
Striga hermonthica Suicidal Germination Activity of Potent Strigolactone Analogs: Evaluation from Laboratory Bioassays to Field Trials

 ,
,  ,
, 
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Striga Seed Germination Bioassays in Response to Various Strigolactone Analogs
2.2. Striga Emergence on Strigolactone Analogs’ Treatment under Greenhouse Conditions
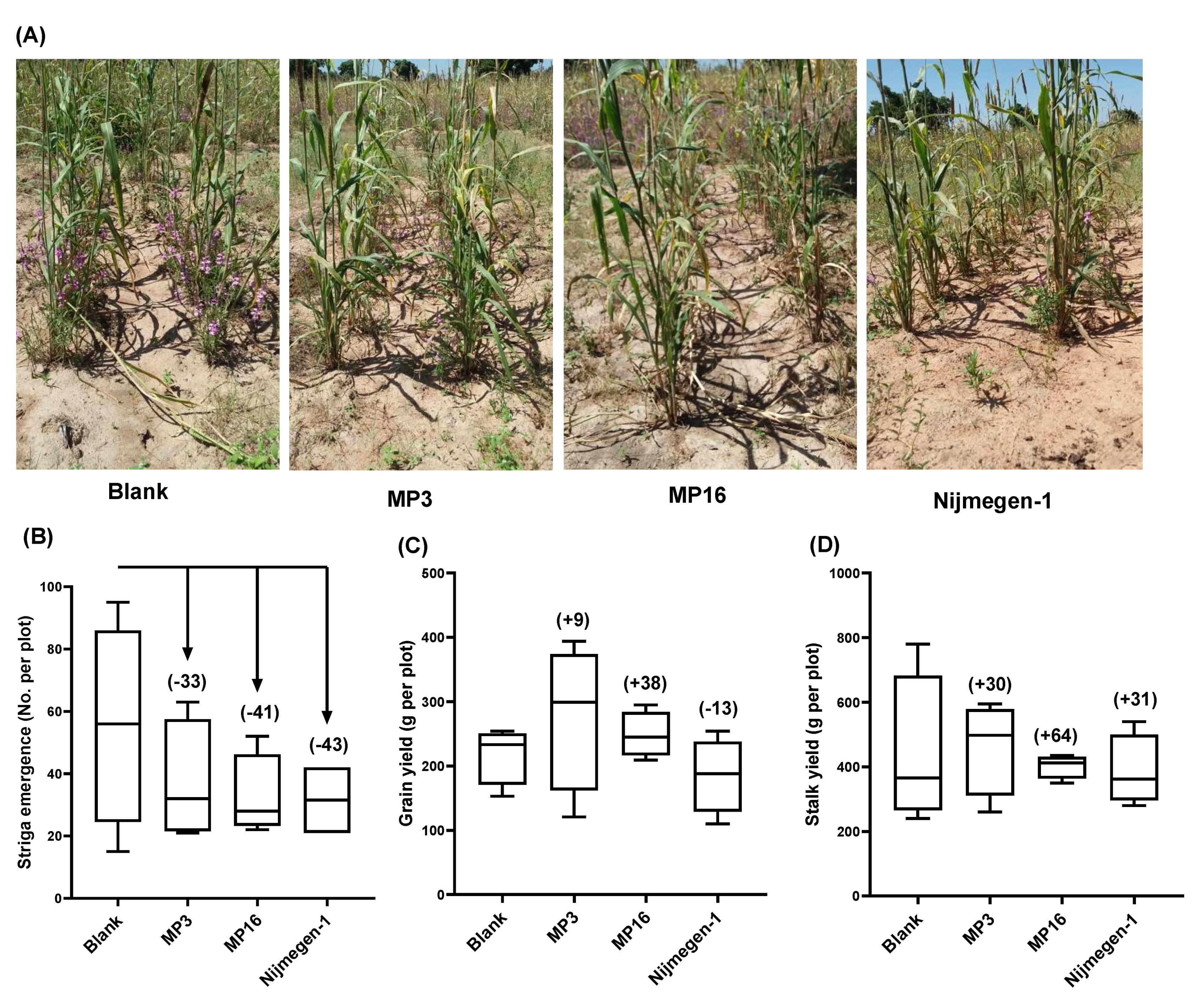
2.3. Effect of Strigolactone Analogs on Striga Infection under Field Conditions
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Chemicals
4.2. Striga Seed Germination Bioassays
4.3. Striga Emergence under Greenhouse Conditions
4.4. Striga Emergence under Field Conditions
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ejeta, G. Breeding for Striga Resistance in Sorghum: Exploitation of an Intricate Host-Parasite Biology. Crop Sci. 2007, 47, 216–227. [Google Scholar] [CrossRef]
- Khan, Z.R.; Midega, C.A.O.; Pittchar, J.O.; Murage, A.W.; Birkett, M.A.; Bruce, T.J.A.; Pickett, J.A. Achieving Food Security for one Million Sub-Saharan African Poor Through Push-Pull Innovation by 2020. Philos. Trans. R. Soc. B-Biol. Sci. 2014, 369, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, D.; Scholes, J.D.; Randrianjafizanaka, M.T.; Randriamampianina, J.A.; Autfray, P.; Freckleton, R.P. Identifying Existing Management Practices in the Control of Striga asiatica within Rice–Maize Systems in Mid-West Madagascar. Ecol. Evol. 2021, 11, 13579–13592. [Google Scholar] [CrossRef] [PubMed]
- Rodenburg, J.; Riches, C.R.; Kayeke, J.M. Addressing Current and Future Problems of Parasitic Weeds in Rice. Crop Prot. 2010, 29, 210–221. [Google Scholar] [CrossRef]
- Gressel, J.; Hanafi, A.; Head, G.; Marasas, W.; Obilana, A.B.; Ochanda, J.; Souissi, T.; Tzotzos, G. Major Heretofore Intractable Biotic Constraints to African Food Security that may be Amenable to Novel Biotechnological Solutions. Crop Prot. 2004, 23, 661–689. [Google Scholar] [CrossRef] [Green Version]
- Ejeta, G. The Striga Scourge in Africa: A Growing Pandemic. In Integrating New Technologies for Striga Control: Towards Ending the Witch-Hunt, 2nd ed.; Ejeta, G., Gressel, J., Eds.; World Scientific: Singapore, 2007; pp. 3–16. [Google Scholar]
- Scholes, J.D.; Press, M.C. Striga Infestation of Cereal Crops–an Unsolved Problem in Resource Limited Agriculture. Curr. Opin. Plant Biol. 2008, 11, 180–186. [Google Scholar] [CrossRef]
- Parker, C. Parasitic Weeds: A World Challenge. Weed Sci. 2012, 60, 269–276. [Google Scholar] [CrossRef]
- Rich, P.J.; Grenier, U.; Ejeta, G. Striga Resistance in the Wild Relatives of Sorghum. Crop Sci. 2004, 44, 2221–2229. [Google Scholar] [CrossRef]
- Pennisi, E. Armed and Dangerous. Science 2010, 327, 804–805. [Google Scholar]
- Rodenburg, J.; Demont, M.; Zwart, S.J.; Bastiaans, L. Parasitic Weed Incidence and Related Economic Losses in Rice in Africa. Agri. Econ. Environ. 2016, 235, 306–317. [Google Scholar] [CrossRef] [Green Version]
- Rodenburg, J.; Bastiaans, L.; Weltzien, E.; Hess, D.E. How can Field Selection for Striga Resistance and Tolerance in Sorghum be Improved? Field Crop. Res. 2005, 93, 34–50. [Google Scholar] [CrossRef] [Green Version]
- Westwood, J.H.; Depamphilis, C.W.; Das, M.; Fernández-Aparicio, M.; Honaas, L.A.; Timko, M.P.; Wafula, E.K.; Wickett, N.J.; Yoder, J.I. The Parasitic Plant Genome Project: New Tools for Understanding the Biology of Orobanche and Striga. Weed Sci. 2012, 60, 295–306. [Google Scholar] [CrossRef]
- Pennisi, E. How Crop-Killing Witchweed Senses its Victims. Science 2015, 350, 146–147. [Google Scholar] [CrossRef] [PubMed]
- Jamil, M.; Kountche, B.A.; Al-Babili, S. Current Progress in Striga Management. Plant Physiol. 2021, 185, 1339–1352. [Google Scholar] [CrossRef] [PubMed]
- Bebawi, F.F.; Eplee, R.E.; Harris, C.E.; Norris, R.S. Longevity of Witchweed (Striga asiatica) Seed. Weed Sci. 1984, 32, 494–497. [Google Scholar] [CrossRef]
- Gbèhounou, G.; Pieterse, A.H.; Verkleij, J.A. Longevity of Striga Seeds Reconsidered: Results of a Field Study on Purple Witchweed (Striga hermonthica) in Benin. Weed Sci. 2003, 51, 940–946. [Google Scholar] [CrossRef]
- Berner, D.; Kling, J.; Singh, B. Striga Research and Control. A Perspective from Africa. Plant Dis. 1995, 79, 652–660. [Google Scholar] [CrossRef]
- Joel, D.M. The Long-term Approach to Parasitic Weeds Control: Manipulation of Specific Developmental Mechanisms of the Parasite. Crop Prot. 2000, 19, 753–775. [Google Scholar] [CrossRef]
- Kim, S.; Adetimirin, V.; The, C.; Dossou, R. Yield Losses in Maize due to Striga hermonthica in West and Central Africa. Int. J. Pest Manag. 2002, 48, 211–217. [Google Scholar] [CrossRef]
- Gbèhounou, G.; Adango, E. Trap Crops of Striga hermonthica: In vitro Identification and Effectiveness in situ. Crop Prot. 2003, 22, 395–404. [Google Scholar] [CrossRef]
- Yoneyama, K.; Awad, A.A.; Xie, X.; Yoneyama, K.; Takeuchi, Y. Strigolactones as Germination Stimulants for Root Parasitic Plants. Plant Cell Physiol. 2010, 51, 1095–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Babili, S.; Bouwmeester, H.J. Strigolactones, a Novel Carotenoid-Derived Plant Hormone. Ann. Rev. Plant Biol. 2015, 66, 161–186. [Google Scholar] [CrossRef] [PubMed]
- Jamil, M.; Kountche, B.A.; Haider, I.; Guo, X.J.; Ntui, V.O.; Jia, K.P.; Ali, S.; Hameed, U.S.; Nakamura, H.; Lyu, Y.; et al. Methyl Phenlactonoates are Efficient Strigolactone Analogs with Simple Structure. J. Exp. Bot. 2018, 69, 2319–2331. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.N.; Yoneyama, K.; Yoneyama, K. The Strigolactone Story. Ann. Rev. Phytopath. 2010, 48, 93–117. [Google Scholar] [CrossRef] [Green Version]
- Felemban, A.; Braguy, J.; Zurbriggen, M.D.; Al-Babili, S. Apocarotenoids Involved in Plant Development and Stress Response. Front. Plant Sci. 2019, 10, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.Y.; Lin, P.-Y.; Al-Babili, S. On the Biosynthesis and Evolution of Apocarotenoid Plant Growth Regulators. Sem. Cell Dev. Biol. 2021, 109, 3–11. [Google Scholar] [CrossRef]
- Fiorilli, V.; Wang, J.Y.; Bonfante, P.; Lanfranco, L.; Al-Babili, S. Apocarotenoids: Old and New Mediators of the Arbuscular Mycorrhizal Symbiosis. Front. Plant Sci. 2019, 10, 1186. [Google Scholar] [CrossRef]
- Conn, C.E.; Nelson, D.C. Evidence that KARRIKIN-INSENSITIVE2 (KAI2) Receptors may Perceive an Unknown Signal that is not Karrikin or Strigolactone. Front. Plant Sci. 2016, 6, 1219–1225. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Yoshimura, M.; Sato, Y.; Kuwata, K.; Toh, S.; Holbrook-Smith, D.; Zhang, H.; Mccourt, P.; Itami, K. Probing strigolactone receptors in Striga hermonthica with fluorescence. Science 2015, 349, 864–868. [Google Scholar] [CrossRef] [Green Version]
- Zarban, R.A.; Hameed, U.F.S.; Jamil, M.; Ota, T.; Wang, J.Y.; Arold, S.; Asami, T.; Al-Babili, S. Rational Design of Striga hermonthica-Specific Seed Germination Inhibitors. Plant Physiol. 2022, 188, 1369–1384. [Google Scholar] [CrossRef]
- Yao, R.; Wang, F.; Ming, Z.; Du, X.; Chen, L.; Wang, Y.; Zhang, W.; Deng, H.; Xie, D. ShHTL7 is a Non-canonical Receptor for Strigolactones in Root Parasitic Weeds. Cell Res. 2017, 27, 838–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, H.; Ye, W.; Hong, L.; Huang, H.; Wang, Z.; Deng, X.; Yang, Q.; Xu, Z. Progress in Parasitic Plant Biology: Host Selection and Nutrient Transfer. Plant Biol. 2006, 8, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Westwood, J.H. The Physiology of the Established Parasite–Host Association. In Parasitic Orobanchaceae; Springer: Berlin/Heidelberg, Germany, 2013; pp. 87–114. [Google Scholar]
- Rubiales, D.; Fernández-Aparicio, M.; Vurro, M.; Eizenberg, H. Advances in Parasitic Weed Research. Front. Plant Sci. 2018, 9, 236–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwanenburg, B.; Pospíšil, T. Structure and Activity of Strigolactones: New Plant Hormones with a Rich Future. Mol. Plant 2013, 6, 38–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamil, M.; Kountche, B.A.; Wang, J.Y.; Haider, I.; Jia, K.-P.; Takahashi, I.; Ota, T.; Asami, T.; Al-Babili, S. A New Series of Carlactonoic Acid Based Strigolactone Analogs for Fundamental and Applied Research. Front. Plant Sci. 2020, 11, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Zwanenburg, B.; Pospisil, T.; Zeljkovic, S.C. Strigolactones: New Plant Hormones in Action. Planta 2016, 243, 1311–1326. [Google Scholar] [CrossRef] [Green Version]
- Kountche, B.A.; Jamil, M.; Yonli, D.; Nikiema, M.P.; Blanco-Ania, D.; Asami, T.; Zwanenburg, B.; Al-Babili, S. Suicidal Germination as a Control Strategy for Striga hermonthica (Benth.) in Smallholder Farms of Sub-Saharan Africa. Plants People Planet 2019, 1, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Eplee, R.E. Ethylene: A Witchweed Seed Germination Stimulant. Weed Sci. 1975, 23, 433–436. [Google Scholar] [CrossRef]
- Samejima, H.; Babiker, A.G.; Takikawa, H.; Sasaki, M.; Sugimoto, Y. Practicality of the Suicidal Germination Approach for Controlling Striga hermonthica. Pest Manag. Sci. 2016, 72, 2035–2042. [Google Scholar] [CrossRef]
- Zwanenburg, B.; Blanco-Ania, D. Strigolactones: New Plant Hormones in the Spotlight. J. Exp. Bot. 2018, 69, 2205–2218. [Google Scholar] [CrossRef]
- Nefkens, G.H.; Thuring, J.W.J.; Beenakkers, M.F.; Zwanenburg, B. Synthesis of a Phthaloylglycine-Derived Strigol Analogue and its Germination Stimulatory Activity Toward Seeds of the Parasitic Weeds Striga hermonthica and Orobanche crenata. J. Agric. Food Chem. 1997, 45, 2273–2277. [Google Scholar] [CrossRef] [Green Version]
- Mwakaboko, A.S.; Zwanenburg, B. Single Step Synthesis of Strigolactone Analogues from Cyclic Keto Enols, Germination Stimulants for Seeds of Parasitic Weeds. Bioorg. Med. Chem. 2011, 19, 5006–5011. [Google Scholar] [CrossRef] [PubMed]
- Jamil, M.; Kountche, B.A.; Haider, I.; Wang, J.Y.; Aldossary, F.; Zarban, R.A.; Jia, K.P.; Yonli, D.; Hameed, U.F.S.; Takahashi, I.; et al. Methylation at the C-3′ in D-Ring of Strigolactone Analogs Reduces Biological Activity in Root Parasitic Plants and Rice. Front. Plant Sci. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliche, E.B.; Screpanti, C.; De Mesmaeker, A.; Munnik, T.; Bouwmeester, H.J. Science and Application of Strigolactones. New Phytol. 2020, 227, 1001–1011. [Google Scholar] [CrossRef] [Green Version]
- Wigchert, S.; Kuiper, E.; Boelhouwer, G.; Nefkens, G.; Verkleij, J.; Zwanenburg, B. Dose—Response of Seeds of the Parasitic Weeds Striga and Orobanche Toward the Synthetic Germination Stimulants GR24 and Nijmegen 1. J. Agric. Food Chem. 1999, 47, 1705–1710. [Google Scholar] [CrossRef]
- Jamil, M.; Wang, J.Y.; Yonli, D.; Patil, R.H.; Riyazaddin, M.; Gangashetty, P.; Berqdar, L.; Chen, G.-T.E.; Traore, H.; Margueritte, O.; et al. A New Formulation for Strigolactone Suicidal Germination Agents, Towards Successful Striga Management. Plants 2022, 11, 808. [Google Scholar] [CrossRef]
- Mangnus, E.M.; Dommerholt, F.J.; De Jong, R.L.; Zwanenburg, B. Improved Synthesis of Strigol Analog GR24 and Evaluation of the Biological Activity of its Diastereomers. J. Agric. Food Chem. 1992, 40, 1230–1235. [Google Scholar] [CrossRef]
- Jamil, M.; Charnikhova, T.; Houshyani, B.; van Ast, A.; Bouwmeester, H.J. Genetic Variation in Strigolactone Production and Tillering in Rice and its Effect on Striga hermonthica Infection. Planta 2012, 235, 473–484. [Google Scholar] [CrossRef] [Green Version]
- Jamil, M.; Kanampiu, F.K.; Karaya, H.; Charnikhova, T.; Bouwmeester, H.J. Striga hermonthica Parasitism in Maize in Response to N and P Fertilisers. Field Crop. Res. 2012, 134, 1–10. [Google Scholar] [CrossRef]
- Braguy, J.; Ramazanova, M.; Giancola, S.; Jamil, M.; Kountche, B.A.; Zarban, R.A.Y.; Felemban, A.; Wang, J.Y.; Lin, P.-Y.; Haider, I.; et al. SeedQuant: A Deep Learning-Based Tool for Assessing Stimulant and Inhibitor Activity on Root Parasitic Seeds. Plant Physiol. 2021, 186, 1632–1644. [Google Scholar] [CrossRef]
- Wang, J.Y.; Jamil, M.; Lin, P.-Y.; Ota, T.; Fiorilli, V.; Novero, M.; Zarban, R.A.; Kountche, B.A.; Takahashi, I.; Martínez, C. Efficient Mimics for Elucidating Zaxinone Biology and Promoting Agricultura Applications. Mol. Plant 2020, 13, 1654–1661. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jamil, M.; Wang, J.Y.; Yonli, D.; Ota, T.; Berqdar, L.; Traore, H.; Margueritte, O.; Zwanenburg, B.; Asami, T.; Al-Babili, S. Striga hermonthica Suicidal Germination Activity of Potent Strigolactone Analogs: Evaluation from Laboratory Bioassays to Field Trials. Plants 2022, 11, 1045. https://doi.org/10.3390/plants11081045
Jamil M, Wang JY, Yonli D, Ota T, Berqdar L, Traore H, Margueritte O, Zwanenburg B, Asami T, Al-Babili S. Striga hermonthica Suicidal Germination Activity of Potent Strigolactone Analogs: Evaluation from Laboratory Bioassays to Field Trials. Plants. 2022; 11(8):1045. https://doi.org/10.3390/plants11081045
Chicago/Turabian StyleJamil, Muhammad, Jian You Wang, Djibril Yonli, Tsuyoshi Ota, Lamis Berqdar, Hamidou Traore, Ouedraogo Margueritte, Binne Zwanenburg, Tadao Asami, and Salim Al-Babili. 2022. "Striga hermonthica Suicidal Germination Activity of Potent Strigolactone Analogs: Evaluation from Laboratory Bioassays to Field Trials" Plants 11, no. 8: 1045. https://doi.org/10.3390/plants11081045