
Intraspecific Diversity in Aquatic Ecosystems: Comparison between Spirodela polyrhiza and Lemna minor in Natural Populations of Duckweed



Abstract
:1. Introduction
2. Results
2.1. Evaluation of AFLP Fingerprinting
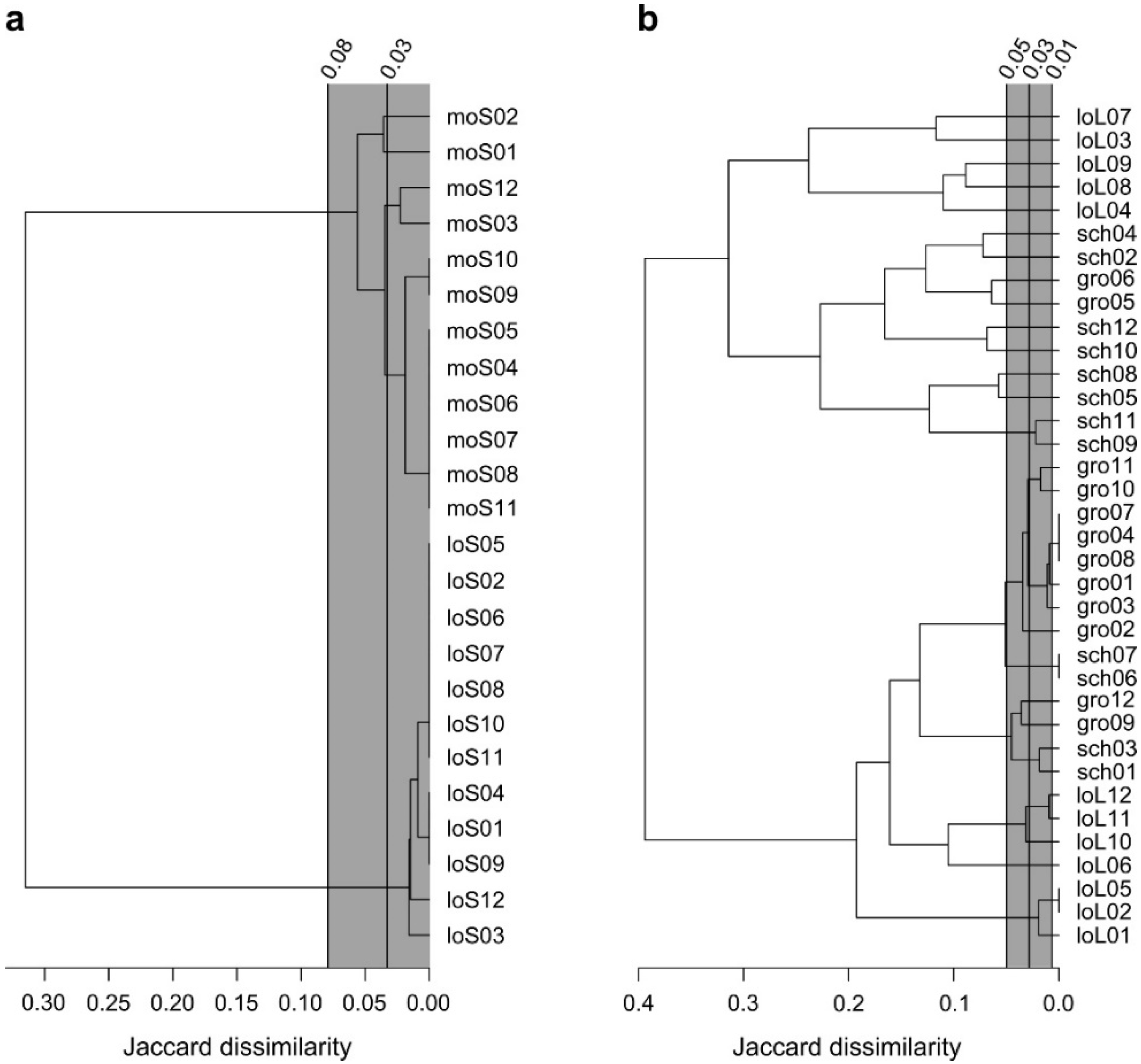
2.2. Average-Linkage Cluster Analysis
3. Discussion
3.1. No Clonal Diversity Could Be Detected within Natural Populations of Spirodela polyrhiza
3.2. Natural Population of Lemna minor Comprises of Several Distinct Clones
3.3. Natural Transport of Clones between Ponds
4. Materials and Methods
4.1. Taxon Sampling and Cultivation of Clones
4.2. DNA Isolation and AFLP Analysis
4.3. Data Analyses
5. Conclusions
- A remarkable intraspecific biodiversity exists in natural populations of Lemna minor, but not in populations of Spirodela polyrhiza, suggesting either a higher mutation rate or a higher rate of sexual reproduction in Lemna minor.
- The intraspecific biodiversity in the ponds was further enhanced by the putative transfer (most probably epizoochorous by birds) of plants between closely spaced ponds.
- The high intraspecific diversity of Lemna minor may have a role in the adaptation of the natural duckweed populations to the changing environmental conditions.
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tippery, N.P.; Les, D.H.; Appenroth, K.J.; Sree, K.S.; Crawford, D.J.; Bog, M. Lemnaceae and Orontiaceae are phylogenetically and morphologically distinct from Araceae. Plants 2021, 10, 2639. [Google Scholar] [CrossRef] [PubMed]
- Sree, K.S.; Bog, M.; Appenroth, K.J. Taxonomy of duckweed (Lemnaceae), potential crop plants. Emir. J. Food Agric. 2016, 28, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Bog, M.; Appenroth, K.J.; Sree, K.S. Key to the determination of taxa of Lemnaceae: An update. Nord. J. Bot. 2020, 38, e02658. [Google Scholar] [CrossRef]
- Bog, M.; Sree, K.S.; Fuchs, J.; Phuong, T.N.H.; Schubert, I.; Kuever, J.; Rabenstein, A.; Paolacci, S.; Jansen, M.A.K.; Appenroth, K.J. A taxonomic revision of Lemna sect. Uninerves (Lemnaceae). Taxon 2020, 69, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Acosta, K.; Appenroth, K.J.; Borisjuk, L.; Edelman, M.; Heinig, U.; Jansen, M.A.K.; Oyama, T.; Pasaribu, B.; Schubert, I.; Sorrels, S.; et al. Return of the Lemnaceae: Duckweed as a model plant system in the genomics and post-genomics era (Review). Plant Cell 2021, 33, 3207–3234. [Google Scholar] [CrossRef]
- Sree, K.S.; Sudakaran, S.; Appenroth, K.J. How fast can angiosperms grow? Species and clonal diversity of growth rates in the genus Wolffia (Lemnaceae). Acta Physiol. Plant. 2015, 37, 204. [Google Scholar] [CrossRef]
- Ziegler, P.; Adelmann, K.; Zimmer, S.; Schmidt, C.; Appenroth, K.J. Relative in vitro growth rates of duckweeds (Lemnaceae)—The most rapidly growing higher plants. Plant Biol. 2015, 17 (Suppl. S1), 33–41. [Google Scholar] [CrossRef]
- Krajncic, B.; Devide, Z. Report on photoperiodic responses in Lemnaceae from Slovenia. Ber. Geobot. Inst. ETH Zur. 1980, 47, 75–86. [Google Scholar]
- Pieterse, A.H. Is flowering in Lemnaceae stress-induced? A review. Aquat. Bot. 2013, 104, 1–4. [Google Scholar] [CrossRef]
- Ho, E.K.H.; Bartkowska, M.; Wright, S.I.; Agrawal, A.F. Population genomics of the facultatively asexual duckweed Spirodela polyrhiza. New Phytol. 2019, 224, 1361–1371. [Google Scholar] [CrossRef]
- Kuehdorf, K.; Jetschke, G.; Ballani, L.; Appenroth, K.J. The clonal dependence of turion formation in the duckweed Spirodela polyrhiza—An ecogeographical approach. Physiol. Plant. 2014, 150, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Appenroth, K.J.; Sree, K.S.; Bog, M.; Ecker, J.; Seeliger, C.; Boehm, V.; Lorkowski, S.; Sommer, K.; Vetter, W.; Tolzin-Basch, K.; et al. Nutritional value of the duckweed species of the genus Wolffia (Lemnaceae) as human food. Front. Chem. 2018, 6, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sree, K.S.; Adelmann, K.; Garcia, C.; Lam, E.; Appenroth, K.-J. Natural variance in salt tolerance and induction of starch accumulation in duckweeds. Planta 2015, 241, 1395–1404. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.B.; Zhu, M.; Yu, C.J.; Wang, Y.; Liu, Y.; Li, M.L.; Sun, Y.D.; Zhao, J.S.; Zhou, G.K. Large-scale screening and characterisation of Lemna aequinoctialis and Spirodela polyrhiza strains for starch production. Plant Biol. 2018, 20, 357–364. [Google Scholar] [CrossRef]
- Appenroth, K.J.; Ziegler, P.; Sree, K.S. Accumulation of starch in duckweeds (Lemnaceae), potential energy plants. Physiol. Mol. Biol. Plants 2021, 27, 2621–2633. [Google Scholar] [CrossRef]
- Appenroth, K.J.; Sree, K.S.; Fakhoorian, T.; Lam, E. Resurgence of duckweed research and applications: Report from the 3rd International Duckweed Conference. Plant Mol. Biol. 2015, 89, 647–654. [Google Scholar] [CrossRef]
- Sree, K.S.; Khurana, J.P. (Eds.) Duckweed: Biological Chemistry and Applications. In Frontiers Abstract Book, Proceedings of the Fourth International Conference on Duckweed Research and Applications, Kerala, India, 23–26 October 2017; Frontiers Media SA: Lausanne, Switzerland, 2018. [Google Scholar]
- Bog, M.; Baumbach, H.; Schween, U.; Hellwig, F.; Landolt, E.; Appenroth, K.-J. Genetic structure of the genus Lemna L. (Lemnaceae) as revealed by amplified fragment length polymorphism. Planta 2010, 232, 609–619. [Google Scholar] [CrossRef]
- Bog, M.; Schneider, P.; Hellwig, F.; Sachse, S.; Kochieva, E.Z.; Martyrosian, E.; Landolt, E.; Appenroth, K.J. Genetic characterization and barcoding of taxa in the genus Wolffia Horkel ex Schleid. (Lemnaceae) as revealed by two plastidic markers and amplified fragment length polymorphism (AFLP). Planta 2013, 237, 1–13. [Google Scholar] [CrossRef]
- Bog, M.; Lautenschlager, U.; Landrock, M.F.; Landolt, E.; Fuchs, J.; Sree, K.S.; Oberprieler, C.; Appenroth, K.-J. Genetic characterization and barcoding of taxa in the genera Landoltia and Spirodela (Lemnaceae) by three plastidic markers and amplified fragment length polymorphism (AFLP). Hydrobiologia 2015, 749, 169–182. [Google Scholar] [CrossRef]
- Bog, M.; Landrock, F.M.; Drefahl, D.; Sree, K.S.; Appenroth, K.J. Fingerprinting by Amplified Fragment Length Polymorphism (AFLP) and barcoding by three plastidic markers in the genus Wolffiella Hegelm. Plant Syst. Evol. 2018, 304, 373–386. [Google Scholar] [CrossRef]
- Les, D.H.; Crawford, D.J.; Landolt, E.; Gabel, J.D.; Kimball, R.T. Phylogeny and systematics of Lemnaceae, the duckweed family. Syst. Bot. 2002, 27, 221–240. [Google Scholar]
- Tippery, N.P.; Les, D.H.; Crawford, D.J. Evaluation of phylogenetic relationships in Lemnaceae using nuclear ribosomal data. Plant Biol. 2015, 17 (Suppl. S1), 50–58. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Ma, S.; Huang, M.; Peng, M.; Bog, M.; Sree, K.S.; Appenroth, K.J.; Zhang, J. Species distribution, genetic diversity and barcoding in the duckweed family (Lemnaceae). Hydrobiologia 2015, 743, 75–87. [Google Scholar] [CrossRef]
- Feng, B.; Fang, Y.; Xu, Z.; Xiang, C.; Thou, C.; Jiang, F.; Wang, T.; Zhao, H. Development of a new marker system for identification of Spirodela polyrhiza and Landoltia punctata. Int. J. Genomics 2017, 2017, 5196763. [Google Scholar] [CrossRef] [Green Version]
- Bog, M.; Appenroth, K.J.; Sree, K.S. Duckweed (Lemnaceae): Its molecular taxonomy. A Review. Front. Sustain. Food Syst. 2019, 3, 117. [Google Scholar] [CrossRef]
- Borisjuk, N.; Chu, P.; Gutierrez, R.; Zhang, H.; Acosta, K.; Friesen, N.; Sree, K.S.; Garcia, C.; Appenroth, K.J.; Lam, E. Assessment, validation and deployment strategy of a two barcode protocol for facile genotyping of duckweed species. Plant Biol. 2015, 17, 42–49. [Google Scholar] [CrossRef]
- Chu, P.; Wilson, G.M.; Michael, T.P.; Vaiciunas, J.; Honig, J.; Lam, E. Sequence-guided approach to genotyping plant clones and species using polymorphic NB-ARC-related genes. Plant Mol. Biol. 2018, 98, 219–231. [Google Scholar] [CrossRef]
- Bog, M.; Xu, S.; Himmelbach, A.; Brandt, R.; Wagner, F.; Appenroth, K.J.; Sree, K.S. Genotyping-by-Sequencing for species delimitation in the section Uninerves Hegelm. (genus Lemna L.). In Compendium of Plant Genomes: The Duckweed Genomes, 1st ed.; Cao, X.H., Fourounjian, P., Wang, W., Eds.; Springer: Berlin/Heidelberg, Germany, 2020; ISBN 455 978-3-030-11045-1. [Google Scholar]
- Xu, N.; Hu, F.; Wu, J.; Zhang, W.; Wang, M.; Zhu, M.; Ke, J. Characterization of 19 polymorphic SSR markers in Spirodela polyrhiza (Lemnaceae) and cross-amplification in Lemna perpusilla. Appl. Plant Sci. 2018, 6, e1153. [Google Scholar] [CrossRef]
- Fu, L.; Ding, Z.; Kumpeangkeaw, A.; Tan, D.; Han, B.; Sun, X.; Zhang, J. De novo assembly, transcriptome characterization, and simple sequence repeat marker development in duckweed Lemna gibba. Physiol. Mol. Biol. Plants 2020, 26, 133–142. [Google Scholar] [CrossRef]
- Cole, C.T.; Voskuil, M.I. Population genetic structure in duckweed (Lemna minor, Lemnaceae). Can. J. Bot. 1996, 74, 220–230. [Google Scholar] [CrossRef]
- Xu, S.; Stapley, J.; Gablenz, S.; Boyer, J.; Appenroth, K.J.; Sree, K.S.; Gershenzon, J.; Widmer, A.; Huber, M. Low mutation rate determines low genetic variation in the greater duckweed. Nat. Commun. 2019, 10, 1857. [Google Scholar] [CrossRef] [PubMed]
- Hoang, P.N.T.; Michael, T.P.; Gilbert, S.; Chu, P.; Motley, T.S.; Appenroth, K.J.; Schubert, I.; Lam, E. Generating a high-confidence reference genome map of the Greater Duckweed by integration of cytogenomic, optical mapping and Oxford Nanopore technologies. Plant J. 2018, 96, 670–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michael, T.P.; Bryant, D.; Gutierrez, R.; Borisjuk, N.; Chu, P.; Zhang, H.; Xia, J.; Zhou, J.; Peng, H.; El Baidouri, M.; et al. Comprehensive definition of genome features in Spirodela polyrhiza by high-depth physical mapping and short-read DNA sequencing strategies. Plant J. 2017, 89, 617–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Kerstetter, R.A.; Michael, T.P. Evolution of Genome Size in Duckweeds (Lemnaceae). J. Bot. 2011, 2011, 570319. [Google Scholar] [CrossRef] [Green Version]
- An, D.; Li, C.S.; Zhou, Y.; Wu, Y.R.; Wang, W.Q. Genomes and transcriptomes of duckweeds. Front. Chem. 2018, 6, 230. [Google Scholar] [CrossRef] [PubMed]
- Braglia, L.; Lauria, M.; Appenroth, K.J.; Bog, M.; Breviario, D.; Grasso, A.; Gavazzi, F.; Morello, L. Duckweed species genotyping and interspecific hybrid discovery by tubulin-based polymorphism fingerprinting. Front. Plant Sci. 2021, 12, 625670. [Google Scholar] [CrossRef]
- Braglia, L.; Breviario, D.; Giani, S.; Gavazzi, F.; De Gregori, J.; Morello, L. New insights into interspecific hybridization in Lemna L. Sect. Lemna (Lemnaceae Martinov). Plants 2021, 12, 2767. [Google Scholar] [CrossRef]
- Tang, J.; Zhang, F.; Cui, W.; Ma, J. Genetic structure of duckweed population of Spirodela, Landoltia and Lemna from Lake Tai, China. Planta 2014, 239, 1299–1307. [Google Scholar] [CrossRef]
- El-Kholy, A.S.; Youssef, M.S.; Eid, E.M. Genetic diversity of Lemna gibba L. and Lemna minor L. populations in Nile delta based in biochemical and ISSR markers. Egypt. J. Exp. Biol. Bot. 2015, 11, 11–19. [Google Scholar]
- Paolaci, S.; Bog, M.; Lautenschlager, U.; Bonfield, R.; Appenroth, K.J.; Oberprieler, C.; Jansen, M.A.K. Clonal diversity amongst island populations of alien, invasive Lemna minuta Kunth. Biol. Invasions 2021, 23, 2649–2660. [Google Scholar] [CrossRef]
- Coughlan, N.E.; Kelly, T.C.; Jansen, M.A.K. Mallard duck (Anas platyrhynchos)-mediated dispersal of Lemnaceae: A contributing factor in the spread of invasive Lemna minuta? Plant Biol. 2015, 17 (Suppl. S1), 108–114. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, N.E.; Kelly, T.C.; Jansen, M.A.K. “Step by step”: High frequency short-distance epizoochorous dispersal of aquatic macrophytes. Biol. Invasions 2017, 19, 625–634. [Google Scholar] [CrossRef]
- Coughlan, N.E.; Kelly, T.C.; Davenport, J.; Jansen, M.A.K. Humid microclimates within the plumage of mallard ducks (Anas platyrhynchos) can potentially facilitate long distance dispersal of propagules. Acta Oecol. 2015, 65–66, 17–23. [Google Scholar] [CrossRef]
- Hicks, L.E. The Lemnaceae of Indiana. Am. Midl. Nat. 1937, 18, 774–789. [Google Scholar] [CrossRef]
- Appenroth, K.J. Sterilization of Duckweed. Duckweed Forum 2015, 3, 90–91. Available online: www.ruduckweed.org/ (accessed on 12 February 2022).
- Appenroth, K.-J.; Teller, S.; Horn, M. Photophysiology of turion formation and germination in Spirodela polyrhiza. Biol. Plant. 1996, 38, 95–106. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; Van de Lee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M.; et al. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef] [Green Version]
- Baumbach, H.; Hellwig, F.H. Genetic differentiation of metallicolous and non-metallicolous Armeria maritima (MILL.) WILLD. Taxa (Plumbaginaceae) in Central Europe. Plant Syst. Evol. 2007, 269, 245–258. [Google Scholar] [CrossRef]
- Schlüter, P.M.; Harris, S.A. Analysis of multilocus fingerprinting data sets containing missing data. Mol. Ecol. Notes 2006, 6, 569–572. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2015. Available online: https://www.R-project.org (accessed on 12 February 2022).



{kind=link}
{kind=link}
| S. polyrhiza | L. minor | ||||
|---|---|---|---|---|---|
| moS | loS | loL | Gro | sch | |
| mean band presence per sample (mean ± SD) | 53 ± 5 | 72 ± 7 | 92 ± 17 | 107 ± 15 | 94 ± 14 |
| number polymorphic bands | 7 | 3 | 70 | 56 | 61 |
| number fixed bands | 52 | 73 | 53 | 68 | 67 |
| number private bands | 5 | 22 | 4 | 0 | 4 |
| number fixed private bands | 2 | 19 | 0 | 0 | 0 |
| Shannon’s index | 2.2 | 0.9 | 28.8 | 16.2 | 28.9 |
| PhiST | 0.992 | 0.241 | |||
| Distinct Clones | Moscow (mo) | Lotschen (lo) | Groeben (gro) | Schloeben (sch) |
|---|---|---|---|---|
| Spirodela1 | moS01–12 | |||
| Spirodela2 | loS01–12 | |||
| Lemna01 | loL01, 02, 05 | |||
| Lemna02 | loL06 | |||
| Lemna03 | loL10–12 | |||
| Lemna04 | gro09, 12 | sch01, 03 | ||
| Lemna05 | sch06–07 | |||
| Lemna06 | gro01–04, 07–08, 10–11 | |||
| Lemna07 | sch09, 11 | |||
| Lemna08 | sch05 | |||
| Lemna09 | sch08 | |||
| Lemna10 | sch10 | |||
| Lemna11 | sch12 | |||
| Lemna12 | gro05 | |||
| Lemna13 | gro06 | |||
| Lemna14 | sch02 | |||
| Lemna15 | sch04 | |||
| Lemna16 | loL04 | |||
| Lemna17 | loL08 | |||
| Lemna18 | loL09 | |||
| Lemna19 | loL03 | |||
| Lemna20 | loL07 |
| Species | Location | Samples | Pond Size |
|---|---|---|---|
| Lemna minor | Groeben, Thuringia, Germany 50°53′06″ N 11°40′53″ E | gro01–gro12 | 900 m2 |
| Lemna minor | Schloeben, Thuringia, Germany 50°53′32″ N 11°41′26″ E | sch01–sch12 | 400 m2 |
| Lemna minor | Lotschen, Thuringia, Germany 50°53′07″ N 11°42′56″ E | loL01–loL12 | 700 m2 |
| Spirodela polyrhiza | Lotschen, 50°53′07″ N 11°42′56″ E | loS01–loS12 | 700 m2 |
| Spirodela polyrhiza | Botanical Garden Moscow, Russia 55°50′36″ N 37°35′23″ E | moS01–moS12 | 100 m2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bog, M.; Appenroth, K.-J.; Schneider, P.; Sree, K.S. Intraspecific Diversity in Aquatic Ecosystems: Comparison between Spirodela polyrhiza and Lemna minor in Natural Populations of Duckweed. Plants 2022, 11, 968. https://doi.org/10.3390/plants11070968
Bog M, Appenroth K-J, Schneider P, Sree KS. Intraspecific Diversity in Aquatic Ecosystems: Comparison between Spirodela polyrhiza and Lemna minor in Natural Populations of Duckweed. Plants. 2022; 11(7):968. https://doi.org/10.3390/plants11070968
Chicago/Turabian StyleBog, Manuela, Klaus-Juergen Appenroth, Philipp Schneider, and K. Sowjanya Sree. 2022. "Intraspecific Diversity in Aquatic Ecosystems: Comparison between Spirodela polyrhiza and Lemna minor in Natural Populations of Duckweed" Plants 11, no. 7: 968. https://doi.org/10.3390/plants11070968