
Mining of Potential Gene Resources for Breeding Nutritionally Improved Maize

 , and
, and
Abstract
:1. Introduction
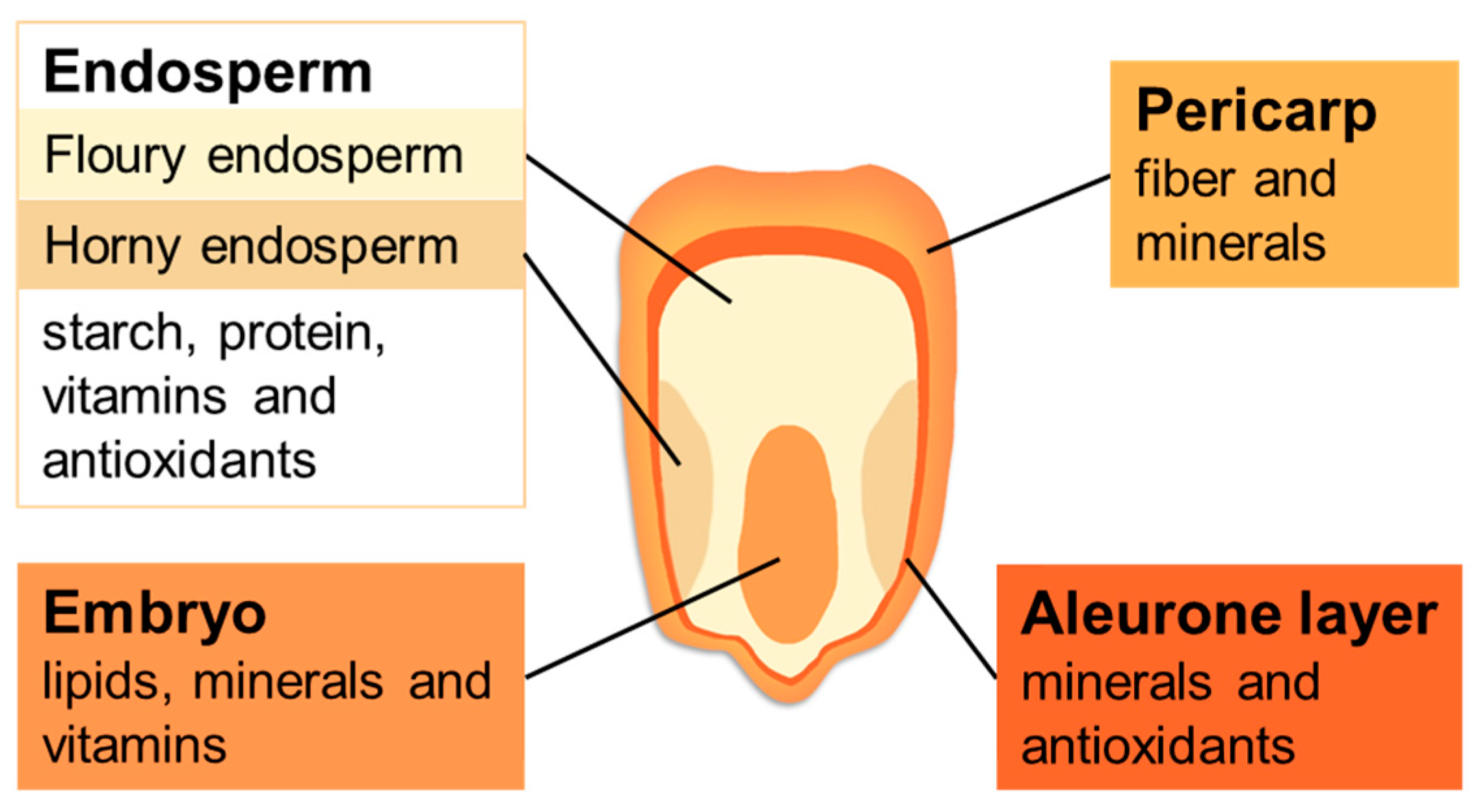
2. Identification of Maize Potential Gene Resources for Starch Content Improvement
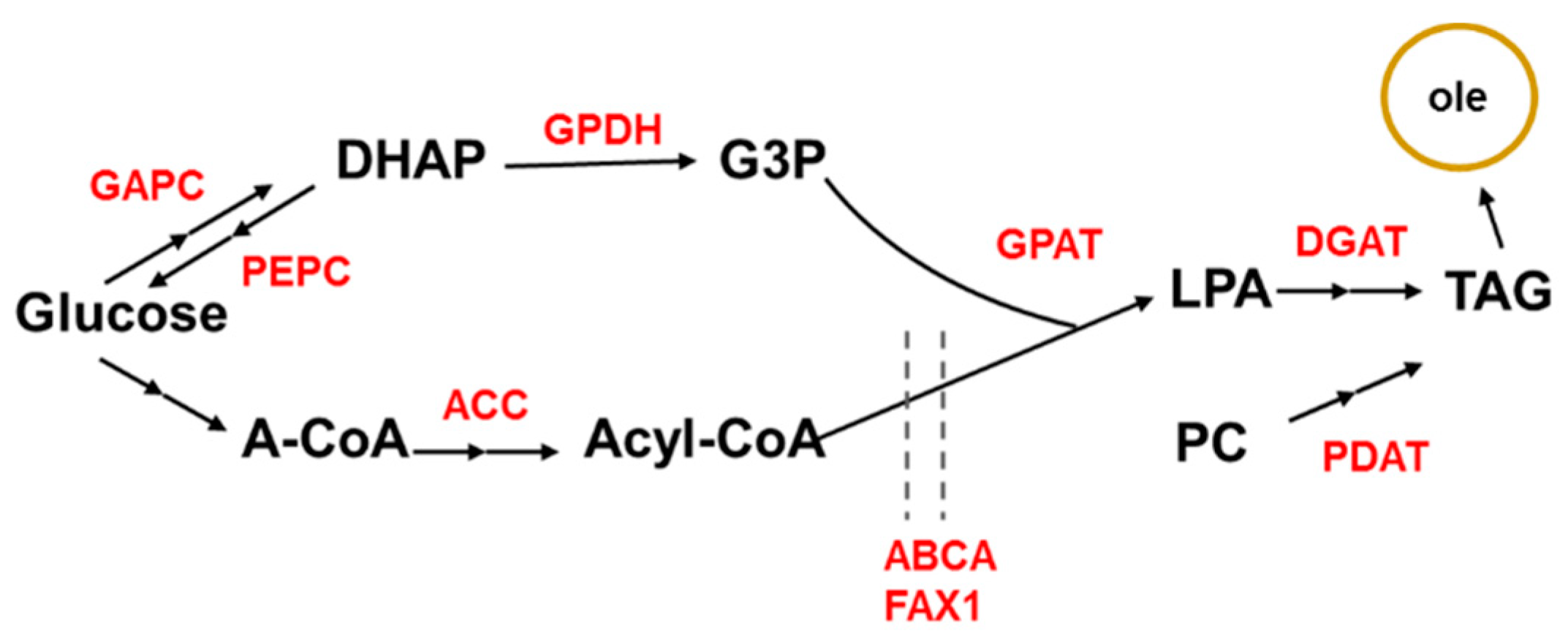
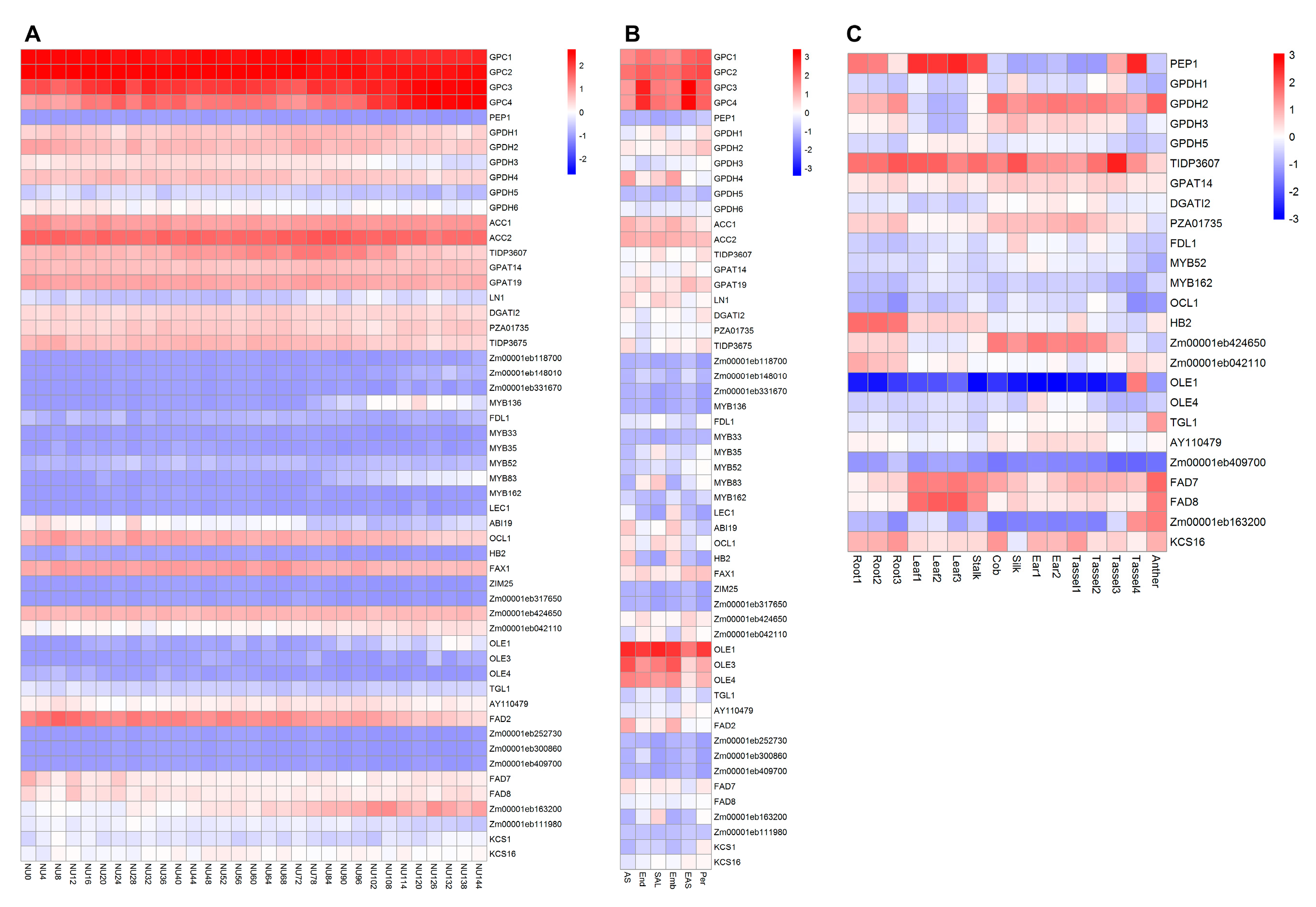
3. Identification of Maize Potential Gene Resources for Oil Content Improvement
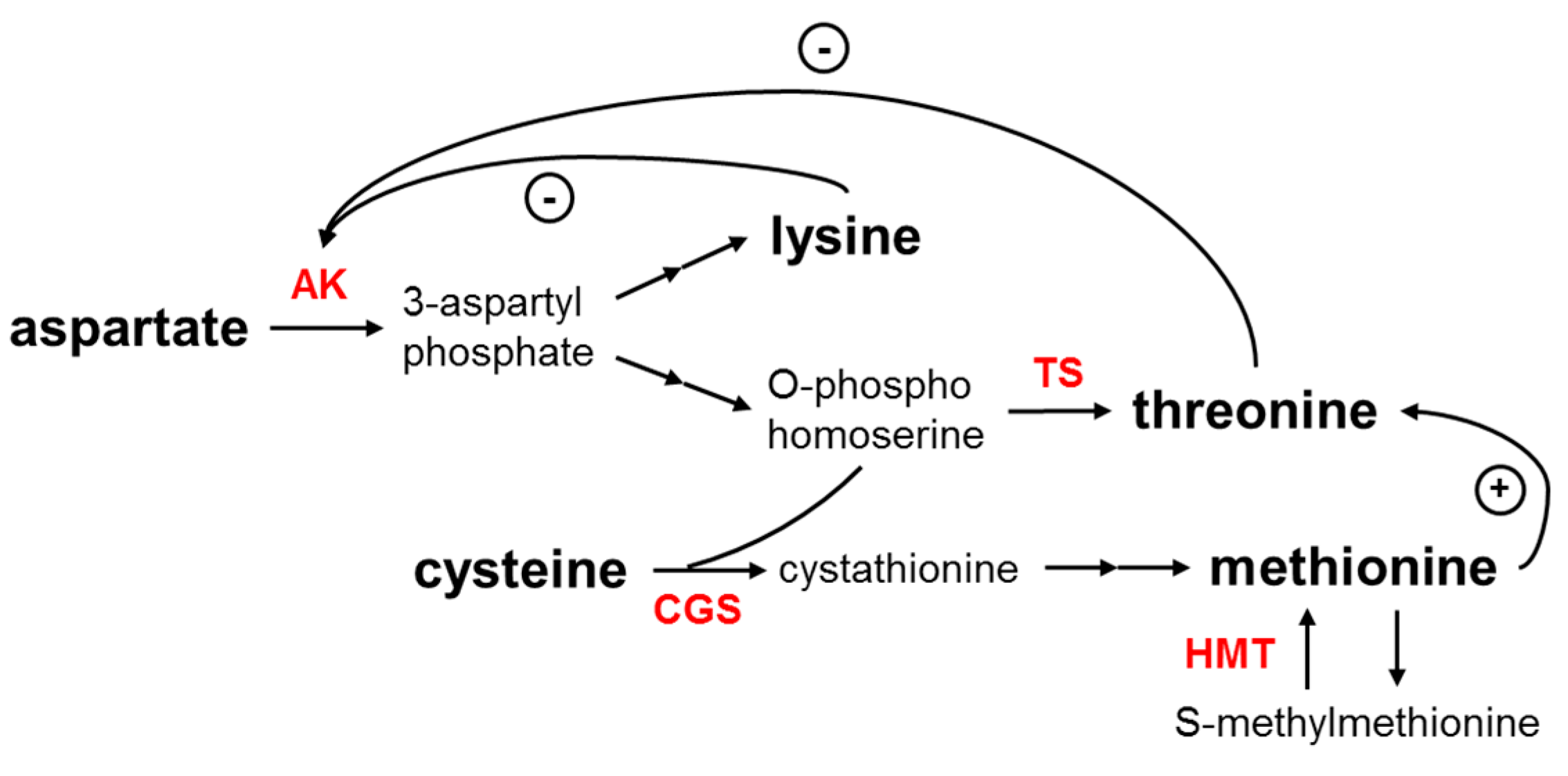
4. Identification of Maize Potential Gene Resources for Essential Amino Acid Content Improvement
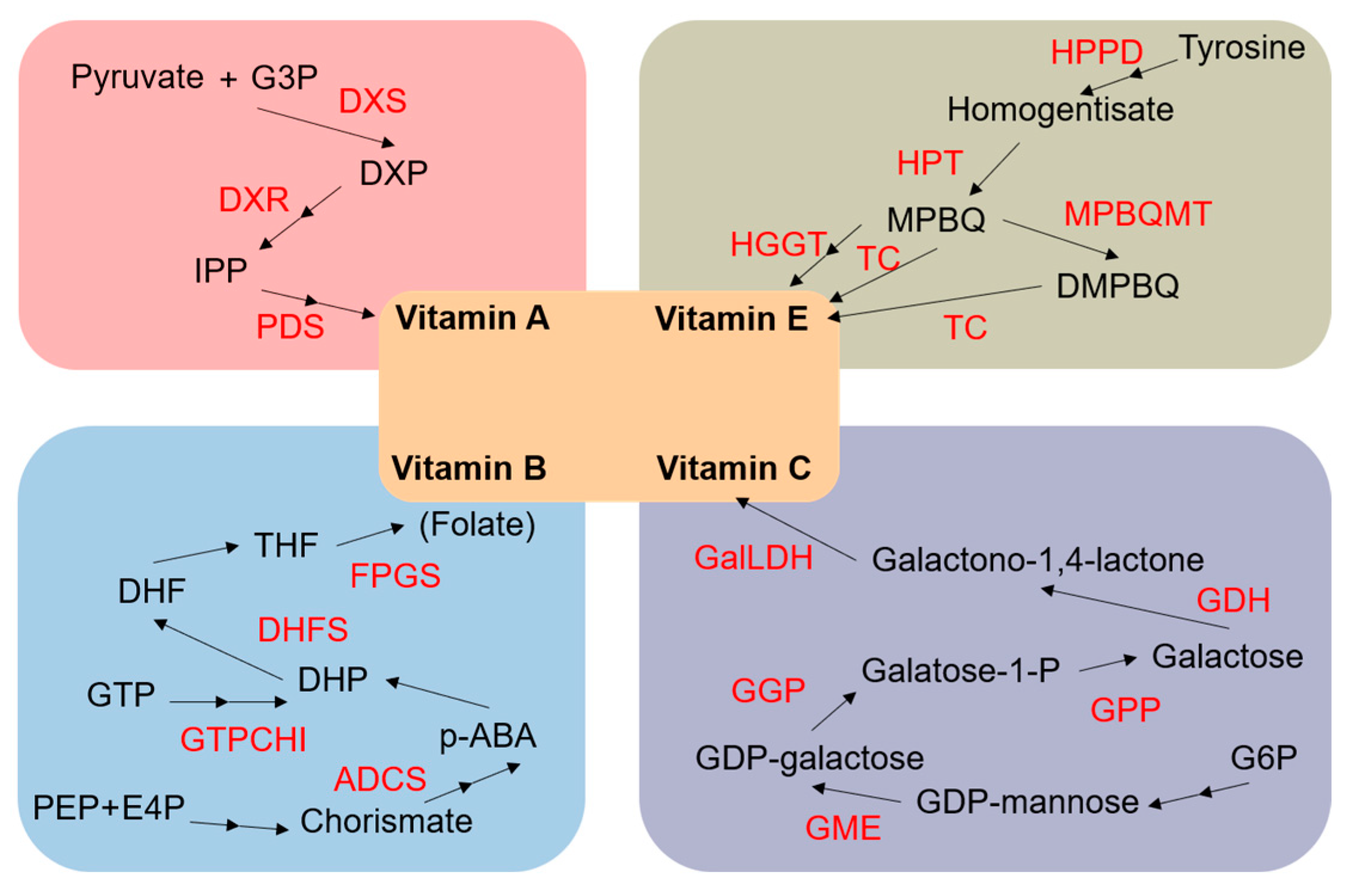
5. Identification of Maize Potential Gene Resources for Vitamin Content Improvement
6. Identification of Maize Potential Gene Resources for Mineral Content Improvement
7. Identification of Maize Potential Gene Resources for Other Secondary Metabolites Content Improvement
8. Expression Patterns of the Putative Nutritional Improvement-Related Maize Genes
9. Discussion
10. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Palacios-Rojas, N.; McCulley, L.; Kaeppler, M.; Titcomb, T.J.; Gunaratna, N.S.; Lopez-Ridaura, S.; Tanumihardjo, S.A. Mining maize diversity and improving its nutritional aspects within agro-food systems. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1809–1834. [Google Scholar] [CrossRef]
- Hou, Q.; Wan, X. Epigenome and Epitranscriptome: Potential Resources for Crop Improvement. Int. J. Mol. Sci. 2021, 22, 12912. [Google Scholar] [CrossRef] [PubMed]
- Chilcoat, D.; Liu, Z.-B.; Sander, J. Use of CRISPR/Cas9 for Crop Improvement in Maize and Soybean. Prog. Mol. Biol. Transl. Sci. 2017, 149, 27–46. [Google Scholar] [CrossRef]
- Yadava, P.; Abhishek, A.; Singh, R.; Singh, I.; Kaul, T.; Pattanayak, A.; Agrawal, P.K. Advances in Maize Transformation Technologies and Development of Transgenic Maize. Front. Plant Sci. 2017, 7, 1949. [Google Scholar] [CrossRef] [Green Version]
- Hannah, L.C.; James, M. The complexities of starch biosynthesis in cereal endosperms. Curr. Opin. Biotechnol. 2008, 19, 160–165. [Google Scholar] [CrossRef]
- Goren, A.; Ashlock, D.; Tetlow, I.J. Starch formation inside plastids of higher plants. Protoplasma 2018, 255, 1855–1876. [Google Scholar] [CrossRef]
- Bello-Perez, L.A.; Flores-Silva, P.C.; Agama-Acevedo, E.; Tovar, J. Starch digestibility: Past, present, and future. J. Sci. Food Agric. 2018, 100, 5009–5016. [Google Scholar] [CrossRef]
- DeMartino, P.; Cockburn, D.W. Resistant starch: Impact on the gut microbiome and health. Curr. Opin. Biotechnol. 2020, 61, 66–71. [Google Scholar] [CrossRef]
- Sang, Y.; Bean, S.; Seib, P.A.; Pedersen, J.; Shi, Y.-C. Structure and Functional Properties of Sorghum Starches Differing in Amylose Content. J. Agric. Food Chem. 2008, 56, 6680–6685. [Google Scholar] [CrossRef]
- Zhu, L.; Gu, M.; Meng, X.; Cheung, S.C.; Yu, H.; Huang, J.; Sun, Y.; Shi, Y.; Liu, Q. High-amylose rice improves indices of animal health in normal and diabetic rats. Plant Biotechnol. J. 2011, 10, 353–362. [Google Scholar] [CrossRef]
- Jiang, H.; Zhang, J.; Wang, J.; Xia, M.; Zhu, S.; Cheng, B. RNA interference-mediated silencing of the starch branching enzyme gene improves amylose content in rice. Genet. Mol. Res. 2013, 12, 2800–2808. [Google Scholar] [CrossRef] [PubMed]
- Regina, A.; Berbezy, P.; Kosar-Hashemi, B.; Li, S.; Cmiel, M.; Larroque, O.; Bird, A.R.; Swain, S.; Cavanagh, C.; Jobling, S.A.; et al. A genetic strategy generating wheat with very high amylose content. Plant Biotechnol. J. 2015, 13, 1276–1286. [Google Scholar] [CrossRef] [PubMed]
- Carciofi, M.; Blennow, A.; Jensen, S.L.; Shaik, S.S.; Henriksen, A.; Buléon, A.; Holm, P.B.; Hebelstrup, K.H. Concerted suppression of all starch branching enzyme genes in barley produces amylose-only starch granules. BMC Plant Biol. 2012, 12, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brummell, A.D.; Watson, L.M.; Zhou, J.; McKenzie, M.J.; Hallett, I.C.; Simmons, L.; Carpenter, M.; Timmerman-Vaughan, G.M. Overexpression of starch branching enzyme II increases short-chain branching of amylopectin and alters the physicochemical properties of starch from potato tuber. BMC Biotechnol. 2015, 15, 28. [Google Scholar] [CrossRef] [Green Version]
- Schwall, G.; Safford, R.; Westcott, R.J.; Jeffcoat, R.; Tayal, A.; Shi, Y.-C.; Gidley, M.J.; Jobling, S.A. Production of very-high-amylose potato starch by inhibition of SBE A and B. Nat. Biotechnol. 2000, 18, 551–554. [Google Scholar] [CrossRef]
- Zhou, W.; Zhao, S.; He, S.; Ma, Q.; Lu, X.; Hao, X.; Wang, H.; Yang, J.; Zhang, P. Production of very-high-amylose cassava by post-transcriptional silencing of branching enzyme genes. J. Integr. Plant Biol. 2020, 62, 832–846. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Cheng, Z.; Zhang, X.; Guo, X.; Su, N.; Jiang, L.; Mao, L.; Wan, J. Double repression of soluble starch synthase genes SSIIa and SSIIIa in rice (Oryza sativa L.) uncovers interactive effects on the physicochemical properties of starch. Genome 2011, 54, 448–459. [Google Scholar] [CrossRef]
- Huang, L.; Gu, Z.; Chen, Z.; Yu, J.; Chu, R.; Tan, H.; Zhao, D.; Fan, X.; Zhang, C.; Li, Q.; et al. Improving rice eating and cooking quality by coordinated expression of the major starch synthesis-related genes, SSII and Wx, in endosperm. Plant Mol. Biol. 2021, 106, 419–432. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Zhang, H.; Zhai, H.; Liu, Q.; He, S. A soluble starch synthase I gene, IbSSI, alters the content, composition, granule size and structure of starch in transgenic sweet potato. Sci. Rep. 2017, 7, 2315. [Google Scholar] [CrossRef] [Green Version]
- Bull, S.E.; Seung, D.; Chanez, C.; Mehta, D.; Kuon, J.-E.; Truernit, E.; Hochmuth, A.; Zurkirchen, I.; Zeeman, S.C.; Gruissem, W.; et al. Accelerated ex situ breeding of GBSS- and PTST1 -edited cassava for modified starch. Sci. Adv. 2018, 4, eaat6086. [Google Scholar] [CrossRef] [Green Version]
- Seung, D.; Soyk, S.; Coiro, M.; Maier, B.A.; Eicke, S.; Zeeman, S.C. Protein targeting to starch is Required for Localising granule-bound starch synthase to Starch Granules and for Normal Amylose Synthesis in Arabidopsis. PLOS Biol. 2015, 13, e1002080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, Y.; Crofts, N.; Abe, M.; Hosaka, Y.; Fujita, N. Characterization of the endosperm starch and the pleiotropic effects of biosynthetic enzymes on their properties in novel mutant rice lines with high resistant starch and amylose content. Plant Sci. 2017, 258, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Dupont, J.; White, P.J.; Carpenter, M.P.; Schaefer, E.J.; Meydani, S.N.; Elson, C.E.; Woods, M.; Gorbach, S.L. Food uses and health effects of corn oil. J. Am. Coll. Nutr. 1990, 9, 438–470. [Google Scholar] [CrossRef]
- Broun, P.; Gettner, S.; Somerville, C. Genetic engineering of plant lipids. Annu. Rev. Nutr. 1999, 19, 197–216. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Ma, F.; Wei, F.; Fanella, B.; Allen, D.; Wang, X. Cytosolic Phosphorylating Glyceraldehyde-3-Phosphate Dehydrogenases Affect Arabidopsis Cellular Metabolism and Promote Seed Oil Accumulation. Plant Cell 2014, 26, 3023–3035. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Huang, Y.; Wang, Y.; Cui, Y.; Liu, Z.; Hua, J. RNA interference of GhPEPC2 enhanced seed oil accumulation and salt tolerance in Upland cotton. Plant Sci. 2018, 271, 52–61. [Google Scholar] [CrossRef]
- Lee, J.; Ridgway, N.D. Substrate channeling in the glycerol-3-phosphate pathway regulates the synthesis, storage and secretion of glycerolipids. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2020, 1865, 158438. [Google Scholar] [CrossRef]
- Yang, S.U.; Kim, J.; Kim, H.; Suh, M.C. Functional Characterization of Physcomitrella patens Glycerol-3-Phosphate Acyltransferase 9 and an Increase in Seed Oil Content in Arabidopsis by Its Ectopic Expression. Plants 2019, 8, 284. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Bi, R.; Li, S.; Zhou, D.; Bai, Y.; Jing, G.; Zhang, K.; Zhang, W. Genome-wide analysis and functional characterization of Acyl-CoA:diacylglycerol acyltransferase from soybean identify GmDGAT1A and 1B roles in oil synthesis in Arabidopsis seeds. J. Plant Physiol. 2019, 242, 153019. [Google Scholar] [CrossRef]
- Izadi-Darbandi, A.; Younessi-Hamzekhanlu, M.; Sticklen, M. Metabolically engineered rice biomass and grain using genes associated with lipid pathway show high level of oil content. Mol. Biol. Rep. 2020, 47, 7917–7927. [Google Scholar] [CrossRef]
- Van Erp, H.; Kelly, A.A.; Menard, G.; Eastmond, P.J. Multigene Engineering of Triacylglycerol Metabolism Boosts Seed Oil Content in Arabidopsis. Plant Physiol. 2014, 165, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Jin, C.; Duan, S.; Zhu, Y.; Qi, S.; Liu, K.; Gao, C.; Ma, H.; Zhang, M.; Liao, Y.; et al. MYB89 Transcription Factor Represses Seed Oil Accumulation. Plant Physiol. 2017, 173, 1211–1225. [Google Scholar] [CrossRef] [Green Version]
- Gil Lee, H.; Kim, H.; Suh, M.C.; Kim, H.U.; Seo, P.J. The MYB96 Transcription Factor Regulates Triacylglycerol Accumulation by Activating DGAT1 and PDAT1 Expression in Arabidopsis Seeds. Plant Cell Physiol. 2018, 59, 1432–1442. [Google Scholar] [CrossRef]
- Tang, G.; Xu, P.; Ma, W.; Wang, F.; Liu, Z.; Wan, S.; Shan, L. Seed-Specific Expression of AtLEC1 Increased Oil Content and Altered Fatty Acid Composition in Seeds of Peanut (Arachis hypogaea L.). Front. Plant Sci. 2018, 9, 260. [Google Scholar] [CrossRef] [Green Version]
- Mu, J.; Tan, H.; Zheng, Q.; Fu, F.; Liang, Y.; Zhang, J.; Yang, X.; Wang, T.; Chong, K.; Wang, X.-J.; et al. Leafy cotyledon1 is a Key Regulator of Fatty Acid Biosynthesis in Arabidopsis. Plant Physiol. 2008, 148, 1042–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angeles-Núñez, J.G.; Tiessen, A. Mutation of the transcription factor leafy cotyledon 2 alters the chemical composition of Arabidopsis seeds, decreasing oil and protein content, while maintaining high levels of starch and sucrose in mature seeds. J. Plant Physiol. 2011, 168, 1891–1900. [Google Scholar] [CrossRef]
- Manan, S.; Ahmad, M.Z.; Zhang, G.; Chen, B.; Haq, B.U.; Gaoyang, Z.; Zhao, J. Soybean LEC2 Regulates Subsets of Genes Involved in Controlling the Biosynthesis and Catabolism of Seed Storage Substances and Seed Development. Front. Plant Sci. 2017, 8, 1604. [Google Scholar] [CrossRef] [Green Version]
- Chai, G.; Bai, Z.; Wei, F.; King, G.J.; Wang, C.; Shi, L.; Dong, C.; Chen, H.; Liu, S. Brassica GLABRA2 genes: Analysis of function related to seed oil content and development of functional markers. Theor. Appl. Genet. 2010, 120, 1597–1610. [Google Scholar] [CrossRef]
- Shen, B.; Sinkevicius, K.W.; Selinger, D.A.; Tarczynski, M.C. The Homeobox Gene GLABRA2 Affects Seed Oil Content in Arabidopsis. Plant Mol. Biol. 2006, 60, 377–387. [Google Scholar] [CrossRef]
- Zhang, M.; Cao, X.; Jia, Q.; Ohlrogge, J. FUSCA3activates triacylglycerol accumulation in Arabidopsis seedlings and tobacco BY2 cells. Plant J. 2016, 88, 95–107. [Google Scholar] [CrossRef]
- Vigeolas, H.; Hühn, D.; Geigenberger, P. Nonsymbiotic Hemoglobin-2 Leads to an Elevated Energy State and to a Combined Increase in Polyunsaturated Fatty Acids and Total Oil Content When Overexpressed in Developing Seeds of Transgenic Arabidopsis Plants. Plant Physiol. 2011, 155, 1435–1444. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhang, H.; Hu, Z.; Chu, S.; Yu, K.; Lv, L.; Yang, Y.; Zhang, X.; Chen, X.; Kan, G.; et al. Artificial selection on GmOLEO1 contributes to the increase in seed oil during soybean domestication. PLoS Genet. 2019, 15, e1008267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanai, M.; Yamada, T.; Hayashi, M.; Mano, S.; Nishimura, M. Soybean (Glycine max L.) triacylglycerol lipase GmSDP1 regulates the quality and quantity of seed oil. Sci. Rep. 2019, 9, 8924. [Google Scholar] [CrossRef] [PubMed]
- Kelly, A.A.; Shaw, E.; Powers, S.J.; Kurup, S.; Eastmond, P.J. Suppression of the sugar-dependent1triacylglycerol lipase family during seed development enhances oil yield in oilseed rape (Brassica napus L.). Plant Biotechnol. J. 2013, 11, 355–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Xia, Y.; Wu, L.; Fu, D.; Hayward, A.; Luo, J.; Yan, X.; Xiong, X.; Fu, P.; Wu, G.; et al. Enhanced seed oil content by overexpressing genes related to triacylglyceride synthesis. Gene 2015, 557, 163–171. [Google Scholar] [CrossRef]
- Roesler, K.; Shintani, D.; Savage, L.; Boddupalli, S.; Ohlrogge, J. Targeting of the Arabidopsis Homomeric Acetyl-Coenzyme A Carboxylase to Plastids of Rapeseeds. Plant Physiol. 1997, 113, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Klaus, D.; Ohlrogge, J.B.; Neuhaus, H.E.; Dormann, P. Increased fatty acid production in potato by engineering of acetyl-CoA carboxylase. Planta 2004, 219, 389–396. [Google Scholar] [CrossRef]
- Tian, Y.; Lv, X.; Xie, G.; Zhang, J.; Xu, Y.; Chen, F. Seed-specific overexpression of AtFAX1 increases seed oil content in Arabidopsis. Biochem. Biophys. Res. Commun. 2018, 500, 370–375. [Google Scholar] [CrossRef]
- Kim, S.; Yamaoka, Y.; Ono, H.; Kim, H.; Shim, D.; Maeshima, M.; Martinoia, E.; Cahoon, E.B.; Nishida, I.; Lee, Y. AtABCA9 transporter supplies fatty acids for lipid synthesis to the endoplasmic reticulum. Proc. Natl. Acad. Sci. USA 2013, 110, 773–778. [Google Scholar] [CrossRef] [Green Version]
- Demorest, Z.L.; Coffman, A.; Baltes, N.J.; Stoddard, T.J.; Clasen, B.M.; Luo, S.; Retterath, A.; Yabandith, A.; Gamo, M.E.; Bissen, J.; et al. Direct stacking of sequence-specific nuclease-induced mutations to produce high oleic and low linolenic soybean oil. BMC Plant Biol. 2016, 16, 225. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Lang, C.; Wang, F.; Wu, X.; Liu, R.; Zheng, T.; Zhang, D.; Chen, J.; Wu, G. Depressed expression of FAE1 and FAD2 genes modifies fatty acid profiles and storage compounds accumulation in Brassica napus seeds. Plant Sci. 2017, 263, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Yu, W.; Wu, H.; Zhang, C.; Sun, S.S.; Liu, Q. Lysine biofortification in rice by modulating feedback inhibition of aspartate kinase and dihydrodipicolinate synthase. Plant Biotechnol. J. 2021, 19, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Arifin, A.G.; Gondo, T.; Akashi, R. Asparaginyl-tRNA synthetase gene (SYNC1) characterized by Lotus corniculatus FOX-superroot lines has effects on plant morphology and amino acid contents of seed in soybean. Plant Biotechnol. 2019, 36, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Guenoune, D.; Amir, R.; Badani, H.; Wolf, S.; Galili, S. Coexpression of the Soybean Vegetative Storage Protein β Subunit (S-VSPβ) Either with the Bacterial Feedback-Insensitive Dihydrodipicolinate Synthase or with S-VSPα Stabilizes the S-VSPβ Transgene Protein and Enhances Lysine Production in Transgenic Tobacco Plants. Transgenic Res. 2003, 12, 123–126. [Google Scholar] [CrossRef]
- Guenoune, D.; Amir, R.; Badani, H.; Wolf, S.; Galili, S. Combined expression of S-VSPalpha in two different organelles enhances its accumulation and total lysine production in leaves of transgenic tobacco plants. J. Exp. Bot. 2002, 53, 1867–1870. [Google Scholar] [CrossRef] [Green Version]
- Kawakatsu, T.; Wang, S.; Wakasa, Y.; Takaiwa, F. Increased Lysine Content in Rice Grains by Over-Accumulation of BiP in the Endosperm. Biosci. Biotechnol. Biochem. 2010, 74, 2529–2531. [Google Scholar] [CrossRef] [Green Version]
- Avraham, T.; Badani, H.; Galili, S.; Amir, R. Enhanced levels of methionine and cysteine in transgenic alfalfa (Medicago sativa L.) plants over-expressing the Arabidopsis cystathionine γ-synthase gene. Plant Biotechnol. J. 2004, 3, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Jander, G. Concurrent Overexpression of Arabidopsis thaliana Cystathionine γ-Synthase and Silencing of Endogenous Methionine γ-Lyase Enhance Tuber Methionine Content in Solanum tuberosum. J. Agric. Food Chem. 2017, 65, 2737–2742. [Google Scholar] [CrossRef]
- Galili, G.; Galili, S.; Lewinsohn, E.; Tadmor, Y. Genetic, Molecular, and Genomic Approaches to Improve the Value of Plant Foods and Feeds. Crit. Rev. Plant Sci. 2002, 21, 167–204. [Google Scholar] [CrossRef]
- Bartlem, D.; Lambein, I.; Okamoto, T.; Itaya, A.; Uda, Y.; Kijima, F.; Tamaki, Y.; Nambara, E.; Naito, S. Mutation in the Threonine Synthase Gene Results in an Over-Accumulation of Soluble Methionine in Arabidopsis. Plant Physiol. 2000, 123, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Zeh, M.; Casazza, A.P.; Kreft, O.; Roessner, U.; Bieberich, K.; Willmitzer, L.; Hoefgen, R.; Hesse, H. Antisense Inhibition of Threonine Synthase Leads to High Methionine Content in Transgenic Potato Plants. Plant Physiol. 2001, 127, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Radwanski, E.R.; Last, R.L. Tryptophan biosynthesis and metabolism: Biochemical and molecular genetics. Plant Cell 1995, 7, 921–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Cai, H.; Xiao, J.; Li, X.; Zhang, Q.; Lian, X. Over-expression of aspartate aminotransferase genes in rice resulted in altered nitrogen metabolism and increased amino acid content in seeds. Theor. Appl. Genet. 2009, 118, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Weichert, N.; Saalbach, I.; Weichert, H.; Kohl, S.; Erban, A.; Kopka, J.; Hause, B.; Varshney, A.; Sreenivasulu, N.; Strickert, M.; et al. Increasing Sucrose Uptake Capacity of Wheat Grains Stimulates Storage Protein Synthesis. Plant Physiol. 2010, 152, 698–710. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Feng, B.; Xu, Z.; Fan, X.; Liu, J.; Liu, Q.; Zhu, P.; Wang, T. TaAAP6-3B, a regulator of grain protein content selected during wheat improvement. BMC Plant Biol. 2018, 18, 71. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Xie, L.; Tian, X.; Liu, S.; Xu, D.; Jin, H.; Song, J.; Dong, Y.; Zhao, D.; Li, G.; et al. TaNAC100 acts as an integrator of seed protein and starch synthesis conferring pleiotropic effects on agronomic traits in wheat. Plant J. 2021, 108, 829–840. [Google Scholar] [CrossRef]
- Cohen, H.; Salmon, A.; Tietel, Z.; Hacham, Y.; Amir, R. The relative contribution of genes operating in the S-methylmethionine cycle to methionine metabolism in Arabidopsis seeds. Plant Cell Rep. 2017, 36, 731–743. [Google Scholar] [CrossRef]
- Tozawa, Y.; Hasegawa, H.; Terakawa, T.; Wakasa, K. Characterization of Rice Anthranilate Synthase α-Subunit GenesOASA1 and OASA2. Tryptophan Accumulation in Transgenic Rice Expressing a Feedback-Insensitive Mutant of OASA1. Plant Physiol. 2001, 126, 1493–1506. [Google Scholar] [CrossRef] [Green Version]
- Tsai, F.-Y.; Brotherton, J.E.; Widholm, J.M. Overexpression of the feedback-insensitive anthranilate synthase gene in tobacco causes tryptophan accumulation. Plant Cell Rep. 2005, 23, 548–556. [Google Scholar] [CrossRef]
- Lee, S.; Park, J.; Lee, J.; Shin, D.; Marmagne, A.; Lim, P.O.; Masclaux-Daubresse, C.; An, G.; Gil Nam, H. OsASN1 Overexpression in Rice Increases Grain Protein Content and Yield under Nitrogen-Limiting Conditions. Plant Cell Physiol. 2020, 61, 1309–1320. [Google Scholar] [CrossRef]
- Strobbe, S.; De Lepeleire, J.; Van Der Straeten, D. From in planta Function to Vitamin-Rich Food Crops: The ACE of Biofortification. Front. Plant Sci. 2018, 9, 1862. [Google Scholar] [CrossRef]
- Arcos, Y.; Godoy, F.; Flores-Ortiz, C.; Arenas-M, A.; Stange, C. Boosting carotenoid content in Malus domestica var. Fuji by expressing AtDXR through an Agrobacterium -mediated transformation method. Biotechnol. Bioeng. 2020, 117, 2209–2222. [Google Scholar] [CrossRef]
- Endo, A.; Saika, H.; Takemura, M.; Misawa, N.; Toki, S. A novel approach to carotenoid accumulation in rice callus by mimicking the cauliflower Orange mutation via genome editing. Rice 2019, 12, 81–85. [Google Scholar] [CrossRef]
- Yazdani, M.; Sun, Z.; Yuan, H.; Zeng, S.; Thannhauser, T.W.; Vrebalov, J.; Ma, Q.; Xu, Y.; Fei, Z.; Van Eck, J.; et al. Ectopic expression of ORANGE promotes carotenoid accumulation and fruit development in tomato. Plant Biotechnol. J. 2019, 17, 33–49. [Google Scholar] [CrossRef] [Green Version]
- Davuluri, G.R.; Van Tuinen, A.; Fraser, P.D.; Manfredonia, A.; Newman, R.; Burgess, D.; Brummell, D.; King, S.R.; Palys, J.; Uhlig, J.; et al. Fruit-specific RNAi-mediated suppression of DET1 enhances carotenoid and flavonoid content in tomatoes. Nat. Biotechnol. 2005, 23, 890–895. [Google Scholar] [CrossRef]
- Wei, S.; Li, X.; Gruber, M.Y.; Li, R.; Zhou, R.; Zebarjadi, A.; Hannoufa, A. RNAi-Mediated Suppression of DET1 Alters the Levels of Carotenoids and Sinapate Esters in Seeds of Brassica napus. J. Agric. Food Chem. 2009, 57, 5326–5333. [Google Scholar] [CrossRef]
- Liu, L.; Jia, C.; Zhang, M.; Chen, D.; Chen, S.; Guo, R.; Guo, D.; Wang, Q. Ectopic expression of aBZR1-1Dtranscription factor in brassinosteroid signalling enhances carotenoid accumulation and fruit quality attributes in tomato. Plant Biotechnol. J. 2013, 12, 105–115. [Google Scholar] [CrossRef]
- Zhang, J.; Hu, Z.; Yao, Q.; Guo, X.; Nguyen, V.; Li, F.; Chen, G. A tomato MADS-box protein, SlCMB1, regulates ethylene biosynthesis and carotenoid accumulation during fruit ripening. Sci. Rep. 2018, 8, 3413. [Google Scholar] [CrossRef]
- Luo, Z.; Zhang, J.; Li, J.; Yang, C.; Wang, T.; Ouyang, B.; Li, H.; Giovannoni, J.; Ye, Z. A stay-green protein S l SGR 1 regulates lycopene and β-carotene accumulation by interacting directly with S l PSY 1 during ripening processes in tomato. New Phytol. 2013, 198, 442–452. [Google Scholar] [CrossRef]
- Ko, M.R.; Song, M.-H.; Kim, J.K.; Baek, S.-A.; You, M.K.; Lim, S.-H.; Ha, S.-H. RNAi-mediated suppression of three carotenoid-cleavage dioxygenase genes, OsCCD1, 4a, and 4b, increases carotenoid content in rice. J. Exp. Bot. 2018, 69, 5105–5116. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Jorge, S.; Ha, S.-H.; Magallanes-Lundback, M.; Gilliland, L.U.; Zhou, A.; Lipka, A.E.; Nguyen, Y.-N.; Angelovici, R.; Lin, H.; Cepela, J.; et al. Carotenoid cleavage dioxygenase4 is a Negative Regulator of β-Carotene Content in Arabidopsis Seeds. Plant Cell 2014, 25, 4812–4826. [Google Scholar] [CrossRef] [Green Version]
- Gayen, D.; Ali, N.; Sarkar, S.N.; Datta, S.K.; Datta, K. Down-regulation of lipoxygenase gene reduces degradation of carotenoids of golden rice during storage. Planta 2015, 242, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Bermudez, L.F.; Del Pozo, T.; Lira, B.S.; De Godoy, F.; Boos, I.; Romanó, C.; Previtali, V.; Almeida, J.; Bréhélin, C.; Asis, R.; et al. A Tomato Tocopherol Binding Protein Sheds Light on Intracellular α-tocopherol Metabolism in Plants. Plant Cell Physiol. 2018, 59, 2188–2203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallie, D.R. Increasing Vitamin C Content in Plant Foods to Improve Their Nutritional Value—Successes and Challenges. Nutrients 2013, 5, 3424–3446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.-S.; Kim, I.-S.; Bae, M.-J.; Choe, Y.-H.; Kim, Y.-H.; Park, H.-M.; Kang, H.-G.; Yoon, H.-S. Homologous expression of cytosolic dehydroascorbate reductase increases grain yield and biomass under paddy field conditions in transgenic rice (Oryza sativa L. japonica). Planta 2013, 237, 1613–1625. [Google Scholar] [CrossRef]
- Wang, Z.; Xiao, Y.; Chen, W.; Tang, K.; Zhang, L. Increased Vitamin C Content Accompanied by an Enhanced Recycling Pathway Confers Oxidative Stress Tolerance inArabidopsis. J. Integr. Plant Biol. 2010, 52, 400–409. [Google Scholar] [CrossRef]
- Dong, W.; Cheng, Z.-J.; Lei, C.-L.; Wang, X.-L.; Wang, J.-L.; Wang, J.; Wu, F.-Q.; Zhang, X.; Guo, X.-P.; Zhai, H.-Q.; et al. Overexpression of Folate Biosynthesis Genes in Rice (Oryza sativa L.) and Evaluation of Their Impact on Seed Folate Content. Mater. Veg. 2014, 69, 379–385. [Google Scholar] [CrossRef]
- Akhtar, T.A.; Orsomando, G.; Mehrshahi, P.; Lara-Núñez, A.; Bennett, M.J.; Iii, J.F.G.; Hanson, A.D. A central role for gamma-glutamyl hydrolases in plant folate homeostasis. Plant J. 2010, 64, 256–266. [Google Scholar] [CrossRef]
- Che, P.; Zhao, Z.-Y.; Glassman, K.; Dolde, D.; Hu, T.X.; Jones, T.J.; Gruis, D.F.; Obukosia, S.; Wambugu, F.; Albertsen, M.C. Elevated vitamin E content improves all-trans β-carotene accumulation and stability in biofortified sorghum. Proc. Natl. Acad. Sci. USA 2016, 113, 11040–11045. [Google Scholar] [CrossRef] [Green Version]
- McQuinn, R.P.; Wong, B.; Giovannoni, J.J. AtPDS overexpression in tomato: Exposing unique patterns of carotenoid self-regulation and an alternative strategy for the enhancement of fruit carotenoid content. Plant Biotechnol. J. 2018, 16, 482–494. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-P.; Charng, Y.-Y. Supraoptimal activity of chlorophyll dephytylase1 results in an increase in tocopherol level in mature arabidopsis seeds. Plant Signal. Behav. 2017, 12, e1382797. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, W.; Ren, G.; Li, D.; Cahoon, R.E.; Chen, M.; Zhou, Y.; Yu, B.; Cahoon, E.B. Chlorophyll Synthase under Epigenetic Surveillance Is Critical for Vitamin E Synthesis, and Altered Expression Affects Tocopherol Levels in Arabidopsis. Plant Physiol. 2015, 168, 1503–1511. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, S.; Farré, G.; Zhu, C.; Sandmann, G.; Capell, T.; Christou, P. Simultaneous expression of Arabidopsis ρ-hydroxyphenylpyruvate dioxygenase and MPBQ methyltransferase in transgenic corn kernels triples the tocopherol content. Transgenic Res. 2010, 20, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Sathish, S.; Preethy, K.S.; Venkatesh, R.; Sathishkumar, R. Rapid enhancement of α-tocopherol content in Nicotiana benthamiana by transient expression of Arabidopsis thaliana Tocopherol cyclase and Homogentisate phytyl transferase genes. 3 Biotech 2018, 8, 485. [Google Scholar] [CrossRef] [PubMed]
- Maruta, T.; Ichikawa, Y.; Mieda, T.; Takeda, T.; Tamoi, M.; Yabuta, Y.; Ishikawa, T.; Shigeoka, S. The Contribution ofArabidopsisHomologs ofL-Gulono-1,4-lactone Oxidase to the Biosynthesis of Ascorbic Acid. Biosci. Biotechnol. Biochem. 2010, 74, 1494–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.-Y.; Liu, R.-R.; Zhang, C.-Q.; Tang, K.-X.; Sun, M.-F.; Yan, G.-H.; Liu, Q.-Q. Manipulation of the Rice L-Galactose Pathway: Evaluation of the Effects of Transgene Overexpression on Ascorbate Accumulation and Abiotic Stress Tolerance. PLoS ONE 2015, 10, e0125870. [Google Scholar] [CrossRef]
- Römheld, V.; Marschner, H. Evidence for a Specific Uptake System for Iron Phytosiderophores in Roots of Grasses. Plant Physiol. 1986, 80, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Johnson, A.A.T.; Kyriacou, B.; Callahan, D.L.; Carruthers, L.; Stangoulis, J.; Lombi, E.; Tester, M. Constitutive Overexpression of the OsNAS Gene Family Reveals Single-Gene Strategies for Effective Iron- and Zinc-Biofortification of Rice Endosperm. PLoS ONE 2011, 6, e24476. [Google Scholar] [CrossRef] [Green Version]
- Masuda, H.; Kobayashi, T.; Ishimaru, Y.; Takahashi, M.; Aung, M.S.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Iron-biofortification in rice by the introduction of three barley genes participated in mugineic acid biosynthesis with soybean ferritin gene. Front. Plant Sci. 2013, 4, 132. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, N.; Beyene, G.; Chauhan, R.D.; Gaitán-Solis, E.; Grusak, M.A.; Taylor, N.; Anderson, P. Overexpression of Arabidopsis VIT1 increases accumulation of iron in cassava roots and stems. Plant Sci. 2015, 240, 170–181. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.-Y.; Gruissem, W.; Bhullar, N.K. Targeting intracellular transport combined with efficient uptake and storage significantly increases grain iron and zinc levels in rice. Plant Biotechnol. J. 2019, 17, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, F.; Yoshihara, T.; Shigemoto, N.; Toki, S.; Takaiwa, F. Iron fortification of rice seed by the soybean ferritin gene. Nat. Biotechnol. 1999, 17, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Masuda, H.; Aung, M.S.; Kobayashi, T.; Hamada, T.; Nishizawa, N.K. Enhancement of Iron Acquisition in Rice by the Mugineic Acid Synthase Gene with Ferric Iron Reductase Gene and osiro2 Confers Tolerance in Submerged and Nonsubmerged Calcareous Soils. Front. Plant Sci. 2019, 10, 1179. [Google Scholar] [CrossRef]
- Kobayashi, T.; Ozu, A.; Kobayashi, S.; An, G.; Jeon, J.-S.; Nishizawa, N.K. OsbHLH058 and OsbHLH059 transcription factors positively regulate iron deficiency responses in rice. Plant Mol. Biol. 2019, 101, 471–486. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.X.; Zhang, J.; Wang, D.W.; Ling, H.Q. AtbHLH29 of Arabidopsis thaliana is a functional ortholog of tomato FER involved in controlling iron acquisition in strategy I plants. Cell Res. 2005, 15, 613–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Gao, W.; Peng, Q.; Zhou, B.; Kong, Q.; Ying, Y.; Shou, H. Two soybean bHLH factors regulate response to iron deficiency. J. Integr. Plant Biol. 2018, 60, 608–622. [Google Scholar] [CrossRef]
- Kobayashi, T.; Ogo, Y.; Itai, R.N.; Nakanishi, H.; Takahashi, M.; Mori, S.; Nishizawa, N.K. The transcription factor IDEF1 regulates the response to and tolerance of iron deficiency in plants. Proc. Natl. Acad. Sci. USA 2007, 104, 19150–19155. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.; Liu, F.; Pan, X.-X.; Zang, Y.-P.; Jin, F.; Zu, W.-X.; Qi, X.-T.; Xiao, W.; Yin, L.-P. CSN6, a subunit of the COP9 signalosome, is involved in early response to iron deficiency in Oryza sativa. Sci. Rep. 2016, 6, 25485. [Google Scholar] [CrossRef]
- Yang, A.; Li, Q.; Chen, L.; Zhang, W.-H. A rice small GTPase, Rab6a, is involved in the regulation of grain yield and iron nutrition in response to CO2 enrichment. J. Exp. Bot. 2020, 71, 5680–5688. [Google Scholar] [CrossRef]
- Aung, M.S.; Kobayashi, T.; Masuda, H.; Nishizawa, N.K. Rice HRZ ubiquitin ligases are crucial for the response to excess iron. Physiol. Plant. 2018, 163, 282–296. [Google Scholar] [CrossRef] [Green Version]
- Menguer, P.K.; Farthing, E.; Peaston, K.A.; Ricachenevsky, F.K.; Fett, J.P.; Williams, L.E. Functional analysis of the rice vacuolar zinc transporter OsMTP1. J. Exp. Bot. 2013, 64, 2871–2883. [Google Scholar] [CrossRef] [Green Version]
- Haydon, M.; Cobbett, C.S. A Novel Major Facilitator Superfamily Protein at the Tonoplast Influences Zinc Tolerance and Accumulation in Arabidopsis. Plant Physiol. 2007, 143, 1705–1719. [Google Scholar] [CrossRef] [Green Version]
- Remy, E.; Cabrito, T.R.; Batista, R.A.; Hussein, M.; Teixeira, M.C.; Athanasiadis, A.; Sa-Correia, I.; Duque, P. Intron Retention in the 5′UTR of the Novel ZIF2 Transporter Enhances Translation to Promote Zinc Tolerance in Arabidopsis. PLoS Genet. 2014, 10, e1004375. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Ogo, Y.; Senoura, T.; Nishizawa, N.K.; Nakanishi, H. The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice. Plant Cell Environ. 2012, 35, 1948–1957. [Google Scholar] [CrossRef]
- Ibuot, A.; Webster, R.E.; Williams, L.E.; Pittman, J.K. Increased metal tolerance and bioaccumulation of zinc and cadmium inChlamydomonas reinhardtiiexpressing a AtHMA4 C-terminal domain protein. Biotechnol. Bioeng. 2020, 117, 2996–3005. [Google Scholar] [CrossRef]
- Kappara, S.; Neelamraju, S.; Ramanan, R. Down regulation of a heavy metal transporter gene influences several domestication traits and grain Fe-Zn content in rice. Plant Sci. 2018, 276, 208–219. [Google Scholar] [CrossRef]
- Ramesh, S.; Choimes, S.; Schachtman, D.P. Over-Expression of an Arabidopsis Zinc Transporter in Hordeum Vulgare Increases Short-Term Zinc Uptake after Zinc Deprivation and Seed Zinc Content. Plant Mol. Biol. 2004, 54, 373–385. [Google Scholar] [CrossRef]
- Lee, S.; Kim, S.A.; Lee, J.; Guerinot, M.L.; An, G. Zinc deficiency-inducible OsZIP8 encodes a plasma membrane-localized zinc transporter in rice. Mol. Cells 2010, 29, 551–558. [Google Scholar] [CrossRef]
- Perera, I.; Seneweera, S.; Hirotsu, N. Manipulating the Phytic Acid Content of Rice Grain Toward Improving Micronutrient Bioavailability. Rice 2018, 11, 4. [Google Scholar] [CrossRef] [Green Version]
- Curie, C.; Alonso, J.M.; Le Jean, M.; Ecker, J.R.; Briat, J.-F. Involvement of NRAMP1 from Arabidopsis thaliana in iron transport. Biochem. J. 2000, 347, 749–755. [Google Scholar] [CrossRef]
- Yoneyama, T.; Ishikawa, S.; Fujimaki, S. Route and Regulation of Zinc, Cadmium, and Iron Transport in Rice Plants (Oryza sativa L.) during Vegetative Growth and Grain Filling: Metal Transporters, Metal Speciation, Grain Cd Reduction and Zn and Fe Biofortification. Int. J. Mol. Sci. 2015, 16, 19111–19129. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.-Y.; Gruissem, W.; Bhullar, N.K. Facilitated citrate-dependent iron translocation increases rice endosperm iron and zinc concentrations. Plant Sci. 2018, 270, 13–22. [Google Scholar] [CrossRef]
- Yokosho, K.; Yamaji, N.; Ueno, D.; Mitani, N.; Ma, J.F. OsFRDL1 Is a Citrate Transporter Required for Efficient Translocation of Iron in Rice. Plant Physiol. 2009, 149, 297–305. [Google Scholar] [CrossRef] [Green Version]
- Ali, N.; Paul, S.; Gayen, D.; Sarkar, S.N.; Datta, S.K.; Datta, K. RNAi mediated down regulation of myo-inositol-3-phosphate synthase to generate low phytate rice. Rice 2013, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Tagashira, Y.; Shimizu, T.; Miyamoto, M.; Nishida, S.; Yoshida, K.T. Overexpression of a Gene Involved in Phytic Acid Biosynthesis Substantially Increases Phytic Acid and Total Phosphorus in Rice Seeds. Plants 2015, 4, 196–208. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, S.; Bhattacharya, S.; Karmakar, A.; Ghosh, S.; Sarkar, S.N.; Gangopadhyay, G.; Datta, K.; Datta, S.K. RNAi-mediated down-regulation of ITPK-2 enhanced inorganic phosphorus and minerals in the transgenic rice. J. Biosci. 2021, 46, 32. [Google Scholar] [CrossRef]
- Ali, N.; Paul, S.; Gayen, D.; Sarkar, S.N.; Datta, K.; Datta, S.K. Development of Low Phytate Rice by RNAi Mediated Seed-Specific Silencing of Inositol 1,3,4,5,6-Pentakisphosphate 2-Kinase Gene (IPK1). PLoS ONE 2013, 8, e68161. [Google Scholar] [CrossRef] [Green Version]
- Pandey, B.K.; Mehra, P.; Verma, L.; Bhadouria, J.; Giri, J. OsHAD1, a Haloacid Dehalogenase-Like APase, Enhances Phosphate Accumulation. Plant Physiol. 2017, 174, 2316–2332. [Google Scholar] [CrossRef] [Green Version]
- Holme, I.B.; Dionisio, G.; Madsen, C.K.; Brinch-Pedersen, H. BarleyHvPAPhy_aas transgene provides high and stable phytase activities in mature barley straw and in grains. Plant Biotechnol. J. 2016, 15, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Li, X.; Ma, J.; Li, W.; Yan, G.; Zhang, C. GmPAP4, a novel purple acid phosphatase gene isolated from soybean (Glycine max), enhanced extracellular phytate utilization in Arabidopsis thaliana. Plant Cell Rep. 2014, 33, 655–667. [Google Scholar] [CrossRef]
- Sosulski, F.; Krygier, K.; Hogge, L. Free, esterified, and insoluble-bound phenolic acids. 3. Composition of phenolic acids in cereal and potato flours. J. Agric. Food Chem. 1982, 30, 337–340. [Google Scholar] [CrossRef]
- Messias, R.D.S.; Galli, V.; Silva, S.D.D.A.E.; Schirmer, M.A.; Rombaldi, C. Micronutrient and Functional Compounds Biofortification of Maize Grains. Crit. Rev. Food Sci. Nutr. 2014, 55, 123–139. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Zheng, Y.; Yang, L.; Yao, Z.; Cheng, J.; Zhang, F.; Jiang, H.; Liu, D. The transcription factor AtGLK1 acts upstream of MYBL2 to genetically regulate sucrose-induced anthocyanin biosynthesis in Arabidopsis. BMC Plant Biol. 2021, 21, 242. [Google Scholar] [CrossRef]
- Bai, Y.; Pattanaik, S.; Patra, B.; Werkman, J.R.; Xie, C.H.; Yuan, L. Flavonoid-related basic helix-loop-helix regulators, NtAn1a and NtAn1b, of tobacco have originated from two ancestors and are functionally active. Planta 2011, 234, 363–375. [Google Scholar] [CrossRef]
- Meng, L.-S. Transcription coactivatorArabidopsis ANGUSTIFOLIA3 modulates anthocyanin accumulation and light-induced root elongation through transrepression of Constitutive Photomorphogenic1. Plant Cell Environ. 2014, 38, 838–851. [Google Scholar] [CrossRef]
- Zhou, C.; Zeng, Z.; Suo, J.; Li, X.; Bian, H.; Wang, J.; Zhu, M.; Han, N. Manipulating a Single Transcription Factor, Ant1, Promotes Anthocyanin Accumulation in Barley Grains. J. Agric. Food Chem. 2021, 69, 5306–5317. [Google Scholar] [CrossRef]
- Gao, R.; Han, T.; Xun, H.; Zeng, X.; Li, P.; Li, Y.; Wang, Y.; Shao, Y.; Cheng, X.; Feng, X.; et al. MYB transcription factors GmMYBA2 and GmMYBR function in a feedback loop to control pigmentation of seed coat in soybean. J. Exp. Bot. 2021, 72, 4401–4418. [Google Scholar] [CrossRef]
- Sawano, H.; Matsuzaki, T.; Usui, T.; Tabara, M.; Fukudome, A.; Kanaya, A.; Tanoue, D.; Hiraguri, A.; Horiguchi, G.; Ohtani, M.; et al. Double-stranded RNA-binding protein DRB3 negatively regulates anthocyanin biosynthesis by modulating PAP1 expression in Arabidopsis thaliana. J. Plant Res. 2016, 130, 45–55. [Google Scholar] [CrossRef]
- Yi, F.; Gu, W.; Chen, J.; Song, N.; Gao, X.; Zhang, X.; Zhou, Y.; Ma, X.; Song, W.; Zhao, H.; et al. High Temporal-Resolution Transcriptome Landscape of Early Maize Seed Development. Plant Cell 2019, 31, 974–992. [Google Scholar] [CrossRef] [Green Version]
- Doll, N.M.; Just, J.; Brunaud, V.; Caïus, J.; Grimault, A.; Depège-Fargeix, N.; Esteban, E.; Pasha, A.; Provart, N.J.; Ingram, G.C.; et al. Transcriptomics at Maize Embryo/Endosperm Interfaces Identifies a Transcriptionally Distinct Endosperm Subdomain Adjacent to the Embryo Scutellum. Plant Cell 2020, 32, 833–852. [Google Scholar] [CrossRef] [Green Version]
- Downs, G.S.; Bi, Y.-M.; Colasanti, J.; Wu, W.; Chen, X.; Zhu, T.; Rothstein, S.J.; Lukens, L.N. A Developmental Transcriptional Network for Maize Defines Coexpression Modules. Plant Physiol. 2013, 161, 1830–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, J.; Cheng, F.; Hu, C.; Quan, S.; Lin, H.; Wang, J.; Chen, G.; Zhao, X.; Alexander, D.; Guo, L.; et al. Metabolic map of mature maize kernels. Metabolomics 2014, 10, 775–787. [Google Scholar] [CrossRef]
- Lai, J.; Li, R.; Xu, X.; Jin, W.; Xu, M.; Zhao, H.; Xiang, Z.; Song, W.; Ying, K.; Zhang, M.; et al. Genome-wide patterns of genetic variation among elite maize inbred lines. Nat. Genet. 2010, 42, 1027–1030. [Google Scholar] [CrossRef]
- Garg, M.; Sharma, A.; Vats, S.; Tiwari, V.; Kumari, A.; Mishra, V.; Krishania, M. Vitamins in Cereals: A Critical Review of Content, Health Effects, Processing Losses, Bioaccessibility, Fortification, and Biofortification Strategies for Their Improvement. Front. Nutr. 2021, 8, 586815. [Google Scholar] [CrossRef]
- Aluru, M.; Xu, Y.; Guo, R.; Wang, Z.; Li, S.; White, W.; Wang, K.; Rodermel, S. Generation of transgenic maize with enhanced provitamin A content. J. Exp. Bot. 2008, 59, 3551–3562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karchi, H.; Shaul, O.; Galili, G. Seed-specific expression of a bacterial desensitized aspartate kinase increases the production of seed threonine and methionine in transgenic tobacco. Plant J. 1993, 3, 721–727. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Protein Function | Maize Orthologs | Gene ID | Strategy | References |
|---|---|---|---|---|---|
| SBE | starch branching enzyme | SBE1 | Zm00001eb228530 | knockout | [10,11,12,13,14,15,16] |
| SBE3 | Zm00001eb357830 | ||||
| SBE4 | Zm00001eb084160 | ||||
| AE1 | Zm00001eb242610 | ||||
| SS | starch synthase | SS1 | Zm00001eb376100 | overexpression | [17,18,19] |
| SS2 | Zm00001eb070230 | ||||
| SS3 | Zm00001eb431240 | ||||
| SS4 | Zm00001eb353810 | ||||
| SS5 | Zm00001eb191890 | ||||
| SS6 | Zm00001eb222830 | ||||
| SS7 | Zm00001eb194550 | ||||
| DU1 | Zm00001eb413290 | ||||
| SU2 | Zm00001eb279740 | ||||
| GBSS | granule bound starch synthase | WX1 GBSS1 | Zm00001eb378140 Zm00001eb305810 | overexpression | [20,21,22] |
| PTST1 | protein targeting to starch | GPM177 | Zm00001eb231700 | overexpression | [20,21] |
| Genes | Protein Function | Maize Orthologs | Gene ID | Strategy | References |
|---|---|---|---|---|---|
| GAPC | glyceraldehyde-3-phosphate dehydrogenase | GPC1 | Zm00001eb173410 | overexpression | [25] |
| GPC2 | Zm00001eb261430 | ||||
| GPC3 | Zm00001eb184000 | ||||
| GPC4 | Zm00001eb246370 | ||||
| PEPC2 | phosphoenolpyruvate carboxylase | PEP1 | Zm00001eb383680 | knockout | [26] |
| GPDH | glycerol-3-phosphate dehydrogenase | GPDH1 | Zm00001eb141610 | overexpression | [45] |
| GPDH2 | Zm00001eb369390 | ||||
| GPDH3 | Zm00001eb352530 | ||||
| GPDH4 | Zm00001eb139850 | ||||
| GPDH5 | Zm00001eb303710 | ||||
| GPDH6 | Zm00001eb419210 | ||||
| ACC1 | acetyl-CoA carboxylase | ACC1 ACC2 TIDP3607 | Zm00001eb419400 Zm00001eb086560 Zm00001eb223980 Zm00001eb028920 Zm00001eb189990 Zm00001eb367400 | overexpression | [46,47] |
| GPAT9 | glycerol-3-phosphate acyltransferase | GPAT14 | Zm00001eb396350 | overexpression | [28] |
| GPAT19 | Zm00001eb323170 | ||||
| DGAT1 | diacylglycerol acyltransferase | LN1 | Zm00001eb277490 | overexpression | [29,30,31] |
| DGATI2 | Zm00001eb284200 | ||||
| PDAT | phospholipid diacylglycerol acyltransferase | PZA01735 TIDP3675 | Zm00001eb100310 Zm00001eb314300 Zm00001eb118700 Zm00001eb148010 Zm00001eb331670 Zm00001eb342120 | overexpression | [30] |
| MYB89 | transcription factor | MYB136 | Zm00001eb128770 | knockout | [32] |
| MYB96 | transcription factor | FDL1 | Zm00001eb328280 | overexpression | [33] |
| MYB33 | Zm00001eb041330 | ||||
| MYB35 | Zm00001eb099570 | ||||
| MYB52 | Zm00001eb392230 | ||||
| MYB70 | Zm00001eb109860 | ||||
| MYB83 | Zm00001eb041320 | ||||
| MYB162 | Zm00001eb312600 | ||||
| LEC1 | transcription factor | LEC1 | Zm00001eb253260 | overexpression | [34,35] |
| LEC2 | transcription factor | ABI19 | Zm00001eb361390 | overexpression | [36,37] |
| GL2 | transcription factor | OCL1 | Zm00001eb126140 | knockout | [38,39] |
| FUS3 | transcription factor | ABI19 | Zm00001eb361390 | overexpression | [40] |
| HB2 | transcription factor | HB2 | Zm00001eb293010 | overexpression | [41] |
| FAX1 | carrier protein | FAX1 ZIM25 | Zm00001eb301150 Zm00001eb379540 Zm00001eb317650 Zm00001eb424650 | overexpression | [48] |
| ABCA9 | carrier protein | Zm00001eb042110 | overexpression | [49] | |
| OLE | delta-9 desaturase | OLE1 | Zm00001eb074940 | overexpression | [30,42] |
| OLE3 | Zm00001eb216880 | ||||
| OLE4 | Zm00001eb053890 | ||||
| SDP1 | sugar dependent | TGL1 | Zm00001eb370460 | knockout | [31,43,44] |
| AY110479 | Zm00001eb062080 | ||||
| FAD2 | delta-12 fatty acid desaturase | FAD2 | Zm00001eb188990 Zm00001eb252730 Zm00001eb300860 Zm00001eb409700 Zm00001eb442020 | knockout | [50,51] |
| FAD3 | delta-12 fatty acid desaturase | FAD7 FAD8 | Zm00001eb397050 Zm00001eb013340 Zm00001eb163200 Zm00001eb111980 | knockout | [50] |
| FAE1 | fatty acid elongase | KCS1 | Zm00001eb344070 | knockout | [51] |
| KCS16 | Zm00001eb296230 |
| Genes | Protein Function | Maize Orthologs | Gene ID | Strategy | References |
|---|---|---|---|---|---|
| AK | aspartate kinase | ASK1 | Zm00001eb064530 | knockout | [52] |
| ASK2 | Zm00001eb094670 | ||||
| SYNC1 | asparaginyl-tRNA synthetase | Zm00001eb341390 | overexpression | [53] | |
| VSP | storage protein | VSP1 | Zm00001eb283460 | overexpression | [54,55] |
| VSP2 | Zm00001eb283450 | ||||
| BIP | storage protein | BIP1 | Zm00001eb229930 | overexpression | [56] |
| BIP2 | Zm00001eb209550 | ||||
| BIP3 | Zm00001eb214940 | ||||
| CGS | cystathionine γ-synthase | CGS1 | Zm00001eb392050 Zm00001eb018300 | overexpression | [57,58] |
| TS1 | threonine synthase | THR1 THR2 THR3 | Zm00001eb156020 Zm00001eb294790 Zm00001eb284240 Zm00001eb022690 Zm00001eb088230 | knockout | [60,61] |
| HMT | homocysteine S-methyltransferase | HMT1 | Zm00001eb399940 | overexpression | [67] |
| ASA | anthranilate synthase | Zm00001eb063220 Zm00001eb211420 | overexpression | [68,69] | |
| AAT | aspartate aminotransferase | GOT1 | Zm00001eb152450 | overexpression | [63] |
| GOT2 | Zm00001eb257910 | ||||
| GOT3 | Zm00001eb238900 | ||||
| GOT4 | Zm00001eb146400 | ||||
| ASN1 | asparagine synthetase | ASN3 | Zm00001eb013430 | overexpression | [70] |
| ASN4 | Zm00001eb396990 | ||||
| SUT1 | carrier protein | SUT1 | Zm00001eb005460 | overexpression | [64] |
| SUT7 | Zm00001eb402200 | ||||
| AAP6 | carrier protein | AAAP21 | Zm00001eb145670 | overexpression | [65] |
| NAC100 | transcription factor | NACTF32 | Zm00001eb080700 | knockout | [66] |
| Genes | Protein Function | Maize Orthologs | Gene ID | Strategy | References |
|---|---|---|---|---|---|
| DXS | 1-deoxyxylulose 5-phosphate synthase | DXS1 | Zm00001eb287860 | overexpression | [89] |
| DXR | 1-deoxy-D-xylulose 5-phosphate reductoisomerase | DXR1 | Zm00001eb126690 | overexpression | [72] |
| DXR2 | Zm00001eb334370 | ||||
| PDS | phytoene desaturase | VP5 | Zm00001eb006300 | overexpression | [90] |
| OR | coactivator | Zm00001eb249060 | overexpression | [73,74] | |
| DET1 | transcription factor | Zm00001eb317230 Zm00001eb341540 | knockout | [75,76] | |
| BZR1 | transcription factor | BES1 | Zm00001eb325550 | overexpression | [77] |
| CMB1 | transcription factor | ZMM6 | Zm00001eb036590 | overexpression | [78] |
| ZMM7 | Zm00001eb317770 | ||||
| ZMM27 | Zm00001eb102450 | ||||
| SGR1 | magnesium dechelatase | NYE1 | Zm00001eb319560 | knockout | [79] |
| NYE2 | Zm00001eb103480 | ||||
| CCD4 | carotenoid cleavage dioxygenase | NCED6 | Zm00001eb188280 | knockout | [80,81] |
| NCED8 | Zm00001eb251990 | ||||
| LOX1 | lipoxygenase | LOX4 | Zm00001eb054050 | knockout | [82] |
| LOX5 | Zm00001eb216870 | ||||
| CLD1 | chlorophyll dephytylase | UMC2173 | Zm00001eb349130 | overexpression | [91] |
| CHLG | chlorophyll synthase | CHLG1 | Zm00001eb286140 | knockout | [92] |
| GPPD | ρ-hydroxyphenylpyruvate dioxygenase | HPPD1 | Zm00001eb232960 Zm00001eb304950 | overexpression | [93] |
| HPT | homogentisate phytyltransferase | HPT1 | Zm00001eb389370 | overexpression | [94] |
| HGGT | homogentisic acid geranylgeranyl transferase | HGGT1 HGGT2 HGGT3 | Zm00001eb386720 Zm00001eb105110 Zm00001eb121230 Zm00001eb382300 | overexpression | [89] |
| MPBQMT | 2-methyl-6-phytylbenzoquinol methyltransferase | APG1 | Zm00001eb031790 | overexpression | [93] |
| TC | tocopherol cyclase | SXD1 | Zm00001eb237270 | overexpression | [94] |
| TBP | tocopherol-binding protein | Zm00001eb197980 Zm00001eb347610 | overexpression | [83] | |
| GULLO | L-gulono-1,4-lactone oxidase | Zm00001eb059530 Zm00001eb072160 Zm00001eb154880 Zm00001eb236290 Zm00001eb421440 Zm00001eb236880 | overexpression | [95] | |
| GME | GDP-mannose 3,5-epimerase | GME1 | Zm00001eb047980 | overexpression | [96] |
| GME2 | Zm00001eb167750 | ||||
| GGP | GDP-L-galactose phosphorylase | SI946084H12 | Zm00001eb144410 | overexpression | [96] |
| GPP | L-galactose-1-phosphate phosphatase | GPP1 | Zm00001eb049310 | overexpression | [96] |
| GDH | L-galactose dehydrogenase | GALDH1 | Zm00001eb408730 | overexpression | [96] |
| GALLDH | L-galactono-1,4-lactone dehydrogenase | GLDH1 | Zm00001eb093120 | overexpression | [96] |
| DHAR1 | dehydroascorbate reductase | DHAR1 | Zm00001eb355540 | overexpression | [85,86] |
| DHAR2 | Zm00001eb355550 | ||||
| DHAR3 | Zm00001eb266260 | ||||
| GTPCHI | GTP cyclohydrolase | GCH1 | Zm00001eb067370 | overexpression | [87] |
| GCH2 | Zm00001eb432940 | ||||
| ADCS | aminodeoxychorismate synthase | ADCS1 | Zm00001eb272970 | overexpression | [87] |
| DHFS | dihydrofolate synthetase | DHFS1 | Zm00001eb410070 | overexpression | [87] |
| DHFS2 | Zm00001eb137120 | ||||
| FPGS | folylpolyglutamate synthase | FGP2 BM4 | Zm00001eb044170 Zm00001eb404110 Zm00001eb299330 Zm00001eb421680 | overexpression | [87] |
| GGH | γ-glutamyl hydrolase | Zm00001eb199250 Zm00001eb353180 | overexpression | [88] |
| Genes | Protein Function | Maize Orthologs | Gene ID | Strategy | References |
|---|---|---|---|---|---|
| NAS | nicotianamine synthase | NAS1 | Zm00001eb396230 | overexpression | [98] |
| NAS2 | Zm00001eb014700 | ||||
| NAS3 | Zm00001eb052890 | ||||
| NAS4 | Zm00001eb218440 | ||||
| NAS6 | Zm00001eb396110 | ||||
| NAS8 | Zm00001eb396250 | ||||
| NAS9 | Zm00001eb014680 | ||||
| NAS10 | Zm00001eb396280 | ||||
| NAAT | nicotianamine aminotransferase | NAAT1 | Zm00001eb203230 | overexpression | [99] |
| PCO115235C | Zm00001eb240650 | ||||
| NRAMP1 | carrier protein | NRAT1 | Zm00001eb224770 | overexpression | [120] |
| NRAMP5 | carrier protein | NRAT5 | Zm00001eb304610 | overexpression | [121] |
| FRD3 | carrier protein | MATE1 | Zm00001eb261140 Zm00001eb143800 Zm00001eb424530 | overexpression | [122] |
| FRDL1 | carrier protein | MATE3 | Zm00001eb008790 | overexpression | [123] |
| VIT1 | carrier protein | Zm00001eb424350 Zm00001eb099160 Zm00001eb312010 | knockout | [100] | |
| NRAMP3 | carrier protein | NRAT3 NRAT4 | Zm00001eb400560 Zm00001eb030050 Zm00001eb051790 | overexpression | [101] |
| FER | storage protein | FER1 | Zm00001eb195010 | overexpression | [102] |
| FER2 | Zm00001eb404870 | ||||
| IRO2 | transcription factor | BHLH54 | Zm00001eb362800 | overexpression | [103] |
| BHLH126 | Zm00001eb140680 | ||||
| BHLH58 | transcription factor | BHLH118 | Zm00001eb289490 | overexpression | [104] |
| BHLH59 | transcription factor | BHLH128 | Zm00001eb209480 | overexpression | [104] |
| BHLH129 | Zm00001eb229950 | ||||
| BHLH29 | transcription factor | BHLH100 | Zm00001eb420910 | overexpression | [105] |
| BHLH101 | Zm00001eb085690 | ||||
| BHLH300 | transcription factor | BHLH54 | Zm00001eb362800 | overexpression | [106] |
| IDEF1 | transcription factor | ABI47 | Zm00001eb198710 | overexpression | [107] |
| ABI49 | Zm00001eb259870 | ||||
| CSN6 | coactivator | SI605023C06B | Zm00001eb199540 Zm00001eb034040 | knockout | [108] |
| RAB6A | small GTPase | IDP871 | Zm00001eb006940 | overexpression | [109] |
| HRZ | Fe-binding ubiquitin ligase | 541975 | Zm00001eb360580 Zm00001eb156300 Zm00001eb294920 | knockout | [110] |
| MTP1 | carrier protein | UMC2311 | Zm00001eb265000 Zm00001eb385520 Zm00001eb420140 Zm00001eb354910 | overexpression | [111] |
| ZIF1 | carrier protein | MFSD1 MFSD2 IDP8516 TOM3 UMC1028 IDP7324 YS3 IDP6979 | Zm00001eb129050 Zm00001eb196170 Zm00001eb038000 Zm00001eb093430 Zm00001eb093440 Zm00001eb128730 Zm00001eb133440 Zm00001eb163460 Zm00001eb129340 Zm00001eb196180 Zm00001eb332620 | overexpression | [112] |
| ZIF2 | carrier protein | PCO099415 | Zm00001eb017730 | overexpression | [113] |
| GPM828 | Zm00001eb017760 | ||||
| HMA2 | carrier protein | HMA2 | Zm00001eb226870 | overexpression | [114] |
| HMA4 | carrier protein | HMA3 | Zm00001eb095020 | overexpression | [115] |
| HMA7 | carrier protein | CSU904 | Zm00001eb327860 | overexpression | [116] |
| ZIP1 | carrier protein | Zm00001eb139810 | overexpression | [117] | |
| ZIP8 | carrier protein | ZIP8 | Zm00001eb303800 | knockout | [118] |
| MIPS | myo-inositol-1-phosphate synthase | MIPS2 | Zm00001eb401220 Zm00001eb276490 Zm00001eb283250 Zm00001eb378070 | knockout | [124] |
| PGK1 | 2-phosphoglycerate kinase | Zm00001eb191270 Zm00001eb259060 | knockout | [125] | |
| ITPK2 | inositol 1,3,4-trisphosphate 5/6-kinase | Zm00001eb399350 | knockout | [126] | |
| IPK1 | inositol 1,3,4,5,6-pentakisphosphate 2-kinase | IDP8938 | Zm00001eb067500 Zm00001eb432760 | knockout | [127] |
| HAD1 | phytase | Zm00001eb063350 Zm00001eb342820 Zm00001eb399750 | overexpression | [128] | |
| PAPHY-A | phytase | PAP2 | Zm00001eb064450 | overexpression | [129] |
| PAP4 | phytase | PAP22 | Zm00001eb048820 | overexpression | [130] |
| Genes | Protein Function | Maize Orthologs | Gene ID | Strategy | References |
|---|---|---|---|---|---|
| GLK1 | transcription factor | G2 | Zm00001eb118900 | overexpression | [133] |
| GLK1 | Zm00001eb371980 | ||||
| AN1 | transcription factor | IN1 | Zm00001eb303250 | overexpression | [134] |
| AN3 | transcription factor | GIF1 | Zm00001eb056300 | overexpression | [135] |
| ANT1 | transcription factor | PL1 | Zm00001eb278680 | overexpression | [136] |
| C1 | Zm00001eb373660 | ||||
| MYBR | transcription factor | MYB31 | Zm00001eb103730 | knockout | [137] |
| MYB42 | Zm00001eb202770 | ||||
| DRB3 | double stranded RNA binding protein | IDP7470 | Zm00001eb102530 | knockout | [138] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, Q.; Zhang, T.; Sun, K.; Yan, T.; Wang, L.; Lu, L.; Zhao, W.; Qi, Y.; Long, Y.; Wei, X.; et al. Mining of Potential Gene Resources for Breeding Nutritionally Improved Maize. Plants 2022, 11, 627. https://doi.org/10.3390/plants11050627
Hou Q, Zhang T, Sun K, Yan T, Wang L, Lu L, Zhao W, Qi Y, Long Y, Wei X, et al. Mining of Potential Gene Resources for Breeding Nutritionally Improved Maize. Plants. 2022; 11(5):627. https://doi.org/10.3390/plants11050627
Chicago/Turabian StyleHou, Quancan, Tianye Zhang, Kangtai Sun, Tingwei Yan, Linlin Wang, Lu Lu, Wei Zhao, Yuchen Qi, Yan Long, Xun Wei, and et al. 2022. "Mining of Potential Gene Resources for Breeding Nutritionally Improved Maize" Plants 11, no. 5: 627. https://doi.org/10.3390/plants11050627