
Drought Stress Responses: Coping Strategy and Resistance

Department of Plant Physiology, Poznan University of Life Sciences, Wołyńska 35, 60-637 Poznań, Poland
Plants 2022, 11(7), 922; https://doi.org/10.3390/plants11070922
Submission received: 17 March 2022
/
Accepted: 28 March 2022
/
Published: 29 March 2022
(This article belongs to the Special Issue 10th Anniversary of Plants—Recent Advances and Perspectives)
Abstract
:Plants’ resistance to stress factors is a complex trait that is a result of changes at the molecular, metabolic, and physiological levels. The plant resistance strategy means the ability to survive, recover, and reproduce under adverse conditions. Harmful environmental factors affect the state of stress in plant tissues, which creates a signal triggering metabolic events responsible for resistance, including avoidance and/or tolerance mechanisms. Unfortunately, the term ‘stress resistance’ is often used in the literature interchangeably with ‘stress tolerance’. This paper highlights the differences between the terms ‘stress tolerance’ and ‘stress resistance’, based on the results of experiments focused on plants’ responses to drought. The ability to avoid or tolerate dehydration is crucial in the resistance to drought at cellular and tissue levels (biological resistance). However, it is not necessarily crucial in crop resistance to drought if we take into account agronomic criteria (agricultural resistance). For the plant user (farmer, grower), resistance to stress means not only the ability to cope with a stress factor, but also the achievement of a stable yield and good quality. Therefore, it is important to recognize both particular plant coping strategies (stress avoidance, stress tolerance) and their influence on the resistance, assessed using well-defined criteria.
1. Introduction
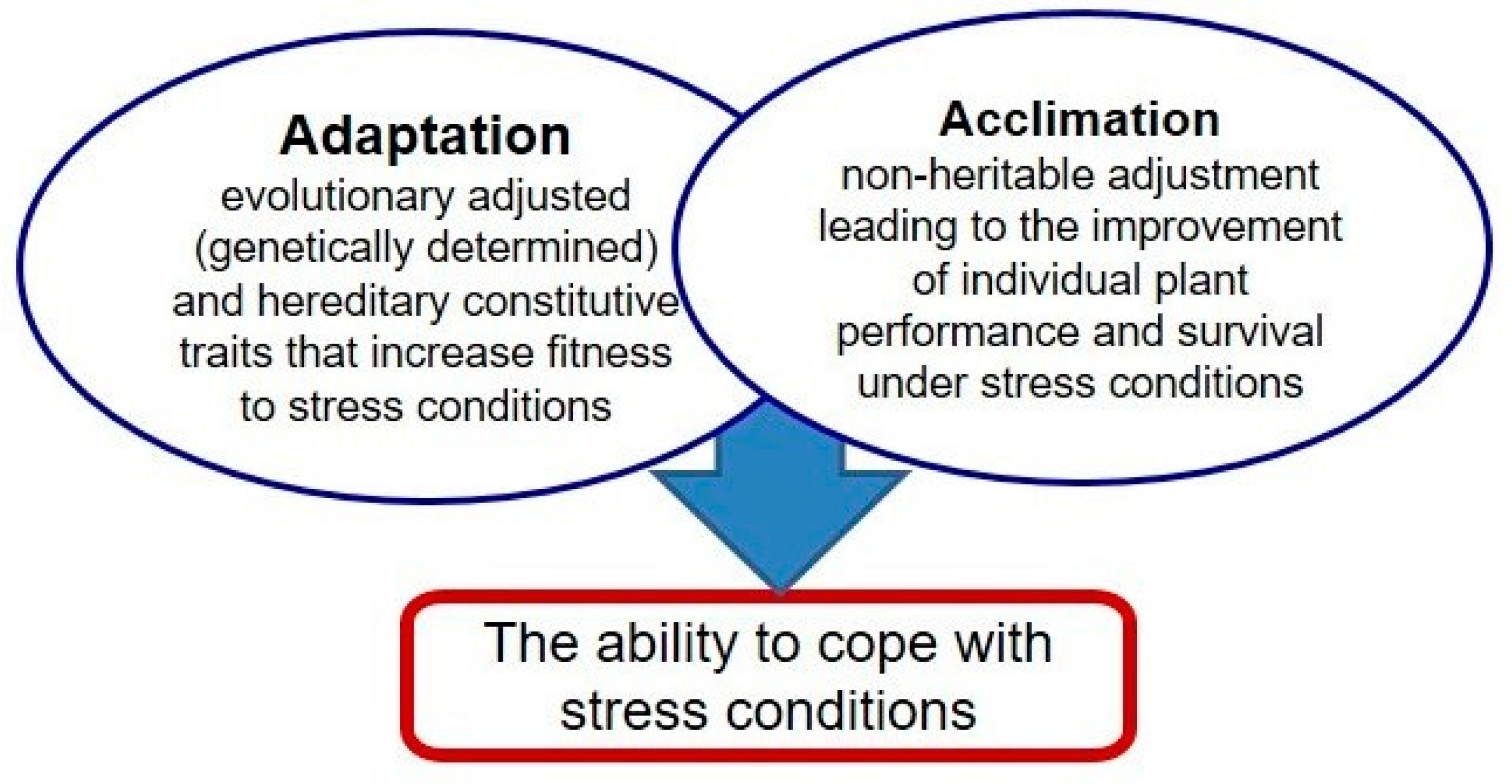
Unfavorable environmental conditions frequently affect plants’ performance, both in natural and agricultural settings. Cramer et al. [1] reported that only 3.5% of the global land area is free from any environmental constraints. Therefore, plants are often exposed to abiotic stress factors which affect their proper development and limit crop production [2,3,4]. Being immobile organisms, plants have been forced to develop specific adaptive traits and ability to adjust (acclimate) to adverse conditions (Figure 1). Adaptation includes developmental, morphological, and physiological traits which help the growth under adverse conditions. Acclimation (hardening) comprises structural, physiological, and biochemical changes responsible for the adjustment to new environmental conditions. It should be distinguished from adaptation, which usually refers to evolutionarily created and genetically determined traits. The ability to acclimate is determined by plant plasticity and includes activation of several complex cellular and molecular responses such as changes in hormone balance and gene expression [5].
The number of papers that focus on the mechanisms of plants response and resistance to stress factors has increased several folds since the beginning of this century [1,6]. It should be highlighted that the understanding of resistance to stress differs depending on the plant’s strategy and plant user’s expectations. Therefore, it is very important to correctly define resistance to stress using clear and appropriate criteria. Stress resistance should not be confused with stress tolerance, which often happens and leads to some misunderstanding which, in my opinion, makes it difficult to define the traits involved in crop resistance to environmental limits. Scientific research focused on identifying the mechanisms or traits responsible for the resistance of crops to stress factors should consider the differences between stress tolerance and stress resistance. The environmental constraint that most often causes the loss of yield throughout the world is drought [2,7]. This paper discusses plant strategies responsible for coping with drought stress and the involvement of the components of these strategies in crop resistance to drought.
2. Concept of Stress and Terminology of Stress Resistance
A stress factor affects the state of stress (strain) in plant cells, which leads to structural or metabolic dysfunctions (growth inhibition, damage of structural and functional proteins, inhibition of enzyme activity) and death, or triggers changes that help the plant to adjust to adverse conditions. The plant response depends on the duration and severity of the stress factor, as well as on genetic traits that determine the ability to cope with stress. Depending on the level of stress and duration, plants can experience the state of eustress or distress (Figure 2). A low dose of stressor causes a slight strain (eustress), which triggers responses that help to cope with harmful conditions. Distress caused by a high dose of stressor rapidly triggers the state of stress in plants, leading to physiological destabilization and death or activation responses that protect against stress damage [8]. Plants’ resistance to stress resulting from either adaptation or acclimation may be the effect of activation of diverse coping strategies including stressor escape, stress avoidance (avoidance of the state of stress in cells), and stress tolerance (tolerance of the state of stress in cells). The strategy of stressor escape (adaptive strategy) relies on the adjustment of the life cycle to the period when plants’ needs are met. It can be observed in drought-sensitive plant species, growing in arid and semi-arid areas with regular water deficit, as well as in early spring plants living in a temperate climate. These plants start to develop at the end of winter (February/March) and complete their life cycle at the beginning of spring. Such a strategy is also observed in perennial plants of a temperate climate, which become dormant at the end of autumn to avoid low winter temperatures. The process of preparing plants to survive winter is autumn leaf senescence, controlled by environmental conditions (light, temperature) which affect the relocation of nitrogen, phosphorus, and other elements from leaves to other organs as well as increased levels of endogenous ABA, responsible for dormancy [9]. Maintaining seed dormancy under harsh conditions, regulated by the interplay between ABA and gibberellins, is also considered a stress escape strategy [10]. Stress avoidance is based on the traits and modifications that prevent the occurrence of the state of stress in plant cells, through retardation or weakening of the action of the stressor—as in, for example, stomatal closure responsible for the restriction of water loss through leaves, as well as osmotic adjustments in plant growing under water deficit conditions [2,11]. Stress tolerance, on the other hand, includes mechanisms responsible for coping with the ongoing state of stress in plant cells, such as the synthesis of compatible substances and proteins, which protects against the negative effect of osmotic and ionic stresses in drought- and salt-stressed plants [11,12]. In other words, it is the capacity to sustain plant functions, thanks to the modifications that counter negative effects of the occurrence of the state of stress, and to repair the damage after stress relief (Figure 2).
Both stress avoidance and stress tolerance are responsible for resistance to stress, understood as the ability to cope with adverse environmental conditions, by keeping a balance between growth, reproduction, and activation of suitable coping strategies [13]. This kind of resistance can be called biological resistance, which is the strategy of an individual plant to tolerate and survive stress conditions. An example of biological resistance is also the stressor escape strategy which occurs in stress-sensitive plants. From the perspective of plant users, crop resistance to environmental stresses should be defined as the ability to cope with stress conditions thanks to defense responses (stress tolerance and/or avoidance) which enables maintenance of stable and good quality yields. Therefore, it can be called agricultural resistance.
3. Plant Responses to Drought
Drought is a meteorological term defined as a period of little or no rainfall, which reduces the amount of water in the soil, and is usually accompanied by high evaporative demand, leading to continuous loss of water by transpiration. It is considered the most frequent climate-related constraint in many regions of the world [3,14]. This stress factor generates the state of stress (strain) in plant cells, which is the reduction in water content (dehydration, water deficit), adversely affecting plant physiological activity, growth, reproduction, and crop productivity [15]. The level of dehydration depends on stress severity and duration, as well as on adaptive traits protecting against water loss (smaller leaves, leaves covered with cuticle or tomentose, as well as leaf folding) and supporting water uptake from deeper soil layers (extensive vertically orientated root system). Another example of an adaptive trait protecting from water loss is stomatal behavior (stomata open at night and closed during day) in crassulacean acid plants (CAM) having an alternative route of carbon assimilation which occurs during the night [5,16]. The above-mentioned adaptations do not usually occur in crop plants, which mostly belong to mesophytes, and are able to grow in an environment with a moderate supply of water. These plants may adjust to water scarcity through the activation of stress avoidance and/or tolerance mechanisms directed at preventing dehydration and/or dehydration damage, and surviving stress [11,13]. A suitable and commonly used marker to evaluate the level of dehydration (state of stress) is relative water content (RWC). In leaves of well-irrigated plants RWC is ≥90%, but with mild drought stress it is in the range 60–70%, with moderate stress 40–60%, and in the case of severe stress it is lower than 40% [17].
Based on the ability to maintain stable leaf hydration under water deficit conditions the water management strategy of plants is classified as isohydric or anisohydric [18]. Isohydric species (‘water savers’) maintain nearly constant RWC through precise control of stomatal behavior. These plants respond to drought by a rapid decrease in stomatal conductance (gs) and restriction of excessive water loss without a reduction in leaf area but at the same time show a decrease in photosynthetic activity. In contrast, anisohydric plants (‘water wasters’) show a decrease in leaf water content and strong leaf area reduction but keep stomata open and maintain a high photosynthetic rate [19,20,21]. The extent of tissue dehydration is a signal triggering, directly or through ABA increase, the activation of appropriate metabolic and physiological changes responsible for plants’ adjustment to drought [22]. Even a slight decrease in RWC triggers upstream signaling events, leading to ABA accumulation and stomatal closure [23]. It was suggested that ABA is primarily synthesized in roots, then it is moved to shoots via xylem vessels and acts as a signal of soil water shortage [24]. Recent research revealed that the source of ABA accumulation in roots under drought stress conditions is its transport from leaves [25,26]. In the model plant Arabidopsis thaliana it was observed that CLE-25 peptide is a root-derived molecule which moves via the vasculature to leaves and transmits a water deficit signal triggering ABA synthesis by the activation of biosynthetic enzymes [27]. The root–shoot signal mediating the effect of soil water deficit on stomata in tomato comprises a dialogue between ABA and strigolactones, ethylene precursor ACC, or sap pH [28]. Stomatal closure in response to soil water deficit in maize and poplar (isohydric species) is regulated by the interaction between the hydraulic signal as a primary message and ABA as a secondary message [18,28,29].
ABA is also involved in several downstream events responsible for the maintenance of tissue hydration (dehydration avoidance strategy), which include osmotic adjustment, comprising the accumulation of organic osmotic compounds (proline, glycine-betaine, soluble proteins, carbohydrates) in leaves and in roots [15,22]. It appears to be necessary for the activation of proline transport and deposition in the root growing region, allowing the maintenance of root growth and undisturbed water uptake under drought conditions [29]. Indeed, ABA plays a central role in plants’ response to drought at different levels of organization; however, it does not act alone but through synergistic or antagonistic crosstalk with other hormones. Crosstalk between ABA, ethylene (ET), and auxin (AUX) regulates root growth and architecture [30]. Guard cells’ aperture and water loss by stomata are controlled by the orchestration of ABA with jasmonic acid (JA), ET, salicylic acid (SA), as well as AUXs and cytokinins (CKs). Increased levels of JA, ET, and SA in drought-stressed plants promote the induction of stomatal closure which is maintained by the decline in CK and AUX levels [31,32,33,34]. Foliage-derived ABA promotes root growth under drought by lowering the level of ET, which is a root growth inhibitor [35]. Additionally, auxin-induced alteration of root architecture, which leads to the creation of more vertical and deeper roots, plays an important role in maintaining better water acquisition under drought conditions [27,36]. Brassinosteroid (BR)-induced root hydrotropism and accumulation of osmoprotectants (proline, trehalose, raffinose) in roots may also improve water uptake under drought [27,37]. The beneficial effect of ABA on water transport and tissue hydration under drought conditions may also be achieved by its influence on the improvement of root hydraulic conductivity through regulation of the activity of membrane water channels—aquaporins [38,39].
The strategy of dehydration avoidance (isohydric behavior) allows plants to sustain physiological functions under stress conditions and recover after stress termination. This strategy is effective in plants exposed to mild or moderate drought that does not last very long but under prolongated drought it affects carbon starvation [40]. Moreover, when stomata are closed plants absorb more light than can be used in carbon fixation, which triggers generation of reactive oxygen species (ROS), affecting secondary stress and damage of PSII, leading to further weakness of photosynthesis [12,41]. What is more, during long-term drought the ability of plants to maintain stomatal closure may be weakened due to a decrease in ABA level and plant behavior changes to anisohydric [42]. The response to drought in anisohydric plants (barley, wheat, sunflower) is mainly regulated hydraulically. The maintenance of stomatal conductance in these plants is supported by the capacity for osmotic adjustment, controlled by the dehydration signal, which enables plants to extract water from soil to maintain tissue hydration [18,40]. In anisohydric wheat genotypes the level of ABA in leaves did not change under water deficit conditions, while in roots it increased but only after 21 days of stress [29]. Therefore, it is possible that, along with tissue dehydration, ABA may also play a role in the response of anisohydric species to prolonged drought. The stomatal conductance in anisohydric plants is also maintained by undisturbed water movement through cell membrane aquaporins responsible for roots’ ability to conduct water [43]. It was reported that ABA increases the activity of aquaporins and improves root hydraulic conductivity [44]. The activity of aquaporins is also regulated by gibberellins (GAs), CKs, methyl jasmonate (MeJA), and AUXs at transcriptional and post-transcriptional levels ([45] and references therein). The anisohydric strategy is beneficial under mild to moderate drought conditions but may be a risk under severe and long-term stress, which may cause hydraulic failure and severe dehydration [20].
In plants exposed to severe and long-term drought, dehydration cannot be avoided, and activation of dehydration tolerance mechanisms becomes important. Dehydration has a deleterious effect on cell membranes and causes the disruption of many biochemical and physiological processes [2,46]. A frequently used indicator of dehydration tolerance is the cell membrane injury index or membrane stability index, which shows the ability to maintain membrane integrity at a given level of dehydration [47,48]. The dehydration tolerance mechanisms enable plants to maintain membrane integrity and cell homeostasis, and to regain physiological activity after stress cessation [12,41]. These mechanisms are controlled by ABA-dependent and -independent pathways and include synthesis of protective proteins (LEA proteins, dehydrins, chaperons) and compatible compounds (proline, glycine-betaine, proline-betaine, trehalose, raffinose mannitol, pinitol) involved in enzyme and membrane protection [2,22,41,49,50]. Dehydration-induced disturbance of the respiratory metabolic pathway exhibits generation of ROS, leading to a state of oxidative stress [2,46,51,52,53]. Moreover, in drought-stressed plants the enhanced build-up of ROS is caused by photosynthesis disruption and increased photorespiration due to the limitation of CO2 uptake [53,54]. Overproduction of ROS (secondary stress), which includes superoxide radicals (O2•−), hydrogen peroxide (H2O2), hydroxyl radical (OH•), and singlet oxygen (1O2), is harmful to organelles through lipid peroxidation and damage to nucleic acids and proteins [2,3,46]. In order to overcome oxidative damage, plants possess enzymatic and non-enzymatic ROS-scavenging systems. Enzymatic antioxidants include superoxide dismutase (SOD), catalase (CAT), and peroxidases (POX). The non-enzymatic components of the antioxidative system comprise ascorbic acid, α-tocopherol, flavonoid, glutathione, carotenoids, proline, and phenolic compounds which mitigate oxidative damage by direct reduction of ROS activity and by working together with antioxidant enzymes [53,55]. Additionally, alternative oxidase (AOX) is involved in avoidance of excess generation of ROS in mitochondrial electron transport chains [54]. ABA plays a pivotal role in the activation of antioxidant enzymes and synthesis of low molecular ROS scavengers [49,50]. Upregulation of the antioxidant system may also be controlled by JA, SA, and BRs [34,51,52,53,54,55,56,57,58].
Thanks to the efficient antioxidative system, plants can keep ROS at non-toxic levels, and these molecules are thought to act as signals for activation of stress defense responses [45,54]. It was also evidenced that NADPH oxidase localized in apoplastic fluid is involved in ROS production for integrating signaling networks involved in stress response processes. An increased level of this enzyme was detected in drought-stressed rice as well as in leaves of ABA- and Ca+-treated maize seedlings [54,59]. Moreover, NADPH oxidase regulates H2O2 production for the signaling cascade which affects ABA-dependent stomatal closure and antioxidant defense. The involvement of NADPH oxidase in brassinosteroid-induced H2O2 production and regulation of stomatal closure/opening and antioxidant defense was also reported [54,60].
Plant responses to drought are governed by a sophisticated regulatory system working at the molecular level. The decrease in turgor pressure leads to tension changes in plasma membranes, which are perceived by membrane proteins including receptor-like kinases (RLKs), histidine kinases (HKs), and integrin-like proteins (ILPs) working as osmotic stress sensors. ATHK1 is an Arabidopsis thaliana His kinase postulated to play a role in water stress perception triggering the mitogen-activated protein kinase (MAPK) signaling cascade both in ABA-dependent and ABA-independent regulatory systems [61]. A crucial role in the signal transduction route is played by transcription factors (TFs) that bind to TF binding sites (TFBS) in the promotor region and regulate gene expression. TF families involved in plants’ response to drought include bZip (AREB/ABF), AP2/ERF (DREB/CBF), MYB/MYC, WRKY, and NAC [3,62]. In the ABA-dependent pathway the perception of ABA by receptor proteins is the primary event that triggers downstream signaling cascades to induce final physiological responses. The receptors for this hormone are small soluble cytosol/nucleus-localized pyrabactin resistance (PYR)/PYR-like (PYL)/regulatory components of ABA receptor (RCAR) proteins. The interaction of ABA with PYR/PYL/RCARs affects deactivation of protein phosphatase enzymes (PP2Cs), which are constitutive negative regulators of ABA-induced responses. The inhibition of PP2Cs leads to auto-phosphorylation of the protein kinases SnRK2s, which induces stomatal closure and stimulates nuclear targets that trigger expression of various water stress associated genes due to activation of TFs [62]. ABA-dependent gene expression systems involve activation of b-ZIP (AREBs/ABFs), MYC/MYB, as well as NAC transcription factors [63].
In ABA-independent responses to drought the dehydration signal from the cell surface to the nucleus is mediated by calcium, JA, and ROS [62]. Water deficit leads to membrane destabilization and Ca2+ influx into the cytoplasm. The calcium signal is detected and transduced through calmodulin (CaM), calcium dependent protein kinases (CDPK), and calcineurin B-like proteins (CBLs) and interacts with the MAPK cascade, leading to activation of TFs (DREB, NAC) and expression of genes coding the synthesis of functional proteins (LEA proteins, chaperones, dehydrins, enzymes of osmolyte biosynthesis). JA, on the other hand, is engaged in activation of the MYC2 transcription factor, which triggers expression of stress-responsive genes [62]. Furthermore, JA along with ROS acts as a stress-signaling unit triggering the expression of genes involved in activation of enzymatic and non-enzymatic scavenging events [62,64]. The widespread plant response to drought is proline accumulation due to the stimulation of its synthesis from glutamate catalyzed by pyrroline-5-carboxylate synthetase (P5CS) and pyrolino-5-carboxylate reductase (P5CR) [65,66]. Synthesis of this amino acid under drought is driven by both ABA-dependent and ABA-independent signaling pathways engaged in triggering expression of P5CS and P5CR genes regulated by many TFs, which are also related to responses to drought controlled by other growth regulators [67].
Important components of the stress-factor-induced regulatory system are epigenetic modifications which are independent of DNA sequence changes. These changes include chromatin remodeling such as DNA methylation and histone modifications altering the structure and accessibility of chromatin, leading to changes in gene expression at the transcriptional and post-transcriptional levels [68]. Drought-stress-induced changes in DNA methylation have been observed in diverse plant species. These changes were related to the expression of genes encoding transcription factors and were involved in drought resistance mechanisms or were linked to drought sensitivity [69,70,71]. It was found that changes in DNA methylation (demethylation) in water deficit stressed rice were responsible for proline accumulation via the upregulation of proline metabolism-related gene expression [72]. In addition to DNA methylation, drought-induced histone modifications (methylation, acetylation) are involved in controlling gene expression in stressed plants [73]. It was observed that drought stress triggered histone H3 lisyne4 tri-methylation (H3K4 me3) in the gene body region of nine cis-epoxycarotenoid dioxygenase 3 (NCED3), which is a key enzyme involved in ABA synthesis. Additionally, some studies reported the increase in H3K4me3 and H3 lisyne9 acetylation (H3K9Ac) in the promotor region of such genes as RD29A, RD29B, RD22, and RELATED TO AP2.4 (RAP2.4) encoding synthesis of LEA proteins. The abundance of histone modification and the number of genes expressed depend on stress duration and degree [70,73]. Most of the epigenetic modifications are removed when the stress is relived, but some of them persist, enabling plants to remember past stress and to prepare for future recurrent stress events which occur during plant life. This is so-called “plant stress memory”, which can also be transferred to further generations during sexual and vegetative reproduction [69,71,74]. Integral components of the stress response at the molecular level also involved in memory pathways are non-coding small RNAs (miRNAs, siRNAs), which can trigger DNA methylation and histone modifications. Plants exposed to drought can memorize stress events through DNA and histone modifications for specific gene expression thanks to up- and downregulation of small RNAs responsible for the increased resistance to future stress events through the control of TFs, ROS, and hormone levels [71,74].
4. Drought Coping Strategies and Resistance
The ability to avoid or tolerate dehydration is crucial in dealing with drought at cellular and tissue levels (biological resistance), which allows plants to survive during water scarcity conditions and recovery. The tolerance and avoidance mechanisms were developed during evolution in order to adjust to environmental conditions but usually do not have beneficial effects in agricultural production. Plants can withstand drought without any visible signs of dehydration and/or dehydration damage, but their growth and yield may be lower than expected. This is an unwanted side effect of plant adjustment to stress, which has a negative impact on biomass accumulation and yield (agricultural resistance).
The activation of coping mechanisms is connected to increased energy and nutrient consumption, which results in the allocation of less energy and assimilates to growth processes, leading to yield reduction [15,75]. Furthermore, many traits associated with drought resistance have a dual effect (positive or negative) on plant productivity which depends on stress intensity and timing as well as on climatic conditions such as light intensity and evaporative demand [76]. The dehydration avoidance strategy, such as stomatal closure, reduces water loss from leaves. However, at the same time it causes the restriction of CO2 uptake, ROS generation, damage of PSII, and the inhibition of photosynthesis, resulting in the reduction of crop production [12,41,77,78,79]. Moreover, changes in the hormonal balance, which is a part of the coping strategy consisting of an increase in the levels of ABA, JA, Et, and SA, and decrease in CKs, AUXs, and GAs, may also bring about photosynthesis inhibition, growth restriction, leaf senescence acceleration, and leaf fall, negatively affecting yield [32,34,80]. Therefore, there is a conflict between plant coping strategies (avoidance, tolerance) and resistance to drought essential for agricultural production. In the agricultural perspective, drought-resistant plants are those that maintain growth and stable yield during water-limited conditions. The priority in breeding research focusing on improving drought resistance is to obtain crop genotypes that can cope with drought stress without growth and yield reduction. Therefore, the research on plant stress physiology should concentrate on finding those features of coping strategies that ensure growth maintenance and stable yield (Table 1).
Many genes and processes involved in plants’ ability to cope and survive drought (biological resistance) in experiments conducted under laboratory conditions have been identified. However, the knowledge about their real function in the resistance to this stress, based on well-defined agronomic criteria (agricultural resistance), is rather poor [11,22,81,82]. It is hard to show the involvement of a particular trait or adjustment to drought in maintaining yield potential in a short-term experiment. The response to short-term drought conditions in soil pot experiments (limited rhizosphere) did not reflect the response to long-term water shortage in the field [83]. Drought resistance is the result of combined processes that happen on different timescales and have a long-term impact on plant performance and yield. Short-term responses to drought include triggering physiological feedback processes responsible for stabilizing plant water and carbon status, which are often not correlated with the long-term effect. The favorable effects of these feedback strategies on yield depend on the drought scenario as well as on scalability and phenotypic distances between traits involved in particular coping strategies and those responsible for yield [28]. It is necessary to search for processes and adjustments that allow crops to continue to grow under water-limited conditions and rapid recovery after stress termination without yield reduction.
The source of traits valuable in developing new drought-resilient crop varieties may be wild genotypes and landraces originating from rainfed areas [3]. Another promising approach is the introduction of new crop species able to cope under water-limited conditions and maintain stable growth. An interesting species in this regard is quinoa (Chenopodium quinoa Willd.), which originated in the Andean region. It has begun to be called ‘the 21st century crop’, and recently it has been introduced into cultivation in many regions of the world. Quinoa has received special attention due to its high nutritional composition of seeds and strong natural ability to cope with drought [84,85]. There is wide diversity among quinoa genotypes in the traits of drought coping strategy (biological resistance) and resistance assessed based on the seed yields (agricultural resistance). The drought response mechanisms in quinoa to 8endure water deficits include accelerated root growth, high water-use efficiency (WUE), osmotic adjustment, turgor maintenance, increased synthesis of osmoprotectants such as amino acid proline, and soluble sugars (glucose, trehalose), ABA biosynthesis, antioxidant defense, heat-shock, and LEA protein synthesis [86]. Field studies have shown no significant yield reduction in the Danish quinoa cultivar Titicaca under water deficit conditions [84]. Soil pot experiments revealed that the capacity for growth in a drought-prone environment in ‘Titicaca’ was associated with the increase in WUE due to higher ABA concentration and nutrient content [87]. Recent studies revealed that drought resistance in quinoa var. Red Faro was due to elevated recovery capacities of PSII and PSI photochemical activities after re-watering [88]. There are numerous studies focused on molecular, biochemical, physiological, and morphological responses of varied quinoa genotypes to drought both under laboratory and field conditions. The sequencing of the quinoa genome creates the possibility of using new molecular tools to fully discover regulatory mechanisms involved in drought resistance of various quinoa genotypes [86].
A promising drought resistance strategy for crops is the ability to optimize water use, along with sustained high photosynthetic activity, which is an essential component of plant productivity [89]. It may be achieved by triggering varied metabolic and physiological responses of the dehydration avoidance strategy, which includes the modification of root conductivity and architecture, regulation of stomatal behavior allowing the maintenance of photosynthetic CO2 fixation, as well as protection against non-stomatal photosynthesis limitation [90,91,92]. Plants with greater WUE assimilate more carbon per unit of transpired water. These plants are less susceptible to drought as they take less water from the soil and may access this water later in the season when a lack of water has become a limiting factor [79]. The improvement in WUE under water-limited conditions without trade-offs in carbon assimilation was revealed in transgenic tomato with overexpression of the gene encoding ABA biosynthesis enzyme (NCED3) as well as in Arabidopsis overexpressing ABA receptors [38,93]. The effect of increased ABA levels in roots and leaves of drought-stressed tomato lines was lower stomatal conductance and greater root conductivity [38]. However, ABA signaling-mediated changes in Arabidopsis transgenic lines affected reduced stomatal conductance, which was compensated by increased CO2 gradients across stomata, allowing maintenance of a CO2 influx [93]. These findings in Arabidopsis are being considered for translation to cereal crops to obtain drought-resistant genotypes through improving WUE [94,95]. A suitable criterion to measure WUE is carbon isotope discrimination (Δ13C), which is used in breeding programs to select drought-resistant crop genotypes [76]. A significant positive relationship between Δ13C and yield was revealed in drought-stressed quinoa cultivars under field conditions [86,96]. However, Tardieu [76] considers that Δ13C is a positive trait for yield under severe water deficit conditions but under mild to medium drought the positive traits that optimize yield are high stomatal conductance and growth maintenance.
Multiple biochemical and physiological changes that are components of drought coping strategies were revealed to have a favorable effect on yield (Table 1). Lower yield reduction under drought conditions was observed in a wheat cultivar that exhibited osmotic adjustment resulting from the accumulation of soluble sugars and proline as well as increased activity of enzymatic and non-enzymatic antioxidants. These changes allow for the maintenance of high photosynthetic CO2 fixation during drought and rapid recovery after re-watering, which are responsible for the final productivity [97]. Barley genotypes that yielded better under drought conditions exhibited increased expression of 34 genes which are involved in stress signaling, carbon metabolism, control of stomatal closure, proline synthesis, activation of the ROS scavenging system, and protective protein synthesis [82]. Elevated osmotic adjustment, increased expression of dehydrin genes, and a significant increase in alpha-tocopherol, which plays an important protective role for PSII, along with a higher photosynthetic rate, were observed in barley genotype, characterized by a smaller decrease in the performance index under drought stress conditions [98]. A large body of evidence has shown a beneficial role of proline in dealing with drought stress (biological resistance). Proline, involved in osmotic adjustment, is a free radical scavenger and acts as a compound that protects enzymes, proteins, and cell membranes against detrimental effects of dehydration and oxidative stress ([66,99] and references therein). It also serves as a carbon and nitrogen reserve after stress relief, and may act as a signaling molecule, able to activate defense responses [100]. Therefore, rapid proline accumulation at the beginning of drought stress may play an essential role in the dehydration avoidance strategy. Its increased level may also protect plants from the detrimental effect of dehydration (dehydration tolerance strategy), and it may be involved in the ability to recover after stress cessation. However, the involvement of this amino acid in the resistance to drought, understood as an adjustment without any negative effects on yield, is still not clear. The possible beneficial effect of greater leaf proline accumulation under drought on agricultural resistance, based on grain yield, was found in wheat [101]. Interesting results were obtained by Frimpong et al. [102], who observed that introgression barley lines, harboring a pyrroline-5-carboxylate synthase (P5cs1) allele, had markedly higher proline content in spikes and leaves, compared with other genotypes. These lines also showed milder drought symptoms, were able to maintain a high photosynthetic rate under drought, and achieved higher final seed production. Moreover, the barley near-isogenic line NIL 143, characterized by higher leaf and root proline content, showed less severe symptoms of drought, higher leaf water content, better stomatal conductance and net CO2 assimilation than other genotypes. This barley line also exhibited increased lateral root growth, probably due to high proline accumulation [103]. Considerable evidence obtained previously revealed that drought-stress-induced expression of proline biosynthetic genes is regulated by TFs related to almost all plant hormones [67].
One recently considered approach in attaining crop resistance to drought is focused on better understanding of the role of plant growth regulators (PGRs) in the coping strategy along with the mitigation of the negative effect of drought on productivity and yield. PGRs play an important role in triggering, directly or through specific signal cascades, a wide range of metabolic and physiological responses of plants to drought. Many of these responses, which are components of the drought stress coping strategy, are the result of positive or negative interactions between diverse PGRs [31,32,104]. Broadening knowledge about the impact of drought on the fluctuation of the level of PGRs and about the crosstalk between them in triggering appropriate responses seems to be essential in identifying components of drought coping strategies, which permit undisturbed growth and stable yield. The hormone that plays a key role in the plant response to drought is ABA, commonly called a “stress hormone”. An increased level of this PGR in drought-stressed plants acts as a signal that regulates multiple responses at physiological and biochemical levels [10,50]. It was suggested that the interaction between plant hormones (ABA, AUXs, CKs, and ET) may play an important role in a diverse drought response of sensitive and resistant wheat lines [105]. The resistant wheat line was able to maintain growth and was characterized by lower yield reduction under drought. This line was temporarily anisohydric and closed the stomata only at a higher level of drought which correlated with the repression of ABA synthesis. At the same time, it had the ability to activate defense responses (ROS protection, LEA proteins, and cuticle synthesis) and to trigger expression of photosynthesis genes as well as genes involved in AUXs, CKs and Et metabolism, and signaling. However, the drought-sensitive wheat line was isohydric, had a higher ABA level, closed stomata at the start of stress and began photosynthesis inhibition. Certain recently obtained results of research focused on crosstalk between ABA, CKs, and BRs at physiological and molecular levels seem to be promising in finding drought coping strategies that prevent yield reduction [27,33]. ABA increase and the reduction in CK level under drought lead to a decrease in stomatal aperture and density, as well as accelerated leaf senescence, along with photosynthesis inhibition [32,106]. The manipulation of endogenous CK level and control of CK signaling pathway components in transgenic rice were effective in restoration of stomatal conductivity, reduction in leaf senescence, and amelioration of yield losses [32]. This transgenic rice also displayed increased expression of BR-related genes and repression of JA-related genes [107]. It was reported that BRs trigger the expression of various stress-related genes important in the maintenance of photosynthetic activity, stimulation of the antioxidant system, and accumulation of osmoprotectants [32,107,108]. Furthermore, overexpression of the BR receptor (BRL3) leads to activation the synthesis of osmoprotectants (i.e., proline, trehalose, sucrose) in roots and overcoming growth arrest as well as modulating the root hydrotropic response during drought [27,37,104]. An interesting and promising mechanism leading to drought resistance appears to be the involvement of BRs in the expression of cell wall extension and release of enzymes, which lead to increased cell expansion [108]. The last several years of research have shown that crosstalk between BRs and other hormones is involved in the network of complex regulatory responses to drought, including stress perception and signaling leading to activation of various coping strategies [109]. Master regulators of abiotic stress responses whose expression is controlled by hormonal balance and crosstalk are TFs [110]. Gaining knowledge about the pattern of appropriate hormonal balance and crosstalk as well as identification of stress-responsive TFs and their role in activation of the components of the drought coping strategy without yield mortality is a powerful approach for achieving drought-resistant crop cultivars [3,62,111].

{kind=link}
{kind=link}
Table 1.
Components of coping strategies and agricultural resistance in crops and model plants.
| Plant Species/Genotypes | Stress Imposition Stress Level | Components of Coping Strategy | Agricultural Resistance | References |
|---|---|---|---|---|
| Arabidopsis thaliana transgenic line RCAR10-4 | soil pot experiment water withholding 8 weeks severe stress RWC—not performed | increased expression of ABA receptor reduced stomatal conductance with maintenance of carbon assimilation | improvement in WUE and growth | [93] |
| tomato transgenic line sp12 | soil pot experiment water withholding at four- or five-leaf stage 5 days RWC—not performed | overexpression of ABA biosynthesis of gene (NCED) increased ABA level in root and leaves reduced stomatal conductance increased root hydraulic conductivity, water status improvement | improvement in WUE without trade-offs in carbon assimilation | [38] |
| wheat ‘Luhan7′ | soil pot experiment, irrigation withheld at tillering and jointing stage 10 days moderate stress RWC 85–89% | osmotic adjustment (proline, sugars) stomata closure activation of antioxidant system | high photosynthetic CO2 fixation high drought index and harvest index | [97] |
| barley H. vulgare ‘Martin’ H. spontaneum HS41-1 | soil pot experiment, water withholding at flowering stage 13 days severe stress RWC—not performed | high expression of signal transduction genes (TFs, CDPK, membrane binding proteins) and functional genes directly involved in coping strategy (stomatal behavior, synthesis of glycine-betaine, proline, antioxidants, dehydrins) | higher chlorophyll content and lower grain yield losses than in genotype without enhanced expression of coping strategy genes | [82] |
| barley ‘Yousof’ and ‘Morocco’ | soil pot experiment water withholding at two weeks seedling stress duration? mild stress RWC ~88% | high level of dehydrin and alpha-tocopherol involved in PSII protection in ‘Yousof’ | lower reduction in CO2 assimilation rate and performance index in ‘Yousof’ | [98] |
| transgenic rice | soil pot experiment water withholding at pre-anthesis and post-anthesis 6–10 days mild stress RWC ~85% | increased CK synthesis, increased expression of BR related genes and repression of JA-related genes modification of source/sink relationships, a stronger sink capacity | higher grain yield with improved quality (nutrients and starch content) | [107] |
| wheat ‘Zagros’ and ‘Marvdaht’ | soil pot experiment soil moisture at about 50% of field capacity 31 days RWC—not performed | higher ABA and proline accumulation in ‘Zagros’ than ‘Marvdaht’ | higher harvest index and lower grain yield reduction in ‘Zagros’ than ‘Marvdaht’ | [101] |
| barley introgression lines with wild allele p5cs1- S42IL-141, S42IL-141 | soil pot experiment reduction in irrigation at booting stage mild stress 15-day RWC ~83% | significantly higher spike and leaf proline level than other line | maintenance of high photosynthetic rate and inherent WUE, high final seed productivity | [102] |
| barley near-isogenic line with wild allele P5cs1- NIL 143 | rhizoboxes filled with soil soil water content decreased from 40% at the beginning to 6% after 17 days (three-leaf seedling) severe stress RWC ~59% | higher root and shoot proline content than in other genotypes, less severe drought symptoms, better stomatal conductance, higher RWC, enhanced root growth | enhanced net assimilation rate | [103] |
| quinoa ‘Titicaca’ | field experiment soil pot experiment mild to severe RWC—not performed | ABA increase, high WUE | no yield reduction | [84,87] |
| quinoa 10 varieties | field experiment RWC—not performed | high carbon isotope discrimination | high yield | [96] |
| wheat drought-tolerant ‘Halberd’ drought-sensitive ‘Cranbrook’ | soil pot experiment growth chamber water withholding drought stress at the young microspore stage RWC—not performed | ABA increase, stomatal closure at the start of stress, inhibition of photosynthesis in ‘Halberd’ delayed stomatal closure and activation of defense response repression of ABA synthesis enrichment of genes involved in AUX, CK and ET metabolism/signaling in ‘Cranbrook’ | lower yield reduction in ‘Cranbrook’ | [105] |
| transgenic Oryza sativa cotton | soil pot experiment in greenhouse and growth chamber irrigation reduction RWC—not performed | increased CK level modifications of source/sink relationships delayed senescence increased expression of BR-related genes | improved grain yield and grain quality improved photosynthesis, biomass accumulation | [106,107] |
5. Conclusions
Drought is the most frequent abiotic stress adversely affecting productivity of crop plants. As sessile organisms, plants have developed sophisticated regulatory mechanisms at molecular and physiological levels to cope with water scarcity conditions. These mechanisms are important for stress survival (biological resistance). However, activation of these mechanisms frequently does not prevent the negative effect of drought on growth and yields (agricultural resistance), which is important for plant users. Therefore, there is a need for continuous and extensive research expanding the knowledge required in breeding drought-resistant and high-yielding crop genotypes. The modern breeding technologies and biotechnological approaches aimed at developing crops resilient to drought with a high yield capacity should include genomic, molecular, and physiological research. It is crucial to determine the relationships between molecular, metabolic, and physiological changes involved in biological resistance to drought and agricultural resistance estimated using agronomic criteria (photosynthetic activity, growth traits, yield). At the molecular level, research based on marker-associated selection, genome-wide association studies, and genome selection with high throughput phenotyping are useful in identifying candidate genes and TFs effective for improving the resistance of crops to drought [3,28,104,112]. Currently used approaches to obtain drought resistance crops include the use of: (a) traditional breeding programs; (b) genetically modified plants; and (c) clustered regularly interspaced short palindromic repeats (CRISPR/Cas) editing strategy [4,113]. Presently, the new strategy with possible future application is the selection of epigenetic phenotypes with increased drought resistance [71].
A slightly different, non-genetic, approach for improving crop resistance to drought is the exogenous application of natural substances, including plant metabolites ([113] and references therein). The favorable effect of such metabolites on biological resistance has been demonstrated. It is reported in the literature that many of these metabolites are also involved in crop yield improvement under drought (agricultural resistance). It is a strategy that is easy and feasible to implement. However, the beneficial effects of application of this strategy depend on the concentration of the used compound, time of application, and crop species. The use of these metabolites by producers should be preceded by long-term experiments under field conditions in order to evaluate the dose, method, and time of application in different plant species as well as the cost of application in the field.
Funding
This publication was co-financed within the framework of the Polish Ministry of Science and Higher Education’s program: “Regional Initiative Excellence” in the years 2019–2022 (No.005/RID/2018/19).
Conflicts of Interest
The author declare no conflict of interest.
References
- Cramer, G.R.; Urano, K.; Delrot, S.; Pezzotti, M.; Shinozaki, K. Effects of abiotic stress on plants: A systems biology perspective. BMC Plant Biol. 2011, 11, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of climate change on crops adaptation and strategies to tackle its outcome: A Review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imran, Q.M.; Falak, N.; Hussain, A.; Mun, B.-G.; Yun, B.-W. Abiotic stress in plants; stress perception to molecular response and role of biotechnological tools in stress resistance. Agronomy 2021, 11, 1579. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin III, F.S.; Pons, T.L. Plant Physiological Ecology, 2nd ed.; Springer Science + Busines Media, LLC: New York, NY, USA, 2008; ISBN 978-0-387-78340-6. [Google Scholar]
- Nuccio, M.; Paul, M.; Bate, N.J.; Cohn, J.; Cutler, S.R. Where are the drought tolerant crops? An assessment of more than two decades of plant biotechnology effort in crop improvement. Plant Sci. 2018, 273, 110–119. [Google Scholar] [CrossRef]
- Yang, S.J.; Vanderbeld, B.; Wan, J.X.; Huang, Y.F. Narrowing down the targets: Towards successful genetic engineering of drought tolerant crops. Mol. Plant 2010, 3, 469–490. [Google Scholar] [CrossRef]
- Hideg, E.; Jansen, M.A.K.; Strid, A. UV-B exposure, ROS, and stress: Inseparable companions or loosely linked associates? Trends Plant Sci. 2013, 18, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Keskitalo, J.; Bergquist, G.; Gardeström, P.; Jansson, S. A cellular timetable of autumn senescence. Plant Physiol. 2005, 139, 1635–1648. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.; Rabara, R. Abscisic acid—An enigma in the abiotic stress tolerance of crop plants. Plant Gene 2017, 11, 90–98. [Google Scholar] [CrossRef]
- Dodd, I.C.; Ryan, A.C. Whole-plant physiological responses to water-deficit stress. In eLS; John Wiley & Sons, Ltd.: Chichester, UK, 2016. [Google Scholar] [CrossRef]
- Dinakar, C.; Djilianov, D.; Bartels, D. Photosynthesis in desiccation tolerant plants: Energy metabolism and antioxidative stress defense. Plant Sci. 2012, 182, 29–41. [Google Scholar] [CrossRef]
- Volaire, F. A unified framework of plant adaptive strategies to drought: Crossing scales and disciplines. Glob. Change Biol. 2018, 24, 2929–2938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, O. Agricultural extreme drought assessment at global level using FAO—Agricultural stress index system (ASIS). Weather Clim. Extrem. 2020, 27, 100184. [Google Scholar] [CrossRef]
- Claeys, H.; Inzé, D. The agony of choice: How plants balance growth and survival under water-limiting conditions. Plant Physiol. 2013, 162, 1768–1779. [Google Scholar] [CrossRef] [PubMed]
- Taiz, L.; Zeiger, E. Plant Physiology, 4th ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2010; ISBN 0-87893-856-7. [Google Scholar]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.-J. The role of the plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Tardieu, F.; Simonneau, T. Variability among species of stomatal control under fluctuating soil water status and evaporative demand: Modelling isohydric and anisohydric behaviours. J. Exp. Bot. 1998, 49, 419–432. [Google Scholar] [CrossRef] [Green Version]
- Maseda, P.H.; Fernández, R.J. Stay wet or else: Three ways in which plants can adjust hydraulically to their environment. J. Exp. Bot. 2006, 57, 3963–3977. [Google Scholar] [CrossRef]
- Sade, N.; Gebremedhin, A.; Moshelion, M. Risk-taking plants: Anisohydric behavior as a stress-resistance trait. Plant Signal. Behav. 2012, 7, 767–770. [Google Scholar] [CrossRef] [Green Version]
- Skelton, R.P.; West, A.G.; Dawson, T.E. Predicting plant vulnerability to drought in biodiverse regions using functional traits. Proc. Natl. Acad. Sci. USA 2015, 112, 5744–5749. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, F.; Kuromori, T.; Sato, H.; Shinozaki, K. Regulatory gene networks in drought stress responses and resistance in plants. In Survival Strategies in Extreme Cold and Desiccation. Advances in Experimental Medicine and Biology; Iwaya-Inoue, M., Sakurai, M., Uemura, M., Eds.; Springer: Singapore, 2018; Volume 1081. [Google Scholar]
- Verslues, P.E.; Zhu, J.-K. Before and beyond ABA: Upstream sensing and internal signals that determine ABA accumulation and response under abiotic stress. Biochem. Soc. Trans. 2005, 33, 375–379. [Google Scholar] [CrossRef] [Green Version]
- Schachtman, D.P.; Goodger, J.Q.D. Chemical root to shoot signaling under drought. Trends Plant Sci. 2008, 13, 281–287. [Google Scholar] [CrossRef]
- McAdam, S.A.N.; Brodribb, T.J.; Ross, J.J. Shoot derived abscisic acid promotes root growth. Plant Cell Environ. 2016, 39, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Castro, P.; Puértolas, J.; Dodd, I. Stem girdling uncouples soybean stomatal conductance from leaf water potential by enhancing leaf xylem ABA concentration. Environ. Exp. Bot. 2018, 159, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, F.; Simonneau, T.; Muller, B. The physiological basis of drought tolerance in crop plants: A scenario-dependent probabilistic approach. Annu. Rev. Plant Biol. 2018, 69, 733–759. [Google Scholar] [CrossRef] [Green Version]
- Gallé, Á.; Csiszár, J.; Benyó, D.; Laskay, G.; Leviczky, T.; Erdei, L.; Tari, I. Isohydric and anisohydric strategies of wheat genotypes under osmotic stress: Biosynthesis and function of ABA in stress responses. J. Plant Physiol. 2013, 170, 1389–1399. [Google Scholar] [CrossRef] [Green Version]
- Sharp, R.E.; Poroyko, V.; Hejlek, L.G.; Spollen, W.G.; Springer, G.K.; Bohnert, H.J.; Nguyen, H.T. Root growth maintenance during water deficits: Physiology to functional genomics. J. Exp. Bot. 2004, 55, 2343–2351. [Google Scholar] [CrossRef] [Green Version]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress respopnses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [Green Version]
- Peleg, Z.; Blumwald, E. Hormone balance and abiotic stress tolerance in crop plants. Curr. Opin. Plant Biol. 2011, 14, 290–295. [Google Scholar] [CrossRef]
- Hai, N.N.; Chuong, N.N.; Tu, N.H.; Kisiala, A.; Hoang, X.L.T.; Thao, N.P. Role and regulation of cytokinins in plant response to drought stress. Plants 2020, 9, 422. [Google Scholar] [CrossRef] [Green Version]
- Ciura, J.; Kruk, J. Phytohormones as targets for improving plant productivity and stress tolerance. J. Plant Physiol. 2018, 229, 32–40. [Google Scholar] [CrossRef]
- Spollen, W.G.; LeNoble, M.E.; Samuels, T.D.; Bernstein, N.; Sharp, R.E. Abscisic acid accumulation maintains maize primary root elongations at low water potentials by restricting ethylene production. Plant Physiol. 2000, 122, 967–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogura, T.; Goeschl, C.; Filiault, D.; Mirea, M.; Slovak, R.; Wolhrab, B.; Satbahai, S.B.; Busch, W. Root system depth in Arabidopsis is shaped by EXOCYST70A3 via the dynamic modulation of auxin transport. Cell 2019, 178, 400–412. [Google Scholar] [CrossRef] [PubMed]
- Fàbregas, N.; Lozano-Elena, F.; Blasco-Escámez, D.; Tohge, T.; Martínez-Andújar, C.; Albacete, A.; Osorio, S.; Bustamante, M.; Riechmann, J.L.; Nomura, T.; et al. Overexpression of the vascular brassinosteroid receptor BRL3 confers drought resistance without penalizing plant growth. Nat. Commun. 2018, 9, 4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, A.J.; Andrews, J.; Mulholland, B.J.; McKee, J.M.T.; Hilton, H.W.; Horridge, J.S.; Farquhar, G.D.; Smeeton, R.C.; Smillie, I.R.A.; Black, C.R.; et al. Overproduction of abscisic acid in tomato increases transpiration efficiency and root hydraulic conductivity and influences leaf expansion. Plant Physiol. 2007, 143, 1905–1917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aroca, R.; Porcel, R.; Ruiz-Lozano, J.M. Regulation of root water uptake under abiotic stress conditions. J. Exp. Bot. 2012, 63, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Blum, A. Towards a conceptual ABA ideotype in plant breeding for water limited environments. Funct. Plant Biol. 2015, 42, 502–513. [Google Scholar] [CrossRef] [PubMed]
- Cruz de Carvalho, H.M. Drought stress and reactive oxygen species: Production, scavenging and signalling. Plant Signal Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef] [Green Version]
- Brodribb, T.J.; McAdam, S.A. Abscisic acid mediates a divergence in the drought response of two conifers. Plant Physiol. 2013, 162, 1370–1377. [Google Scholar] [CrossRef] [Green Version]
- Vandeleur, R.K.; Mayo, G.; Shelden, M.C.; Gilliham, M.; Kaiser, B.N.; Tyerman, S.D. The role of plasma membrane intrinsic protein aquaporins in water transport through roots: Diurnal and drought stress responses reveal different strategies between isohydric and anisohydric cultivars of grapevine. Plant Physiol. 2009, 149, 445–460. [Google Scholar] [CrossRef] [Green Version]
- Sharipova, G.; Ivanov, R.; Veselov, D.; Akhiyarova, G.; Shishova, M.; Nuzhnaya, T.; Kdoyarova, G. Involvement of reactive oxygen species in ABA-induced increase in hydraulic conductivity and aquaporin abundance. Int. J. Mol. Sci. 2021, 22, 9144. [Google Scholar] [CrossRef]
- Ahmed, S.; Kouser, S.; Asgher, M.; Gandhi, S.G. Plant aquaporins: A frontward to make crop plants drought resistant. Physiol. Plant. 2021, 172, 1089–1105. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Bhushan, A.; Rama, J.; Dubey, S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Bajji, M.; Kinet, J.-M.; Lutts, S. The use of electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant Growth Regul. 2002, 36, 61–70. [Google Scholar] [CrossRef]
- Blum, A.; Tuberosa, R. Dehydration survival of crop plants and its measurement. J. Exp. Bot. 2018, 69, 975–981. [Google Scholar] [CrossRef]
- Rajasheker, G.; Jawahar, G.; Jalaja, N.; Kumar, S.A.; Kumari, P.H.; Punita, D.L.; Karumanchi, A.R.; Palakolanu, S.R.; Polavarapu, R.; Sreenivasulu, N.; et al. Chapter 27—Role and Regulation of Osmolytes and ABA Interaction in Salt and Drought Stress Tolerance. In Plant Signaling Molecules; Khan, M.I.R., Reddy, P.S., Ferrante, A., Khan, N.A., Eds.; Woodhead Publishing: Sawston, UK, 2019; pp. 417–436. [Google Scholar] [CrossRef]
- Cardoso, A.A.; Gori, A.; Da-Silva, C.J.; Brunetti, C. Abscisic acid biosynthesis and signaling in plants: Key targets to improve water use efficiency and drought tolerance. Appl. Sci. 2020, 10, 6322. [Google Scholar] [CrossRef]
- Bernardo, S.; Dinis, L.T.; Machado, N.; Moutinho-Pereira, J. Grapevine abiotic stress assessment and search for sustainable adaptation strategies in Mediterranean-like climates. A review. Agron. Sustain. Dev. 2018, 38, 66. [Google Scholar] [CrossRef] [Green Version]
- Gómez, R.; Vicino, P.; Carrillo, N.; Lodeyro, A.F. Manipulation of oxidative stress responses as a strategy to generate stress-tolerant crops. From damage to signaling to tolerance. Crit. Rev. Biotechnol. 2019, 39, 693–708. [Google Scholar] [CrossRef]
- Nadarajah, K.K. ROS Homeostasis in abiotic stress tolerance in plants. Int. J. Mol. Sci. 2020, 21, 5208. [Google Scholar] [CrossRef]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [Green Version]
- Meena, M.; Divyanshu, K.; Kumar, S.; Swapnil, P.; Zehra, A.; Shukla, V.; Yadav, M.; Upadhyay, R.S. Regulation of L-proline biosynthesis, signal transduction, transport, accumulation and its vital role in plants during variable environmental conditions. Heliyon 2019, 5, e02952. [Google Scholar] [CrossRef] [Green Version]
- Anjum, S.A.; Wamg, L.; Farooq, M.; Khan, I.; Xue, L. Methyl jasmonate-induced alteration in lipid peroxidation, antioxidative defence system and yield in soybean under drought. J. Agron. Crop Sci. 2011, 197, 296–301. [Google Scholar] [CrossRef]
- Bandurska, H. Salicylic acid: An update on biosynthesis and action in plant response to water deficit and performance under drought. In Salicylic Acid Plant Growth and Development; Hayat, A., Ahmad, A., Alyemeni, M.N., Eds.; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; New York, NY, USA; London, UK, 2013; pp. 1–14. [Google Scholar]
- Chaudhry, S.; Sidhu, G.P.S. Climate change regulated abiotic stress mechanisms in plants: A comprehensive review. Plant Cell Rep. 2022, 41, 1–31. [Google Scholar] [CrossRef]
- Duan, Z.-Q.; Bai, L.; Zhao, Z.-G.; Zhang, G.-P.; Cheng, F.-M.; Jiang, L.-X.; Chen, K.-M. Drought-stimulated activity of plasma membrane nicotin amide adenine dinucleotide phosphate oxidase and its catalytic properties in rice. J. Integr. Plant Biol. 2009, 51, 1104–1115. [Google Scholar] [CrossRef]
- Wen, F.; Qin, T.; Wang, Y.; Dong, W.; Zhang, A.; Tan, M.; Jiang, M. OsHK3 is a crucial regulator of abscisic acid signaling involved in antioxidant defense in rice. J. Integr. Plant Biol. 2015, 57, 213–228. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, F.; Kuromori, T.; Urano, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Drought stress responses and resistance in plants: From cellular responses to long-distance intercellular communication. Front. Plant Sci. 2020, 11, 556972. [Google Scholar] [CrossRef]
- Mahmood, T.; Khalid, S.; Abdullah, M.; Ahmed, Z.; Shah, M.K.N.; Ghafoor, A.; Du, X. Insights into drought stress signaling in plants and the molecular genetic basis of cotton drought tolerance. Cells 2020, 9, 105. [Google Scholar] [CrossRef] [Green Version]
- Abhilasha, A.; Choudhury, R.S. Molecular and physiological perspectives of abscisic acid mediated drought adjustment strategies. Plants 2021, 10, 2769. [Google Scholar] [CrossRef]
- Ullah, A.; Sun, H.; Yang, X.; Zhang, X. Drought coping strategies in cotton: Increased crop per drop. Plant Biotechnol. J. 2017, 15, 271–284. [Google Scholar] [CrossRef]
- Per, T.S.; Khan, N.A.; Reddy, P.S.; Masood, A.; Hasanuzzaman, M.; Khan, M.I.R.; Anjum, N.A. Approaches in modulating proline metabolism in plants for salt and drought stress tolerance: Phytohormones, mineral nutrients and transgenics. Plant Physiol. Biochem. 2017, 115, 126–140. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: Multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Zarattini, M.; Forlani, G. Toward unveiling the mechanisms for transcriptional regulation of proline biosynthesis in the plant cell response to biotic and abiotic stress conditions. Front. Plant Sci. 2017, 8, 927. [Google Scholar] [CrossRef] [Green Version]
- Kakoulidou, I.; Avramidou, E.V.; Baránek, M.; Brunel-Muguet, S.; Farrona, S.; Johannes, F.; Kaiserli, E.; Lieberman-Lazarovich, M.; Martinelli, F.; Mladenov, V.; et al. Epigenetics for crop improvement in times of global change. Biology 2021, 10, 766. [Google Scholar] [CrossRef]
- Mozgova, I.; Mikulski, P.; Pecinka, A.; Farrona, S. Epigenetic mechanisms of abiotic stress response and memory in plants. In Epigenetics in Plants of Agronomic Importance: Fundamentals and Applications; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–64. [Google Scholar] [CrossRef]
- Miryeganeh, M. Plants’ Epigenetic mechanisms and abiotic stress. Genes 2021, 12, 1106. [Google Scholar] [CrossRef]
- Sun, C.; Ali, K.; Yan, K.; Fiaz, S.; Dormatey, R.; Bi, Z.; Bai, J. Exploration of Epigenetics for improvement of drought and other stress resistance in crops: A Review. Plants 2021, 10, 1226. [Google Scholar] [CrossRef]
- Zhang, C.Y.; Wang, N.N.; Zhang, Y.H.; Feng, Q.Z.; Yang, C.W.; Liu, B. DNA methylation involved in proline accumulation in response to osmotic stress in rice (Oryza sativa). Genet. Mol. Res. 2013, 12, 1269–1277. [Google Scholar] [CrossRef]
- Chang, Y.N.; Zhu, C.; Jiang, J.; Zhang, H.; Zhu, J.K.; Duan, C.G. Epigenetic regulation in plant abiotic stress responses. J. Integr. Plant Biol. 2020, 62, 563–580. [Google Scholar] [CrossRef]
- Thiebaut, F.; Hemerly, A.S.; Ferreira, P.C.G. A Role for epigenetic regulation in the adaptation and stress responses of non-model plants. Front. Plant Sci. 2019, 10, 246. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, J.; Inzé, D.; Nelissen, H.; Saibo, N.J.M. Source–sink regulation in crops under water deficit. Trends Plant Sci. 2019, 24, 652–663. [Google Scholar] [CrossRef]
- Tardieu, F. Any trait or trait-related allele can confer drought tolerance: Just design the right drought scenario. J. Exp. Bot. 2012, 63, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T. Diffusive and metabolic limitations to photosynthesis under drought and salinity in C3 plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef]
- Flexas, J.; Ribas-Carbó, M.; Bot, J.; Galmés, J.; Henkle, M.; Martínez-Cañellas, S.; Medrano, H. Decreased Rubisco activity during water stress is not induced by decreased relative water content but related to conditions of low stomatal conductance and chloroplast CO2 concentration. New Phytol. 2006, 172, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Gilliham, M.; Able, J.A.; Roy, S.J. Translating knowledge about abiotic stress tolerance to breeding programmes. Plant J. 2017, 90, 898–917. [Google Scholar] [CrossRef] [Green Version]
- Munné-Bosch, S.; Leonor, A. Die and let live: Leaf senescence contributes to plant survival under drought stress. Funct. Plant Biol. 2004, 31, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Obidiegwu, J.E.; Bryan, G.J.; Jones, H.G.; Prashar, A. Coping with drought: Stress and adaptive responses in potato and perspectives for improvement. Front. Plant Sci. 2015, 6, 542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, P.G.; Baum, M.; Grando, S.; Ceccarelli, S.; Bai, G.; Li, R.; von Korff, M.; Varshney, R.K.; Graner, A.; Volkoun, J. Differentially expressed genes between drought-tolerant and drought-sensitive barley genotypes in response to drought stress during the reproductive stage. J. Exp. Bot. 2009, 60, 3531–3544. [Google Scholar] [CrossRef]
- Perlikowski, D.; Kosmala, A. Mechanisms of drought resistance in introgression forms of Lolium multiflorum/Festuca arundinacea. Biol. Plant. 2020, 64, 497–503. [Google Scholar] [CrossRef]
- Pulvento, C.; Riccardi, M.; Lavini, A.; Iafelice, G.; Marconi, E.; d’Andria, R. Yield and quality characteristicsof quinoa grown in open field under different saline and non-saline irrigation regimes. J. Agron. Crop Sci. 2012, 198, 254–263. [Google Scholar] [CrossRef]
- Zurita-Silva, A.; Fuentes, F.; Zamora, P.; Jacobsen, S.-E.; Schwember, A.R. Breeding quinoa (Chenopodium quinoa Willd.): Potential and perspectives. Mol. Breed. 2014, 34, 13–30. [Google Scholar] [CrossRef]
- Hinojosa, L.; González, J.A.; Barrios-Masias, F.H.; Fuentes, F.; Murphy, K.M. Quinoa abiotic stress responses: A review. Plants 2018, 7, 106. [Google Scholar] [CrossRef] [Green Version]
- Yang, A.; Akhtar, S.S.; Amjad, M.; Iqbal, S.; Jacobsen, S.-E. Growth and physiological responses of quinoa to drought and temperature stress. J. Agron. Crop Sci. 2016, 202, 445–453. [Google Scholar] [CrossRef]
- Manaa, A.; Goussi, R.; Derbali, W.; Cantamessa, S.; Essemine, J.; Barbato, R. Photosynthetic performance of quinoa (Chenopodium quinoa Willd.) after exposure to a gradual drought stress followed by a recovery period. Biochim. Biophys. Acta (BBA) Bioenerg. 2021, 1862, 148383. [Google Scholar] [CrossRef] [PubMed]
- Simkin, A.J.; López-Calcagno, P.E.; Raines, C.A. Feeding the world: Improving photosynthetic efficiency for sustainable crop production. J. Exp. Bot. 2019, 70, 1119–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, A. Osmotic adjustment is a prime drought stress adaptive engine in support of plant production. Plant Cell Environ. 2017, 40, 4–10. [Google Scholar] [CrossRef]
- Ruggiero, A.; Punzo, P.; Landi, S.; Costa, A.; Van Oosten, M.J.; Grillo, S. Improving plant water use efficiency through molecular genetics. Horticulturae 2017, 3, 31. [Google Scholar] [CrossRef] [Green Version]
- Turner, N.C. Turgor maintenance by osmotic adjustment: 40 years of progress. J. Exp. Bot. 2018, 69, 3223–3233. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Liu, J.; Tischer, S.V.; Christmann, A.; Windisch, W.; Schnyder, H.; Grill, E. Leveraging abscisic acid receptors for efficient water use in Arabidopsis. Proc. Natl. Acad. Sci. USA 2016, 113, 6791–6796. [Google Scholar] [CrossRef] [Green Version]
- Dresselhaus., T.; Hückelhoven, R. Biotic and abiotic stress responses in crop plants. Agronomy 2018, 8, 267. [Google Scholar] [CrossRef] [Green Version]
- Blankenagel, S.; Yang, Z.; Avramova, V.; Schön, C.-C.; Grill, E. Generating plants with improved water use efficiency. Agronomy 2018, 8, 194. [Google Scholar] [CrossRef] [Green Version]
- González, J.A.; Bruno, M.; Valoy, M.; Prado, F.E. Genotypic variation of gas exchange parameters and leaf stable carbon and nitrogen isotopes in ten quinoa cultivars grown under drought. J. Agron. Crop Sci. 2011, 197, 81–93. [Google Scholar] [CrossRef]
- Abid, M.; Ali, S.; Qi, L.K.; Zahoor, R.; Tian, Z.; Jiang, D.; Snider, J.L.; Dai, T. Physiological and biochemical changes during drought and recovery periods at tillering and jointing stages in wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 4615. [Google Scholar] [CrossRef]
- Ghotbi-Ravandi, A.A.; Shahbazi, M.; Shariati, M.; Mulo, P. Effects of mild and severe drought stress on photosynthetic efficiency in tolerant and susceptible barley (Hordeum vulgare L.) genotypes. J. Agron. Crop Sci. 2014, 200, 403–415. [Google Scholar] [CrossRef]
- Kavi Kishor, P.B.; Sreenivasulu, N. Is proline accumulation per se correlated with stress tolerance or is proline homeostasis a more critical issue? Plant Cell Environ. 2014, 37, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Hare, P.D.; Cress, W.A. Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Regul. 1997, 21, 79–102. [Google Scholar] [CrossRef]
- Saeedipour, S. Relationship of grain yield, ABA and proline accumulation in tolerant and sensitive wheat cultivars as affected by water stress. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2013, 83, 311–315. [Google Scholar] [CrossRef]
- Frimpong, F.; Windt, C.W.; van Dusschoten, D.; Naz, A.A.; Frei, M.; Fiorani, F. A wild allele of pyrroline-5-carboxylate synthase1 leads to proline accumulation in spikes and leaves of barley contributing to improved performance under reduced water availability. Front. Plant Sci. 2021, 12, 633448. [Google Scholar] [CrossRef]
- Frimpong, F.; Anokye, M.; Windt, C.W.; Naz, A.A.; Frei, M.; van Dusschoten, D.; Fiorani, F. Proline mediated drought tolerance in the barley (Hordeum vulgare L.) isogenic line is associated with lateral root growth at the early seedlings stage. Plants 2021, 10, 2177. [Google Scholar] [CrossRef]
- Kundu, S.; Gantait, S. Abscisic acid signal crosstalk during abiotic stress response. Plant Gene 2017, 11, 61–69. [Google Scholar] [CrossRef]
- Onyemaobi, O.; Sangma, H.; Garg, G.; Wallace, X.; Kleven, S.; Suwanchaikasem, P.; Roessner, U.; Dolferus, R. Reproductive stage drought tolerance in wheat: Importance of stomatal conductance and plant growth regulators. Genes 2021, 12, 1742. [Google Scholar] [CrossRef]
- Kuppu, S.; Mishra, N.; Hu, R.; Sun, L.; Zhu, X.; Blumwald, E.; Payton, P.; Zhang, H. Water-deficit inducible expression of a cytokinin biosynthetic gene IPT improves drought tolerance in cotton. PLoS ONE 2013, 8, e64190. [Google Scholar] [CrossRef] [Green Version]
- Peleg, Z.; Reguera, M.; Tumimbang, E.; Walia, H.; Blumwald, E. Cytokinin-mediated source/sink modifications improve drought tolerance and increase grain yield in rice under water-stress. Plant Biotechnol. J. 2011, 9, 747–758. [Google Scholar] [CrossRef]
- Sharma, I.; Kaur, N.; Pati, P.K. Brassinosteroids: A promising option in deciphering remedial strategies for abiotic stress tolerance in rice. Front. Plant Sci. 2017, 8, 2151. [Google Scholar] [CrossRef] [PubMed]
- Nolan, T.M.; Vukašinović, N.; Liu, D.; Russinova, E.; Yanhai, Y. Brassinosteroids: Multidimensional regulators of plant growth, development, and stress responses. Plant Cell 2020, 32, 295–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hrmova, M.; Hussain, S.S. Plant transcription factors involved in drought and associated stresses. Int. J. Mol. Sci. 2021, 22, 5662. [Google Scholar] [CrossRef] [PubMed]
- Hoang, X.; Nhi, D.; Thu, N.; Thao, N.P.; Tran, L.P. Transcription factors and their roles in signal transduction in plants under abiotic stresses. Curr. Genom. 2017, 18, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Younis, A.; Ramzan, F.; Ramzan, Y.; Zulfiqar, F.; Ahsan, M.; Lim, K.B. Molecular markers improve abiotic stress tolerance in crops: A Review. Plants 2020, 9, 1374. [Google Scholar] [CrossRef]
- Godoy, F.; Olivos-Hernández, K.; Stange, C.; Handford, M. Abiotic stress in crop species: Improving tolerance by applying plant metabolites. Plants 2021, 10, 186. [Google Scholar] [CrossRef]
Figure 1.
Stress adjustment developed in plants.

Figure 2.
Plant responses to abiotic stress factors, coping strategy, and resistance.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bandurska, H. Drought Stress Responses: Coping Strategy and Resistance. Plants 2022, 11, 922. https://doi.org/10.3390/plants11070922
AMA Style
Bandurska H. Drought Stress Responses: Coping Strategy and Resistance. Plants. 2022; 11(7):922. https://doi.org/10.3390/plants11070922
Chicago/Turabian StyleBandurska, Hanna. 2022. "Drought Stress Responses: Coping Strategy and Resistance" Plants 11, no. 7: 922. https://doi.org/10.3390/plants11070922
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.