
Role of Melatonin in Apple Fruit during Growth and Ripening: Possible Interaction with Ethylene


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Experiment 1: Evolution of the Growth and Ripening Parameters, and Changes in the Melatonin Content
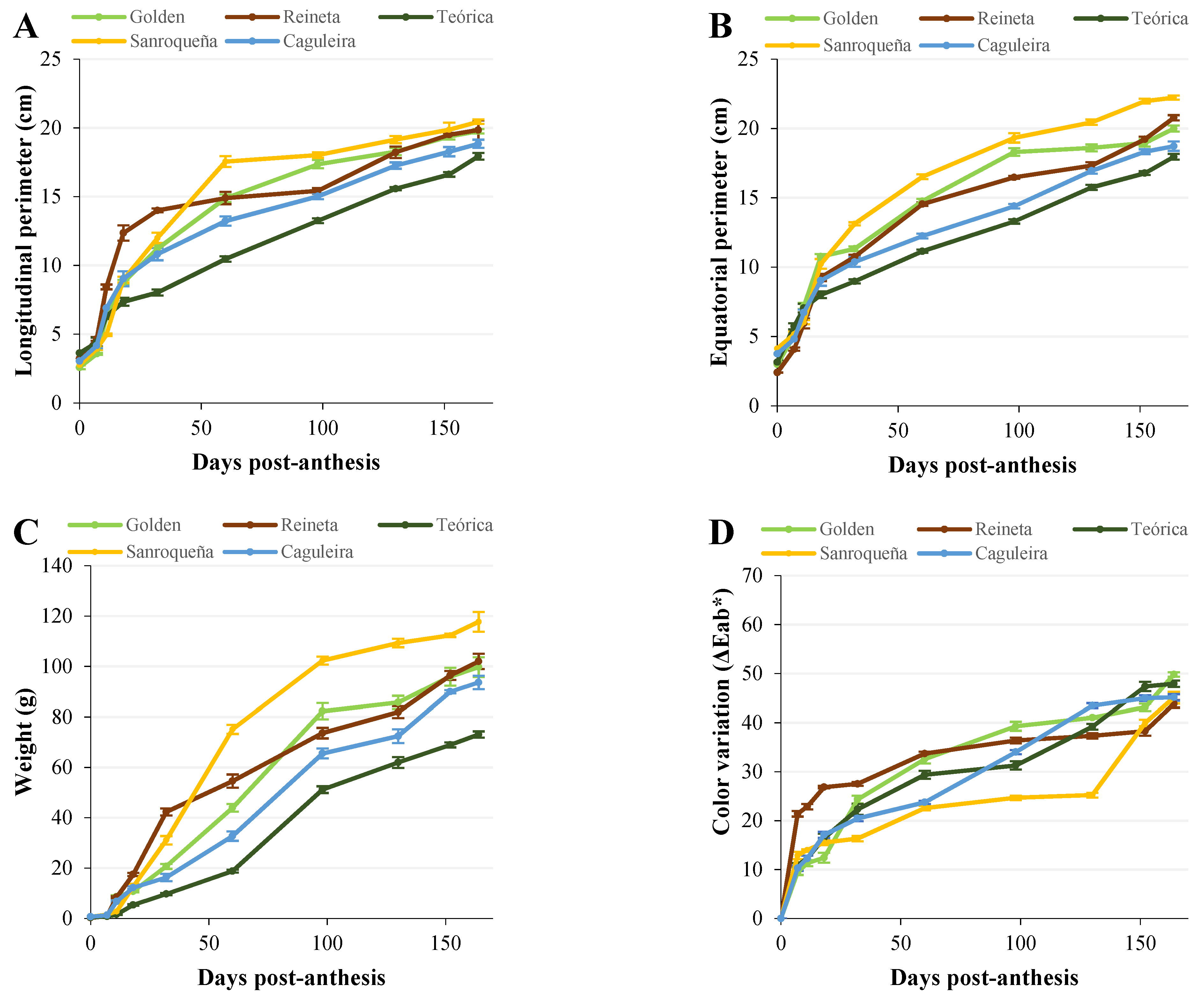
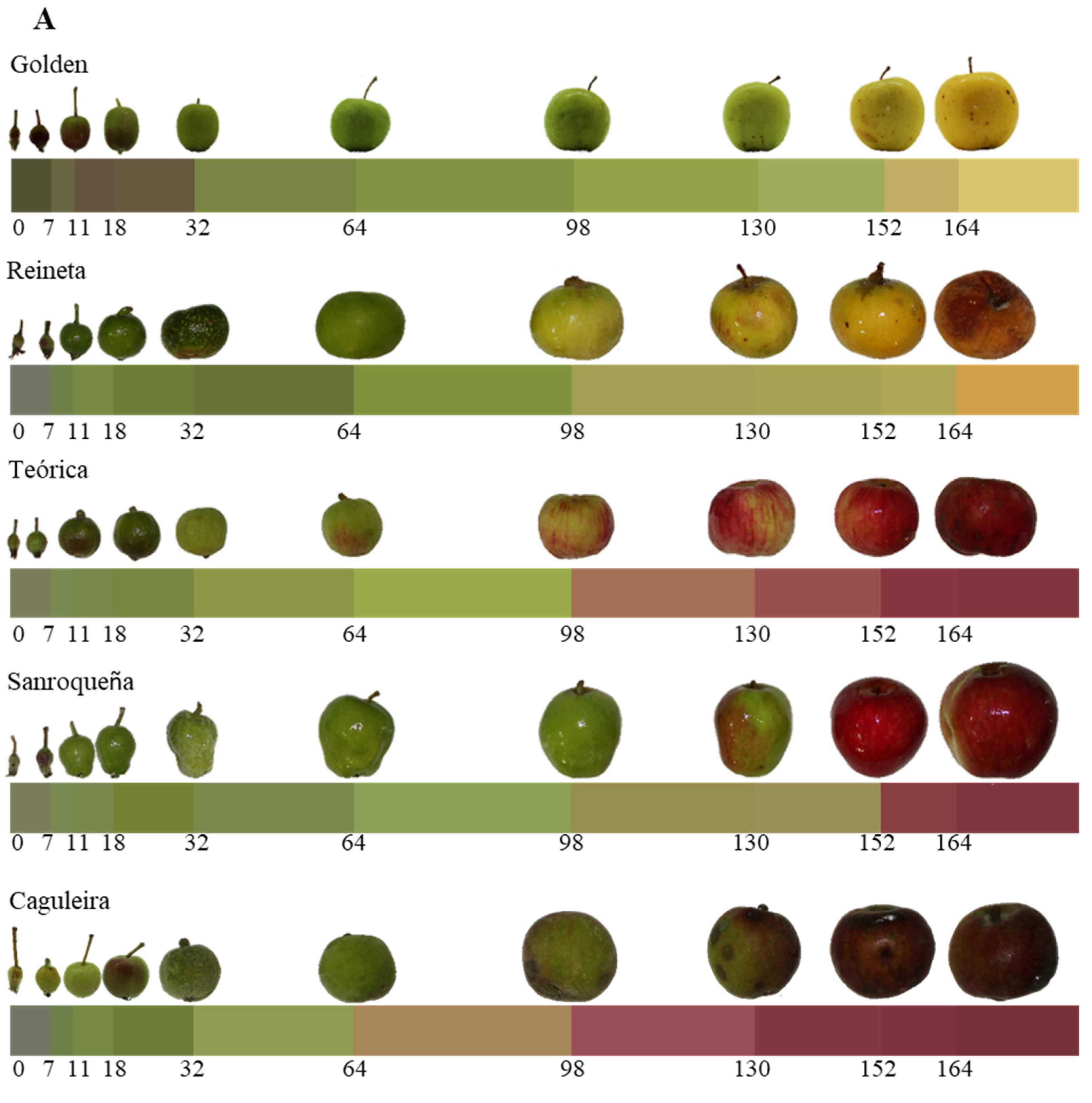
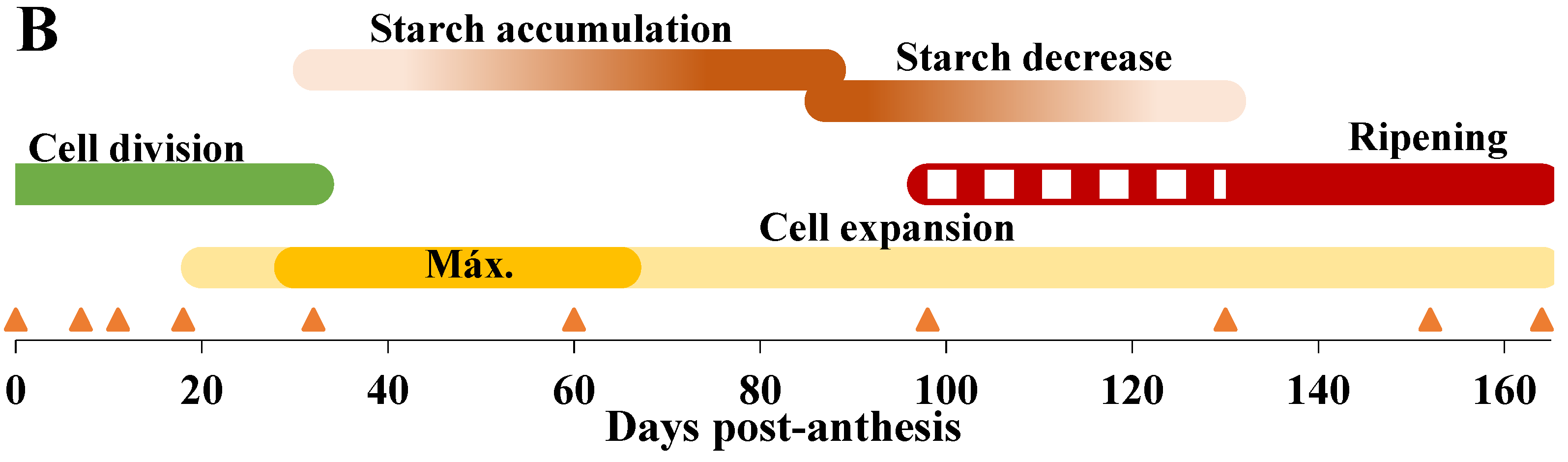
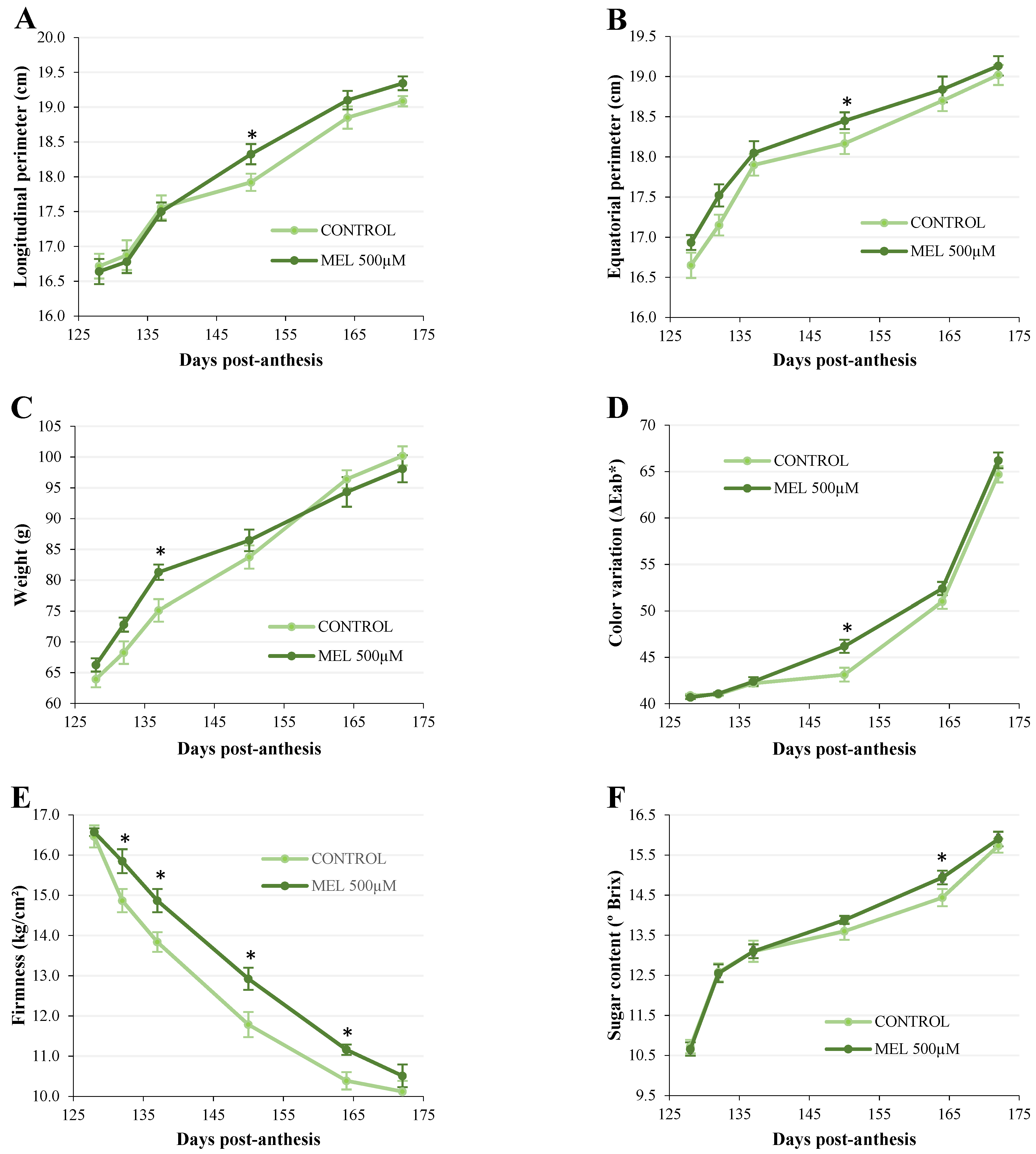
2.1.1. Analysis of the Physiological Parameters of Apples
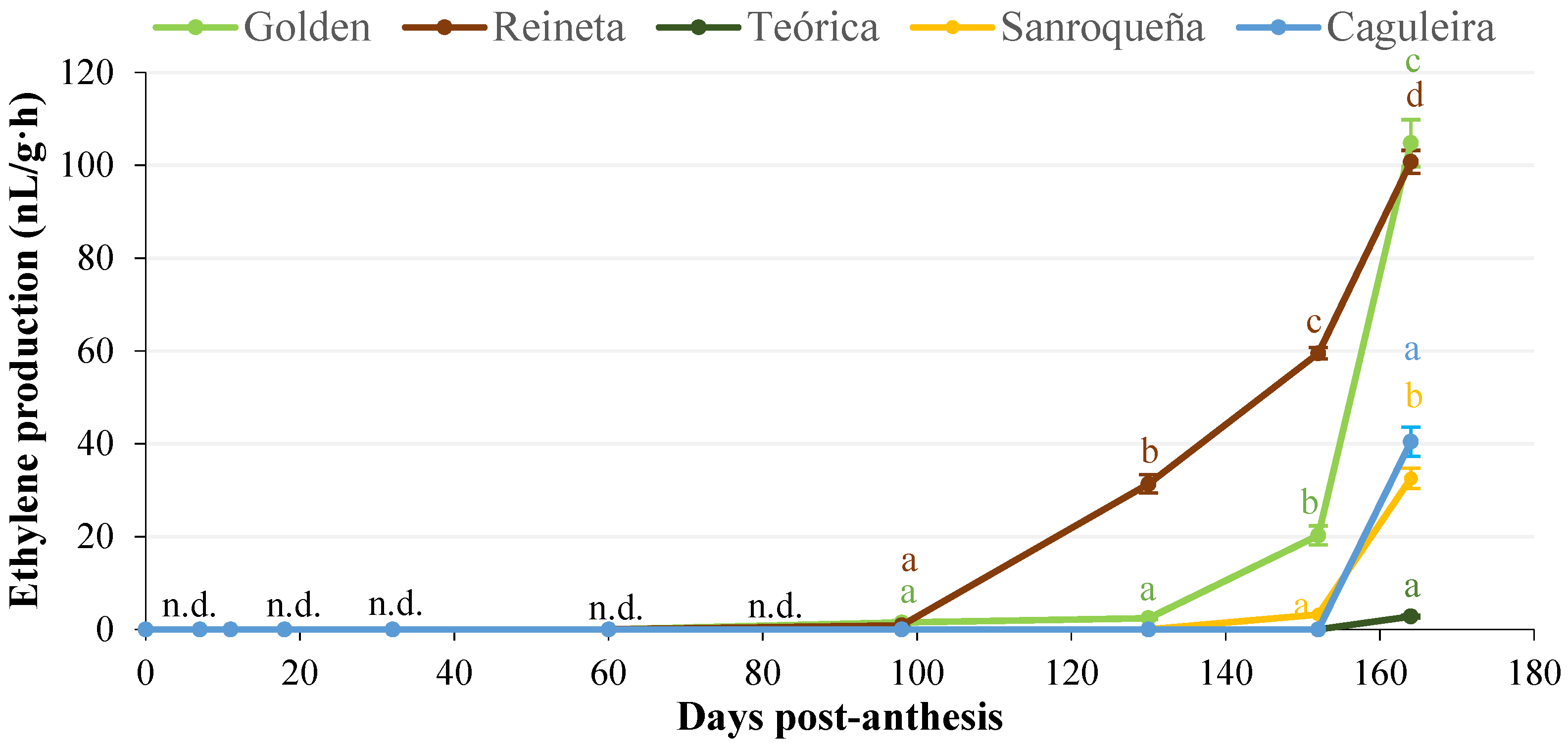
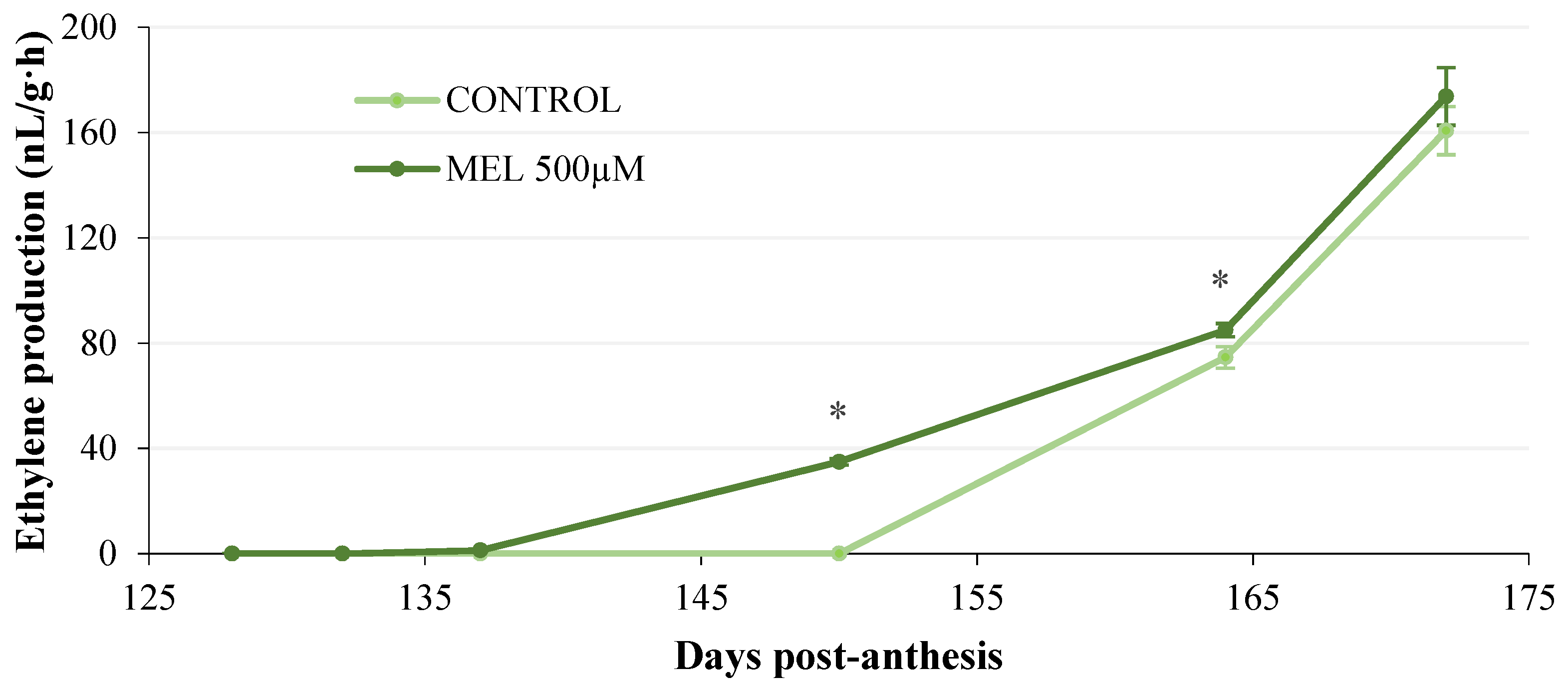
2.1.2. Evolution of Ethylene Production
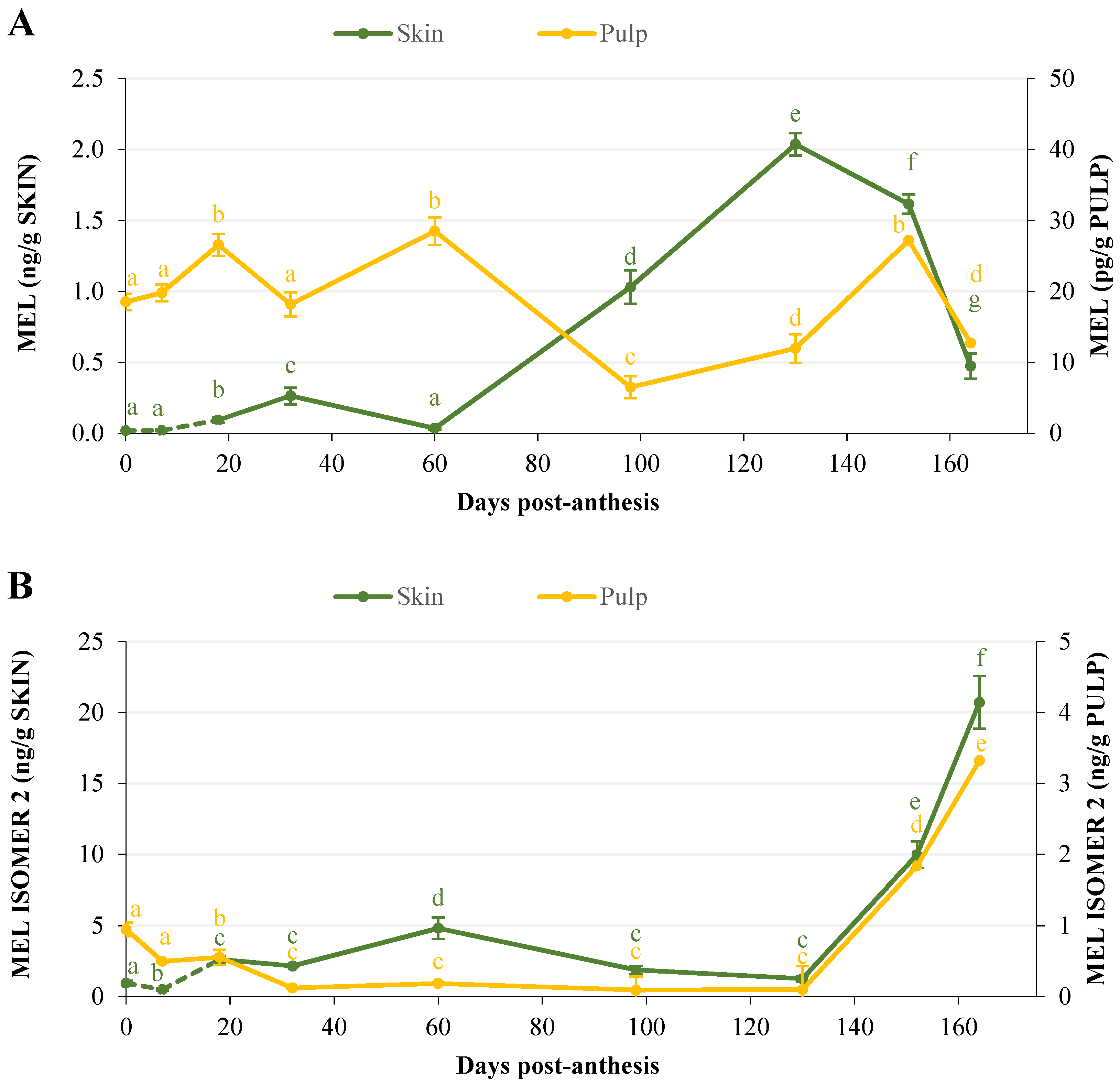
2.1.3. Evolution of Melatonin and Melatonin Isomer 2 Contents
2.2. Experiment 2: Effect of the In Vivo Application of Melatonin to the Tree upon Fruit Ripening
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Design
4.1.1. Plant Material
4.1.2. Experiment 1: Evolution of the Apple Fruit Growth and Ripening Parameters, and Changes in Melatonin Content
4.1.3. Experiment 2: Effect of the In Vivo Application of Melatonin to the Tree on Fruit Ripening
4.2. Determination of the Apple Development and Ripening Parameters
4.3. Analysis of Ethylene Production
4.4. Determination of Melatonin and Its Isomer Content
4.4.1. Sample Extraction
4.4.2. Analysis of Melatonin and Its Isomer by HPLC
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khanizadeh, S.; Tsao, R.; Rekika, D.; Yang, R.; Charles, M.T.; Vasantha Rupasinghe, H.P. Polyphenol Composition and Total Antioxidant Capacity of Selected Apple Genotypes for Processing. J. Food Comp. Anal. 2008, 21, 396–401. [Google Scholar] [CrossRef]
- Goulas, V.; Kourdoulas, P.; Makris, F.; Theodorou, M.; Fellman, J.K.; Manganaris, G.A. Comparative Polyphenolic Antioxidant Profile and Quality of Traditional Apple Cultivars as Affected by Cold Storage. Int. J. Food Sci. Technol. 2014, 49, 2037–2044. [Google Scholar] [CrossRef]
- Schempp, H.; Christof, S.; Mayr, U.; Treutter, D. Phenolic Compounds in Juices of Apple Cultivars and Their Relation to Antioxidant Activity. J. Appl. Bot. Food Qual. 2016, 89, 11–20. [Google Scholar] [CrossRef]
- Alberti, A.; Zielinski, A.A.F.; Couto, M.; Judacewski, P.; Mafra, L.I.; Nogueira, A. Distribution of Phenolic Compounds and Antioxidant Capacity in Apples Tissues during Ripening. J. Food Sci. Technol. 2017, 54, 1511–1518. [Google Scholar] [CrossRef] [Green Version]
- Stanger, M.C.; Steffens, C.A.; Soethe, C.; Moreira, M.A.; do Amarante, C.V.T. Phenolic Content and Antioxidant Activity during the Development of ‘Brookfield’ and ‘Mishima’ Apples. J. Agric. Food Chem. 2017, 65, 3453–3459. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, X.; Chen, T.; Ji, Y.; Shi, K.; Wang, L.; Zheng, X.; Kong, J. Melatonin in Apples and Juice: Inhibition of Browning and Microorganism Growth in Apple Juice. Molecules 2018, 23, 521. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Khurana, A.; Sharma, A.K. Role of Plant Hormones and Their Interplay in Development and Ripening of Fleshy Fruits. J. Exp. Bot. 2014, 65, 4561–4575. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.-H.; Janssen, B.-J.; Bieleski, L.R.F.; Atkinson, R.G.; Morris, B.A.M.; Gardner, R.C. Isolating and Characterizing Genes Differentially Expressed Early in Apple Fruit Development. J. Am. Soc. Hortic. Sci. 1997, 122, 752–757. [Google Scholar] [CrossRef] [Green Version]
- Janssen, B.J.; Thodey, K.; Schaffer, R.J.; Alba, R.; Balakrishnan, L.; Bishop, R.; Bowen, J.H.; Crowhurst, R.N.; Gleave, A.P.; Ledger, S. Global Gene Expression Analysis of Apple Fruit Development from the Floral Bud to Ripe Fruit. BMC Plant Biol. 2008, 8, 16. [Google Scholar] [CrossRef] [Green Version]
- Eccher, G.; Ferrero, S.; Populin, F.; Colombo, L.; Botton, A. Apple (Malus domestica L. Borkh) as an Emerging Model for Fruit Development. Plant Biosyst. 2014, 148, 157–168. [Google Scholar] [CrossRef]
- Harada, T.; Kurahashi, W.; Yanai, M.; Wakasa, Y.; Satoh, T. Involvement of Cell Proliferation and Cell Enlargement in Increasing the Fruit Size of Malus Species. Sci. Hortic. 2005, 105, 447–456. [Google Scholar] [CrossRef]
- Li, M.; Feng, F.; Cheng, L. Expression Patterns of Genes Involved in Sugar Metabolism and Accumulation during Apple Fruit Development. PLoS ONE 2012, 7, e33055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gapper, N.E.; McQuinn, R.P.; Giovannoni, J.J. Molecular and Genetic Regulation of Fruit Ripening. Plant Mol. Biol. 2013, 82, 575–591. [Google Scholar] [CrossRef] [PubMed]
- Seymour, G.B.; Østergaard, L.; Chapman, N.H.; Knapp, S.; Martin, C. Fruit Development and Ripening. Annu. Rev. Plant Biol. 2013, 64, 219–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, J.W.; Gunaseelan, K.; Pidakala, P.; Wang, M.; Schaffer, R.J. Co-Ordination of Early and Late Ripening Events in Apples Is Regulated through Differential Sensitivities to Ethylene. J. Exp. Bot. 2009, 60, 2689–2699. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Song, J.; Campbell-Palmer, L.; Fillmore, S.; Zhang, Z. Effect of Ethylene and 1-MCP on Expression of Genes Involved in Ethylene Biosynthesis and Perception during Ripening of Apple Fruit. Postharvest Biol. Technol. 2013, 78, 55–66. [Google Scholar] [CrossRef]
- Busatto, N.; Tadiello, A.; Trainotti, L.; Costa, F. Climacteric Ripening of Apple Fruit Is Regulated by Transcriptional Circuits Stimulated by Cross-Talks between Ethylene and Auxin. Plant Signal Behav. 2017, 12, e1268312. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Tan, D.; Takahashi, A.; Zhong Li, T.; Harada, T. MdERFs, Two Ethylene-Response Factors Involved in Apple Fruit Ripening. J. Exp. Bot. 2007, 58, 3743–3748. [Google Scholar] [CrossRef]
- Costa, F.; Alba, R.; Schouten, H.; Soglio, V.; Gianfranceschi, L.; Serra, S.; Musacchi, S.; Sansavini, S.; Costa, G.; Fei, Z.; et al. Use of Homologous and Heterologous Gene Expression Profiling Tools to Characterize Transcription Dynamics during Apple Fruit Maturation and Ripening. BMC Plant Biol. 2010, 10, 229. [Google Scholar] [CrossRef] [Green Version]
- Carrillo-Vico, A.; Lardone, P.; Álvarez-Sánchez, N.; Rodríguez-Rodríguez, A.; Guerrero, J. Melatonin: Buffering the Immune System. Int. J. Mol. Sci. 2013, 14, 8638–8683. [Google Scholar] [CrossRef] [Green Version]
- Cipolla-Neto, J.; Amaral, F.G. do Melatonin as a Hormone: New Physiological and Clinical Insights. Endocr. Rev. 2018, 39, 990–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Tassel, D.L.; O’Neill, S.D. Putative Regulatory Molecules in Plants: Evaluating Melatonin: Putative Regulators in Plants. J. Pineal Res. 2001, 31, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Iriti, M.; Varoni, E.M.; Vitalini, S. Melatonin in Traditional Mediterranean Diets: Melatonin in Mediterranean Plant Foods. J. Pineal Res. 2010, 49, 101–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawaz, M.A.; Huang, Y.; Bie, Z.; Ahmed, W.; Reiter, R.J.; Niu, M.; Hameed, S. Melatonin: Current Status and Future Perspectives in Plant Science. Front Plant Sci. 2016, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verde, A.; Míguez, J.M.; Leao-Martins, J.M.; Gago-Martínez, A.; Gallardo, M. Melatonin Content in Walnuts and Other Commercial Nuts. Influence of Cultivar, Ripening and Processing (Roasting). J. Food Compos Anal 2022, 105, 104180. [Google Scholar] [CrossRef]
- Tan, D.-X.; Hardeland, R.; Manchester, L.C.; Korkmaz, A.; Ma, S.; Rosales-Corral, S.; Reiter, R.J. Functional Roles of Melatonin in Plants, and Perspectives in Nutritional and Agricultural Science. J. Exp. Bot. 2012, 63, 577–597. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Functions of Melatonin in Plants: A Review. J. Pineal Res. 2015, 59, 133–150. [Google Scholar] [CrossRef] [Green Version]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin in Flowering, Fruit Set and Fruit Ripening. Plant Reprod. 2020, 33, 77–87. [Google Scholar] [CrossRef]
- Mukherjee, S. Recent Advancements in the Mechanism of Nitric Oxide Signaling Associated with Hydrogen Sulfide and Melatonin Crosstalk during Ethylene-Induced Fruit Ripening in Plants. Nitric Oxide 2019, 82, 25–34. [Google Scholar] [CrossRef]
- Posmyk, M.M.; Janas, K.M. Melatonin in Plants. Acta Physiol. Plant 2009, 31, 1. [Google Scholar] [CrossRef]
- Zhai, R.; Liu, J.; Liu, F.; Zhao, Y.; Liu, L.; Fang, C.; Wang, H.; Li, X.; Wang, Z.; Ma, F.; et al. Melatonin Limited Ethylene Production, Softening and Reduced Physiology Disorder in Pear (Pyrus communis L.) Fruit during Senescence. Postharvest Biol. Technol. 2018, 139, 38–46. [Google Scholar] [CrossRef]
- Xu, L.; Yue, Q.; Xiang, G.; Bian, F.; Yao, Y. Melatonin Promotes Ripening of Grape Berry via Increasing the Levels of ABA, H2O2, and Particularly Ethylene. Hortic. Res. 2018, 5, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okazaki, M.; Ezura, H. Profiling of Melatonin in the Model Tomato (Solanum lycopersicum L.) Cultivar Micro-Tom. J. Pineal Res. 2009, 46, 338–343. [Google Scholar] [CrossRef]
- Wang, C.; Yin, L.-Y.; Shi, X.-Y.; Xiao, H.; Kang, K.; Liu, X.-Y.; Zhan, J.-C.; Huang, W.-D. Effect of Cultivar, Temperature, and Environmental Conditions on the Dynamic Change of Melatonin in Mulberry Fruit Development and Wine Fermentation: Melatonin in Mulberry Fruits and Wine. J. Food Sci. 2016, 81, M958–M967. [Google Scholar] [CrossRef] [PubMed]
- Tijero, V.; Muñoz, P.; Munné-Bosch, S. Melatonin as an Inhibitor of Sweet Cherries Ripening in Orchard Trees. Plant Physiol. Biochem. 2019, 140, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Yue, Q.; Bian, F.; Sun, H.; Zhai, H.; Yao, Y. Melatonin Enhances Phenolics Accumulation Partially via Ethylene Signaling and Resulted in High Antioxidant Capacity in Grape Berries. Front Plant Sci. 2017, 8, 1426. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yue, R.; Si, M.; Wu, M.; Cong, L.; Zhai, R.; Yang, C.; Wang, Z.; Ma, F.; Xu, L. Effects of Exogenous Application of Melatonin on Quality and Sugar Metabolism in ‘Zaosu’ Pear Fruit. J. Plant Growth Regul. 2019, 38, 1161–1169. [Google Scholar] [CrossRef]
- Çolak, A.M. Effect of Melatonin and Gibberellic Acid Foliar Application on the Yield and Quality of Jumbo Blackberry Species. Saudi J. Biol. Sci. 2018, 25, 1242–1246. [Google Scholar] [CrossRef]
- Sun, Q.; Zhang, N.; Wang, J.; Zhang, H.; Li, D.; Shi, J.; Li, R.; Weeda, S.; Zhao, B.; Ren, S. Melatonin Promotes Ripening and Improves Quality of Tomato Fruit during Postharvest Life. J. Exp. Bot. 2015, 66, 657–668. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Zhang, Z.K.; Chai, H.K.; Cheng, N.; Yang, Y.; Wang, D.N.; Yang, T.; Cao, W. Melatonin Treatment Delays Postharvest Senescence and Regulates Reactive Oxygen Species Metabolism in Peach Fruit. Postharvest Biol. Technol. 2016, 118, 103–110. [Google Scholar] [CrossRef]
- Hu, W.; Yang, H.; Tie, W.; Yan, Y.; Ding, Z.; Liu, Y.; Wu, C.; Wang, J.; Reiter, R.J.; Tan, D.-X.; et al. Natural Variation in Banana Varieties Highlights the Role of Melatonin in Postharvest Ripening and Quality. J. Agric. Food Chem. 2017, 65, 9987–9994. [Google Scholar] [CrossRef] [PubMed]
- Aghdam, M.S.; Fard, J.R. Melatonin Treatment Attenuates Postharvest Decay and Maintains Nutritional Quality of Strawberry Fruits (Fragaria×anannasa Cv. Selva) by Enhancing GABA Shunt Activity. Food Chem. 2017, 221, 1650–1657. [Google Scholar] [CrossRef] [PubMed]
- Hattori, A.; Migitaka, H.; Iigo, M.; Itoh, M.; Yamamoto, K.; Ohtani-Kaneko, R.; Hara, M.; Suzuki, T.; Reiter, R.J. Identification of Melatonin in Plants and Its Effects on Plasma Melatonin Levels and Binding to Melatonin Receptors in Vertebrates. Biochem. Mol. Biol. Int. 1995, 35, 627–634. [Google Scholar] [PubMed]
- Badria, F.A. Melatonin, Serotonin, and Tryptamine in Some Egyptian Food and Medicinal Plants. J. Med. Food 2002, 5, 153–157. [Google Scholar] [CrossRef]
- Azcón-Bieto, J.; Talón, M. Fundamentos de Fisiología Vegetal; McGraw-Hill Interamericana: New York, NY, USA, 2008. [Google Scholar]
- Prasanna, V.; Prabha, T.N.; Tharanathan, R.N. Fruit Ripening Phenomena—An Overview. Crit. Rev. Food Sci. Nutr. 2007, 47, 1–19. [Google Scholar] [CrossRef]
- Rugkong, A.; McQuinn, R.; Giovannoni, J.J.; Rose, J.K.C.; Watkins, C.B. Expression of Ripening-Related Genes in Cold-Stored Tomato Fruit. Postharvest Biol. Technol. 2011, 61, 1–14. [Google Scholar] [CrossRef]
- Gupta, A.K.; Das, S.; Sahu, P.P.; Mishra, P. Design and Development of IDE Sensor for Naringin Quantification in Pomelo Juice: An Indicator of Citrus Maturity. Food Chem. 2022, 377, 131947. [Google Scholar] [CrossRef]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Trivellini, A.; Francini, A.; Khan, M.I.R. Ethylene Role in Plant Growth, Development and Senescence: Interaction with Other Phytohormones. Front Plant Sci. 2017, 08. [Google Scholar] [CrossRef] [Green Version]
- Vitalini, S.; Gardana, C.; Zanzotto, A.; Simonetti, P.; Faoro, F.; Fico, G.; Iriti, M. The Presence of Melatonin in Grapevine (Vitis vinifera L.) Berry Tissues: Melatonin in Grape Berry Tissues. J. Pineal Res. 2011, 51, 331–337. [Google Scholar] [CrossRef]
- Wang, S.-Y.; Shi, X.-C.; Wang, R.; Wang, H.-L.; Liu, F.; Laborda, P. Melatonin in Fruit Production and Postharvest Preservation: A Review. Food Chem. 2020, 320, 126642. [Google Scholar] [CrossRef]
- Zhao, Y.; Tan, D.-X.; Lei, Q.; Chen, H.; Wang, L.; Li, Q.; Gao, Y.; Kong, J. Melatonin and Its Potential Biological Functions in the Fruits of Sweet Cherry. J. Pineal Res. 2013, 55, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Shen, Y.; Shen, T.; Wang, X.; Zhang, X.; Hu, P.; Liang, D.; Lin, L.; Deng, H.; Wang, J. Melatonin Accumulation in Sweet Cherry and Its Influence on Fruit Quality and Antioxidant Properties. Molecules 2020, 25, 753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Q.; Zhang, N.; Wang, J.; Cao, Y.; Li, X.; Zhang, H.; Zhang, L.; Tan, D.-X.; Guo, Y.-D. A Label-Free Differential Proteomics Analysis Reveals the Effect of Melatonin on Promoting Fruit Ripening and Anthocyanin Accumulation upon Postharvest in Tomato. J. Pineal Res. 2016, 61, 138–153. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.-X.; Hardeland, R.; Manchester, L.C.; Rosales-Corral, S.; Coto-Montes, A.; Boga, J.A.; Reiter, R.J. Emergence of Naturally Occurring Melatonin Isomers and Their Proposed Nomenclature: Naturally Occurring Melatonin Isomers. J. Pineal Res. 2012, 53, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.-X.; Zheng, X.; Kong, J.; Manchester, L.; Hardeland, R.; Kim, S.; Xu, X.; Reiter, R. Fundamental Issues Related to the Origin of Melatonin and Melatonin Isomers during Evolution: Relation to Their Biological Functions. Int. J. Mol. Sci. 2014, 15, 15858–15890. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.-F.; Shi, T.-C.; Song, S.; Zhang, Z.-W.; Fang, Y.-L. Melatonin in Grapes and Grape-Related Foodstuffs: A Review. Food Chem. 2017, 231, 185–191. [Google Scholar] [CrossRef]
- Okatan, V.; Bulduk, I.; Sekara, A.; Colak, A.M.; Kaki, B.; Gundogdu, M. Bioactive Components and Market Quality of Apple (Malus x Domestica Borkh.) Fruits Could Be Effectively Controlled by Trees Pretreatment with Boric Acid, Melatonin and Gibberellic Acid. Fresenius Environ. Bull. 2018, 27, 6933–6944. [Google Scholar]
- International Commission on Illumination. Recommendations on Uniform Color Spaces, Color-Difference Equations, Psychometric Color Terms; CIE Publication; Bureau Central de la CIE: Vienna, Austria, 1978. [Google Scholar]
- de Dios, P.; Matilla, A.J.; Gallardo, M. Flower Fertilization and Fruit Development Prompt Changes in Free Polyamines and Ethylene in Damson Plum (Prunus insititia L.). J. Plant Physiol 2006, 163, 86–97. [Google Scholar] [CrossRef]
- Verde, A.; Míguez, J.M.; Gallardo, M. Melatonin and Related Bioactive Compounds in Commercialized Date Palm Fruits (Phoenix dactylifera L.): Correlation with Some Antioxidant Parameters. Eur. Food Res. Technol. 2019, 245, 51–59. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verde, A.; Míguez, J.M.; Gallardo, M. Role of Melatonin in Apple Fruit during Growth and Ripening: Possible Interaction with Ethylene. Plants 2022, 11, 688. https://doi.org/10.3390/plants11050688
Verde A, Míguez JM, Gallardo M. Role of Melatonin in Apple Fruit during Growth and Ripening: Possible Interaction with Ethylene. Plants. 2022; 11(5):688. https://doi.org/10.3390/plants11050688
Chicago/Turabian StyleVerde, Antía, Jesús M. Míguez, and Mercedes Gallardo. 2022. "Role of Melatonin in Apple Fruit during Growth and Ripening: Possible Interaction with Ethylene" Plants 11, no. 5: 688. https://doi.org/10.3390/plants11050688