
Heritability and Associations among Grain Yield and Quality Traits in Quality Protein Maize (QPM) and Non-QPM Hybrids

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Variance Components and Heritability for Grain Yield, and Agronomic and Quality Traits
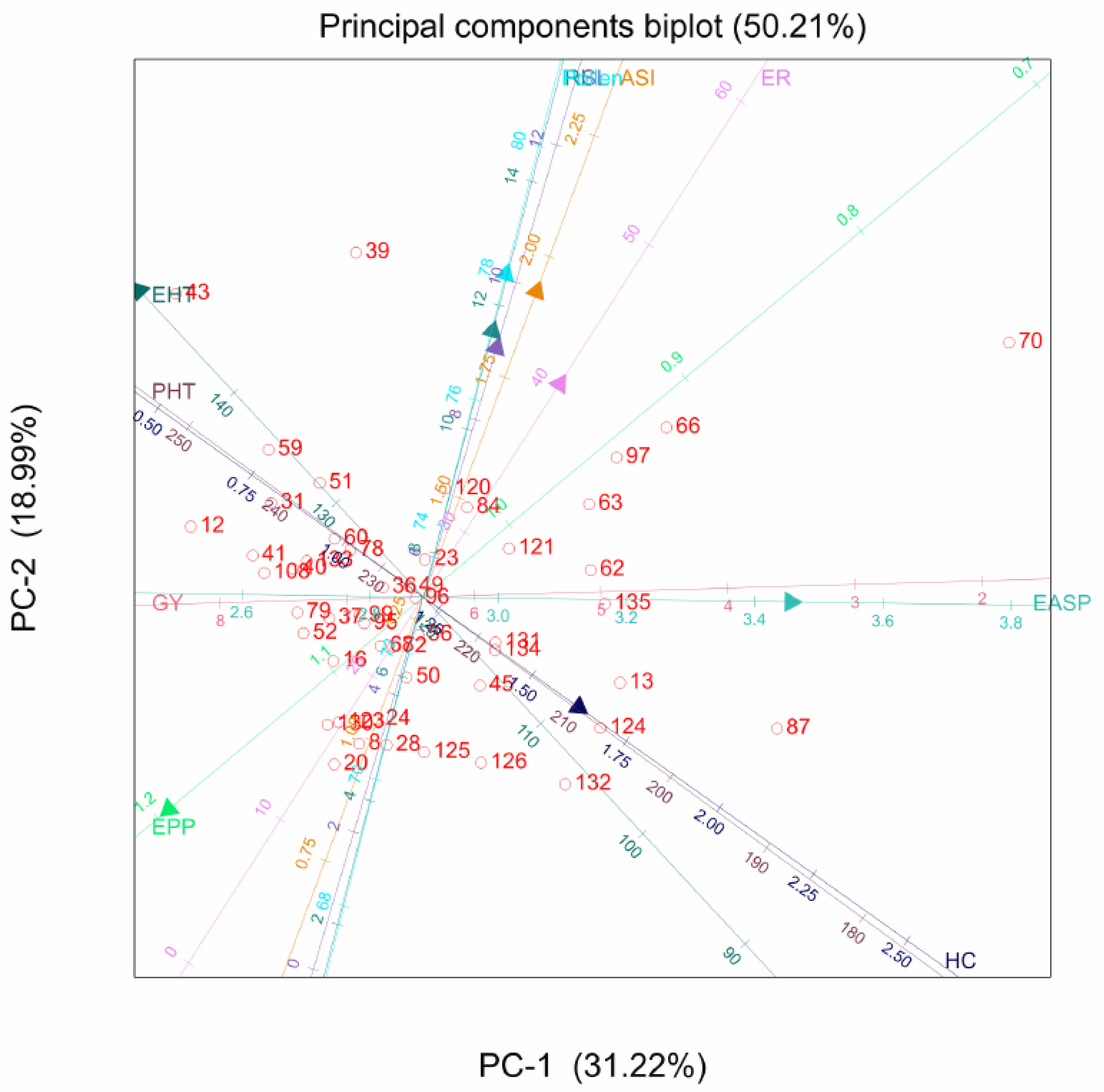
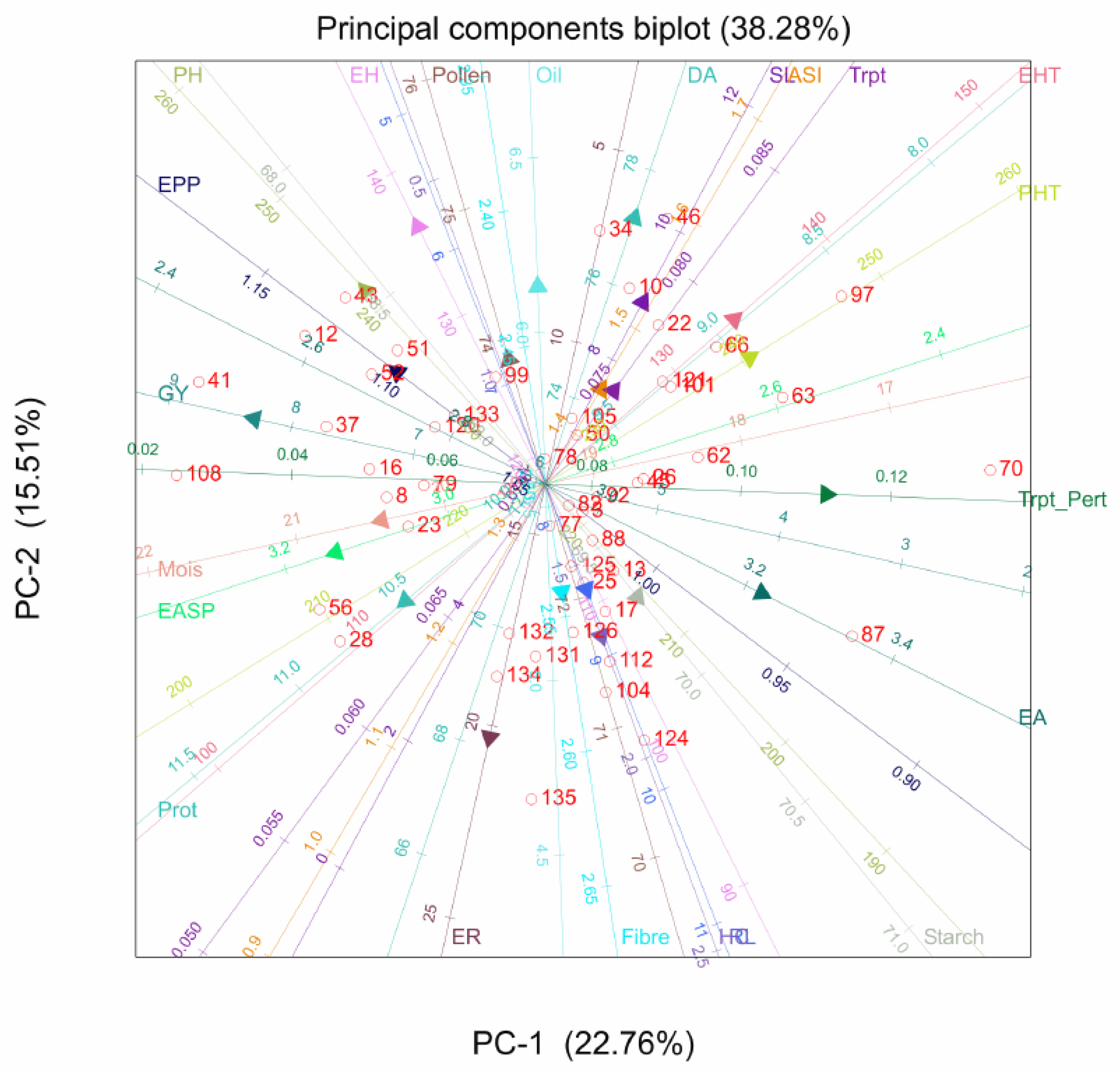
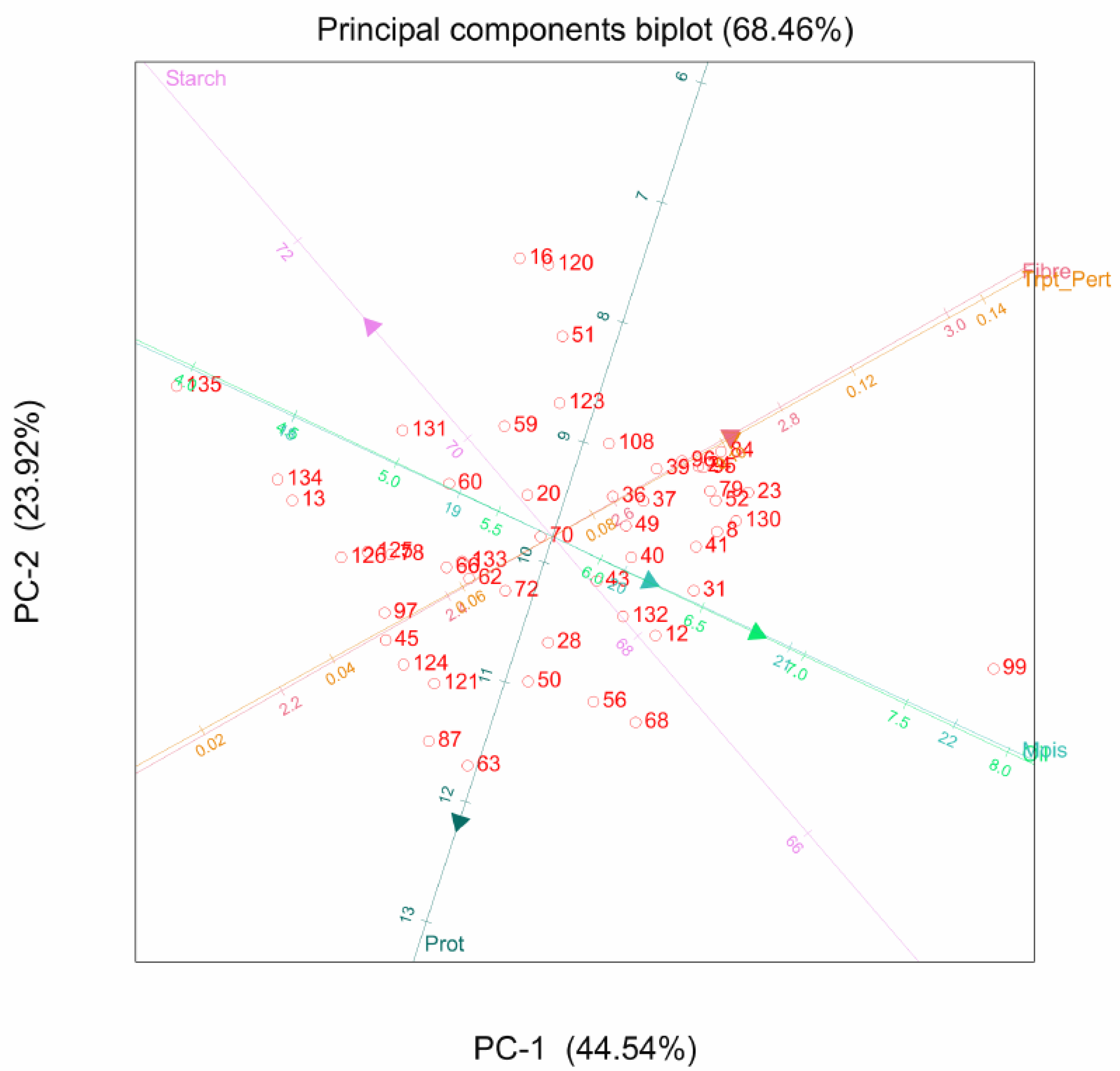
2.2. Principal Component Analysis
2.3. Genotypic and Phenotypic Correlation between Grain Yield and Other Agronomic Traits
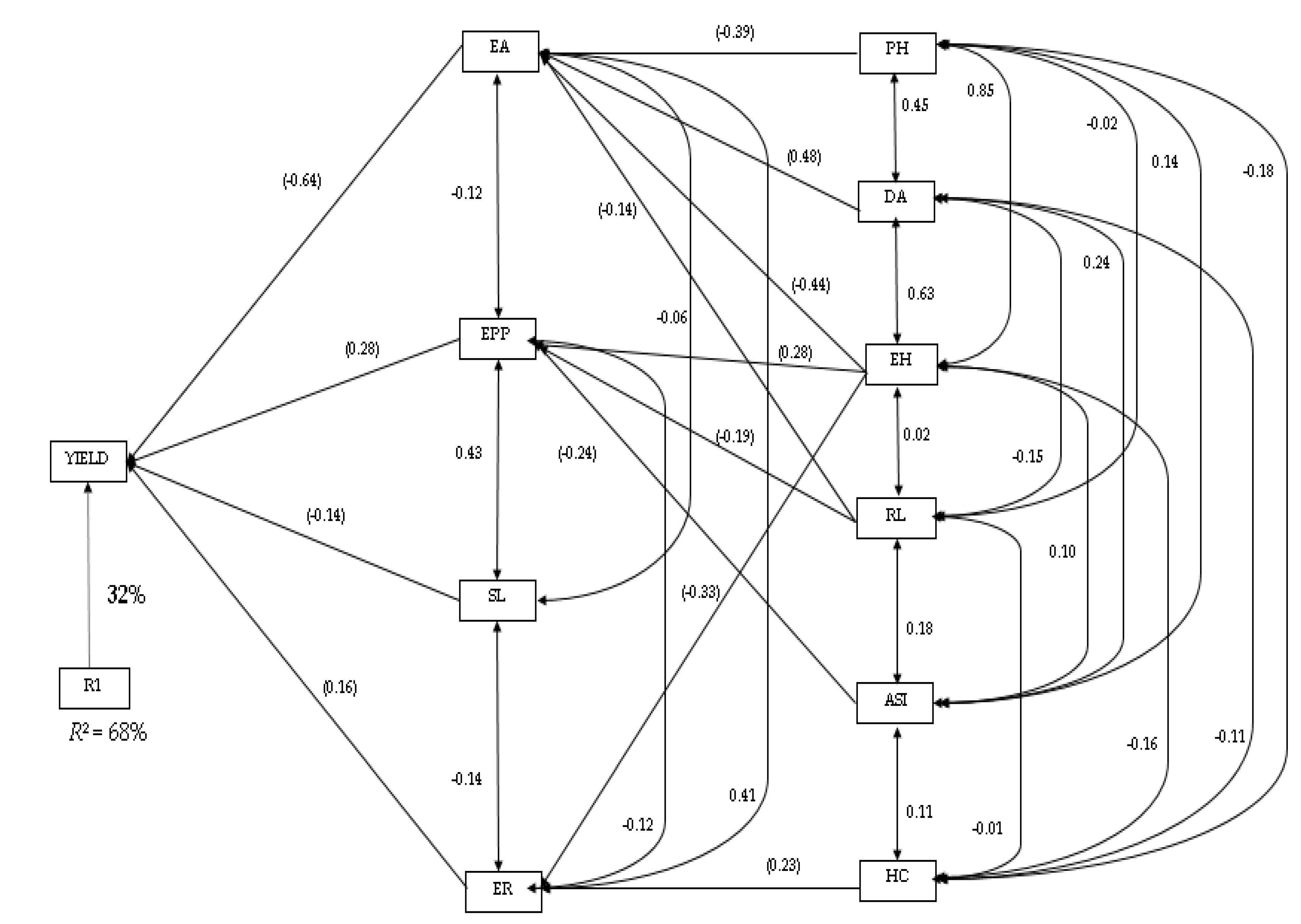
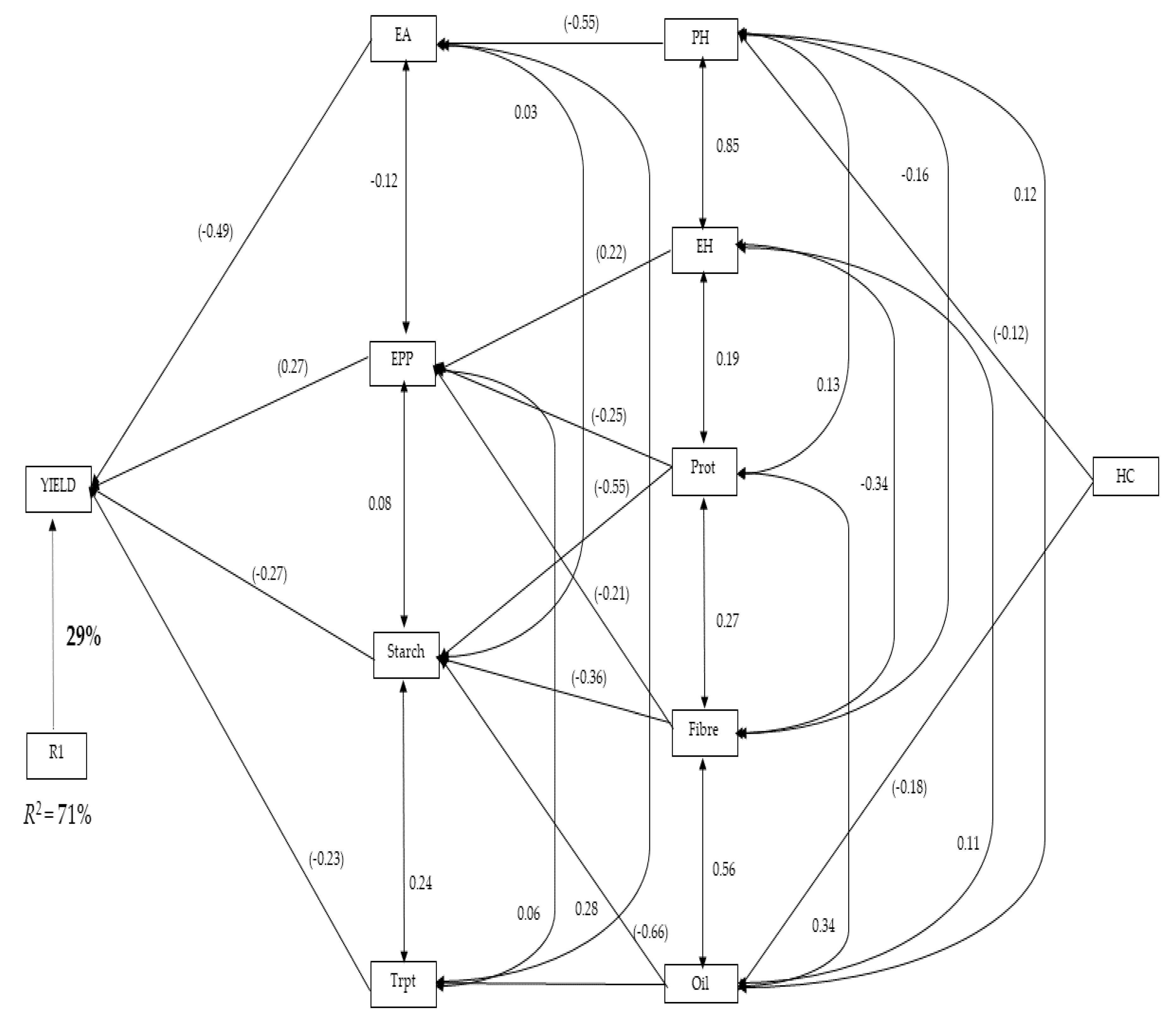
2.4. Path Coefficient Analysis for Grain Yield and Agronomic Traits
3. Discussion
3.1. Heritability and Variance Components for Agronomic and Quality Traits
3.2. Principal Component Analysis for Agronomic and Quality Traits
3.3. Correlation Coefficients and Path Analysis of Grain Yield, Agronomic and Quality Traits
4. Materials and Methods
4.1. Field Trials
4.2. Determination of Tryptophan and Starch
4.3. Determination of Protein, Oil, Moisture, and Fibre
4.4. Heritability Estimates
4.5. Estimation of Variance Components
4.6. Principal Component Analysis
4.7. Genetic and Phenotypic Correlation Estimations
4.8. Regression and Path Coefficient Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maqbool, M.A.; Issa, B.A.; Khokhar, E.S. Quality protein maize (QPM): Importance, genetics, timeline of different events, breeding strategies and varietal adoption. Plant Breed. 2021, 140, 375–399. [Google Scholar] [CrossRef]
- Mbuya, K.; Nkongolo, K.K.; Kalonji-Mbuyi, A. Nutritional analysis of quality protein maize varieties selected for agronomic characteristics in a breeding program. Int. J. Plant Breed. Gen. 2011, 5, 317–327. [Google Scholar] [CrossRef] [Green Version]
- Giwa, E.O.; Ikujenlola, A.V. Application of quality protein maize in the formulation of broiler’s finisher feed. J. Sci. Food Hosp. 2009, 1, 47–50. [Google Scholar]
- Blum, A. Heterosis, stress, and the environment: A possible road map towards the general improvement of crop yield. J. Exp. Bot. 2013, 64, 4829–4837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.; Li, Z.H.; Jiang, L.L.; Yu, X.M.; Ngezahayo, F.; Liu, B. Grain-yield heterosis in Zea mays L. shows positive correlation with parental difference in CHG Methylation. Crop Sci. 2010, 50, 2338–2346. [Google Scholar] [CrossRef]
- Machikowa, T.; Laosuwan, P. Path coefficient analysis for yield of early maturing soybean. Sonklanakarin J. Sci. Technol. 2011, 33, 365–368. [Google Scholar]
- Wegary, D.; Vivek, B.; Labuschagne, M. Association of parental genetic distance with heterosis and specific combining ability in quality protein maize. Euphytica 2013, 191, 205–216. [Google Scholar] [CrossRef]
- Amini, Z.; Khodambashi, M.; Houshmand, S. Correlation and path coefficient analysis of seed yield related traits in maize. Int. J. Agric. Crop Sci. 2013, 5, 2217–2220. [Google Scholar]
- Adesoji, A.G.; Abubakar, I.U.; Labe, D.A. Character association and path coefficient analysis of maize (Zea mays L.) grown under incorporated legumes and nitrogen. J. Agron. 2015, 14, 158–163. [Google Scholar] [CrossRef] [Green Version]
- Muturi, P.W.; Mgonja, M.; Rubaihayo, P. Gene action conditioning resistance traits to spotted stem borer, Chilo partellus, in grain sorghum. Int. J. Trop. Insect Sci. 2019, 39, 147–155. [Google Scholar] [CrossRef]
- Jilo, T. Nutritional benefit and development of quality protein maize (QPM) in Ethiopia: Review article. Cereal Res. Commun. 2021, 1–14. [Google Scholar] [CrossRef]
- Aman, J.; Bantte, K.; Alamerew, S.; Tolera, B. Evaluation of quality protein maize (Zea mays L.) hybrids at Jimma, Western Ethiopia. J. Forensic Anthropol. 2016, 1, 1–6. [Google Scholar]
- Sharma, P.; Punia, M.S.; Kamboj, M.C.; Singh, N.; Chand, M. Evaluation of quality protein maize crosses through line x tester analysis for grain yield and quality traits. Agric. Sci. Dig. 2017, 37, 42–45. [Google Scholar]
- Aliu, S.; Rusinovci, I.; Fetahu, S.; Simeonovska, E. Genetic diversity and correlation estimates for grain yield and quality traits in Kosovo local maize (Zea mays L.) populations. Not. Sci. Biol. 2012, 4, 121–128. [Google Scholar] [CrossRef]
- Pixley, K.V.; Bjarnason, M.S. Stability of grain yield, endosperm modification, and protein quality of hybrid and open-pollinated quality protein maize cultivars. Crop Sci. 2002, 42, 1882–1890. [Google Scholar] [CrossRef]
- Meseka, S.; Fakorede, M.A.; Ajala, S.; Badu-Apraku, B.; Menkir, A. Introgression of alleles from maize landraces to improve drought tolerance in an adapted germplasm. J. Crop Improv. 2013, 27, 96–112. [Google Scholar] [CrossRef]
- Sreckov, Z.; Nastasic, A.; Bocanski, J.; Djalovic, I.; Vukosavljev, M.; Jockovic, B. Correlation and path analysis of grain yield and morphological traits in test–cross populations of maize. Pak. J. Bot. 2011, 43, 1729–1731. [Google Scholar]
- Bizeti, H.S.; Carvalho, C.G.P.; Souza, J.R.P.; Destro, D. Path analysis under multicollinearity in soybean. Braz. Arch. Biol. Technol. 2004, 47, 669–676. [Google Scholar] [CrossRef] [Green Version]
- Udensi, O.; Ikpeme, E.V. Correlation and path coefficient analyses of seed yield and its contributing traits in Cajanus cajan (L.). Millsp. Am. J. Exp. Agric. 2012, 2, 351–358. [Google Scholar] [CrossRef] [Green Version]
- Halilu, A.D.; Ado, S.G.; Aba, D.A.; Usman, I.S. Genetics of carotenoids for provitamin A biofortification in tropical-adapted maize. Crop J. 2016, 4, 313–322. [Google Scholar] [CrossRef] [Green Version]
- Ababulgu, D.; Shimelis, H.; Laing, M.; Beyene, A. Phenotypic characterization of elite quality protein maize (QPM) inbred lines adapted to tropical-highlands and the association studies using SSR markers. Aus. J. Crop Sci. 2018, 12, 22–31. [Google Scholar] [CrossRef]
- Dutta, R.; Kumar, A.; Chandra, S.; Ngachan, S.V. Genetic divergence, path coefficient, principal component and cluster analyses of maize genotypes in the mid-altitudes of Meghalaya. Maydica 2017, 62, 1–5. [Google Scholar]
- Mastrodomenico, A.; Hendrix, C.; Below, F. Nitrogen use efficiency and the genetic variation of maize expired plant variety protection germplasm. Agriculture 2017, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Nyine, M.; Uwimana, B.; Blavet, N.; Hřibová, E.; Vanrespaille, H.; Batte, M.; Akech, V.; Brown, A.; Lorenzen, J.; Swennen, R.; et al. Genomic Prediction in a Multiploid Crop: Genotype by Environment Interaction and Allele Dosage Effects on Predictive Ability in Banana. Plant Genome 2018, 11, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Setimela, P.S.; Gasura, E.; Tarekegne, A.T. Evaluation of grain yield and related agronomic traits of quality protein maize hybrids in Southern Africa. Euphytica 2017, 213, 1–14. [Google Scholar] [CrossRef]
- Causse, M.; Chaïb, J.; Lecomte, L.; Buret, M.; Hospital, F. Both additivity and epistasis control the genetic variation for fruit quality traits in tomato. Theor. Appl. Genet. 2007, 115, 429–442. [Google Scholar] [CrossRef]
- Liu, H.; Pandey, M.K.; Xu, Z.; Rao, D.; Huang, Z.; Chen, M.; Feng, D.; Varshney, R.K.; Hong, Y. Analysis and evaluation of quality traits of peanut varieties with near infra-red spectroscopy technology. Int. J. Agric. Biol. 2019, 21, 491–498. [Google Scholar] [CrossRef]
- Sarika, K.; Hossain, F.; Muthusamy, M.; Zunjare, R.U.; Baveja, A.; Goswami, R.; Bhat, J.S.; Saha, S.; Gupta, H.S. Marker-assisted pyramiding of opaque2 and novel opaque16 genes for further enrichment of lysine and tryptophan in sub-tropical maize. Plant Sci. 2018, 272, 142–152. [Google Scholar] [CrossRef]
- Izzam, A.; Rehman, H.; Sohail, A.; Ali, S.; Manzoor, A.; Hussain, Q. Genetic variability and correlation studies for morphological and yield traits in maize (Zea mays L.). Pure Appl. Biol. 2017, 6, 1234–1243. [Google Scholar] [CrossRef]
- Singh, G.; Kumar, R. Genetic parameters and character association study for yield traits in maize (Zea mays L.). J. Pharmacogn. Phytochem. 2017, 6, 808–813. [Google Scholar]
- Babu, R.; Prasanna, B.M. Molecular breeding for quality protein maize (QPM). In Genomics of Plant Genetic Resources; Tuberosa, R., Graner, A., Frison, E., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 490–505. [Google Scholar] [CrossRef] [Green Version]
- Bhusal, T.N.; Lal, G.M. Relationship among heterosis, combining ability and SSR based genetic distance in single cross hybrids of maize (Zea Mays L.). Vegetos—Int. J. Plant Res. 2017, 30, 1–17. [Google Scholar] [CrossRef]
- Pixley, K.V.; Bjarnason, M.S. Combining ability for yield and protein quality among modified-endosperm opaque-2 tropical maize inbreds. Crop Sci. 1993, 33, 1229–1234. [Google Scholar] [CrossRef]
- Kumar, A.; Kumari, J.; Rana, J.C.; Chaudhary, D.P.; Kumar, R.; Singh, H.; Singh, T.P.; Dutta, M. Diversity among maize landraces in North West Himalayan region of India assessed by agro-morphological and quality traits. Indian J. Gen. 2015, 75, 188–195. [Google Scholar] [CrossRef]
- Mutiga, S.K.; Morales, L.; Angwenyi, S.; Wainaina, J.; Harvey, J.; Das, B.; Nelson, B.J. Field crops research association between agronomic traits and aflatoxin accumulation in diverse maize lines grown under two soil nitrogen levels in Eastern Kenya. Field Crops Res. 2017, 205, 124–134. [Google Scholar] [CrossRef]
- Bocianowski, J.; Górczak, K.; Nowosad, K.; Rybiński, W.; Piesik, D. Path analysis and estimation of additive and epistatic gene effects of barley SSD lines. J. Integr. Agric. 2016, 15, 1983–1990. [Google Scholar] [CrossRef] [Green Version]
- Raza, I.; Khan, A.; Masood, A.; Abid, S. Path analysis for potato crisps. Int. J. Sci. Eng. Res. 2018, 9, 1565–1570. [Google Scholar]
- Badu-Apraku, B.; Fakorede, M.A.B.; Annor, B.; Talabi, A.O. Improvement in grain yield and low-nitrogen tolerance in maize cultivars of three eras. Exp. Agric. 2017, 54, 1–9. [Google Scholar] [CrossRef]
- Barrera, S.F.; Ribeiro, L.P.; Rodrigues, E.V.; Lopes, B.L.; Eduardo, T.P. Correlations and path analysis in cherry tomato genotypes. Funct. Plant Breed. J. 2019, 1, 37–44. [Google Scholar] [CrossRef]
- John, K.; Santhoshi, M.V.M.; Rajasekhar, P. Correlation and path analysis for yield and yield attributes in groundnut (Arachis hypogaea L.). Legume Res. 2019, 42, 518–522. [Google Scholar] [CrossRef]
- Kanwar, S.S.; Nag, Y.K. To study correlation and path analysis in rice breeding lines. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 2481–2487. [Google Scholar] [CrossRef]
- Nyamapfene, K.W. The Soils of Zimbabwe; Nehanda Publishers: Harare, Zimbabwe, 1991; p. 179. [Google Scholar]
- Nurit, E.; Tiessen, A.; Pixley, K.; Palacios-Rojas, N. Reliable and inexpensive colorimetric method for deter- mining protein-bound tryptophan in maize kernels. J. Agric. Food Chem. 2009, 57, 7233–7238. [Google Scholar] [CrossRef] [PubMed]
- Caprita, R.; Caprita, A.; Cretescu, I. Effective extraction of soluble non-starch polysaccharides and viscosity determination of aqueous extracts from wheat and barley. Proc. World Cong. Eng. Compt. Sci. 2011, 2, 1–4. [Google Scholar]
- Dabholkar, A.R. Elements of Biometrical Genetics; Revised and Enlarged Edition; Concept Publishing Company: New Delhi, India, 1999; pp. 138–140. [Google Scholar]
- GenStat. GenStat for Windows 20th Edition (2019); VSN International: Hemel Hempstead, UK, 2019. [Google Scholar]
- Alvarado, G.; López, M.; Vargas, M.; Pacheco, Á.; Rodríguez, F.; Burgueño, J.; Crossa, J. META-R (Multi Environment Trail Analysis with R for Windows); Version 6.04, hdl:11529/10201; CIMMYT Research Data and Software Repository Network, V23. 2015. Available online: https://data.cimmyt.org/dataset.xhtml?persistentId=hdl:11529/10201 (accessed on 15 January 2022).
- Yu, R.; Zheng, Y.; Abdel-Aty, M.; Gao, Z. Exploring crash mechanisms with microscopic traffic flow variables: A hybrid approach with latent class logit and path analysis models. Accid. Anal. Prev. 2019, 125, 70–78. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Variance Components | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Line Variance | Tester Variance | Line x Tester Variance | Genotype Variance | Additive Variance | Dominance Variance | Environmental Variance | Broad-Sense Heritability (%) | Narrow-Sense Heritability (%) | |
| GY | 0.030 | 0.1800 | 0.43000 | 0.5800 | 2.320000 | 1.72000 | 0.13000 | 0.970 | 0.560 |
| ASI | 0.002 | 0.0200 | 0.02400 | 0.0420 | 0.167000 | 0.09700 | 0.04500 | 0.854 | 0.541 |
| EH | 56.020 | 67.9600 | 24.04000 | 130.6200 | 522.460000 | 96.17000 | 6.61000 | 0.990 | 0.840 |
| EPP | 0.001 | 0.0010 | 0.00300 | 0.0050 | 0.019000 | 0.01200 | 0.00100 | 0.955 | 0.578 |
| ER | 5.320 | 2.6500 | 0.74000 | 7.8700 | 31.490000 | 2.96000 | 7.04000 | 0.830 | 0.760 |
| EA | 0.004 | 0.0060 | 0.01700 | 0.0250 | 0.101000 | 0.06800 | 0.01000 | 0.946 | 0.563 |
| HC | 0.117 | 0.0000 | 0.02900 | 0.1470 | 0.590000 | 0.11600 | 0.26500 | 0.727 | 0.608 |
| PH | 54.170 | 54.7300 | 52.73000 | 147.7000 | 590.780000 | 210.92000 | 9.28000 | 0.990 | 0.730 |
| DA | 3.720 | 2.5400 | 0.68000 | 6.2900 | 25.160000 | 2.73000 | 0.24000 | 0.990 | 0.900 |
| RL | 3.060 | 0.0000 | 0.00000 | 2.9000 | 11.580000 | 0.00000 | 7.02000 | 0.620 | 0.620 |
| SL | 1.210 | 1.6500 | 0.46000 | 2.8700 | 11.470000 | 1.83000 | 4.66000 | 0.740 | 0.640 |
| Fibre | 0.005 | 0.0110 | 0.00500 | 0.0180 | 0.074000 | 0.01900 | 0.00200 | 0.978 | 0.777 |
| Moisture | 0.132 | 0.0590 | 0.12800 | 0.2980 | 1.191000 | 0.51300 | 0.23600 | 0.878 | 0.614 |
| Oil | 0.133 | 0.0890 | 0.01400 | 0.2120 | 0.849000 | 0.05600 | 0.04400 | 0.954 | 0.895 |
| Protein | 0.133 | 0.1880 | 0.06900 | 0.3440 | 1.375000 | 0.27400 | 0.06300 | 0.963 | 0.803 |
| Starch | 0.305 | 0.1950 | 0.09900 | 0.5470 | 2.187000 | 0.39400 | 0.15200 | 0.944 | 0.800 |
| Tryptophan | 0.000 | 0.0003 | 0.00001 | 0.0003 | 0.001020 | 0.00003 | 0.00002 | 0.983 | 0.956 |
| Traits | Eigenvectors | |||
|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC4 | |
| Grain yield | 0.018 | 0.012 | 0.019 | −0.021 |
| Plant height | 0.718 | 0.657 | −0.211 | −0.011 |
| Ear height | 0.682 | −0.610 | 0.364 | 0.003 |
| ASI | 0.003 | 0.005 | −0.014 | 0.019 |
| Days-to-shed | 0.095 | −0.202 | −0.058 | −0.067 |
| Root lodging | −0.001 | 0.111 | 0.199 | 0.893 |
| Stalk lodging | 0.002 | −0.146 | −0.191 | 0.427 |
| Husk cover | −0.006 | 0.009 | 0.023 | −0.007 |
| Ears per plant | 0.001 | −0.002 | 0.000 | −0.003 |
| Ear rot | −0.105 | 0.347 | 0.861 | −0.119 |
| Ear aspect | −0.006 | −0.001 | 0.008 | −0.001 |
| Eigenvalue | 273.987 | 23.479 | 18.174 | 14.102 |
| Proportion (%) | 80.10 | 6.90 | 5.30 | 4.10 |
| Cumulative (%) | 80.10 | 87.00 | 92.30 | 96.40 |
| Traits | Eigenvectors | |||
|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC4 | |
| Tryptophan | 0.002 | −0.003 | −0.011 | −0.002 |
| Moisture | 0.982 | −0.065 | 0.157 | 0.077 |
| Protein | −0.048 | 0.811 | 0.534 | 0.205 |
| Oil | 0.048 | 0.185 | −0.602 | 0.756 |
| Starch | −0.172 | −0.551 | 0.568 | 0.557 |
| Fibre | 0.030 | −0.026 | −0.063 | −0.266 |
| Eigenvalue | 3.562 | 1.149 | 0.837 | 0.029 |
| Proportion (%) | 63.663 | 20.540 | 14.961 | 0.521 |
| Cumulative (%) | 63.66 | 84.20 | 99.16 | 99.68 |
| Eigenvectors | ||||||
|---|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | |
| Grain yield | 0.018 | 0.013 | 0.020 | −0.020 | −0.002 | 0.215 |
| Plant height | 0.718 | 0.657 | −0.211 | −0.014 | 0.068 | −0.040 |
| Ear height | 0.681 | −0.609 | 0.365 | 0.007 | −0.038 | 0.069 |
| ASI | 0.003 | 0.005 | −0.014 | 0.019 | 0.006 | 0.010 |
| Days-to-anthesis | 0.095 | −0.201 | −0.059 | −0.065 | 0.108 | −0.266 |
| Root lodging | −0.001 | 0.112 | 0.194 | 0.892 | −0.363 | −0.027 |
| Stalk lodging | 0.002 | −0.144 | −0.195 | 0.429 | 0.849 | −0.041 |
| Husk cover | −0.006 | 0.010 | 0.023 | −0.007 | 0.034 | 0.024 |
| Ears per plant | 0.001 | −0.002 | 0.000 | −0.003 | 0.005 | 0.002 |
| Ear rot | −0.105 | 0.348 | 0.859 | −0.112 | 0.317 | −0.096 |
| Ear aspect | −0.006 | −0.001 | 0.008 | −0.001 | −0.002 | −0.034 |
| Tryptophan | 0.000 | 0.001 | −0.001 | 0.001 | 0.001 | 0.000 |
| Moisture | −0.010 | 0.043 | 0.031 | 0.014 | 0.107 | 0.912 |
| Protein | 0.007 | −0.010 | 0.046 | −0.017 | −0.128 | 0.045 |
| Oil | 0.006 | 0.010 | −0.026 | 0.030 | 0.010 | 0.022 |
| Starch | −0.007 | −0.014 | 0.021 | −0.024 | 0.016 | −0.171 |
| Fibre | −0.002 | 0.009 | −0.004 | 0.003 | 0.006 | 0.019 |
| Eigenvalue | 274.053 | 23.527 | 18.247 | 14.127 | 8.378 | 3.628 |
| Proportion (%) | 78.80 | 6.800 | 5.30 | 4.10 | 2.40 | 1.00 |
| Cumulative (%) | 78.80 | 85.60 | 90.80 | 94.90 | 97.30 | 98.40 |
| Traits | GY | DA | ASI | PH | EH | RL | SL | EPP | HC | ER | EA | Trpt | Mois | Prot | Oil | Fibre | Starch |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GY | - | −0.16 | −0.14 | 0.29 ** | 0.19 * | 0.18 * | −0.19 * | 0.42 ** | −0.1 | 0.06 | −0.54 ** | −0.27 ** | 0.26 ** | 0.25 * | 0.07 | −0.06 | −0.17 |
| DA | −0.19 ** | - | 0.35 ** | 0.39 ** | 0.60 ** | 0.02 | 0.22 * | −0.04 | −0.33 ** | −0.29 ** | 0.01 | −0.10 | 0.03 | −0.07 | 0.13 | −0.27 ** | 0.02 |
| ASI | −0.23 ** | 0.78 ** | - | 0.09 | 0.14 | 0.15 | 0.32 ** | −0.29 ** | 0.08 | −0.09 | 0.14 | 0.10 | 0.19* | −0.24 * | 0.14 | 0.18 * | −0.01 |
| PH | 0.40 ** | 0.43 ** | 0.30 ** | - | 0.82 ** | 0.20 * | 0.14 | 0.15 | −0.15 | −0.17 | −0.53 ** | −0.09 | 0.01 | 0.08 | 0.14 | −0.07 | −0.09 |
| EH | 0.27 ** | 0.65 ** | 0.37** | 0.86 ** | - | 0.20 * | 0.19 * | 0.14 | −0.23 * | −0.24 * | −0.44 ** | −0.23 * | −0.07 | 0.18 * | 0.04 | −0.26 ** | −0.01 |
| RL | NA | NA | NA | NA | NA | - | 0.13 | −0.03 | 0.23 * | −0.03 | −0.18 * | −0.18 * | 0.01 | 0.14 | 0.14 | −0.07 | 0.04 |
| SL | −0.28 ** | 0.47 ** | 1.00 ** | 0.37 ** | 0.43 ** | NA | - | −0.08 | 0.03 | −0.20 ** | −0.13 | 0.19 * | 0.04 | −0.26 ** | −0.02 | −0.09 | 0.21 * |
| EPP | 0.64 ** | −0.08 | −0.76 ** | 0.19 * | 0.20 * | NA | −0.16 | - | −0.20 * | 0.05 | −0.34 ** | −0.10 | 0.01 | 0.05 | −0.04 | −0.17 | 0.07 |
| HC | −0.16 | −0.68 ** | 0.21 * | −0.24 * | −0.41 ** | NA | 0.06 | −0.63 ** | - | 0.18 * | 0.06 | 0.11 | −0.05 | −0.01 | −0.10 | 0.16 | 0.06 |
| ER | 0.29 ** | −0.47 ** | −0.35 ** | −0.20 * | −0.39 ** | NA | −0.88 ** | 0.31 ** | 0.36 ** | - | 0.28 ** | −0.03 | 0.12 | 0.06 | −0.03 | 0.16 | −0.09 |
| EA | −0.83 ** | 0.01 | 0.27 ** | −0.83 ** | −0.74 ** | NA | −0.68 ** | −0.58 * | −0.02 | 0.11 | - | 0.19* | −0.02 | −0.14 | 0.05 | 0.14 | −0.08 |
| Trpt | −0.39 ** | −0.12 | 0.16 | −0.12 | −0.26 ** | NA | 0.42 ** | −0.18 * | 0.24 * | −0.04 | 0.35 ** | - | 0.15 | −0.56 ** | 0.51 ** | 0.39 ** | −0.30 ** |
| Mois | 0.50 ** | 0.05 | 0.38 ** | 0.02 | −0.10 | NA | 0.05 | 0.06 | −0.06 | 0.15 | −0.07 | 0.21 * | - | −0.32 ** | 0.20 * | 0.48 ** | −0.20 * |
| Prot | 0.38 ** | −0.04 | −0.45 ** | 0.11 | 0.23 * | NA | −0.54 ** | 0.15 | −0.01 | 0.07 | −0.31 ** | −0.64 ** | −0.45 ** | - | −0.15 | −0.40 ** | −0.30 ** |
| Oil | 0.04 | 0.14 | 0.21 * | 0.14 | 0.03 | NA | −0.16 | −0.09 | −0.23 * | −0.05 | 0.13 | 0.58 ** | 0.32 ** | −0.17 * | - | 0.29 ** | −0.69 ** |
| Fibre | −0.10 | −0.31 ** | 0.28 ** | −0.09 | −0.31 ** | NA | −0.19 * | −0.26 ** | 0.31 ** | 0.24 ** | 0.20 * | 0.44 ** | 0.64 ** | −0.48 ** | 0.33 ** | - | −0.32 ** |
| Starch | −0.25* | 0.01 | 0.01 | −0.12 | −0.02 | NA | 0.44 ** | 0.04 | 0.16 | −0.13 | −0.08 | −0.35 ** | −0.29 ** | −0.21 * | −0.78 ** | −0.36 ** | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amegbor, I.K.; van Biljon, A.; Shargie, N.; Tarekegne, A.; Labuschagne, M.T. Heritability and Associations among Grain Yield and Quality Traits in Quality Protein Maize (QPM) and Non-QPM Hybrids. Plants 2022, 11, 713. https://doi.org/10.3390/plants11060713
Amegbor IK, van Biljon A, Shargie N, Tarekegne A, Labuschagne MT. Heritability and Associations among Grain Yield and Quality Traits in Quality Protein Maize (QPM) and Non-QPM Hybrids. Plants. 2022; 11(6):713. https://doi.org/10.3390/plants11060713
Chicago/Turabian StyleAmegbor, Isaac Kodzo, Angeline van Biljon, Nemera Shargie, Amsal Tarekegne, and Maryke T. Labuschagne. 2022. "Heritability and Associations among Grain Yield and Quality Traits in Quality Protein Maize (QPM) and Non-QPM Hybrids" Plants 11, no. 6: 713. https://doi.org/10.3390/plants11060713