
Silicon Application Promotes Productivity, Silicon Accumulation and Upregulates Silicon Transporter Gene Expression in Rice



Abstract
:1. Introduction
2. Results
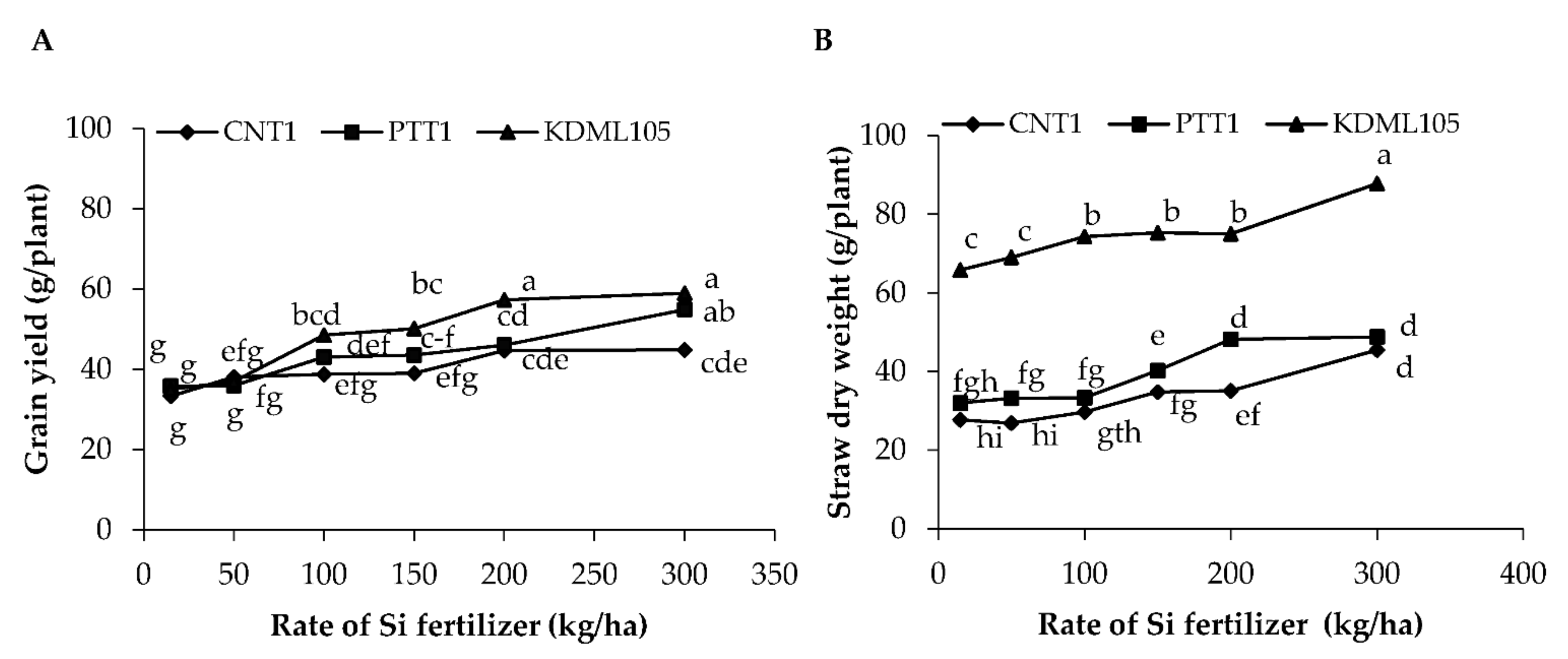
2.1. Grain Yield, Straw Dry Weight, and Yield Components
Yield
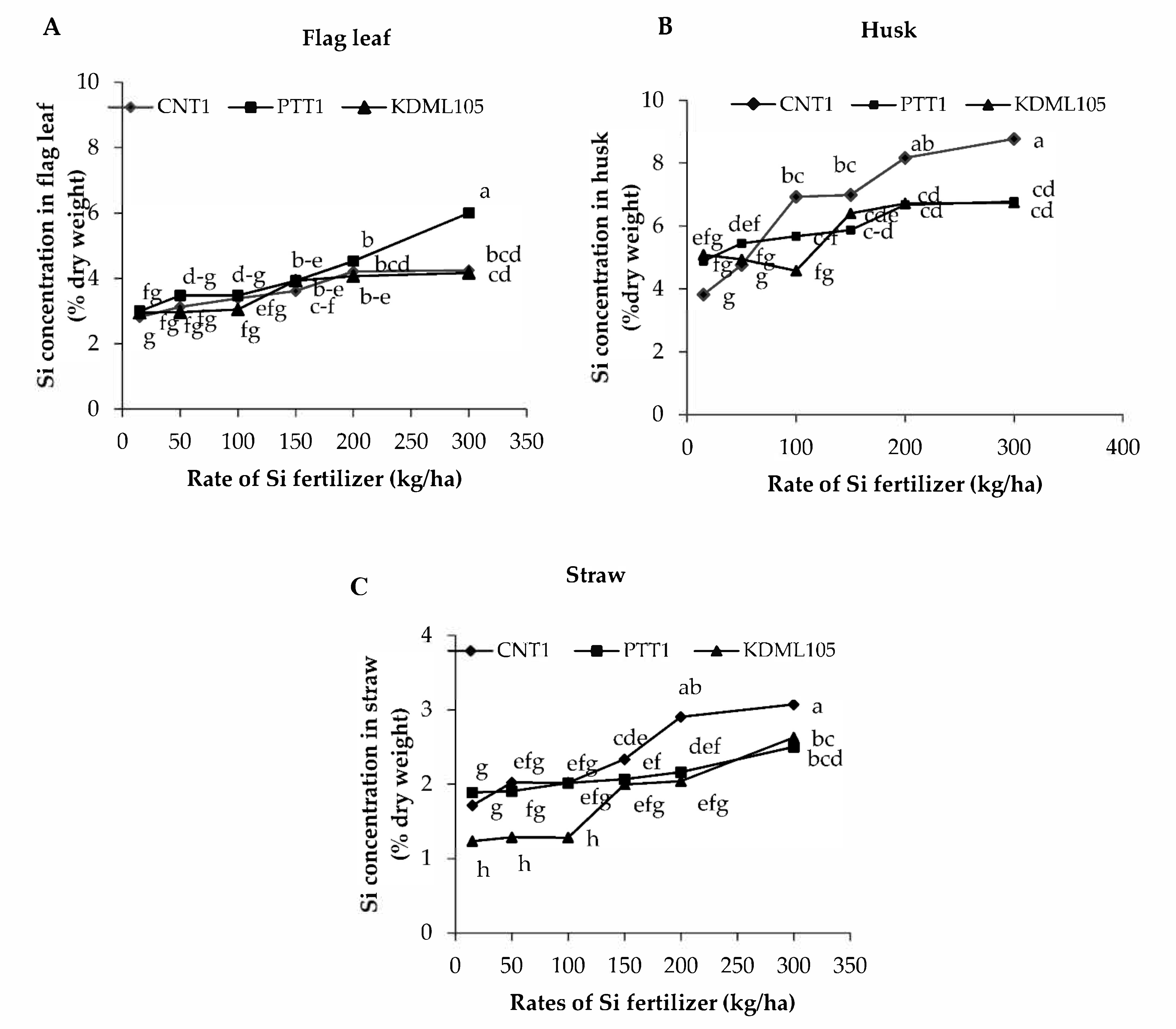
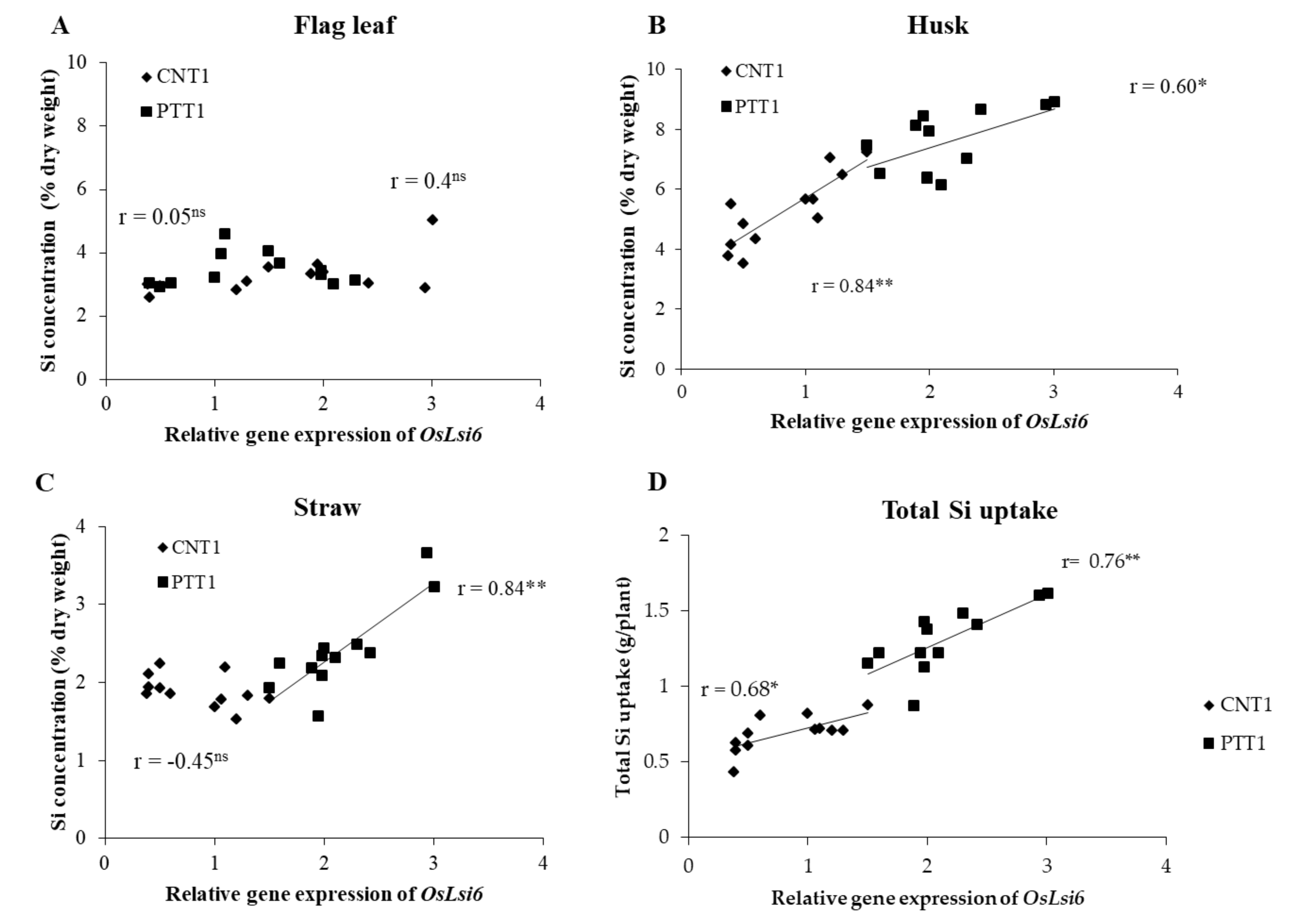
2.2. Silicon Concentration in Different Plant Parts
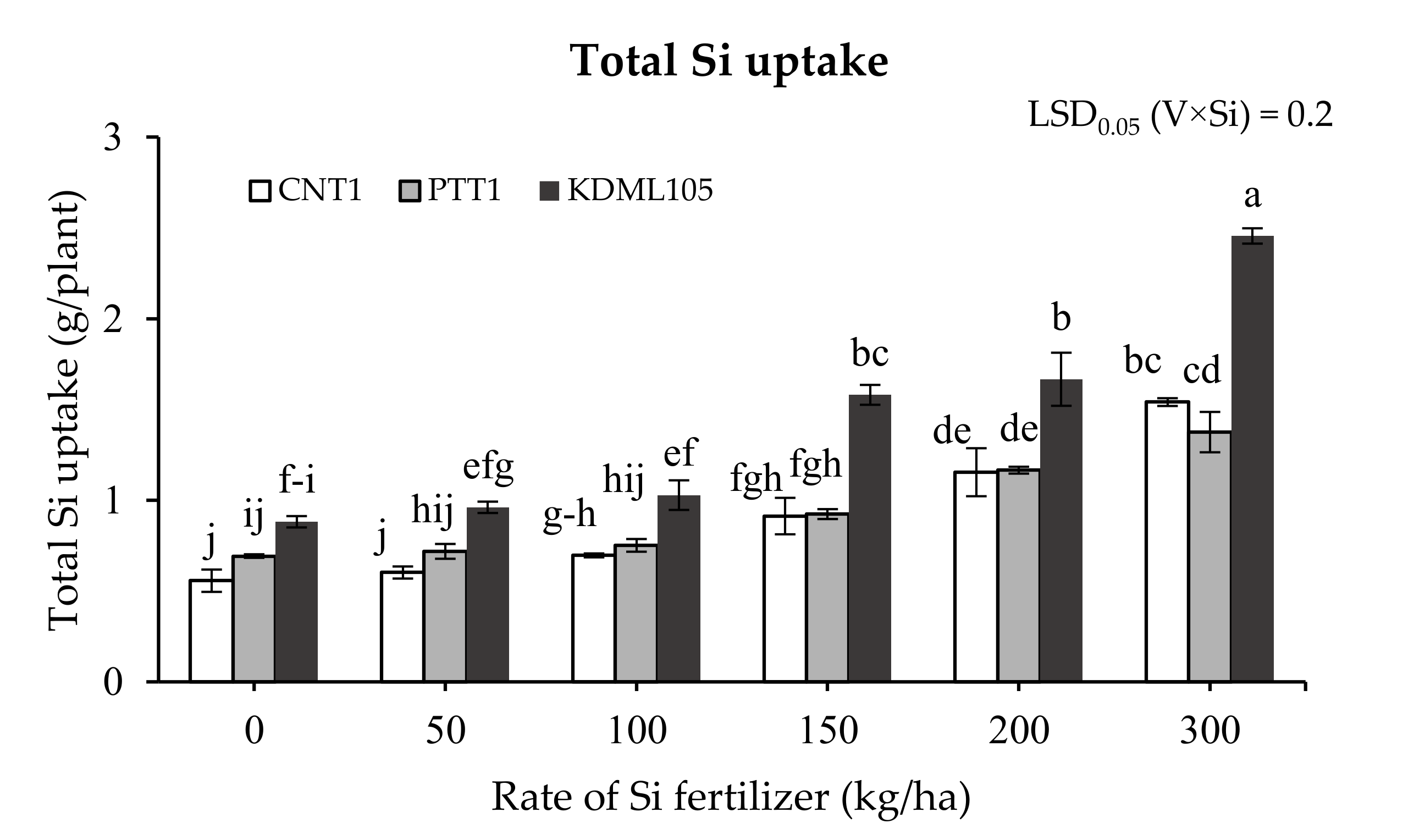
2.3. Total Silicon Uptake
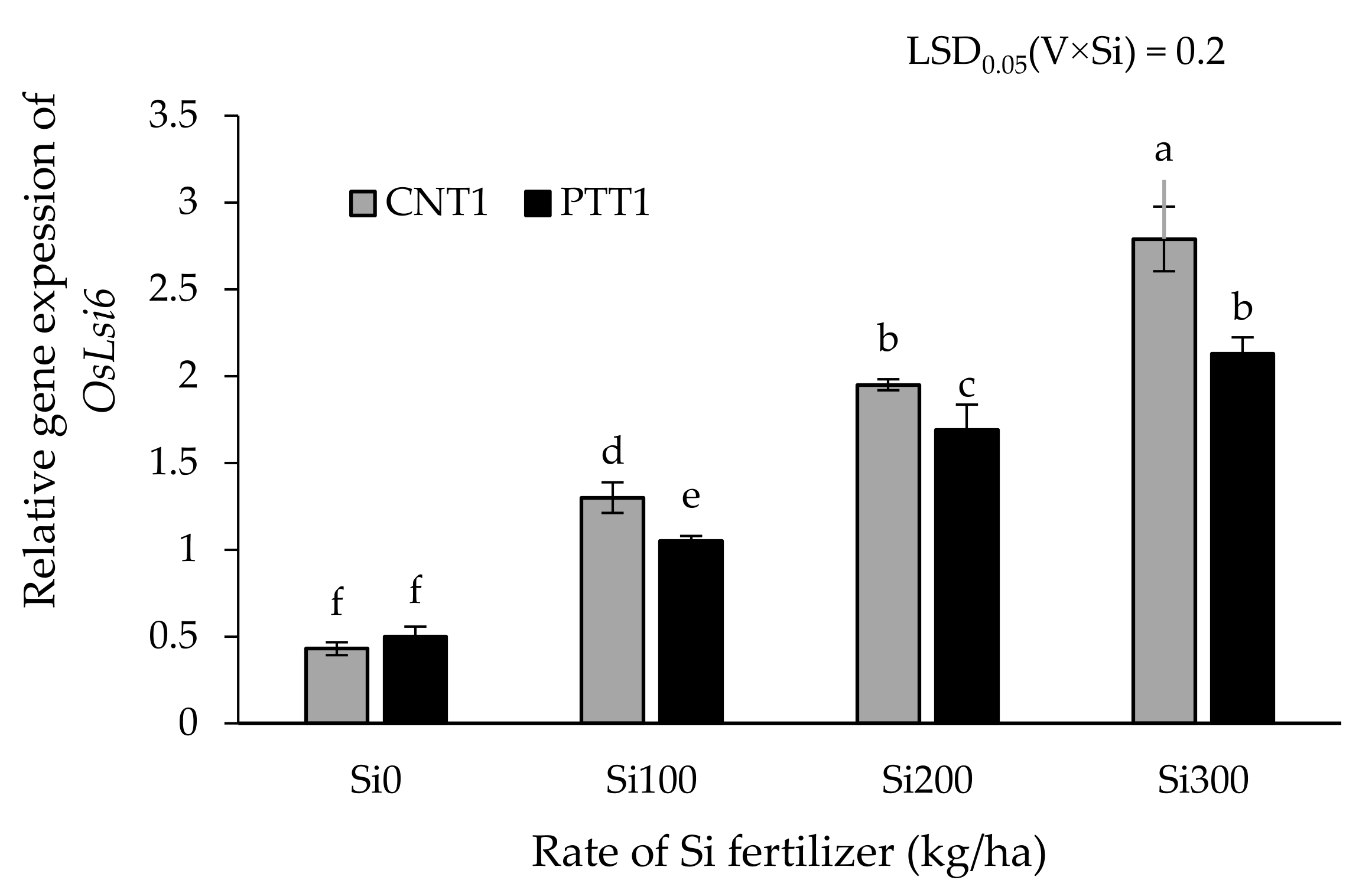
2.4. Expression of OsLsi6 Gene
3. Materials and Methods
3.1. Plant Culture
3.2. Data Collection and Sample Preparation
3.3. Silicon Concentration Analysis
3.4. Gene Expression by Semi-Quantitative RT-PCR Analysis
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Khush:, G.S. What it will take to feed 5.0 billion rice consumers in 2030. Plant Mol. Biol. 2005, 59, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Delvaux, B. Phytolith-rich biochar: A potential Si fertilizer in desilicated soils. Gcb Bioenergy 2019, 11, 1264–1282. [Google Scholar] [CrossRef] [Green Version]
- Haynes, R.J. A contemporary overview of silicon availability in agricultural soils. J. Plant Nutr. Soil Sci. 2014, 177, 831–844. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Rahman, M.M.; Ansary, M.M.U.; Keya, S.S.; Abdelrahman, M.; Miah, M.G.; Phan Tran, L.S. Silicon in mitigation of abiotic stress-induced oxidative damage in plants. Crit. Rev. Biotechnol. 2021, 41, 918–934. [Google Scholar] [CrossRef]
- Etesami, H.; Jeong, B.R. Silicon (Si): Review and future prospects on the action mechanisms in alleviating biotic and abiotic stresses in plants. Ecotoxicol. Environ. Saf. 2018, 147, 881–896. [Google Scholar] [CrossRef]
- Singh, A.K.; Singh, R.; Singh, K. Growth, yield, and economics of rice (Oryza sativa) as influenced by level and time of silicon application. Ind. J. Agron. 2005, 50, 90–193. [Google Scholar]
- Ma, J.F.; Nishimura, K.; Takahashi, E. Effect of silicon on the growth of rice plant at different growth stages. Soil Sci. Plant Nutr. 1989, 5, 347–356. [Google Scholar] [CrossRef]
- Detmann, K.C.; Araújo, W.L.; Martins, S.C.; Sanglard, L.M.V.P.; Reis, J.V.; Detmann, E.; Rodrigues, F.Á.; Nunes-Nesi, A.; Fernie, A.R.; DaMatta, F.M. Silicon nutrition increases grain yield, which, in turn, exerts a feed-forward stimulation of photosynthetic rates via enhanced mesophyll conductance and alters primary metabolism in rice. New Phytol. 2012, 196, 752–762. [Google Scholar] [CrossRef] [Green Version]
- Tamai, K.; Ma, J.F. Reexamination of silicon effects on rice growth and production under field conditions using a low silicon mutant. Plant Soil 2008, 307, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Berahim, Z.; Omar, H.M.; Zakaria, N.; Ismail, M.R.; Rosle, R.; Roslin, N.A.; Che’ya, N.N. Silicon improves yield performance by enhancement in physiological responses, crop imagery, and leaf and culm sheath morphology in new rice line, PadiU Putra. BioMed Res. Int. 2021, 2021, 6679787. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N.; Tamai, K.; Mitani, N. Genotypic difference in silicon uptake and expression of silicon transporter genes in rice. Plant Physiol. 2007, 145, 919–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.F.; Yamaji, N.; Mitani-Ueno, N. Transport of silicon from roots to panicles in plant. Proc. Jpn. Acad. Ser. B 2011, 87, 377–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaji, N.; Sakurai, G.; Mitani-Ueno, N.; Ma, J.F. Orchestration of three transporters and distinct vascular structures in node for intervascular transfer of silicon in rice. Proc. Natl. Acad. Sci USA 2015, 112, 11401–11406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.F.; Tamai, K.; Yamaji, N.; Mitani, N.; Konishi, S.; Katsuhara, M.; Ishiguro, M.; Murata, Y.; Yano, M. A silicon transporter in rice. Nature 2006, 440, 688–691. [Google Scholar] [CrossRef]
- Yamaji, N.; Ma, J.F. The node, a hub for mineral nutrient distribution in graminaceous plants. Trends Plant Sci. 2014, 19, 556–563. [Google Scholar] [CrossRef]
- Yamaji, N.; Mitatni, N.; Ma, J.F. A transporter regulating silicon distribution in rice shoots. Plant Cell. 2008, 20, 1381–1389. [Google Scholar] [CrossRef] [Green Version]
- Chaiwong, N.; Rerkasem, B.; Pusadee, T.; Prom-u-thai, C. Silicon application improves caryopsis development and yield in rice. J. Sci. Food Agric. 2021, 10, 220–228. [Google Scholar] [CrossRef]
- Pati, S.; Pal, B.; Badole, S.; Hazra, G.C.; Mandal, B. Effect of silicon fertilization on growth, yield, and nutrient uptake of rice. Commun. Soil Sci Plant Anal. 2016, 47, 284–290. [Google Scholar] [CrossRef]
- Cuong, T.X.; Ullah, H.; Datta, A.; Hanh, T.C. Effects of silicon-based fertilizer on growth, yield and nutrient uptake of rice in tropical zone of Vietnam. Rice Sci. 2017, 24, 283–290. [Google Scholar] [CrossRef]
- Wu, C.Y.; Chen, D.; Luo, H.W.; Yao, Y.M.; Wang, Z.W.; Matsui, T.; Tian, X.H. Effects of exogenous silicon on the pollination and fertility characteristics of hybrid rice under heat stress during anthesis. Chin. J. Appl. Ecol. 2013, 24, 3113–3122. [Google Scholar]
- International Rice Research Institute (IRRI). Annual Report 1965; IRRI: Los Banos, Laguna, Philippines, 1966. [Google Scholar]
- Malav, J.K.; Ramani, V.P.; Sajid, M.; Patel, K. Yield and nutrient content of rice as influenced by silicon and nitrogen application. Ann. Plant Soil Res. 2016, 18, 413–417. [Google Scholar]
- Bityutskii, N.; Pavlovic, J.; Yakkonen, K.; Maksimović, V.; Nikolic, M. Contrasting effect of silicon on iron, zinc and manganese status and accumulation of metal-mobilizing compounds in micronutrient-deficient cucumber. Plant. Physiol. Biochem. 2014, 74, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.M.; Guo, F.S.; Cornelis, J.; Song, Z.L.; Wang, X.D.; Delvaux, B. Combined silicon-phosphorus fertilization affects the biomass and phytolith stock of rice plants. Front. Plant Sci. 2020, 11, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heckman, J.R. Soil fertility test interpretation: Phosphorus, potassium, magnesium and calcium. In Fact Sheet FS719 Rutgers Cooperative Extension, New Jersey Agricultural Experimental Extension; Rutgers, State University of New Jersey: New Brunswick, NJ, USA, 2006; Volume 4. [Google Scholar]
- Pavlovic, J.; Kostic, L.; Bosnic, P.; Kirkby, E.A.; Nikolic, M. Interactions of silicon with essential and beneficial elements in plants. Front. Plant Sci. 2021, 12, 1224. [Google Scholar] [CrossRef]
- Wu, Q.S.; Wan, X.Y.; Su, N.; Cheng, Z.J.; Wang, J.K.; Lei, C.L.; Zhang, X.; Jiang, L.; Ma, J.F.; Wana, J.M. Genetic dissection of silicon uptake ability in rice (Oryza sativa L.). Plant Sci. 2006, 171, 441–448. [Google Scholar] [CrossRef]
- Kim, Y.H.; Khan, A.L.; Waqas, M.; Lee, I.J. Silicon regulates antioxidant activities of crop plants under abiotic-induced oxidative stress:a review. Front. Plant Sci. 2017, 8, 510. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.B.; Shen, J.N.; Hu, S.S. Effect of silicon on plant traits and yield of cold region rice. North Rice 2014, 44, 12–15. [Google Scholar]
- Zhang, L.; Chen, X.; Chang, B.; Gu, X.; Li, W.; Wang, S. Application effect of silicon fertilizer on rice. Heilongjiang Agric. Sci. 2014, 12, 43–46. [Google Scholar]
- Sheikhani, A.R.; Aminpanah, H.; Firouzi, S. Effects of inoculation method of plant growth-promoting rhizobacteria and silicon rate on rice grain yield. Thai J. Agric. Sci. 2014, 47, 227–234. [Google Scholar]
- Sun, X.; Liu, Q.; Tang, T.; Chen, X.; Luo, X. Silicon fertilizer application promotes phytolith accumulation in rice plants. Front. Plant Sci. 2019, 10, 425. [Google Scholar] [CrossRef]
- Tripathi, P.; Subedi, S.; Khan, A.L.; Chung, Y.S.; Kim, Y. Silicon effects on the root system of diverse crop species using root phenotyping technology. Plants. 2021, 10, 885. [Google Scholar] [CrossRef] [PubMed]
- Patil, A.A.; Durgude, A.G.; Pharande, A.L.; Kadlag, A.D.; Nimbalkar, C.A. Effect of calcium silicate as a silicon source on growth and yield of rice plants. Int. J. Chem. Stud. 2017, 5, 545–549. [Google Scholar]
- Jinger, D.; Devi, M.T.; Dhar, S.; Dass, A.; Rajanna, G.A.; Upadhaya, P.; Raj, R. Silicon in mitigating biotic stresses in rice (Oryza sativa L.)—A review. Ann. Agric. Sci. 2017, 38, 1. [Google Scholar]
- Takahashi, E.; Ma, J.F.; Miyake, Y. The possibility of silicon as an essential element for higher plants. J. Agric. Food Chem. 1990, 2, 99–122. [Google Scholar]
- Deren, C.W. Plant genotype, silicon concentration and silicon-related responses. In Silicon in Agriculture; Datnoff, L.E., Snyder, G.H., Korndorfer, G.H., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2001; Volume 8, pp. 149–169. [Google Scholar]
- Ma, J.F.; Yamaji, N. Silicon uptake and accumulation in higher plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef]
- Crooks, R.; Prentice, P. Extensive investigation into field based responses to a siliccon fertiliser. Silicon 2017, 9, 301–304. [Google Scholar] [CrossRef]
- Yan, G.C.; Nikolic, M.; Ye, M.J.; Xiao, Z.X.; Liang, Y.C. Silicon acquisition and accumulationin plant and its significance for agriculture. J. Interg. Agric. 2018, 17, 2138–2150. [Google Scholar] [CrossRef]
- Bokor, B.; Bokorová, S.; Ondoš, S.; Švubová, R.; Lukačová, Z.; Hýblová, M.; Szemes, T.; Lux, A. Ionome and expression level of Si transporter genes (Lsi1, Lsi2, and Lsi6) affected by Znand Si interaction in maize. Environ. Sci. Pollut. Res. 2015, 22, 6800–6811. [Google Scholar] [CrossRef]
- Lavinsky, A.O.; Detmann, K.C.; Reis, J.V.; Ávila, R.T.; Sanglard, M.L.; Pereira, L.F.; DaMatta, F.M. Silicon improves rice grain yield and photosynthesis specifically when supplied during the reproductive growth stage. J. Plant. Physiol. 2016, 206, 125–132. [Google Scholar] [CrossRef]
- Chaiwong, N.; Lordkaew, S.; Yimyam, N.; Rerkasem, B.; Prom-u-thai, C. Silicon nutrition and distribution in plants of different Thai rice varieties. Int. J. Agric. Biol. 2018, 20, 669–675. [Google Scholar] [CrossRef]
- Dai, W.; Zhang, K.; Duan, B.; Sun, C.; Zheng, K.; Cai, R.; Zhuang, J. Rapid determination of silicon content in rice. Rice Sci. 2005, 12, 145–147. [Google Scholar]
- Wangkaew, B.; Prom-u-thai, C.; Jamjod, S.; Rerkasem, B.; Pusadee, T. Silicon concentration and expression of silicon transport genes in two Thai rice Varieties. CMU J. Nat. Sci. 2019, 18, 358–372. [Google Scholar] [CrossRef]
- Yamaji, N.; Ma, J.F. A transporter at the node responsible for intervascular transfer of silicon in rice. Plant Cell 2009, 21, 2878–2883. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grain Yield | Straw Dry Weight | |
|---|---|---|
| Variety (V) | p < 0.01 | p < 0.01 |
| Si application (Si) | p < 0.01 | p < 0.01 |
| V × Si | p < 0.05 | p < 0.05 |
| LSD0.05 (V × Si) | 6.8 | 5.1 |
| Tiller/Plant | Panicle/Plant | Filled Grain (%) | 1000 Grain Weight (g) | |
|---|---|---|---|---|
| Si application rate (kg/ha) | ||||
| Si0 | 12.44 b | 12.44 c | 90.58 e | 28.78 d |
| Si50 | 12.44 b | 12.59 c | 92.61 d | 30.59 cd |
| Si100 | 15.89 a | 16.10 ab | 93.33 cd | 32.22 cd |
| Si150 | 15.71 a | 15.56 ab | 94.31 bc | 32.90 bc |
| Si200 | 16.89 a | 16.41 ab | 95.49 ab | 35.60 ab |
| Si300 | 17.86 a | 18.00 a | 96.50 a | 36.81 a |
| Rice variety | ||||
| CNT1 | 17.90 a | 17.49 a | 92.68 b | 30.00 b |
| PTT1 | 15.21 b | 15.20 b | 93.11 b | 33.87 a |
| KDML105 | 12.41 c | 12.90 c | 95.60 a | 30.00 b |
| F-test | ||||
| Variety (V) | p < 0.01 | p < 0.01 | p < 0.01 | p < 0.01 |
| LSD0.05 (V) | 1.5 | 1.5 | 11.4 | 2.4 |
| Si application (Si) | p < 0.01 | p < 0.01 | p < 0.01 | p < 0.01 |
| LSD0.05 (Si) | 2.1 | 2.1 | 16.2 | 3.3 |
| V × Si | - | - | - | - |
| Si Concentration (% Dry Weight) | Total Si Uptake (g/Plant) | |||
|---|---|---|---|---|
| Flag Leaf | Husk | Straw | ||
| Variety (V) | p < 0.01 | p < 0.01 | p < 0.01 | p < 0.01 |
| Si application (Si) | p < 0.01 | p < 0.01 | p < 0.01 | p < 0.01 |
| V × Si | p < 0.05 | p < 0.05 | p < 0.05 | p < 0.01 |
| LSD0.05 (V × Si) | 0.7 | 1.35 | 0.4 | 0.2 |
| Si Concentration (% Dry Weight) | |||
|---|---|---|---|
| Flag Leaf | Husk | Straw | |
| Grain yield (g/plant) | 0.66 ** | 0.48 * | 0.42 ns |
| Straw dry weight (g/plant) | 0.91 ** | 0.04 ns | 0.19 ns |
| Filled grain (%) | 0.65 ** | 0.60 ** | 0.08 ns |
| Number of tillers/plant | 0.37 ns | 0.30 ns | 0.44 ns |
| Number of panicles/plant | 0.40 ns | 0.29 ns | 0.41 ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaiwong, N.; Pusadee, T.; Jamjod, S.; Prom-u-thai, C. Silicon Application Promotes Productivity, Silicon Accumulation and Upregulates Silicon Transporter Gene Expression in Rice. Plants 2022, 11, 989. https://doi.org/10.3390/plants11070989
Chaiwong N, Pusadee T, Jamjod S, Prom-u-thai C. Silicon Application Promotes Productivity, Silicon Accumulation and Upregulates Silicon Transporter Gene Expression in Rice. Plants. 2022; 11(7):989. https://doi.org/10.3390/plants11070989
Chicago/Turabian StyleChaiwong, Nanthana, Tonapha Pusadee, Sansanee Jamjod, and Chanakan Prom-u-thai. 2022. "Silicon Application Promotes Productivity, Silicon Accumulation and Upregulates Silicon Transporter Gene Expression in Rice" Plants 11, no. 7: 989. https://doi.org/10.3390/plants11070989