
Impact of Grazing on Diversity of Semi-Arid Rangelands in Crete Island in the Context of Climatic Change


Abstract
:1. Introduction
- (a)
- Do patterns of species diversity indices and composition differ among rangelands exposed to different grazing intensities?
- (b)
- Do these differences vary among rangelands with different altitudes and climatic conditions?
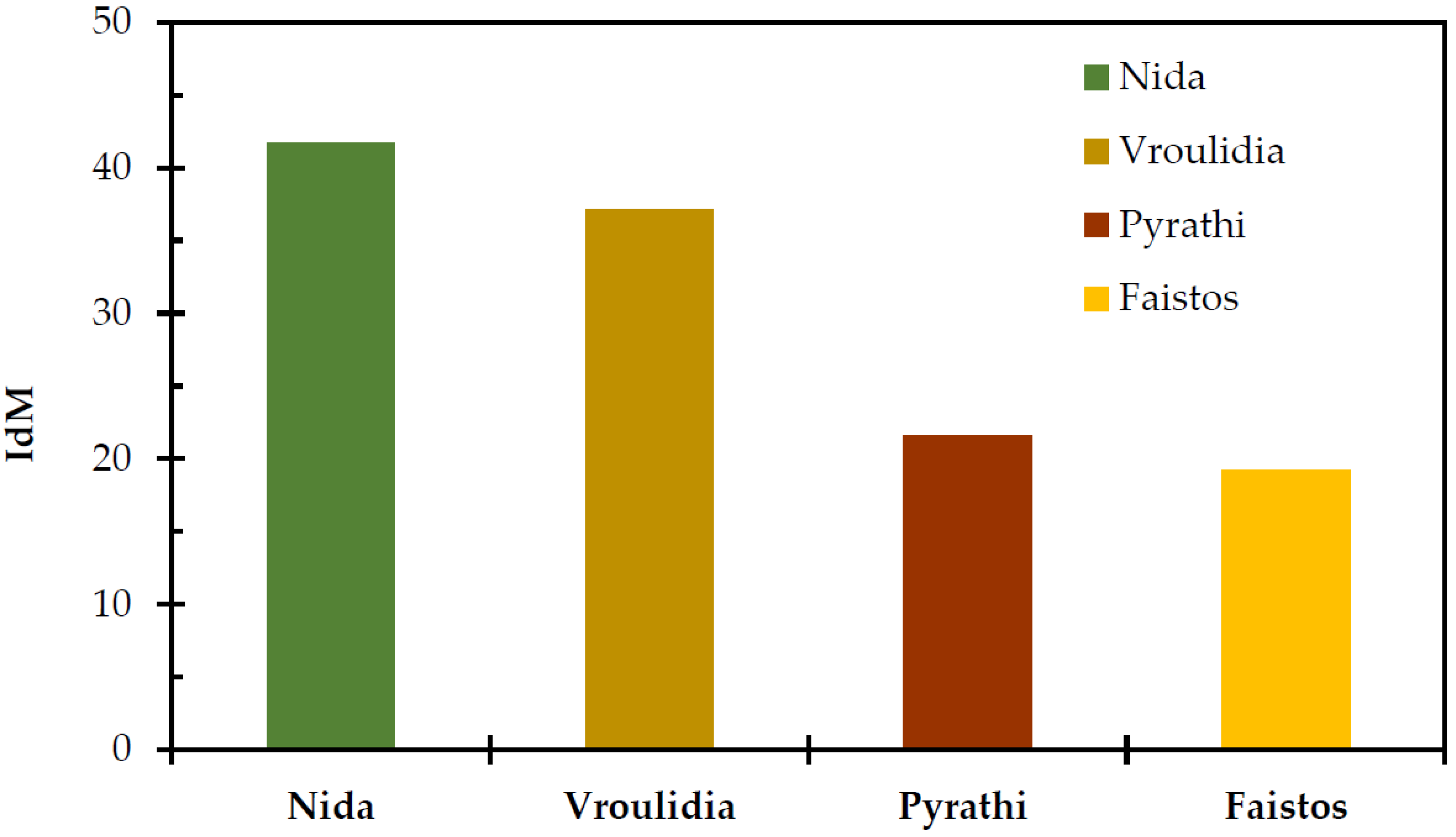
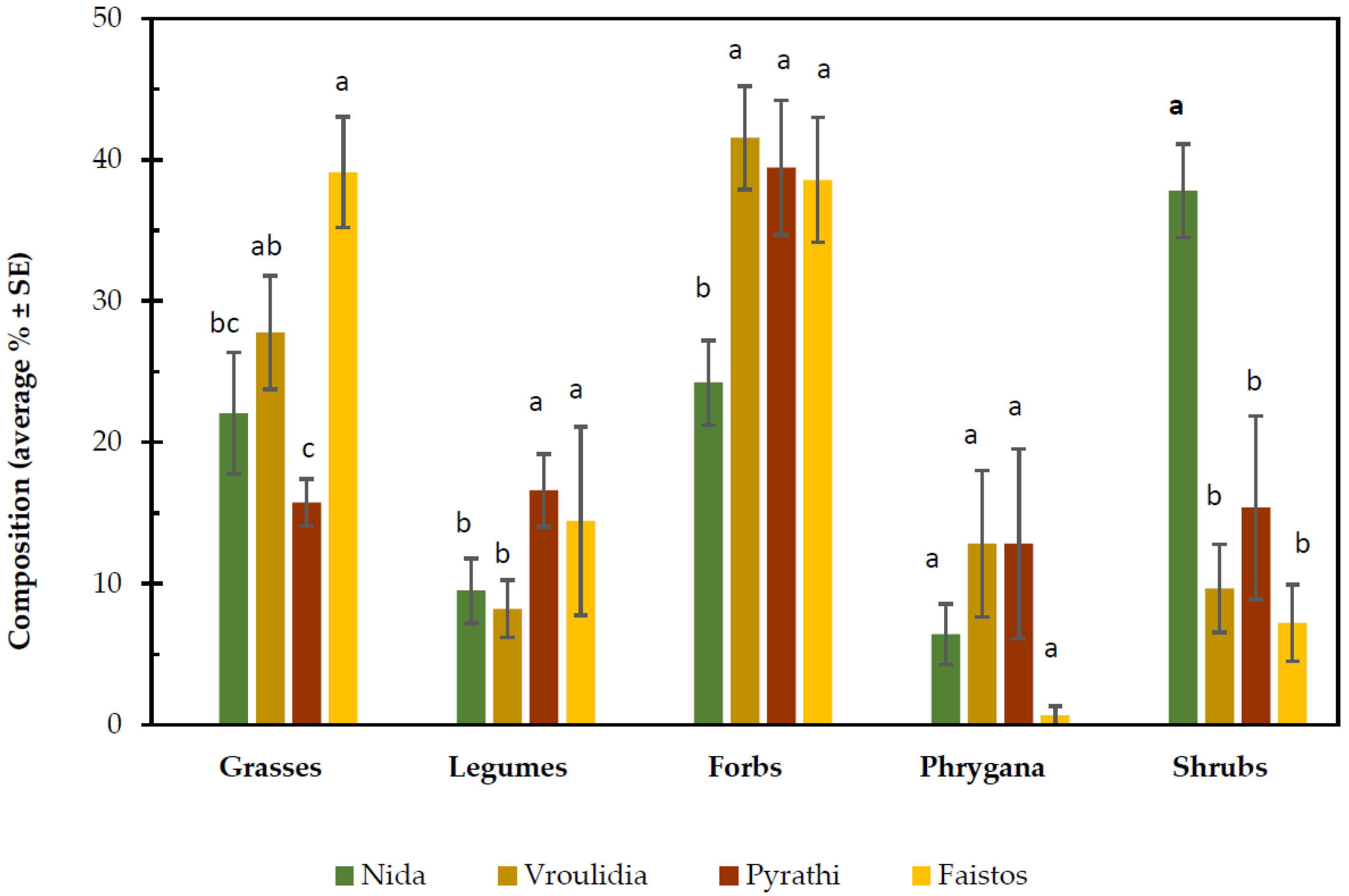
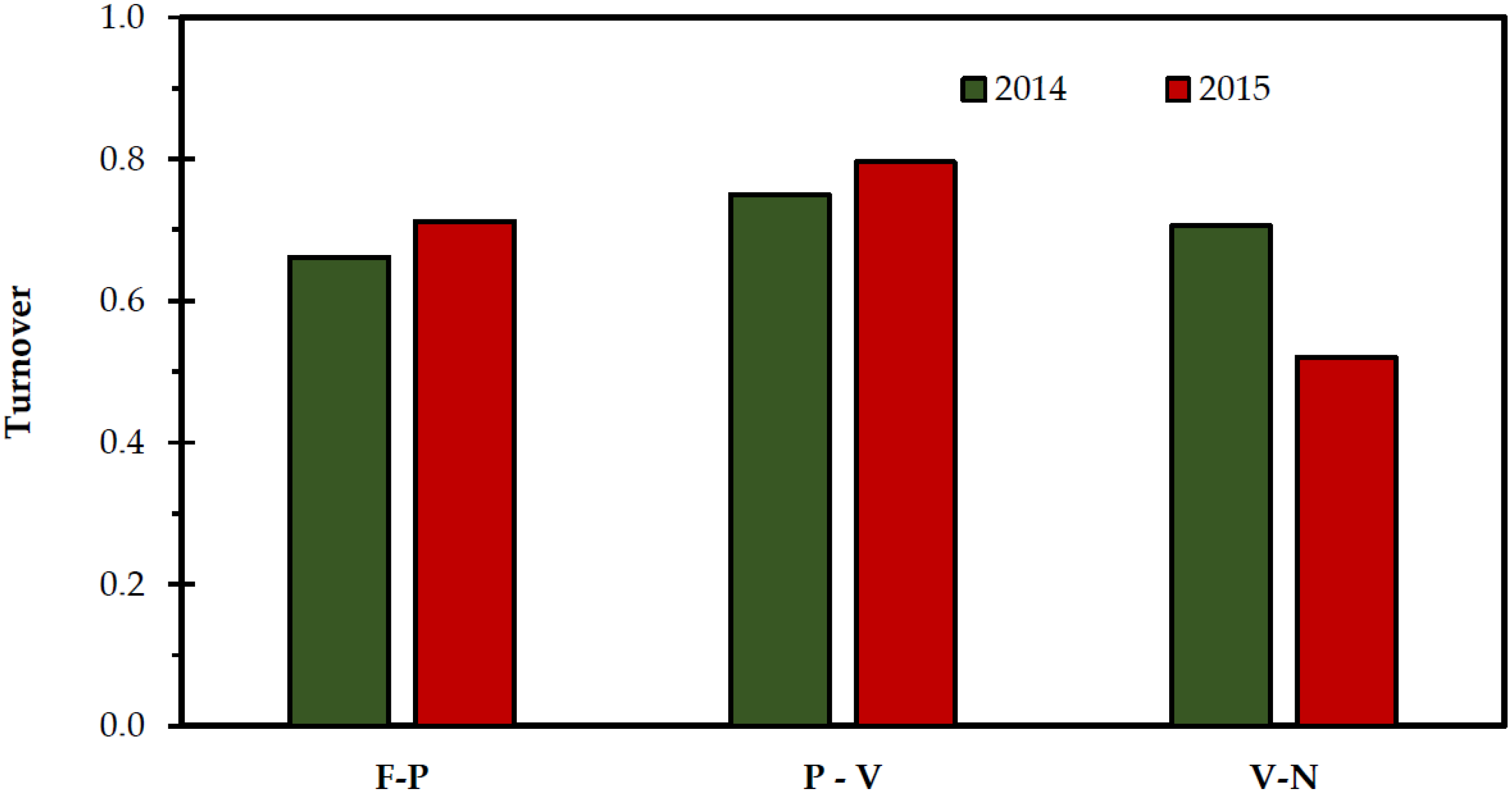
2. Results and Discussion
3. Materials and Methods
3.1. Study Area
3.2. Field Data
3.3. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- White, R.P.; Murray, S.; Rohweder, M. Pilot Analyses of Global Ecosystems: Grassland Ecosystems; Edeburn, M., Ed.; World Resources Institute: Washington, DC, USA, 2000. [Google Scholar]
- Reynolds, J.F.; Smith, D.M.; Lambin, E.F.; Turner, B.L., II; Mortimore, M.; Batterbury, S.P.; Downing, T.E.; Dowlatabadi, H.; Fernandez, R.J.; Herrick, J.E.; et al. Global desertification: Building a science for dryland development. Science 2007, 316, 847–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, W.; Mooney, H.; Cropper, A.; Capistrano, D.; Carpenter, S.; Chopra, K.; Dasgupta, P.; Dietz, T.; Duraiappah, A.; Hassan, R.; et al. Millenium Ecosystem Assessment Synthesis Report; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- Neely, C.; Bunning, S.; Wilkes, A. Review of Evidence on Drylands Pastoral Systems and Climate Change. Implications and Opportunities for Mitigation and Adaptation; FAO: Rome Italy, 2009. [Google Scholar]
- Allen-Diaz, B.; Chapin, F.S.; Díaz, S.; Howden, S.M.; Puigdefábregas, J.; Stafford Smith, M. Rangelands in a Changing Climate: Impacts, Adaptations, and Mitigation. In Climate Change 1995: Impacts, Adaptations and Mitigation of Climate Change: Scientific-Technical Analyses; Watson, R.T., Zinyowera, M.C., Moss, R.H., Dokken, D.J., Eds.; Cambridge University Press: Cambridge, UK, 1996; pp. 131–158. [Google Scholar]
- Olff, H.; Ritchie, M.E.; Prins, H.H. Global environmental controls of diversity in large herbivores. Nature 2002, 415, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkemade, R.; Reid, R.S.; Van den Berg, M.; De Leeuw, J.; Jeuken, M. Assessing the impacts of livestock production on biodiversity in rangeland ecosystems. PNAS 2013, 110, 20900–20905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, M. Experience with Grazing in Flemish Nature Reserves (Northern Belgium). Proceedings of Grazing as a Conservation Management Tool in Peatland, Goniadz, Poland, 22–26 April 2022; pp. 49–53. [Google Scholar]
- Hayati, D.; Ranjbar, Z.; Karami, E. Measuring agricultural sustainability. In Biodiversity, Biofuels, Agroforestry and Conservation Agriculture; Lichtfause, E., Ed.; Springer: Dordrecht, The Netherlands, 2010; Volume 5. [Google Scholar]
- Eteraf, H.; Telvari, A.A.R. Effects of animal grazing on some physical characteristics of loose soil in Maravetapeh Rangelands, Golestan, Iran. Pajouhesh-Va-Sazandegi 2005, 17, 8–13. [Google Scholar]
- Rahmati, O.; Samani, A.N.; Mahmoodi, N.; Mahdavi, M. Assessment of the Contribution of N-Fertilizers to Nitrate Pollution of Groundwater in Western Iran (Case Study: Ghorveh–Dehgelan Aquifer). Water Quall. Expos. Hea. 2015, 7, 143–151. [Google Scholar] [CrossRef]
- Marușca, T.; Roman, A.; Taulescu, E.; Ursu, T.M.; Popa, R.D. Detecting trends in the quality and productivity of grasslands by analyzing the historical vegetation relevés: A case study from Southeastern Carpathians, Vlădeasa Mountains (Romania). Not. Bot. Horti Agrobot. Cluj-Na. 2021, 49. [Google Scholar] [CrossRef]
- Peters, D.P.C.; Bestelmeyer, B.T.; Havstad, K.M.; Rango, A.; Archer, S.; Comrie, A.; Gimblett, R.; López-Hoffman, L.; Sala, O.E.; Vivoni, E.R. Desertification of rangelands. In Climate Vulnerability: Understanding and Addressing Threats to Essential Resources; Elsevier Inc.: Dublin, Ireland, 2013; pp. 239–258. [Google Scholar]
- Karatassiou, M.; Koukoura, Z. Protection from grazing: A way to restore vegetation in semiarid grasslands in Northern Greece. Options Mediterr. Serie A 2009, 85, 99–104. [Google Scholar]
- Adler, P.B.; Levine, J.M. Contrasting relationships between precipitation and species richness in space and time. Oikos 2007, 116, 221–232. [Google Scholar] [CrossRef]
- Herrero-Jáuregui, C.; Oesterheld, M. Effects of grazing intensity on plant richness and diversity: A meta-analysis. Oikos 2018, 127, 757–766. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, Z.; Han, G.; Schellenberg, M.P.; Wu, Q.; Gu, C. Grazing induced changes in plant diversity is a critical factor controlling grassland productivity in the Desert Steppe, Northern China. Agric. Ecosys. Environ. 2018, 265, 73–83. [Google Scholar] [CrossRef]
- Pinto, H.V.; Villa, P.M.; de Menezes, L.F.T.; Pereira, M.C.A. Effect of climate and altitude on plant community composition and richness in Brazilian inselbergs. J. Mt. Sci 2020, 17, 1931–1941. [Google Scholar] [CrossRef]
- Koerner, S.E.; Burkepile, D.E.; Fynn, R.W.S.; Burns, C.E.; Eby, S.; Govender, N.; Hagenah, N.; Matchett, K.J.; Thompson, D.I.; Wilcox, K.R.; et al. Plant community response to loss of large herbivores differs between North American and South African savanna grasslands. Ecology 2014, 95, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Biggs, J.R.; VanLeeuwen, D.M.; Holechek, J.L.; Valdez, R. Multi-Scale Analyses of Habitat Use by Elk Following Wildfire. Northwest Sci. 2010, 84, 20–32. [Google Scholar] [CrossRef]
- Holechek, J.L. An approach for setting the stocking rate. Rangel. Archives 1988, 10, 10–14. [Google Scholar]
- Austin, M.P. Searching for a model for use in vegetation analysis. Vegetatio 1980, 42, 11–21. [Google Scholar] [CrossRef]
- Austin, M.P.; Smith, T.M. A New Model for the Continuum Concept. Vegetatio 1989, 83, 35–47. [Google Scholar] [CrossRef]
- Roukos, C.; Koutsoukis, C.; Akrida-Demertzi, K.; Karatassiou, M.; Demertzis, G.P.; Kandrelis, S. The effect of altitudinal zone on soil properties, species composition and forage production in a subalpine grassland in northwest Greece. Appl. Eco. Environ. Res. 2017, 15, 609–626. [Google Scholar] [CrossRef]
- Begon, M.; Harper, J.L.; Townsend, C.R. Ecology: Individuals, Populations and Communities; Blackwell Scientific: Oxford, UK, 1996. [Google Scholar]
- Odland, A.; Birks, H.J.B. The altitudinal gradient of vascular plant richness in Aurland, western Norway. Ecography 1999, 22, 548–566. [Google Scholar] [CrossRef]
- Trigas, P.; Panitsa, M.; Tsiftsis, S. Elevational gradient of vascular plant species richness and endemism in Crete--the effect of post-isolation mountain uplift on a continental island system. PLoS ONE 2013, 8, e59425. [Google Scholar] [CrossRef]
- Rahbek, C. The elevational gradient of species richness: A uniform pattern? Ecography 1995, 18, 200–205. [Google Scholar] [CrossRef]
- Grabherr, G.; Gottfried, M.; Gruber, A.; Pauli, H. Patterns and current changes in alpine plant diversity. In Arctic And Alpine Biodiversity: Patterns, Causes and Ecosystem Consequences; Chapin, F.S., Körner, C., Eds.; Springer: Heidelberg/Berlin, Germany, 1995; Volume 113. [Google Scholar]
- Sætersdal, M.; Birks, H.J.B.; Peglar, S. Predicting changes in Fennoscandian vascular-plant species richness as a result of future climatic change. J. Biogeogr. 1998, 25, 111–112. [Google Scholar] [CrossRef]
- Kluge, J.; Bach, K.; Kessler, M. Elevational distribution and zonation of tropical pteridophyte assemblages in Costa Rica. Basic Appl. Ecol. 2008, 9, 35–43. [Google Scholar] [CrossRef]
- Sandel, B.; Arge, L.; Dalsgaard, B.; Davies, R.; Gaston, K.; Sutherland, W.; Svenning, J.-C. The Influence of Late Quaternary Climate-Change Velocity on Species Endemism. Science 2011, 334, 660–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Schneider, H.; Zhang, X.C.; Xiang, Q.P. The rise of the Himalaya enforced the diversification of SE Asian ferns by altering the monsoon regimes. BMC Plant Biol. 2012, 12, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, C.; Atchison, G. The ubiquity of alpine plant radiations: From the Andes to the Hengduan Mountains. New Phytol. 2015, 207. [Google Scholar] [CrossRef]
- Olivares, I.; Kessler, M. Regional species richness determines local species turnover in ferns. Front. Biogeogr. 2020, 12, e46818. [Google Scholar] [CrossRef]
- Yan, H.; Liang, C.; Li, Z.; Liu, Z.; Miao, B.; He, C.; Sheng, L. Impact of precipitation patterns on biomass and species richness of annuals in a dry steppe. PLoS ONE 2015, 10, e0125300. [Google Scholar] [CrossRef]
- Belgacem, A.O.; Louhaichi, M. The vulnerability of native rangeland plant species to global climate change in the West Asia and North African regions. Clim. Change 2013, 119, 451–463. [Google Scholar] [CrossRef] [Green Version]
- Hudson, L.N.; Newbold, T.; Contu, S.; Hill, S.L.L.; Lysenko, I.; De Palma, A.; Phillips, H.R.P.; Senior, R.A.; Bennett, D.J.; Booth, H.; et al. The PREDICTS database: A global database of how local terrestrial biodiversity responds to human impacts. Ecol. Evol. 2014, 4, 4701–4735. [Google Scholar] [CrossRef]
- Adeel, Z.; Safriel, U.; Niemeijer, D.; White, R. Ecosystems and Human Well-Being: Desertification Synthesis; World Resources Institute (WRI): Washington, DC, USA, 2005. [Google Scholar]
- Sala, O.E.; Chapin, F.S.; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef]
- Hanke, W.; Böhner, J.; Dreber, N.; Jürgens, N.; Schmiedel, U.; Wesuls, D.; Dengler, J. The impact of livestock grazing on plant diversity: An analysis across dryland ecosystems and scales in southern Africa. Ecol. Appl. 2014, 24, 1188–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Liu, J.; Wang, D.; Wang, H.; Wu, Y.; Lü, Z. Fencing for conservation?-The impacts of fencing on grasslands and the endangered Przewalski’s gazelle on the Tibetan Plateau. Sci. China Life Sci. 2018, 61, 1593–1595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metera, E.; Sakowski, T.; Sloniewski, K.; Romanowicz, B. Grazing as a tool to maintain biodiversity of grassland—A review. Anim. Sci. Pap. Rep. 2010, 28, 315–334. [Google Scholar]
- Wan, H.W.; Bai, Y.F.; Hooper, D.U.; Schonbach, P.; Gierus, M.; Schiborra, A.; Taube, F. Selective grazing and seasonal precipitation play key roles in shaping plant community structure of semi-arid grasslands. Landsc. Ecol. 2015, 30, 1767–1782. [Google Scholar] [CrossRef]
- Rook, A.J.; Dumont, B.; Isselstein, J.; Osoro, K.; WallisDeVries, M.F.; Parente, G.; Mills, J. Matching type of livestock to desired biodiversity outcomes in pastures—A review. Biol. Conserv. 2004, 119, 137–150. [Google Scholar] [CrossRef]
- Rahmanian, S.; Hejda, M.; Ejtehadi, H.; Farzam, M.; Pysek, P.; Memariani, F. Effects of livestock grazing on plant species diversity vary along a climatic gradient in northeastern Iran. Appl. Veg. Sci. 2020, 23, 551–561. [Google Scholar] [CrossRef]
- Schultz, N.L.; Morgan, J.W.; Lunt, I.D. Effects of grazing exclusion on plant species richness and phytomass accumulation vary across a regional productivity gradient. J. Veg. Sci. 2011, 22, 130–142. [Google Scholar] [CrossRef]
- Purvis, A.; Hector, A. Getting the measure of biodiversity. Nature 2000, 405, 212–219. [Google Scholar] [CrossRef]
- Tuomisto, H. Commentary: Do we have a consistent terminology for species diversity? Yes, if we choose to use it. Oecologia 2011, 167, 903–911. [Google Scholar] [CrossRef]
- Tuomisto, H. A consistent terminology for quantifying species diversity? Yes, it does exist. Oecologia 2010, 164, 853–860. [Google Scholar] [CrossRef]
- Jost, L. Partitioning diversity into independent alpha and beta components. Ecology 2007, 88, 2427–2439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Jost, L. The relation between evenness and diversity. Diversity 2010, 2, 207–232. [Google Scholar] [CrossRef]
- Cao, Y.; Hawkins, C.P. Weighting effective number of species measures by abundance weakens detection of diversity responses. J. Appl. Ecol. 2019, 56, 1200–1209. [Google Scholar] [CrossRef] [Green Version]
- Beck, J.; Schwanghart, W. Comparing measures of species diversity from incomplete inventories: An update. Methods Ecol. Evol. 2010, 1, 38–44. [Google Scholar] [CrossRef]
- Daly, A.J.; Baetens, J.M.; De Baets, B. Ecological Diversity: Measuring the Unmeasurable. Mathematics 2018, 6, 119. [Google Scholar] [CrossRef] [Green Version]
- Jost, L.; DeVries, P.; Walla, T.; Greeney, H.; Chao, A.; Ricotta, C. Partitioning diversity for conservation analyses. Divers. Distrib. 2010, 16, 65–76. [Google Scholar] [CrossRef]
- Tsakona, M.; Gekas, V. Desertification in Crete and the effect of global warming. In Proceedings of the 5th WSEAS, International Conference on Environment, Ecosystems and Development, Tenerife, Spain, 14–16 December 2007; pp. 211–216. [Google Scholar]
- Morianou, G.G.; Kourgialas, N.N.; Psarras, G.; Koubouris, G.C. Mapping sensitivity to desertification in Crete (Greece), the risk for agricultural areas. J. Water Clim. Change 2018, 9, 691–702. [Google Scholar] [CrossRef]
- Li, Z.; Liang, M.; Li, Z.; Mariotte, P.; Tong, X.; Zhang, J.; Dong, L.; Zheng, Y.; Ma, W.; Zhao, L.; et al. Plant functional groups mediate effects of climate and soil factors on species richness and community biomass in grasslands of Mongolian Plateau. J. Plant Ecol. 2021, 14, 679–691. [Google Scholar] [CrossRef]
- Mallen-Cooper, M.; Eldridge, D.J.; Delgado-Baquerizo, M. Livestock grazing and aridity reduce the functional diversity of biocrusts. Plant Soil 2018, 429, 175–185. [Google Scholar] [CrossRef]
- Agrawal, A.A. Overcompensation of plants in response to herbivory and the by-product benefits of mutualism. Trends Plant Sci. 2000, 5, 309–313. [Google Scholar] [CrossRef]
- Louthan, A.M.; Doak, D.F.; Goheen, J.R.; Palmer, T.M.; Pringle, R.M. Climatic stress mediates the impacts of herbivory on plant population structure and components of individual fitness. J. Ecol. 2013, 101, 1074–1083. [Google Scholar] [CrossRef]
- Milchunas, D.G.; Sala, O.E.; Lauenroth, W.K. A generalized model of the effects of grazing by large herbivores on grassland community structure. Am. Nat. 1988, 132, 87–106. [Google Scholar] [CrossRef]
- Oñatibia, G.R.; Amengual, G.; Boyero, L.; Aguiar, M.R. Aridity exacerbates grazing-induced rangeland degradation: A population approach for dominant grasses. J. Appl. Ecol 2020, 57, 1999–2009. [Google Scholar] [CrossRef]
- Stuth, J.W. Foraging behavior. In Grazing Management: An Ecological Perspective; Heitschmidt, R.K., Stuth, J.W., Eds.; Timber Press: Portland, OR, USA, 1991. [Google Scholar]
- Eldridge, D.J.; Delgado-Baquerizo, M.; Travers, S.K.; Val, J.; Oliver, I.; Dorrough, J.W.; Soliveres, S. Livestock activity increases exotic plant richness, but wildlife increases native richness, with stronger effects under low productivity. J. Appl. Ecol. 2018, 55, 766–776. [Google Scholar] [CrossRef]
- Bhandari, J.; Zhang, Y. Effect of altitude and soil properties on biomass and plant richness in the grasslands of Tibet, China, and Manang District, Nepal. Ecosphere 2019, 10, e02915. [Google Scholar] [CrossRef]
- Abraham, E.; Karatassiou, M.; Parissi, Z.; Koukoura, Z.; Tsiouvaras, C. Long-term effects of grazing on composition in various habitats of a mountainous area in Central Greece. Options Méditerr. Série A 2009, 73–78. [Google Scholar]
- Tsiourlis, G.; Kasapidis, P.; Konstantinidis, P. The role of grazing on the maintain and degradation of Mediterranean Ecosystems in Central Crete, Greece. In Proceedings of the Forest Research: A Challenge for an Integrated European Approach, Thessaloniki, Greece, 27 August–1 September 2001; pp. 715–721. [Google Scholar]
- Papanastasis, V.P.; Kyriakakis, S.; Kazakis, G. Plant diversity in relation to overgrazing and burning in mountain Mediterranean ecosystems. J. Mediterr. Ecol. 2002, 3, 53–64. [Google Scholar]
- Ojima, D.S.; Aicher, R.; Archer, S.R.; Bailey, D.W.; Casby-Horton, S.M.; Cavallaro, N.; Reyes, J.J.; Tanaka, J.A.; Washington-Allen, R.A. A climate change indicator framework for rangelands and pastures of the USA. Clim. Change 2020, 163, 1733–1750. [Google Scholar] [CrossRef]
- Lund, H.G. Accounting for the World’s Rangelands. Rangelands 2007, 29, 3–10. [Google Scholar] [CrossRef]
- Margaris, N.S.; Vokou, D. Structural and physiological features of woody plants in phryganic ecosystems related to adaptive mechanisms. Ecol. Mediterr. 1982, 8, 449–459. [Google Scholar] [CrossRef]
- Thompson, J.D. Plant Evolution in the Mediterranean: Insights for Conservation; Oxford University Press: New York, NY, USA, 2020. [Google Scholar]
- Atherden, M.; Hall, J. Human impact on vegetation in the White Mountains of Crete since AD 500. Holocene 1999, 9, 183–193. [Google Scholar] [CrossRef]
- Papanastasis, V.P.; Kyriakakis, S.; Kazakis, G.; Abid, M.; Doulis, A. Plant cover as a tool for monitoring desertification in mountain Mediterranean rangelands. Manag. Environ. Qual. 2003, 14, 69–81. [Google Scholar] [CrossRef]
- Vogiatzakis, I.; Griffiths, G.H.; Mannion, A.M. Environmental factors and vegetation composition, Lefka Ori massif, Crete, S. Aegean. Glob. Ecol. Biogeogr. 2003, 12, 131–146. [Google Scholar] [CrossRef]
- Sklenár, P.; Ramsay, P.M. Diversity of zonal páramo plant communities in Ecuador. Divers. Distrib. 2001, 7, 113–124. [Google Scholar] [CrossRef]
- Mekonnen, Z.A.; Riley, W.J.; Randerson, J.T.; Grant, R.F.; Rogers, B.M. Expansion of high-latitude deciduous forests driven by interactions between climate warming and fire. Nat. Plants 2019, 5, 952–958. [Google Scholar] [CrossRef] [Green Version]
- Zupo, T.; Daibes, L.F.; Pausas, J.G.; Fidelis, A. Post-fire regeneration strategies in a frequently burned Cerrado community. J. Veg. Sci. 2021, 32, e12968. [Google Scholar] [CrossRef]
- Gatti, R.C.; Amoroso, N.; Monaco, A. Estimating and comparing biodiversity with a single universal metric. Ecol. Mod. 2020, 424, 109020. [Google Scholar] [CrossRef]
- Mena, J.L.; Vázquez-Domínguez, E. Species turnover on elevational gradients in small rodents. Glob. Ecol. Biogeogr. 2005, 14, 539–547. [Google Scholar] [CrossRef]
- Xiang, M.; Wu, J.; Wu, J.; Guo, Y.; Lha, D.; Pan, Y.; Zhang, X. Heavy Grazing Altered the Biodiversity–Productivity Relationship of Alpine Grasslands in Lhasa River Valley, Tibet. Front. Ecol. Evol. 2021, 9, 698707. [Google Scholar] [CrossRef]
- Zawierucha, K.; Smykla, J.; Michalczyk, Ł.; Gołdyn, B.; Kaczmarek, Ł. Distribution and diversity of Tardigrada along altitudinal gradients in the Hornsund, Spitsbergen (Arctic). Polar Res. 2015, 34, 24168. [Google Scholar] [CrossRef]
- Lantz, T.C.; Gergel, S.E.; Henry, G.H.R. Response of green alder (Alnus viridis subsp. fruticosa) patch dynamics and plant community composition to fire and regional temperature in north-western Canada. J. Biogeogr. 2010, 37, 1597–1610. [Google Scholar] [CrossRef]
- Coop, J.D.; Massatti, R.T.; Schoettle, A.W. Subalpine vegetation pattern three decades after stand-replacing fire: Effects of landscape context and topography on plant community composition, tree regeneration, and diversity. J. Veg. Sci. 2010, 21, 472–487. [Google Scholar] [CrossRef]
- McKenzie, D.A.; Tinker, D.B. Fire-induced shifts in overstory tree species composition and associated understory plant composition in Glacier National Park, Montana. Plant Ecol. 2012, 213, 207–224. [Google Scholar] [CrossRef]
- Staver, A.C.; Botha, J.; Hedin, L. Soils and fire jointly determine vegetation structure in an African savanna. New Phytol. 2017, 216, 1151–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omidipour, R.; Tahmasebi, P.; Faal Faizabadi, M.; Faramarzi, M.; Ebrahimi, A. Does β diversity predict ecosystem productivity better than species diversity? Ecol. Indic. 2021, 122, 107212. [Google Scholar] [CrossRef]
- Morris, E.K.; Caruso, T.; Buscot, F.; Fischer, M.; Hancock, C.; Maier, T.S.; Meiners, T.; Müller, C.; Obermaier, E.; Prati, D.; et al. Choosing and using diversity indices: Insights for ecological applications from the German Biodiversity Exploratories. Ecol. Evol. 2014, 4, 3514–3524. [Google Scholar] [CrossRef] [Green Version]
- Gibson, D.J.; Newman, J.A. Grasslands and climate change: An overview. In Grasslands and Climate Change; Gibson, D.J., Newman, J.A., Eds.; Cambridge University Press: Cambridge, UK, 2019; pp. 3–18. [Google Scholar]
- Sanaei, A.; Ali, A.; Chahouki, M.A.Z.; Jafari, M. Plant coverage is a potential ecological indicator for species diversity and aboveground biomass in semi-steppe rangelands. Ecol. Indic. 2018, 93, 256–266. [Google Scholar] [CrossRef]
- WallisDeVries, M.F.; Poschlod, P.; Willems, J.H. Challenges for the conservation of calcareous grasslands in northwestern Europe: Integrating the requirements of flora and fauna. Biol. Conserv. 2002, 104, 265–273. [Google Scholar] [CrossRef]
- Benthien, O.; Braun, M.; Riemann, J.C.; Stolter, C. Long-term effect of sheep and goat grazing on plant diversity in a semi-natural dry grassland habitat. Heliyon 2018, 4, e00556. [Google Scholar] [CrossRef] [Green Version]
- Mohapatra, J.; Singh, C.P.; Hamid, M.; Khuroo, A.A.; Malik, A.H.; Pandya, H.A. Assessment of the alpine plant species biodiversity in the western Himalaya using Resourcesat-2 imagery and field survey. J. Earth Syst. Sci. 2019, 128, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Kazakis, G.; Ghosn, D.; Vogiatzakis, I.; Papanastasis, V. Vascular plant diversity and climate change in the alpine zone of the Lefka Ori, Crete. Biodivers. Conserv. 2007, 16, 1603–1615. [Google Scholar] [CrossRef]
- Ibanez, T.; Hart, P.; Ainsworth, A.; Gross, J.; Monello, R. Factors associated with alien plant richness, cover and composition differ in tropical island forests. Divers. Distrib. 2019, 25, 1910–1923. [Google Scholar] [CrossRef] [Green Version]
- Todd, S.W. Gradients in vegetation cover, structure and species richness of Nama-Karoo shrublands in relation to distance from livestock watering points. J. Appl. Ecol. 2006, 43, 293–304. [Google Scholar] [CrossRef]
- Stirling, G.; Wilsey, B. Empirical Relationships between Species Richness, Evenness, and Proportional Diversity. Am. Nat. 2001, 158, 286–299. [Google Scholar] [CrossRef] [PubMed]
- Nagendra, H. Opposite trends in response for the Shannon and Simpson indices of landscape diversity. Appl. Geogr. 2002, 22, 175–186. [Google Scholar] [CrossRef]
- Cook, C.W.; Stubbendieck, J. Range Research: Basic Problems and Techniques; Society for Range Management: Denver, CO, USA, 1986. [Google Scholar]
- Bonham, C.D. Measurements for Terrestrial Vegetation; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Deléglise, C.; Loucougaray, G.; Alard, D. Effects of grazing exclusion on the spatial variability of subalpine plant communities: A multiscale approach. Basic Appl. Ecol. 2011, 12, 609–619. [Google Scholar] [CrossRef]
- Hill, M.O. Diversity and evenness: A unifying notation and its consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.N.; Chiu, C.H.; Jost, L. Unifying Species Diversity, Phylogenetic Diversity, Functional Diversity, and Related Similarity and Differentiation Measures Through Hill Numbers. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 297–324. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Gurevitch, J.; Scheiner, S.M.; Fox, G.A. The Ecology of Plants, 3rd ed.; Sinauer Associates Sunderland, Oxfrord University Press: Cary, NC, USA, 2021. [Google Scholar]
- Heady, H.F.; Child, R.D. Rangeland Ecology and Management; Routledge: New York, NY, USA, 2019. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| R | SE | GS | IdM | T | P | Altitude | |
|---|---|---|---|---|---|---|---|
| R | 1 | ||||||
| SE | −0.073 | 1 | |||||
| GS | −0.280 | 0.861 ** | 1 | ||||
| IdM | 0.059 | −0.876 ** | −0.678 | 1 | |||
| T | −0.295 | 0.894 ** | 0.795 * | −0.954 ** | 1 | ||
| P | −0.174 | −0.766 * | −0.498 | 0.947 ** | −0.809 * | 1 | |
| Altitude | 0.218 | −0.888 ** | −0.763 * | 0.980 ** | −0.986 ** | 0.882 ** | 1 |
| Forage Production (g m−2) | FUP (%) | |||||
|---|---|---|---|---|---|---|
| Fenced Plots | Grazed Sites | |||||
| Rangeland (R) | 2014 | 2015 | 2014 | 2015 | 2014 | 2015 |
| Nida | 122.2 ± 6.03 b | 120.2 ± 5.5 bc | 15.8 ± 1.4 d | 18.8 ± 1.1 d | 87.1 ± 0.9 a | 84.4 ± 0.5 a |
| Vroulidia | 126.7 ± 9.5 b | 116.5 ± 7.3 c | 29.9 ± 1.7 c | 30.2 ± 2.2 c | 76.1 ± 0.8 b | 74.1 ± 1.0 b |
| Pyrathi | 189.2 ± 24.2 a | 173.6 ± 22.2 a | 45.7 ± 6.5 b | 42.2 ± 4.9 b | 76 ± 0.7 b | 75.1± 1.8 b |
| Faistos | 134.4 ± 4.0 b | 151.4 ± 5.3 b | 111.5 ± 3.3 a | 128.9 ± 4.8 a | 17.00 ± 0.8 c | 14.9 ± 0.8 c |
| R | <0.001 ** | <0.001 ** | <0.001 ** | |||
| Year | 0.748 ns | 0.084 ns | 0.003 * | |||
| R X Year | 0.567 ns | 0.017 * | 0.761 ns | |||
| Wald Chi-Square | df | p-Value | |
|---|---|---|---|
| Year | 0.033 | 1 | p ≥ 0.05 ns |
| Rangeland | 0.045 | 3 | p ≥ 0.05 ns |
| Functional group | 116.816 | 4 | <0.001 * |
| Year * Rangeland | 0.014 | 3 | p ≥ 0.05 ns |
| Rangeland * Functional group | 80.120 | 12 | <0.001 * |
| Lowland Rangelands | Highland Rangelands | |||||||
|---|---|---|---|---|---|---|---|---|
| Faistos | Pyrathi | Vroulidia | Nida | |||||
| 2014 | 2015 | 2014 | 2015 | 2014 | 2015 | 2014 | 2015 | |
| * HE/R | 0.74 | 0.75 | 0.50 | 0.58 | 0.66 | 0.59 | 0.43 | 0.31 |
| * GS/R | 0.60 | 0.55 | 0.29 | 0.41 | 0.49 | 0.46 | 0.23 | 0.23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karatassiou, M.; Parissi, Z.M.; Panajiotidis, S.; Stergiou, A. Impact of Grazing on Diversity of Semi-Arid Rangelands in Crete Island in the Context of Climatic Change. Plants 2022, 11, 982. https://doi.org/10.3390/plants11070982
Karatassiou M, Parissi ZM, Panajiotidis S, Stergiou A. Impact of Grazing on Diversity of Semi-Arid Rangelands in Crete Island in the Context of Climatic Change. Plants. 2022; 11(7):982. https://doi.org/10.3390/plants11070982
Chicago/Turabian StyleKaratassiou, Maria, Zoi M. Parissi, Sampson Panajiotidis, and Afroditi Stergiou. 2022. "Impact of Grazing on Diversity of Semi-Arid Rangelands in Crete Island in the Context of Climatic Change" Plants 11, no. 7: 982. https://doi.org/10.3390/plants11070982