
Plastid Phylogenomic Analysis of Tordylieae Tribe (Apiaceae, Apioideae)


Abstract
:1. Introduction
2. Results and Discussion
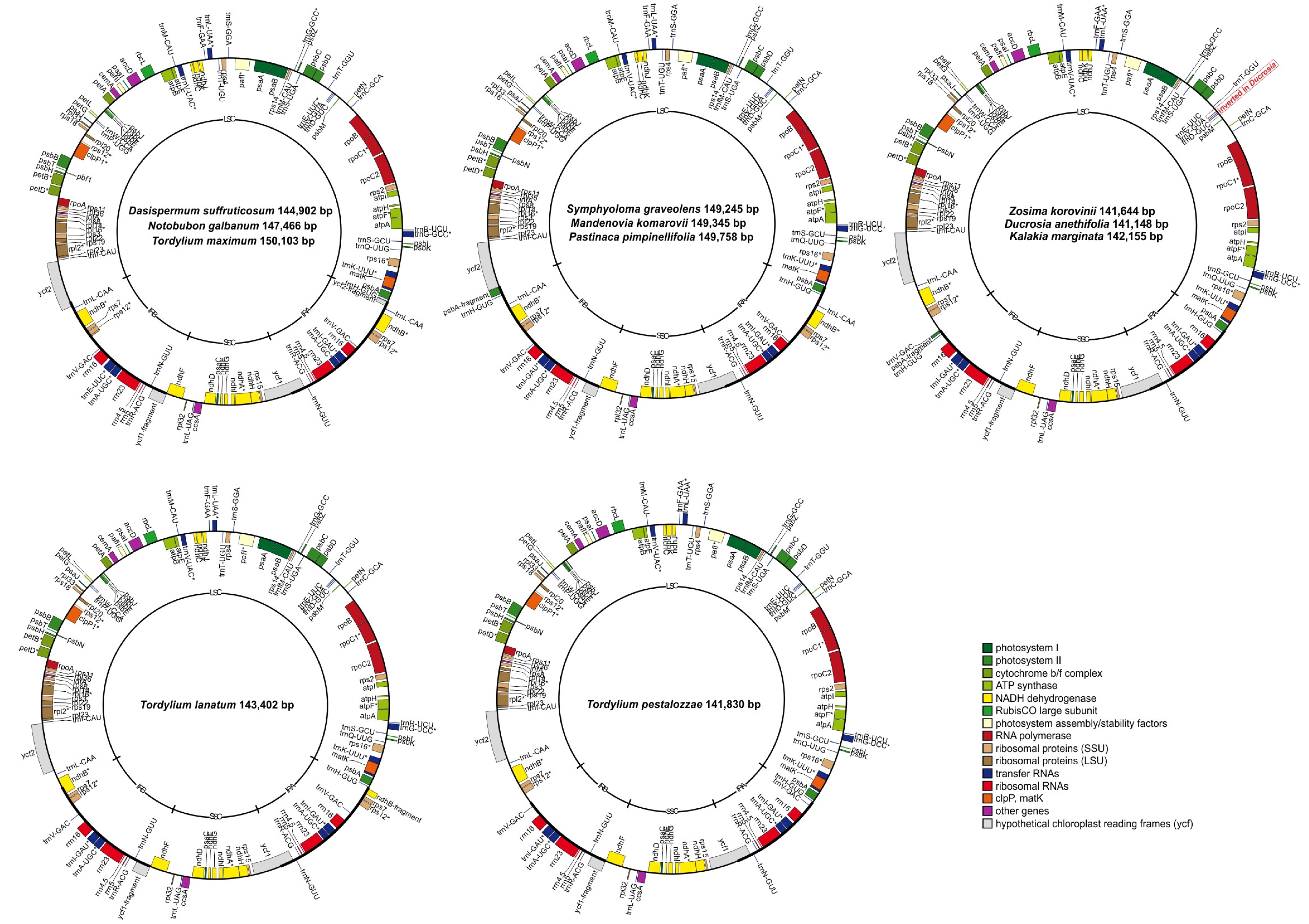
2.1. General Overview of Plastomes
2.2. Inverted Repeat Contractions and Expansions
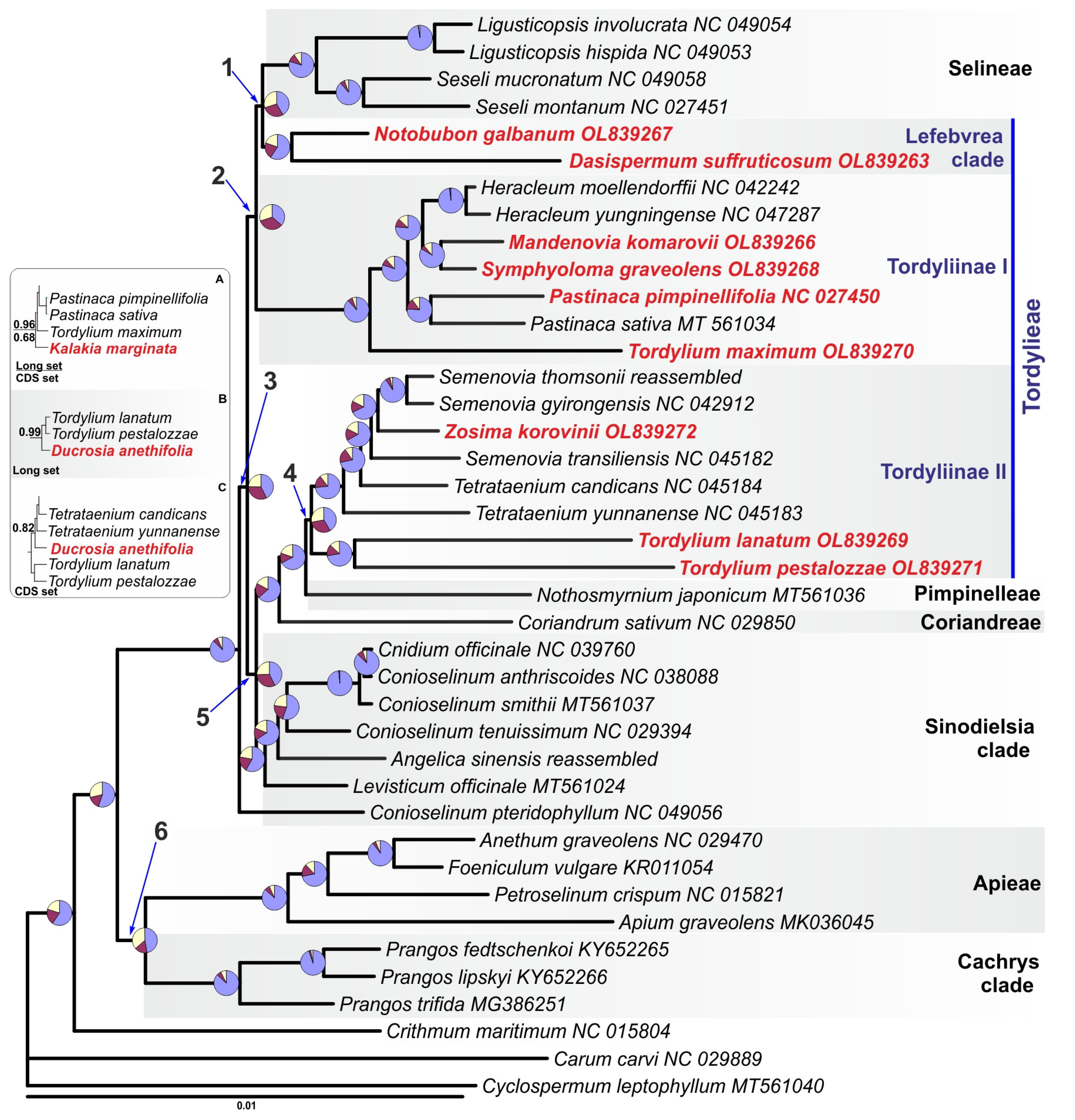
2.3. Phylogenetic Analysis of Tordylieae Plastomes
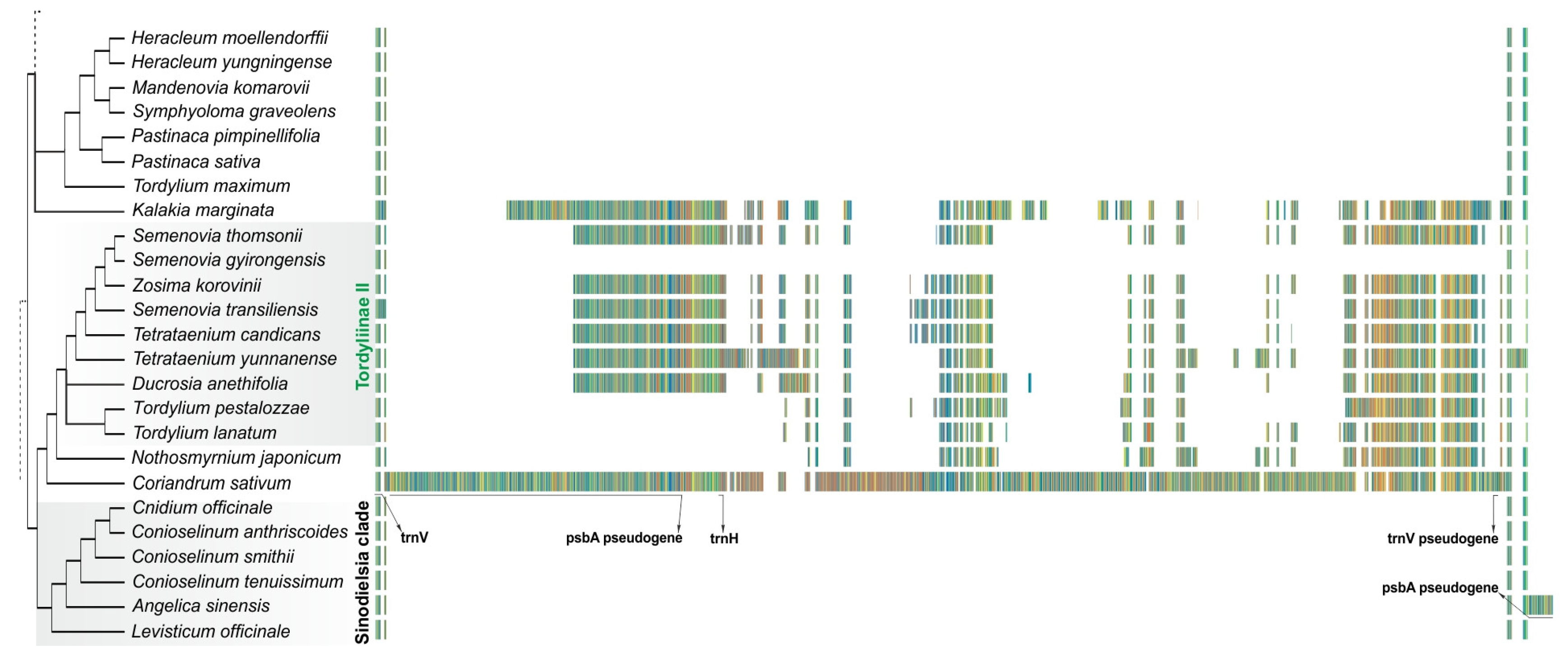
2.4. Distribution of Insertion in the trnV-rrn16 Spacer across Tordylieae and Its Allies
3. Materials and Methods
3.1. Plant Material and DNA Extraction
3.2. Genome Sequencing, Plastome Assembly, and psbA-trnH Insertion Survey
3.3. Phylogenetic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mower, J.P.; Vickrey, T.L. Structural Diversity Among Plastid Genomes of Land Plants. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2018; Volume 85, pp. 263–292. ISBN 978-0-12-813457-3. [Google Scholar]
- Ruhlman, T.A.; Jansen, R.K. Plastid Genomes of Flowering Plants: Essential Principles. In Chloroplast Biotechnology; Humana: New York, NY, USA, 2021; pp. 3–47. [Google Scholar]
- Ruhlman, T.A.; Jansen, R.K. Aberration or Analogy? The Atypical Plastomes of Geraniaceae. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2018; Volume 85, pp. 223–262. ISBN 978-0-12-813457-3. [Google Scholar]
- Wicke, S.; Naumann, J. Molecular Evolution of Plastid Genomes in Parasitic Flowering Plants. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2018; Volume 85, pp. 315–347. ISBN 978-0-12-813457-3. [Google Scholar]
- Goulding, S.E.; Wolfe, K.H.; Olmstead, R.G.; Morden, C.W. Ebb and Flow of the Chloroplast Inverted Repeat. MGG Mol. Gen. Genet. 1996, 252, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Logacheva, M.D.; Krinitsina, A.A.; Belenikin, M.S.; Khafizov, K.; Konorov, E.A.; Kuptsov, S.V.; Speranskaya, A.S. Comparative Analysis of Inverted Repeats of Polypod Fern (Polypodiales) Plastomes Reveals Two Hypervariable Regions. BMC Plant Biol. 2017, 17, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruhlman, T.; Lee, S.-B.; Jansen, R.K.; Hostetler, J.B.; Tallon, L.J.; Town, C.D.; Daniell, H. Complete Plastid Genome Sequence of Daucus Carota: Implications for Biotechnology and Phylogeny of Angiosperms. BMC Genom. 2006, 7, 222. [Google Scholar] [CrossRef] [PubMed]
- Downie, S.R.; Peery, R.M.; Jansen, R.K. Another First for the Apiaceae: Evidence for Mitochondrial DNA Transfer into the Plastid Genome. J. Fac. Pharm. Istanb. Univ. 2015, 44, 131–144. [Google Scholar]
- Downie, S.R.; Jansen, R.K. A Comparative Analysis of Whole Plastid Genomes from the Apiales: Expansion and Contraction of the Inverted Repeat, Mitochondrial to Plastid Transfer of DNA, and Identification of Highly Divergent Noncoding Regions. Syst. Bot. 2015, 40, 336–351. [Google Scholar] [CrossRef]
- Samigullin, T.H.; Logacheva, M.D.; Terenteva, E.I.; Degtjareva, G.V.; Vallejo-Roman, C.M. Plastid Genome of Seseli Montanum: Complete Sequence and Comparison with Plastomes of Other Members of the Apiaceae Family. Biochem. Mosc. 2016, 81, 981–985. [Google Scholar] [CrossRef]
- Yuan, C.; Zhong, W.; Mou, F.; Gong, Y.; Pu, D.; Ji, P.; Huang, H.; Yang, Z.; Zhang, C. The Complete Chloroplast Genome Sequence and Phylogenetic Analysis of Chuanminshen (Chuanminshen Violaceum Sheh et Shan). Physiol. Mol. Biol. Plants 2017, 23, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Yue, M.; Niu, C.; Ma, X.-F.; Li, Z.-H. Comparative Analysis of the Complete Chloroplast Genome of Four Endangered Herbals of Notopterygium. Genes 2017, 8, 124. [Google Scholar] [CrossRef]
- Spooner, D.M.; Ruess, H.; Iorizzo, M.; Senalik, D.; Simon, P. Entire Plastid Phylogeny of the Carrot Genus (Daucus, Apiaceae): Concordance with Nuclear Data and Mitochondrial and Nuclear DNA Insertions to the Plastid. Am. J. Bot. 2017, 104, 296–312. [Google Scholar] [CrossRef] [Green Version]
- Samigullin, T.H.; Logacheva, M.D.; Degtjareva, G.V.; Terentieva, E.I.; Vallejo-Roman, C.M. Complete Plastid Genome of Critically Endangered Plant Prangos Trifida (Apiaceae: Apioideae). Conserv. Genet. Resour. 2018, 10, 847–849. [Google Scholar] [CrossRef]
- Mustafina, F.U.; Yi, D.; Choi, K.; Shin, C.H.; Tojibaev, K.S.; Downie, S.R. A Comparative Analysis of Complete Plastid Genomes from Prangos Fedtschenkoi and Prangos Lipskyi (Apiaceae). Ecol. Evol. 2018, 9, 364–377. [Google Scholar] [CrossRef] [PubMed]
- Park, I.; Yang, S.; Kim, W.J.; Noh, P.; Lee, H.O.; Moon, B.C. The Complete Chloroplast Genome of Cnidium Officinale Makino. Mitochondrial DNA Part B Resour. 2018, 3, 490–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Wang, X.-F.; Cao, D.; Niu, J.-F.; Wang, Z.-Z. The Complete Chloroplast Genome Sequence of Angelica Tsinlingensis (Apioideae). Mitochondrial DNA Part B Resour. 2018, 3, 480–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, I.; Yang, S.; Kim, W.; Song, J.-H.; Lee, H.-S.; Lee, H.; Lee, J.-H.; Ahn, S.-N.; Moon, B. Sequencing and Comparative Analysis of the Chloroplast Genome of Angelica Polymorpha and the Development of a Novel Indel Marker for Species Identification. Molecules 2019, 24, 1038. [Google Scholar] [CrossRef] [Green Version]
- Kang, L.; Yu, Y.; Zhou, S.-D.; He, X.-J. Sequence and Phylogenetic Analysis of Complete Plastid Genome of a Medicinal Plant Heracleum Moellendorffii. Mitochondrial DNA Part B 2019, 4, 1251–1252. [Google Scholar] [CrossRef] [Green Version]
- Kang, L.; Xie, D.; Xiao, Q.; Peng, C.; Yu, Y.; He, X. Sequencing and Analyses on Chloroplast Genomes of Tetrataenium Candicans and Two Allies Give New Insights on Structural Variants, DNA Barcoding and Phylogeny in Apiaceae Subfamily Apioideae. PeerJ 2019, 7, e8063. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Q.-Y.; Feng, T.; Yu, Y.; Luo, Q.; He, X.-J. The Complete Chloroplast Genome of Semenovia Gyirongensis (Tribe Tordylieae, Apiaceae). Mitochondrial DNA Part B 2019, 4, 1863–1864. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Yu, Y. The Complete Chloroplast Genome of Cryptotaenia Japonica. Mitochondrial DNA Part B 2019, 4, 1650–1651. [Google Scholar] [CrossRef] [Green Version]
- Gou, W.; Jia, S.-B.; Price, M.; Guo, X.-L.; Zhou, S.-D.; He, X.-J. Complete Plastid Genome Sequencing of Eight Species from Hansenia, Haplosphaera and Sinodielsia (Apiaceae): Comparative Analyses and Phylogenetic Implications. Plants 2020, 9, 1523. [Google Scholar] [CrossRef]
- Ren, T.; Li, Z.-X.; Xie, D.-F.; Gui, L.-J.; Peng, C.; Wen, J.; He, X.-J. Plastomes of Eight Ligusticum Species: Characterization, Genome Evolution, and Phylogenetic Relationships. BMC Plant Biol. 2020, 20, 519. [Google Scholar] [CrossRef]
- Wang, M.; Wang, X.; Sun, J.; Wang, Y.; Ge, Y.; Dong, W.; Yuan, Q.; Huang, L. Phylogenomic and Evolutionary Dynamics of Inverted Repeats across Angelica Plastomes. BMC Plant Biol. 2021, 21, 26. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Xie, D.-F.; Price, M.; Ren, T.; Deng, Y.-Q.; Gui, L.-J.; Guo, X.-L.; He, X.-J. Backbone Phylogeny and Evolution of Apioideae (Apiaceae): New Insights from Phylogenomic Analyses of Plastome Data. Mol. Phylogenet. Evol. 2021, 161, 107183. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.; Sha, X.; Xiong, M.; Zhong, W.; Wei, Y.; Li, M.; Tao, S.; Mou, F.; Peng, F.; Zhang, C. Uncovering Dynamic Evolution in the Plastid Genome of Seven Ligusticum Species Provides Insights into Species Discrimination and Phylogenetic Implications. Sci. Rep. 2021, 11, 8844. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.-Y.; Feng, T.; Luo, Q.; He, X.-J. The Complete Chloroplast Genome of Semenovia Thomsonii (Tordylieae: Apiaceae), a New Record from Xizang, China. Mitochondrial DNA Part B Resour. 2021, 6, 1911–1913. [Google Scholar] [CrossRef]
- Zhu, A.; Guo, W.; Gupta, S.; Fan, W.; Mower, J.P. Evolutionary Dynamics of the Plastid Inverted Repeat: The Effects of Expansion, Contraction, and Loss on Substitution Rates. New Phytol. 2016, 209, 1747–1756. [Google Scholar] [CrossRef] [Green Version]
- Plunkett, G.M.; Downie, S.R. Expansion and Contraction of the Chloroplast Inverted Repeat in Apiaceae Subfamily Apioideae. Syst. Bot. 2000, 25, 648. [Google Scholar] [CrossRef]
- Downie, S.R.; Spalik, K.; Katz-Downie, D.S.; Reduron, J.-P. Major Clades within Apiaceae Subfamily Apioideae as Inferred by Phylogenetic Analysis of nrDNA ITS Sequences. Plant Divers. Evol. 2010, 128, 111–136. [Google Scholar] [CrossRef] [Green Version]
- Plunkett, G.M.; Pimenov, M.G.; Reduron, J.-P.; Kljuykov, E.V.; van Wyk, B.-E.; Ostroumova, T.A.; Henwood, M.J.; Tilney, P.M.; Spalik, K.; Watson, M.F.; et al. Apiaceae. In Flowering Plants. Eudicots; Springer: Cham, Switzerland, 2018; pp. 9–206. [Google Scholar]
- Koch, W.D.J. Generum Tribuumque Plantarum Umbelliferarum Nova Dispositio. Nova Acta Acad. Caesareae Leopold. Carol. Ger. Nat. Curiosorum 1824, 12, 55–156. [Google Scholar]
- Pimenov, M.G.; Leonov, M.V. The Genera of the Umbelliferae. A Nomenclator; Royal Botanic Gardens, Kew: London, UK, 1993; ISBN 978-0-947643-58-4. [Google Scholar]
- Downie, S.R.; Katz-Downie, D.S.; Watson, M.F. A Phylogeny of the Flowering Plant Family Apiaceae Based on Chloroplast DNA rpl16 and rpoC1 Intron Sequences: Towards a Suprageneric Classification of Subfamily Apioideae. Am. J. Bot. 2000, 87, 273–292. [Google Scholar] [CrossRef] [Green Version]
- Downie, S.R.; Watson, M.F.; Spalik, K.; Katz-Downie, D.S. Molecular Systematics of Old World Apioideae (Apiaceae): Relationships among Some Members of Tribe Peucedaneae Sensu Lato, the Placement of Several Island-Endemic Species, and Resolution within the Apioid Superclade. Can. J. Bot. 2000, 78, 506–528. [Google Scholar] [CrossRef]
- Downie, S.R.; Plunkett, G.M.; Watson, M.F.; Spalik, K.; Katz-Downie, D.S.; Valiejo-Roman, C.M.; Terentieva, E.I.; Troitsky, A.V.; Lee, B.-Y.; Lahham, J.; et al. Tribes and Clades within Apiaceae Subfamily Apioideae: The Contribution of Molecular Data. Edinb. J. Bot. 2001, 58, 301–330. [Google Scholar] [CrossRef]
- Spalik, K.; Downie, S.R. Intercontinental Disjunctions in Cryptotaenia (Apiaceae, Oenantheae): An Appraisal Using Molecular Data. J. Biogeogr. 2007, 34, 2039–2054. [Google Scholar] [CrossRef]
- Ajani, Y.; Ajani, A.; Cordes, J.M.; Watson, M.F.; Downie, S.R. Phylogenetic Analysis of nrDNA ITS Sequences Reveals Relationships within Five Groups of Iranian Apiaceae Subfamily Apioideae. Taxon 2008, 57, 383–401. [Google Scholar] [CrossRef]
- Winter, P.J.D.; Magee, A.R.; Phephu, N.; Tilney, P.M.; Downie, S.R.; van Wyk, B.-E. A New Generic Classification for African Peucedanoid Species (Apiaceae). Taxon 2008, 57, 347–364. [Google Scholar] [CrossRef]
- Zhou, J.; Peng, H.; Downie, S.R.; Liu, Z.-W.; Gong, X. A Molecular Phylogeny of Chinese Apiaceae Subfamily Apioideae Inferred from Nuclear Ribosomal DNA Internal Transcribed Spacer Sequences. Taxon 2008, 57, 402–416. [Google Scholar] [CrossRef]
- Logacheva, M.D.; Valiejo-Roman, C.M.; Pimenov, M.G. ITS Phylogeny of West Asian Heracleum Species and Related Taxa of Umbelliferae–Tordylieae W.D.J.Koch, with Notes on Evolution of Their psbA-trnH Sequences. Plant Syst. Evol. 2008, 270, 139–157. [Google Scholar] [CrossRef]
- Magee, A.R.; van Wyk, B.-E.; Tilney, P.M.; Downie, S.R. Generic Delimitations and Relationships of the Cape Genera Capnophyllum, Dasispermum, and Sonderina, the North African Genera Krubera and Stoibrax, and a New Monotypic Genus of the Subfamily Apioideae (Apiaceae). Syst. Bot. 2009, 34, 580–594. [Google Scholar] [CrossRef]
- Logacheva, M.D.; Valiejo-Roman, C.M.; Degtjareva, G.V.; Stratton, J.M.; Downie, S.R.; Samigullin, T.H.; Pimenov, M.G. A Comparison of nrDNA ITS and ETS Loci for Phylogenetic Inference in the Umbelliferae: An Example from Tribe Tordylieae. Mol. Phylogenet. Evol. 2010, 57, 471–476. [Google Scholar] [CrossRef]
- Yu, Y.; Downie, S.R.; He, X.; Deng, X.; Yan, L. Phylogeny and Biogeography of Chinese Heracleum (Apiaceae Tribe Tordylieae) with Comments on Their Fruit Morphology. Plant Syst. Evol. 2011, 296, 179–203. [Google Scholar] [CrossRef]
- Banasiak, Ł.; Piwczyński, M.; Uliński, T.; Downie, S.R.; Watson, M.F.; Shakya, B.; Spalik, K. Dispersal Patterns in Space and Time: A Case Study of Apiaceae Subfamily Apioideae. J. Biogeogr. 2013, 40, 1324–1335. [Google Scholar] [CrossRef]
- Mousavi, S.; Mozaffarian, V.; Mummenhoff, K.; Downie, S.R.; Zarre, S. An Updated Lineage-Based Tribal Classification of Apiaceae Subfamily Apioideae with Special Focus on Iranian Genera. Syst. Biodivers. 2021, 19, 89–109. [Google Scholar] [CrossRef]
- Clarkson, J.J.; Zuntini, A.R.; Maurin, O.; Downie, S.R.; Plunkett, G.M.; Nicolas, A.N.; Smith, J.F.; Feist, M.A.E.; Gutierrez, K.; Malakasi, P.; et al. A Higher-Level Nuclear Phylogenomic Study of the Carrot Family (Apiaceae). Am. J. Bot. 2021, 108, 1252–1269. [Google Scholar] [CrossRef] [PubMed]
- Downie, S.R.; Ramanath, S.; Katz-Downie, D.S.; Llanas, E. Molecular Systematics of Apiaceae Subfamily Apioideae: Phylogenetic Analyses of Nuclear Ribosomal DNA Internal Transcribed Spacer and Plastid rpoC1 Intron Sequences. Am. J. Bot. 1998, 85, 563–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doğru-Koca, A.; Bagheri, A.; Moradi, A. Investigations on the Phylogenetic Position of the Ditypic Genus Froriepia Reveal Yildirimlia, a New Genus of Apiaceae. Taxon 2020, 69, 1259–1272. [Google Scholar] [CrossRef]
- Wang, W.-C.; Chen, S.-Y.; Zhang, X.-Z. Chloroplast Genome Evolution in Actinidiaceae: clpP Loss, Heterogenous Divergence and Phylogenomic Practice. PLoS ONE 2016, 11, e0162324. [Google Scholar] [CrossRef]
- Sharpe, R.M.; Williamson-Benavides, B.; Edwards, G.E.; Dhingra, A. Methods of Analysis of Chloroplast Genomes of C3, Kranz Type C4 and Single Cell C4 Photosynthetic Members of Chenopodiaceae. Plant Methods 2020, 16, 119. [Google Scholar] [CrossRef]
- de Santana Lopes, A.; Pacheco, T.G.; dos Santos, K.G.; Vieira, L.d.N.; Guerra, M.P.; Nodari, R.O.; de Souza, E.M.; de Oliveira Pedrosa, F.; Rogalski, M. The Linum usitatissimum L. Plastome Reveals Atypical Structural Evolution, New Editing Sites, and the Phylogenetic Position of Linaceae within Malpighiales. Plant Cell Rep. 2018, 37, 307–328. [Google Scholar] [CrossRef]
- Chen, H.; Shao, J.; Zhang, H.; Jiang, M.; Huang, L.; Zhang, Z.; Yang, D.; He, M.; Ronaghi, M.; Luo, X.; et al. Sequencing and Analysis of Strobilanthes Cusia (Nees) Kuntze Chloroplast Genome Revealed the Rare Simultaneous Contraction and Expansion of the Inverted Repeat Region in Angiosperm. Plant Sci. 2018, 324. [Google Scholar] [CrossRef] [Green Version]
- Peery, R. Understanding Angiosperm Genome Interactions and Evolution: Insights from Sacred Lotus (Nelumbo Nucifera) and the Carrot Family (Apiaceae). Ph.D. Thesis, University of Illinois at Urbana-Champaign, Champaign, IL, USA, 2015. [Google Scholar]
- Jansen, R.K.; Ruhlman, T.A. Plastid Genomes of Seed Plants. In Genomics of Chloroplasts and Mitochondria; Bock, R., Knoop, V., Eds.; Springer: Dordrecht, The Netherlands, 2012; Volume 35, pp. 103–126. ISBN 978-94-007-2919-3. [Google Scholar]
- Hoot, S.; Palmer, J. Structural Rearrangements, Including Parallel Inversions, within the Chloroplast Genome of Anemone and Related Genera. J. Mol. Evol. 1994, 38, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-L.; Jansen, R.K.; Chumley, T.W.; Kim, K.-J. Gene Relocations within Chloroplast Genomes of Jasminum and Menodora (Oleaceae) Are Due to Multiple, Overlapping Inversions. Mol. Biol. Evol. 2007, 24, 1161–1180. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.-J.; Choi, K.-S.; Jansen, R.K. Two Chloroplast DNA Inversions Originated Simultaneously During the Early Evolution of the Sunflower Family (Asteraceae). Mol. Biol. Evol. 2005, 22, 1783–1792. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.E.; Rousseau-Gueutin, M.; Cordonnier, S.; Lima, O.; Michon-Coudouel, S.; Naquin, D.; de Carvalho, J.F.; Aïnouche, M.; Salmon, A.; Aïnouche, A. The First Complete Chloroplast Genome of the Genistoid Legume Lupinus Luteus: Evidence for a Novel Major Lineage-Specific Rearrangement and New Insights Regarding Plastome Evolution in the Legume Family. Ann. Bot. 2014, 113, 1197–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, E.N.; Ruhlman, T.A.; Sabir, J.S.M.; Hajrah, N.H.; Alharbi, N.S.; Al-Malki, A.L.; Bailey, C.D.; Jansen, R.K. Plastid Genome Sequences of Legumes Reveal Parallel Inversions and Multiple Losses of rps16 in Papilionoids. J. Syst. Evol. 2015, 53, 458–468. [Google Scholar] [CrossRef]
- Charboneau, J.L.M.; Cronn, R.C.; Liston, A.; Wojciechowski, M.F.; Sanderson, M.J. Plastome Structural Evolution and Homoplastic Inversions in Neo-Astragalus (Fabaceae). Genome Biol. Evol. 2021, 13, evab215. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.S.; Son, O.; Park, S. The Chloroplast Genome of Elaeagnus Macrophylla and trnH Duplication Event in Elaeagnaceae. PLoS ONE 2015, 10, e0138727. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zuo, Y.; Zhu, X.; Liao, S.; Ma, J. Complete Chloroplast Genomes and Comparative Analysis of Sequences Evolution among Seven Aristolochia (Aristolochiaceae) Medicinal Species. Int. J. Mol. Sci. 2019, 20, 1045. [Google Scholar] [CrossRef] [Green Version]
- Cai, Z.; Penaflor, C.; Kuehl, J.V.; Leebens-Mack, J.; Carlson, J.E.; dePamphilis, C.W.; Boore, J.L.; Jansen, R.K. Complete Plastid Genome Sequences of Drimys, Liriodendron, and Piper: Implications for the Phylogenetic Relationships of Magnoliids. BMC Evol. Biol. 2006, 6, 77. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Liu, G.-M.; Zhang, Z.-R.; Corlett, R.T.; Yu, W.-B. Characteristics of the Complete Chloroplast Genome Sequences of Stylidium Debile and Stylidium Petiolare (Stylidiaceae). Mitochondrial DNA Part B Resour. 2021, 6, 3134–3136. [Google Scholar] [CrossRef]
- Bai, H.-R.; Oyebanji, O.; Zhang, R.; Yi, T.-S. Plastid Phylogenomic Insights into the Evolution of Subfamily Dialioideae (Leguminosae). Plant Divers. 2021, 43, 27–34. [Google Scholar] [CrossRef]
- Graham, S.W.; Reeves, P.A.; Burns, A.C.E.; Olmstead, R.G. Microstructural Changes in Noncoding Chloroplast DNA: Interpretation, Evolution, and Utility of Indels and Inversions in Basal Angiosperm Phylogenetic Inference. Int. J. Plant Sci. 2000, 161, S83–S96. [Google Scholar] [CrossRef] [Green Version]
- Borsch, T.; Quandt, D. Mutational Dynamics and Phylogenetic Utility of Noncoding Chloroplast DNA. Plant Syst. Evol. 2009, 282, 169–199. [Google Scholar] [CrossRef]
- Escobari, B.; Borsch, T.; Quedensley, T.S.; Gruenstaeudl, M. Plastid Phylogenomics of the Gynoxoid Group (Senecioneae, Asteraceae) Highlights the Importance of Motif-Based Sequence Alignment amid Low Genetic Distances. Am. J. Bot. 2021, 108, 2235–2256. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-J.; Lee, H.-L. Widespread Occurrence of Small Inversions in the Chloroplast Genomes of Land Plants. Mol. Cells 2005, 19, 104–113. [Google Scholar] [PubMed]
- Aberer, A.J.; Krompass, D.; Stamatakis, A. Pruning Rogue Taxa Improves Phylogenetic Accuracy: An Efficient Algorithm and Webservice. Syst. Biol. 2013, 62, 162–166. [Google Scholar] [CrossRef] [Green Version]
- Al-Eisawi, D.; Jury, S.L. A Taxonomic Revision of the Genus Tordylium L. (Apiaceae). Bot. J. Linn. Soc. 1988, 97, 357–403. [Google Scholar] [CrossRef]
- Samigullin, T.; Vallejo-Roman, C.; Degtjareva, G.; Terentieva, E. Structural Rearrangements in Plastid Genomes of Apiaceae as Phylogenetic Markers. BIO Web Conf. 2021, 38, 107. [Google Scholar] [CrossRef]
- Calviño, C.I.; Teruel, F.E.; Downie, S.R. The Role of the Southern Hemisphere in the Evolutionary History of Apiaceae, a Mostly North Temperate Plant Family. J. Biogeogr. 2016, 43, 398–409. [Google Scholar] [CrossRef]
- Zhou, J.; Gao, Y.; Wei, J.; Liu, Z.-W.; Downie, S.R. Molecular Phylogenetics of Ligusticum (Apiaceae) Based on nrDNA ITS Sequences: Rampant Polyphyly, Placement of the Chinese Endemic Species, and a Much-Reduced Circumscription of the Genus. Int. J. Plant Sci. 2019, 181, 306–323. [Google Scholar] [CrossRef]
- Wang, R.-J.; Cheng, C.-L.; Chang, C.-C.; Wu, C.-L.; Su, T.-M.; Chaw, S.-M. Dynamics and Evolution of the Inverted Repeat-Large Single Copy Junctions in the Chloroplast Genomes of Monocots. BMC Evol. Biol. 2008, 8, 36. [Google Scholar] [CrossRef] [Green Version]
- Maréchal, A.; Brisson, N. Recombination and the Maintenance of Plant Organelle Genome Stability. New Phytol. 2010, 186, 299–317. [Google Scholar] [CrossRef]
- Hastings, P.J.; Ira, G.; Lupski, J.R. A Microhomology-Mediated Break-Induced Replication Model for the Origin of Human Copy Number Variation. PLoS Genet. 2009, 5, e1000327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, T.; Huq, E.; Herrin, D.L. Microhomology-Mediated and Nonhomologous Repair of a Double-Strand Break in the Chloroplast Genome of Arabidopsis. Proc. Natl. Acad. Sci. USA 2010, 107, 13954–13959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmonds, S.E.; Smith, J.F.; Davidson, C.; Buerki, S. Phylogenetics and Comparative Plastome Genomics of Two of the Largest Genera of Angiosperms, Piper and Peperomia (Piperaceae). Mol. Phylogenet. Evol. 2021, 163, 107229. [Google Scholar] [CrossRef] [PubMed]
- Haberle, R.C.; Fourcade, H.M.; Boore, J.L.; Jansen, R.K. Extensive Rearrangements in the Chloroplast Genome of Trachelium Caeruleum Are Associated with Repeats and tRNA Genes. J. Mol. Evol. 2008, 66, 350–361. [Google Scholar] [CrossRef]
- Lyskov, D.; Kljuykov, E.; Terentieva, E.; Ukrainskaja, U.; Samigullin, T. The End of the Long Road: Iranian Endemic Seseli Elbursense (Apiaceae) Has Taken Its Place in the Genus Semenovia. Phytotaxa 2020, 435, 1–15. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Leung, H.C.M.; Yiu, S.M.; Chin, F.Y.L. IDBA-UD: A de Novo Assembler for Single-Cell and Metagenomic Sequencing Data with Highly Uneven Depth. Bioinformatics 2012, 28, 1420–1428. [Google Scholar] [CrossRef] [Green Version]
- Milne, I.; Stephen, G.; Bayer, M.; Cock, P.J.A.; Pritchard, L.; Cardle, L.; Shaw, P.D.; Marshall, D. Using Tablet for Visual Exploration of Second-Generation Sequencing Data. Brief. Bioinform. 2013, 14, 193–202. [Google Scholar] [CrossRef]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and Accurate Annotation of Organelle Genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef]
- Carver, T.; Berriman, M.; Tivey, A.; Patel, C.; Böhme, U.; Barrell, B.G.; Parkhill, J.; Rajandream, M.-A. Artemis and ACT: Viewing, Annotating and Comparing Sequences Stored in a Relational Database. Bioinformatics 2008, 24, 2672–2676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) Version 1.3.1: Expanded Toolkit for the Graphical Visualization of Organellar Genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K. MAFFT: A Novel Method for Rapid Multiple Sequence Alignment Based on Fast Fourier Transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Anisimova, M.; Gil, M.; Dufayard, J.-F.; Dessimoz, C.; Gascuel, O. Survey of Branch Support Methods Demonstrates Accuracy, Power, and Robustness of Fast Likelihood-Based Approximation Schemes. Syst. Biol. 2011, 60, 685–699. [Google Scholar] [CrossRef]
- Minh, B.Q.; Hahn, M.W.; Lanfear, R. New Methods to Calculate Concordance Factors for Phylogenomic Datasets. Mol. Biol. Evol. 2020, 37, 2727–2733. [Google Scholar] [CrossRef]
- Wolfe, K.H.; Li, W.H.; Sharp, P.M. Rates of Nucleotide Substitution Vary Greatly among Plant Mitochondrial, Chloroplast, and Nuclear DNAs. Proc. Natl. Acad. Sci. USA 1987, 84, 9054–9058. [Google Scholar] [CrossRef] [Green Version]
- Crotty, S.M.; Minh, B.Q.; Bean, N.G.; Holland, B.R.; Tuke, J.; Jermiin, L.S.; Haeseler, A.V. GHOST: Recovering Historical Signal from Heterotachously Evolved Sequence Alignments. Syst. Biol. 2019, 69, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Akaike, H. A New Look at the Statistical Model Identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Swofford, D. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4; Sinauer Associates: Sunderland, MA, USA, 2003. [Google Scholar]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Xia, X. DAMBE5: A Comprehensive Software Package for Data Analysis in Molecular Biology and Evolution. Mol. Biol. Evol. 2013, 30, 1720–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, J.; Yu, Y.; Xie, D.-F.; Peng, C.; Liu, Q.; Zhou, S.-D.; He, X.-J. A Transcriptome-Based Study on the Phylogeny and Evolution of the Taxonomically Controversial Subfamily Apioideae (Apiaceae). Ann. Bot. 2020, 125, 937–953. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Taxon | Size (bp) | LSC Size (bp) | SSC Size (bp) | IR Size (bp) | GC Content (%) | Total Genes | Protein-Coding Genes (Pseudogenes) | tRNA Genes | rRNA Genes | GenBank Accession Number | Mean Coverage Depth (X) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Dasispermum suffruticosum (P.J.Bergius) B.L.Burtt | 144,902 | 91,681 | 16,931 | 18,145 | 35.2 | 126 | 82 (2) | 36 | 8 | OL839263 | 61 |
| Ducrosia anethifolia Boiss. | 141,148 | 98,931 | 17,523 | 12,347 | 37.4 | 122 | 79 (2) | 35 | 8 | OL839264 | 25.8 |
| Kalakia marginata (Boiss.) Alava | 142,155 | 98,758 | 17,497 | 12,950 | 37.5 | 122 | 79 (2) | 35 | 8 | OL839265 | 23.7 |
| Mandenovia komarovii (Manden.) Alava | 149,345 | 92,332 | 17,485 | 19,764 | 37.4 | 127 | 82 (2) | 37 | 8 | OL839266 | 121 |
| Notobubon galbanum (L.) Magee | 147,466 | 93,641 | 17,443 | 18,191 | 37.5 | 126 | 82 (2) | 36 | 8 | OL839267 | 51.9 |
| Pastinaca pimpinellifolia M.Bieb. | 149,758 | 92,242 | 17,654 | 19,931 | 37.4 | 127 | 82 (2) | 37 | 8 | NC_027450 | 82.8 |
| Symphyoloma graveolens C.A.Mey. | 149,245 | 92,159 | 17,516 | 19,785 | 37.5 | 127 | 82 (2) | 37 | 8 | OL839268 | 115 |
| Tordylium lanatum Boiss. | 143,402 | 94,157 | 17,521 | 15,862 | 37.2 | 124 | 81 (2) | 35 | 8 | OL839269 | 89.7 |
| Tordylium maximum L. | 150,103 | 91,637 | 17,676 | 20,395 | 37.3 | 126 | 82 (2) | 36 | 8 | OL839270 | 80.6 |
| Tordylium pestalozzae Boiss. | 141,830 | 99,355 | 17,488 | 12,493 | 37.1 | 122 | 79 (1) | 35 | 8 | OL839271 | 33.1 |
| Zosima korovinii Pimenov | 141,644 | 99,620 | 17,498 | 12,263 | 37.4 | 122 | 79 (2) | 35 | 8 | OL839272 | 173 |
| Branch | CDS, PP/aBayes | Unpartitioned, PP/aBayes | 4 Partitions, PP/aBayes | 3 Partitions, PP/aBayes | 4 Partitions + Heterotachy, aBayes/BS | sCF/sDF1/sDF2 | Putative Synapomorphies |
|---|---|---|---|---|---|---|---|
| 1 | 1/1 | 1/1 | 1/1 | 1/1 | 1/97 | 43.5/28.6/28.9 | 9 |
| 2 | 0.97/0.95 | 1/0.99 | 0.99/ | 1/1 | 1/93 | 36.4/33.2/30.4 | 2 |
| 3 | 1/1 | 1/1 | 1/1 | 1/1 | 1/100 | 42.6/32.8/25.6 | 20 |
| 4 | 0.99/0.99 | 1/1 | 1/1 | 1/1 | 1/99 | 41.4/30.5/28.1 | 12 |
| 5 | 1/1 | 1/1 | 1/1 | 1/1 | 1/100 | 42.6/31.9/25.5 | 6 |
| 6 | 0.82/0.84 | 1/1 | 1/1 | 1/1 | 1/99 | 47.3/16.2/36.5 | 46 |
| Species Name | Voucher Number | Locality | Collector, Date |
|---|---|---|---|
| Dasispermum suffruticosum | MW0589014 | Republic of South Africa, 34°21′ S, 18°55′ E | Pimenov et al., 12 January 2003 |
| Ducrosia anethifolia | MW0744172 | Iran, prov. Fars, 29°41′ N, 52°45′ E | Pimenov et al., 8 June 2001 |
| Kalakia marginata | E №3567 | Iran, Iranshakr | Lamond, 1 June 1971 |
| Mandenovia komarovii | MW0701533 | Russia, Daghestan, left bank of the river Avarskoe Kojsu, near Tlyarata village | Pimenov et al., 15 August 1978 |
| Notobubon galbanum | MW0589116 | Republic of South Africa, 34°05′ S, 18°25′ E | Pimenov et al., 13 January 2003 |
| Pastinaca pimpinellifolia | * | Russia, North Caucasus | Kljuykov et al., 5 August 2005 |
| Symphyoloma graveolens | MW0700962 | Russia, Daghestan, Andijski distr., Danukh village | Amirhanov, 5 August 1989 |
| Tordylium lanatum | * | Turkey, Antalya, Elmali | Pimenov et al., 11 July 2007 |
| Tordylium maximum | * | Turkey, prov. Kastamonu, Kure-Inebolu, Ercisler dere | Pimenov et al., 21 August 2008 |
| Tordylium pestalozzae | MW0745191 | Turkey, Ephesus C1 Izmir: between Ephesus and Mariamane, 37°55′ N, 27°20′ E | Pimenov and Kljuykov, 27 May 1995 |
| Zosima korovinii | MW0864736 | Kyrgyzstan, bank of river At-Bashi, Baybichetau range, tract Kara-Terek | Pimenov and Kljuykov, 1 August 1987 |
| Purpose | Primer: Name and Sequence |
|---|---|
| Insertion in assembled genomes | trnV-rrn16_U: AGTTCGAGCCTGATTATCC |
| trnV-rrn16_L: ATTACTTATAGCTTCCTTGTT | |
| Survey of 16 Tordylieae species | trnV-rrn16_U: AGTTCGAGCCTGATTATCC |
| trnV-trnH_L: CAATCCACTGCCTTGATCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samigullin, T.; Logacheva, M.; Terentieva, E.; Degtjareva, G.; Pimenov, M.; Valiejo-Roman, C. Plastid Phylogenomic Analysis of Tordylieae Tribe (Apiaceae, Apioideae). Plants 2022, 11, 709. https://doi.org/10.3390/plants11050709
Samigullin T, Logacheva M, Terentieva E, Degtjareva G, Pimenov M, Valiejo-Roman C. Plastid Phylogenomic Analysis of Tordylieae Tribe (Apiaceae, Apioideae). Plants. 2022; 11(5):709. https://doi.org/10.3390/plants11050709
Chicago/Turabian StyleSamigullin, Tahir, Maria Logacheva, Elena Terentieva, Galina Degtjareva, Michael Pimenov, and Carmen Valiejo-Roman. 2022. "Plastid Phylogenomic Analysis of Tordylieae Tribe (Apiaceae, Apioideae)" Plants 11, no. 5: 709. https://doi.org/10.3390/plants11050709