
Prescriptions for the Control of a Clonal Invasive Species Using Demographic Models

 , , and
, , and
Abstract
:1. Introduction
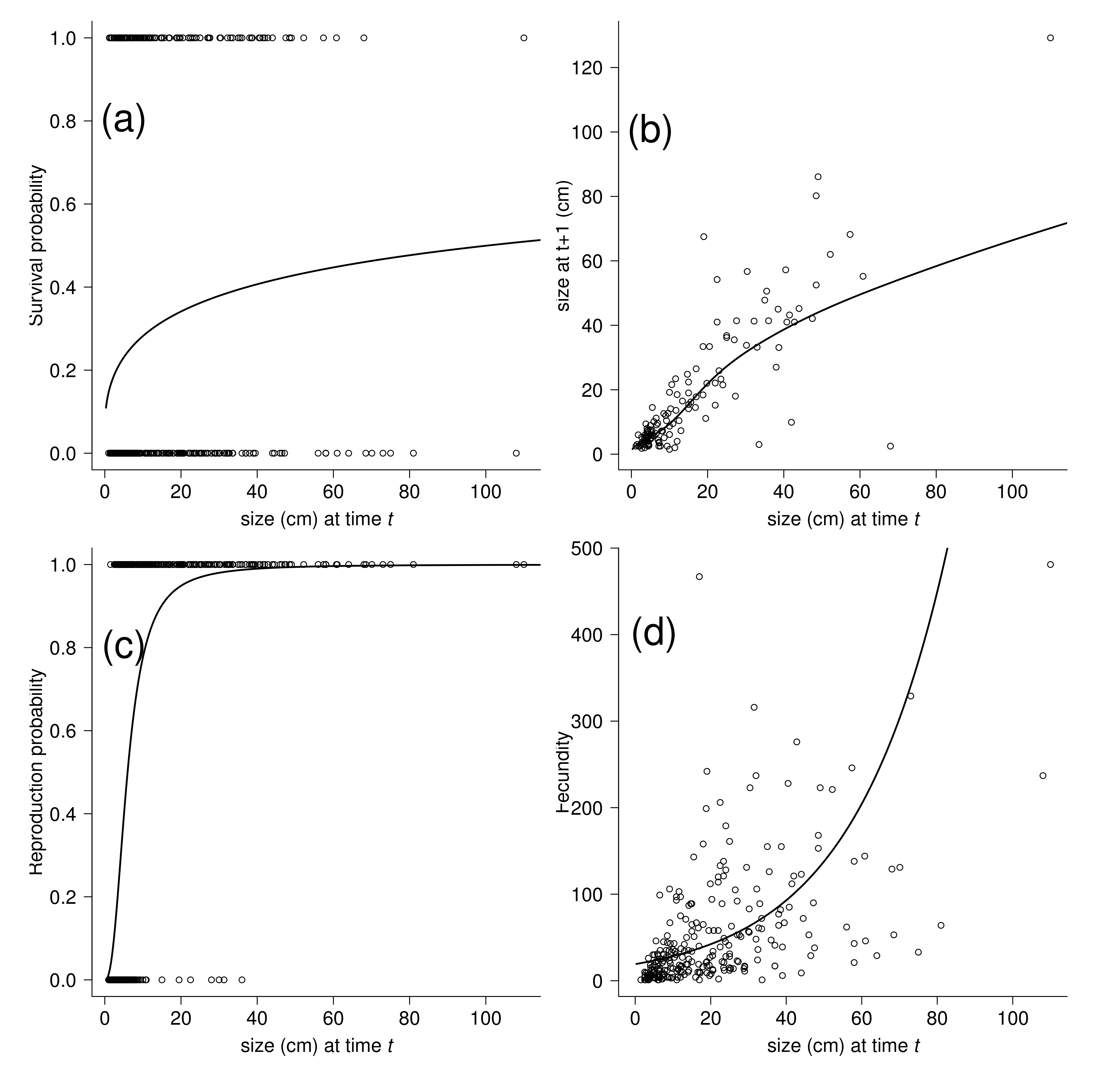
2. Results
Vital Rates
3. Discussion
Management Scenarios
4. Materials and Methods
4.1. Species
4.2. Population Dynamics
4.3. Population Growth Rate
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Genovesi, H.; Carboneras, C.; Vilà, M.; Walton, P. EU adopts innovative legislation on invasive species: A step towards a global response to biological invasions. Biol. Invasions 2015, 117, 1307–1311. [Google Scholar] [CrossRef]
- Pimentel, D.; Zuniga, R.; Morrison, D. Update on the environmental and economic costs associated with aline-invasive species in the United States. Ecol. Econ. 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Bradshaw, C.; Leroy, B.; Bellard, C.; Roiz, D.; Albert, C.; Fournier, A.; Babet-Massin, M.; Salles, J.; Simard, F.; Courchamp, F. Massive yet grossly underestimated global costs of invasive insects. Nat. Commun. 2016, 7, 12986. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, B.D.; Broadhurst, L.M. The economic cost of managing invasive species in Australia. NeoBiota 2016, 31, 18. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, T.M.; Pyšek, P.; Bacher, S.; Carlton, J.T.; Duncan, R.P.; Jarosik, V.; Wilson, J.R.U.; Richardson, D.M. A unified framework for biological invasions. Trends Ecol. Evol. 2011, 26, 333–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evangelista, P.H.; Kumar, S.; Stohlgren, T.J.; Jarnevich, C.S.; Crall, A.W.; Norman, J.B.; Barnett, D.T. Modelling invasion for a habitat generalist and a specialist plant species. Divers. Distrib. 2008, 14, 808–817. [Google Scholar] [CrossRef]
- Duncan, R.P.; Blackburn, T.M.; Rossinelli, S.; Bacher, S. Quantifying invasion risk: The relationship between establishment probability and founding population size. Methods Ecol. Evol. 2014, 5, 1255–1263. [Google Scholar] [CrossRef] [Green Version]
- Lockwood, J.L.; Cassey, P.; Blackburn, T. The role of propagule pressure in explaining species invasions. Trends Ecol. Evol. 2005, 20, 223–228. [Google Scholar] [CrossRef]
- Richardson, D.M.; Allsop, N.; D’Antonio, C.M.; Milton, S.J.; Rejmánek, M. Plant invasions—The role of mutualisms. Biol. Rev. 2000, 75, 65–93. [Google Scholar] [CrossRef]
- Richards, C.; Bossdorf, O.; Muth, N.; Gurevitch, J.; Pigliucci, M. Jack of all trades, master of some? On the role of phenotypic plasticity in plant invasions. Ecol. Lett. 2006, 9, 981–993. [Google Scholar] [CrossRef] [Green Version]
- Daehler, C.C. Performance comparison of co-occurring native and aliens invasive plants: Implications for conservation and restoration. Annu. Rev. Ecol. Syst. 2003, 34, 183–211. [Google Scholar] [CrossRef]
- Liu, J.; Dong, M.; Miao, S.L.; Li, Z.Y.; Song, M.H.; Wang, R.Q. Invasive alien plants in China: Role of clonality and geographical origin. Biol. Invasions 2006, 8, 1461–1470. [Google Scholar] [CrossRef]
- Pyšek, P. Clonality and plant invasions: Can a trait make a difference. In The Ecology and Evolution of Clonal Plants; de Kroon, H.J., van Groenendael, H., Eds.; Backhuys Publishers: Leiden, The Netherlands, 1997; pp. 405–427. [Google Scholar]
- Lloret, F.; Médial, F.; Brundu, G.; Camarada, J.; Moragues, E.; Rita, J.; Lambdon, P.; Hulme, P. Species attributes and invasion success by alien plants on Mediterranean islands. J. Ecol. 2005, 93, 512–520. [Google Scholar] [CrossRef]
- Ramula, A.; Knight, T.M.; Burns, J.H.; Buckley, Y.M. General guidelines for invasive plant management based in comparative demography of invasive and native plant populations. J. Appl. Ecol. 2008, 45, 1124–1133. [Google Scholar] [CrossRef]
- Jelbert, K.; Buss, D.; McDonald, J.; Townley, S.; Franco, M.; Stott, I.; Jones, O.; Salguero-Gómez, R.; Buckley, Y.; Knight, T.; et al. Demographic amplification is a predictor of invasiveness among plants. Nat. Commun. 2019, 10, 5602. [Google Scholar] [CrossRef]
- Daehler, C.C.; Carino, D.A. Predicting invasive plants: Prospects for a general screening system based on current regional models. Biol. Invasions 2000, 2, 93–102. [Google Scholar] [CrossRef]
- Krivanek, M.; Pyšek, P. Predicting invasions by woody species in a temperate zone: A test of three risk assessment schemes in the Czech Republic (Central Europe). Divers. Distrib. 2006, 12, 319–327. [Google Scholar] [CrossRef]
- Koop, A.L.; Fowler, L.; Newton, L.P.; Caton, B.P. Development and validation of a weed screening tool for the United States. Biol. Invasions 2012, 14, 273–294. [Google Scholar] [CrossRef]
- Suehls, C.M.; Affra, L.; Médail, F. Invasion dynamics of two Carpobrotus (Aizoaceae) taxa on a Mediterranean island: I. Genetic diversity and introgression. Heredity 2004, 93, 31–40. [Google Scholar] [CrossRef]
- Li, S.L.; Ramula, S. Demographic strategies of plant invaders in temporally varying environments. Popul. Ecol. 2015, 57, 373–380. [Google Scholar] [CrossRef]
- Sebert-Cuvillier, E.; Paccaut, F.; Chabrerie, O.; Endels, P.; Goubet, O.; Cecocq, G. Local population dynamics of invasive tree species with a complex life history cycle: A stochastic matrix model. Ecol. Model. 2007, 201, 127–143. [Google Scholar] [CrossRef]
- Williams, J.L.; Auge, H.; Maron, J.L. Testing hypotheses for exotic plant success: Parallel experiments in the native and introduced ranges. Ecology 2010, 91, 1355–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramula, S. Annual mowing has the potential to reduce the invasion of herbaceous Lupinus polyphyllus. Biol. Invasions 2020, 22, 3163–3173. [Google Scholar] [CrossRef]
- Zucaratto, R.; Pires, A.S.; Bergallo, H.G.; de Cássia Quitete Portela, R. Felling the giants: Integral projection models indicate adult management to control an exotic invasive palm. Plant Ecol. 2021, 222, 93–105. [Google Scholar] [CrossRef]
- Kerr, N.Z.; Baxter, P.W.J.; Salguero-Gómez, R.; Wardle, G.M.; Buckley, Y.M. Ecological predictions and risk assessment for alien fishes in North America. J. Appl. Ecol. 2016, 53, 305–316. [Google Scholar] [CrossRef]
- Iles, D.T.; Salguero-Gomez, R.; Adler, P.; Koons, P.B. Linking transient dynamics and life history in biological invasion success. J. Ecol. 2016, 104, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Horvitz, C.C.; Denslow, J.S.; Johnson, T.; Gaoue, O.; Uowolo, A. Unexplained variability among spatial replicates in transient elasticity: Implications for evolutionary ecology and management of invasive species. Popul. Ecol. 2018, 60, 61–75. [Google Scholar] [CrossRef]
- Claridge, K.; Franklin, S. Compensation and plasticity in an invasive plant species. Biol. Invasions 2002, 4, 339–347. [Google Scholar] [CrossRef]
- Aillie, R.; Reshi, Z.; Wafai, B. Demographic plasticity in relation to growth and resource allocation pattern in Anthemis cotula- an alien invasive species in Kashmir Himalaya, India. Appl. Environ. Res. 2005, 4, 63–74. [Google Scholar] [CrossRef]
- Ramula, S. Linking vital rates to invasiveness of a perennial herb. Oecologia 2014, 174, 1255–1264. [Google Scholar] [CrossRef]
- Sultan, S. Phenotypic plasticity for fitness components in Polygonum spcecies of constrasting ecological breadth. Ecology 2001, 82, 328–343. [Google Scholar] [CrossRef] [Green Version]
- Shea, K.; Kelly, D. Estimating biocontrol agent impact with matrix models: Carduus nutans in New Zealand. Ecol. Appl. 1998, 8, 824–832. [Google Scholar] [CrossRef]
- Herrera, I.; Chacón, N.; Flores, S.; Benzo, D.; Marínez, J.; García, B.; Hernández-Rosas, J.I. La planta exótica Kalanchoe daigremontiana incrementa el reservorio y flujo de carbono en el suelo. Interciencia 2011, 36, 937–942. [Google Scholar]
- Buhle, E.; Margolis, M.; Ruesink, J. Bang for buck: Cost-effective control of invasive species with different life histories. Ecol. Econ. 2005, 52, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Easterling, M.R.; Ellner, S.P.; Dixon, P.M. Size-specific sensitivity: Applying a new structured population model. Ecology 2000, 81, 694–708. [Google Scholar] [CrossRef]
- Gonzáles, E.; Child, D.; Quintana-Ascencio, P.F.; Salguero-Gómez, R. Integral projection models. In Demographic Methods across the Tree of Life; Salguero-Gómez, R., Gamelon, M., Eds.; Oxford University Press: Oxford, UK, 2021; pp. 181–195. [Google Scholar]
- Quintana-Ascencio, P.F.; Koontz, S.M.; Smith, S.A.; Sclater, V.L.; David, A.S.; Menges, E.S. Predicting landscape-level distribution and abundance: Integrating demography, fire, elevation and landscape habitat configuration. J. Ecol. 2018, 106, 2395–2408. [Google Scholar] [CrossRef]
- Ramula, S.; Rees, M.; Buckley, Y.M. Integral projection models perform better for small demographic data sets than matrix population models: A case study of two perennial herbs. J. Appl. Ecol. 2009, 46, 1048–1053. [Google Scholar] [CrossRef]
- Quintana-Ascencio, P.F.; Menges, E.S.; Cook, G.S.; Ehrlé, J.; Afkhami, M. Drivers of demography: Past challenges and a promise for a changed future. In Demographic Methods across the Tree of Life; Salguero-Gómez, R., Gamelon, M., Eds.; Oxford University Press: Oxford, UK, 2021; pp. 115–129. [Google Scholar]
- Hannan-Jones, M.A.; Playford, J. The biology of Australian Weeds 40. Bryophyllum Salisb. species. Plant Prot. Q. 2002, 17, 42–57. [Google Scholar]
- Guillot-Ortiz, D.; Lopez-Pujol, J.; Lumbreras, E.L.; Puche, C. Kalanchoe daigremontiana Raym. Hamet & H. Perrier. ’Iberian Coast’. Bouteloua 2015, 21, 35–48. [Google Scholar]
- Guerra-Garcia, A.; Golubov, J.; Mandujano, M.C. Invasion of Kalanchoe by clonal spread. Biol. Invasions 2015, 44, 1–8. [Google Scholar] [CrossRef]
- Herrera, I.; Hernández, M.J.; Lampo, M.; Nassar, J. Plantlet recruitment is the key demographic transition in invasion by Kalanchoe daigremontiana. Popul. Ecol. 2012, 54, 225–237. [Google Scholar] [CrossRef]
- Williams, J.L.; Crone, E.E. The impact of invasive grasses on the population growth of Anemone patens, a long lived native forb. Ecology 2006, 87, 3200–3208. [Google Scholar] [CrossRef] [Green Version]
- Ren, M.X.; Zhang, D.G.; Zhang, D.Y. Random amplified polymorphic DNA markers reveal low genetic variation and a single dominant genotype in Eichhornia crassipes populations throughout China. Weed Res. 2005, 45, 236–244. [Google Scholar] [CrossRef]
- Geng, Y.; Pan, X.; Xu, C.; Zhang, W.; Li, B.; Chen, J.; Lu, B.; Song, Z. Phenotypic plasticity rather than locally adapted ecotypes allow the invasiivse alligator weed to colonize a wide range of habitats. Biol. Invasions 2007, 9, 245–256. [Google Scholar] [CrossRef]
- Lambertini, C.; Riis, T.; Olesen, B.; Clayton, J.S.; Sorrell, B.K.; Brix, H. Genetic diversity in three invasive clonal aquatic species in New Zealand. BMC Genet. 2010, 11, 52. [Google Scholar] [CrossRef] [Green Version]
- Pichancourt, J.P.; van Klinken, R.D. Phenotypic plasticity influences the size, shape and dynamics of the geographic distribution of an invasive plant. PLoS ONE 2012, 7, e32323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera, I.; Nassar, N. Reproductive and recruitment traits as indicators of the invasive potential of Kalanchoe daigremontiana (Crassulaceae) and Stapelia gigantea (Apocynaceae) in a Neotropical arid zone. J. Arid. Environ. 2009, 73, 978–986. [Google Scholar] [CrossRef]
- Schutzenhofer, M.R.; Valone, T.J.; Knight, T.M. Herbivory and population dynamics of invasive and native Lespedeza. Oecologia 2009, 161, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Hahn, M.A.; Buckley, Y.M.; Müller-Scharer, H. Increased population growth rate in invasive polyploid Centaurea stoebe in a common garden. Ecol. Lett. 2012, 15, 947–954. [Google Scholar] [CrossRef]
- Münzbergová, Z.; Hadincová, V.; Wild, J.; Kindlmannová, J. Variation in the contribution of different life stages to population growth as a key factor in the invasion success of Pinus strobus. PLoS ONE 2015, 8, e56953. [Google Scholar]
- Tenhumberg, B.; Suwa, T.; Tyre, A.J.; Russell, L.; Louda, S.M. Integral projection models show exotic thistle is more limited than native thistle by ambient competition and herbivory. Ecosphere 2015, 6, 1–18. [Google Scholar] [CrossRef]
- Van Kleuken, G.; Johnson, S.D. Effects of self-compatiblility on the distribution of invasive European plants in North America. Conserv. Biol. 2007, 21, 1537–1544. [Google Scholar]
- Knight, T.; Havens, K.; Vitt, P. Will the use of less defund cultivars reduce the invasiveness of perennial plants? BioScience 2011, 61, 816–822. [Google Scholar] [CrossRef]
- González de León, S.; Herrera, I.; Guevara, R. Mating system, population growth, and management for Kalanchoe pinnata in an invaded seasonal dry tropical forest. Ecol. Evol. 2016, 6, 4541–4550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SEMARNAT. Acuerdo por el que se Determina la Lista de Especies Exóticas Invasoras para México; Diario Oficial de la Federación: Ciudad de México, Mexico, 2016. [Google Scholar]
- Eggli, U. Crassulaceae; Springer: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Johnson, M.A. The origin of the foliar pseudobulbils in Kalanchoe daigremontiana. J. Torrey Bot. Soc. 1934, 61, 355–366. [Google Scholar] [CrossRef]
- Guerra-García, A. Evaluación del éxito clonal en una especie invasora Kalanchoe delagoensis. Bachelor Thesis, Facultad de Ciencias, Universidad Nacional Autónoma de México, Ciudad de México, Mexico, 2011. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Ellner, S.P.; Rees, M. Integral projection models for species with complex demography. Am. Nat. 2006, 167, 410–428. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Stasinopoulos, D.; Rigby, R. Generalized additive models for location scale and shape (GAMLSS) in R. J. Stat. Softw. 2007, 23, 1–46. [Google Scholar] [CrossRef] [Green Version]
- Jongejans, E.; Jorritsma-Wienk, L.D.; Becker, U.; Dostal, P.; Milden, M.; de Kroon, H. Region versus site variation in the population dynamics of three short-lived perennials. J. Ecol. 2010, 98, 279–289. [Google Scholar] [CrossRef]
- Guerra-García, A.; Barrales-Alcalá, D.; Argueta-Guzmán, M.; Cruz, A.; Mandujano, M.C.; Arévalo-Ramírez, J.A.; Milligan, B.G.; Golubov, J. Biomass allocation, plantlet survival, and chemical control of the invasive Chandelier plant (Kalanchoe delagoensis) (Crassulaceae). Invasive Plant Sci. Manag. 2018, 11, 33–39. [Google Scholar] [CrossRef]
- Griffith, A. Perturbation approaches for integral projection models. Oikos 2017, 126, 1675–1686. [Google Scholar] [CrossRef]
- Stubben, C.J.; Milligan, B.G. Estimating and analyzing demographic models using the popbio package in R. J. Stat. Softw. 2007, 22, 1–23. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Treatment 1 | Proportion | Pmatrix | Fmatrix | |
|---|---|---|---|---|
| of Plantlet Survival | (Confidence Interval) | |||
| Control 2 | 0.12 | 1.28 (1.094–1.640) | 0.556 | 0.444 |
| 25% Water | 0.14 | 1.37 (1.228–1.836) | 0.545 | 0.455 |
| 50% Water | 0.25 | 1.81 (1.531–2.869) | 0.502 | 0.498 |
| 100% Water | 0.84 | 3.51 (2.234–5.142) | 0.399 | 0.601 |
| G/2-4D | 0 | 0.29 (0.282–0.415) | 1.000 | 0.000 |
| 2-4D | 0.01 | 0.55 (0.245–0.717) | 0.776 | 0.224 |
| Other herbicides | >0.33 | 2.08 (1.860–2.410) | 0.480 | 0.520 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arroyo-Cosultchi, G.; Golubov, J.; Solórzano, J.V.; Mandujano, M.C. Prescriptions for the Control of a Clonal Invasive Species Using Demographic Models. Plants 2022, 11, 689. https://doi.org/10.3390/plants11050689
Arroyo-Cosultchi G, Golubov J, Solórzano JV, Mandujano MC. Prescriptions for the Control of a Clonal Invasive Species Using Demographic Models. Plants. 2022; 11(5):689. https://doi.org/10.3390/plants11050689
Chicago/Turabian StyleArroyo-Cosultchi, Gabriel, Jordan Golubov, Jonathan V. Solórzano, and Maria C. Mandujano. 2022. "Prescriptions for the Control of a Clonal Invasive Species Using Demographic Models" Plants 11, no. 5: 689. https://doi.org/10.3390/plants11050689