herzogiana (Cactaceae). Haseltonia 1997, 5, 37–50. [Google Scholar]
- Mauseth, J.D. Theoretical aspects of surface-to-volume ratios and water-storage capacities of succulent shoots. Am. J. Bot. 2000, 88, 1107–1115. [Google Scholar] [CrossRef]
- Anderson, E.F. The Cactus Family; Timber Press: Portland, OR, USA, 2001. [Google Scholar]
- Terrazas, S.T.; Mauseth, J.D. Shoot Anatomy and Morphology. In Cacti: Biology and Uses; Nobel, P.S., Ed.; University of California Press: Berkeley, CA, USA, 2002; pp. 23–40. [Google Scholar]
- Horn, J.W.; van Ee, B.W.; Morawetz, J.J.; Riina, R.; Steinmann, V.W.; Berry, P.E.; Wurdack, K.J. Phylogenetics and the evolution of major structural characters in the giant genus Euphorbia L. (Euphorbiaceae). Mol. Phylogenet. Evol. 2012, 63, 305–326. [Google Scholar] [CrossRef]
- Frodin, D.G. History and concepts of big plant genera. Taxon 2004, 53, 753–776. [Google Scholar]
- Dorsey, B.L.; Haevermans, T.; Aubriot, X.; Morawetz, J.J.; Riina, R.G.; Steinmann, V.W.; Berry, P.E. Phylogenetics, morphological evolution, and classification of Euphorbia subgenus Euphorbia. Taxon 2013, 62, 291–315. [Google Scholar] [CrossRef] [Green Version]
- Edwards, E.J.; Nyffeler, R.; Donoghue, M.J. Basal cactus phylogeny: Implications of the Pereskia (Cactaceae) paraphyly for the transition to the cactus life form. Am. J. Bot. 2005, 92, 1177–1188. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Hernández, T.; Hernández, H.M.; Arturo De-Nova, J.; Puente, R.; Eguiarte, L.E.; Magallón, S. Phylogenetic relationships and evolution of growth form in Cactaceae (Caryophyllales, Eudicotyledoneae). Am. J. Bot. 2011, 98, 44–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinmann, V.W. The Evolution of Succulence in the New World Species of Euphorbia (Euphorbiaceae). Ph.D. Thesis, Claremont Graduate University, Claremont, CA, USA, 2001. [Google Scholar]
- Hurbath, F. Sistemática, Estrutura Genética de Populações e Filogeografia de Euphorbia sect. Brasilienses V.W. Steinm. & Dorsey (Euphorbiaceae). Ph.D. Thesis, Instituto de Botânica, São Paulo, SP, Brazil, 2018. [Google Scholar]
- Solereder, H. Systematic Anatomy of the Dicotyledons, 2nd ed.; Clarendon Press: Oxford, UK, 1908. [Google Scholar]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledons; Clarendon Press: Oxford, UK, 1950. [Google Scholar]
- Bobich, E.G.; North, G.B. Structural implications of succulence: Architecture, anatomy, and mechanics of photosynthetic stem succulents, pachycauls, and leaf succulents. In Perspectives in Biophysical Plant Ecophysiology: A Tribute to Park S. Nobel; De la Barrera, E., Smith, W.K., Eds.; Universidad Nacional Autónoma de México: Mexico City, Mexico, 2009; pp. 3–37. [Google Scholar]
- Rowley, G.D. Caudiciform and Pachycaul Succulents: Pachycauls, Bottle-, Barrel- and Elephant Trees and Their Kin; Strawberry Press: Mill Valley, CA, USA, 1987. [Google Scholar]
- Eames, A.J.; MacDaniels, L.H. An Introduction to Plant Anatomy; McGraw-Hill: New York, NY, USA, 1947. [Google Scholar]
- Esau, K. Anatomy of Seed Plants, 2nd ed.; Wiley: New York, NY, USA, 1977. [Google Scholar]
- Kaplan, D.R. Fundamental concepts of leaf morphology and morphogenesis: A contribution to the interpretation of molecular genetic mutants. Int. J. Plant. Sci. 2001, 162, 465–474. [Google Scholar] [CrossRef]
- Ogburn, R.M.; Edwards, E.J. Repeated origin of three-dimensional leaf venation releases constraints on the evolution of succulence in plants. Curr. Biol. 2013, 23, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandes-Lopes, J.; Oliveira-Neto, M.A.; Melo-de-Pinna, G.F.A. Different ways to build succulent leaves in Portulacineae (Caryophyllales). Int. J. Plant Sci. 2016, 177, 198–208. [Google Scholar] [CrossRef]
- Kniep, H. Über die Bedeutung des Milchsafts der Pflanzen. Flora Allg. Bot. Ztg. 1905, 94, 129–205. [Google Scholar]
- Trumpke, H. Beiträge zur Anatomie der Sukkulen Euphorbien; Schlesische Volkszeitung: Breslau, Poland, 1914. [Google Scholar]
- Sajeva, M.; Mauseth, J.D. Leaf-like structure in the photosynthetic, succulent stems of cacti. Ann. Bot. 1991, 68, 405–411. [Google Scholar] [CrossRef]
- Mauseth, J.D. Blossfeldia lacks cortical bundles and persistent epidermis; is it basal within Cactoideae? Bradleya 2006, 24, 73–82. [Google Scholar] [CrossRef]
- Terrazas, T.; Escamilla-Molina, R.; Vázquez-Sánchez, M. Variation in the tracheary elements in species of Coryphantha (Cacteae-Cactoideae) with contrasting morphology: The bottleneck model. Braz. J. Bot. 2016, 39, 669–678. [Google Scholar] [CrossRef]
- Schwager, H.; Neinhuis, C.; Mauseth, J.D. Secondary growth of the leaf and bud traces in Hylocereus undatus (Cactaceae) during the formation of branches or flowers. Int. J. Plant. Sci. 2015, 176, 762–769. [Google Scholar] [CrossRef]
- Mauseth, J. Many cacti have clusters of terminal tracheids on their cortical bundles. Haseltonia 2018, 24, 64–74. [Google Scholar] [CrossRef]
- Mauseth, J.D.; Sajeva, M. Cortical bundles in the persistent, photosynthetic stems of Cacti. Ann. Bot. 1992, 70, 317–324. [Google Scholar] [CrossRef]
- Peirson, J.; Bruyns, P.; Riina, R.; Morawetz, J.; Berry, P. A molecular phylogeny and classification of the largely succulent and mainly African Euphorbia subg. Athymalus (Euphorbiaceae). Taxon 2013, 62, 1178–1199. [Google Scholar] [CrossRef]
- Boke, N.H. Developmental morphology and anatomy in Cactaceae. BioScience 1980, 30, 605–610. [Google Scholar] [CrossRef]
- Nobel, P.S. Water relations and photosynthesis of a barrel cactus, Ferocactus acanthodes, in the Colorado desert. Oecologia 1977, 27, 117–133. [Google Scholar] [CrossRef] [PubMed]
- Nobel, P.S. Interception of photosynthetically active radiation by cacti of different morphology. Oecologia 1980, 45, 160–166. [Google Scholar] [CrossRef]
- Darling, M.S. 1989. Epidermis and hypodermis of the saguaro cactus (Cereus giganteus): Anatomy and spectral properties. Am. J. Bot. 1989, 76, 1698–1706. [Google Scholar] [CrossRef]
- Boke, N.H. Histogenesis of the vegetative shoot in Echinocereus. Am. J. Bot. 1951, 38, 23–38. [Google Scholar] [CrossRef]
- Barcikowski, W.; Nobel, P.S. Water relations of cacti during desiccation: Distribution of water in tissues. Bot. Gaz. 1984, 145, 110–115. [Google Scholar] [CrossRef]
- Mauseth, J.D.; Ross, R.G. Systematic anatomy of the primitive cereoid cactus Leptocereus quadricostatus. Bradleya 1988, 6, 49–64. [Google Scholar] [CrossRef]
- Mauseth, J.D. Comparative structure-function studies within a strongly dimorphic plant, Melocactus intortus (Cactaceae). Bradleya 1989, 1, 1–12. [Google Scholar] [CrossRef]
- Mauseth, J.D. Morphogenesis in a highly reduced plant; the endophyte of Tristerix aphyllus (Loranthaceae). Bot. Gaz. 1990, 151, 348–353. [Google Scholar] [CrossRef]
- Mitra, G.C.; Majumdar, G.P. The leaf-base and the internode: Their true morphology. Palaeobotanist 1952, 1, 351–367. [Google Scholar]
- Kadereit, G.; Mucina, I.; Freitag, H. Phylogeny of Salicornioideae (Chenopodiaceae): Diversification, biogeography, and evolutionary trends in leaf and flower morphology. Taxon 2006, 55, 617–642. [Google Scholar] [CrossRef]
- Klak, C.; Reeves, G.; Hedderson, T.A.J. Unmatched tempo of evolution in southern African semi-desert ice plants. Nature 2004, 427, 63–65. [Google Scholar] [CrossRef]
- Valente, L.M.; Britton, A.W.; Powell, M.P.; Papadopulos, A.S.T.; Burgoyne, P.M.; Savolainen, V. Correlates of hyperdiversity in southern African ice plants. Bot. J. Linn. Soc. 2014, 174, 110–129. [Google Scholar] [CrossRef]
- Jäger-Zürn, I. Shoot apex and spathella: Two problematical structures of Podostemaceae–Podostemoideae. Plant Syst. Evol. 2005, 253, 209–218. [Google Scholar] [CrossRef]
- Mauseth, J.D. Structure–function relationships in highly modified shoots of Cactaceae. Ann. Bot. 2006, 98, 901–926. [Google Scholar] [CrossRef] [Green Version]
- Howard, R. The stem-node-leaf continuum of the dicotyledoneae. J. Arnold Arbor. 1974, 55, 125–181. [Google Scholar] [CrossRef]
- Gibson, A.C.; Horak, K.E. Systematic anatomy and phylogeny of Mexican columnar cacti. Ann. Mo. Bot. Gard. 1978, 65, 999–1057. [Google Scholar] [CrossRef]
- Majumdar, G.P. Stipules, stipels, ligules and leaf-sheath. Proc. Indian Acad. Sci. 1956, 43, 9–22. [Google Scholar] [CrossRef]
- Kellogg, E.A. Are Macroevolution and Microevolution Qualitatively Different? Evidence from Poaceae and Other Families. In Developmental Genetics and Plant Evolution; Cronk, Q.C.B., Bateman, R.M., Hawkins, J.A., Eds.; Taylor & Francis: London, UK, 2002; pp. 70–84. [Google Scholar]
- Baum, D.A.; Donoghue, M.J. Transference of Function, Heterotopy and the Evolution of Plant Development. In Developmental Genetics and Plant Evolution; Cronk, Q.C.B., Bateman, R.M., Hawkins, J.A., Eds.; Taylor & Francis: London, UK, 2002; pp. 52–69. [Google Scholar]
- Aloni, R. The Induction of Vascular Tissues by Auxin. In Plant Hormones; Davies, P.J., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 485–506. [Google Scholar]
- Aloni, R. Ecophysiological implications of vascular differentiation and plant evolution. Trees 2015, 29, 1–16. [Google Scholar]
- Johansen, D.A. Plant Microtechnique; McGraw-Hill: New York, NY, USA, 1940. [Google Scholar]
- Kraus, J.E.; Arduin, M. Manual Básico de Métodos em Morfologia Vegetal; Editora da Universidade Federal Rural do Rio de Janeiro: Seropédica, Brazil, 1997. [Google Scholar]
- Gerlach, D. Botanische Mikrotechnik: Eine Einführung, 3rd ed.; Georg Thieme: Stuttgart, Germany, 1984. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arévalo-Rodrigues, G.; Hurbath, F.; Prado, E.; Galvão, I.; Cordeiro, I.; Demarco, D. Adnate Leaf-Base and the Origin of Ribs in Succulent Stems of Euphorbia L. Plants 2022, 11, 1076. https://doi.org/10.3390/plants11081076
Arévalo-Rodrigues G, Hurbath F, Prado E, Galvão I, Cordeiro I, Demarco D. Adnate Leaf-Base and the Origin of Ribs in Succulent Stems of Euphorbia L. Plants. 2022; 11(8):1076. https://doi.org/10.3390/plants11081076
Chicago/Turabian StyleArévalo-Rodrigues, Gustavo, Fernanda Hurbath, Erika Prado, Isabella Galvão, Inês Cordeiro, and Diego Demarco. 2022. "Adnate Leaf-Base and the Origin of Ribs in Succulent Stems of Euphorbia L." Plants 11, no. 8: 1076. https://doi.org/10.3390/plants11081076