
Circumscription and Phylogenetic Position of Two Propagulose Species of Syntrichia (Pottiaceae, Bryophyta) Reveals Minor Realignments within the Tribe Syntricheae


Abstract
:1. Introduction
2. Materials and Methods
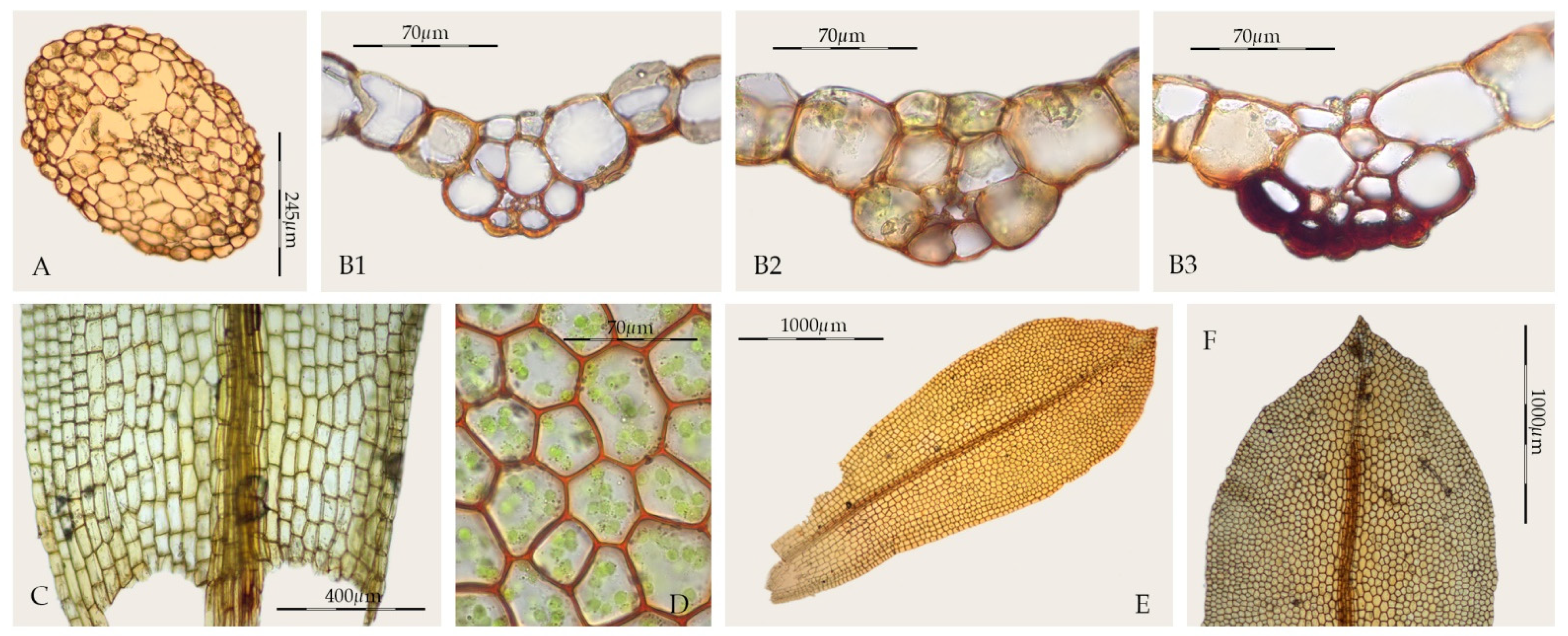
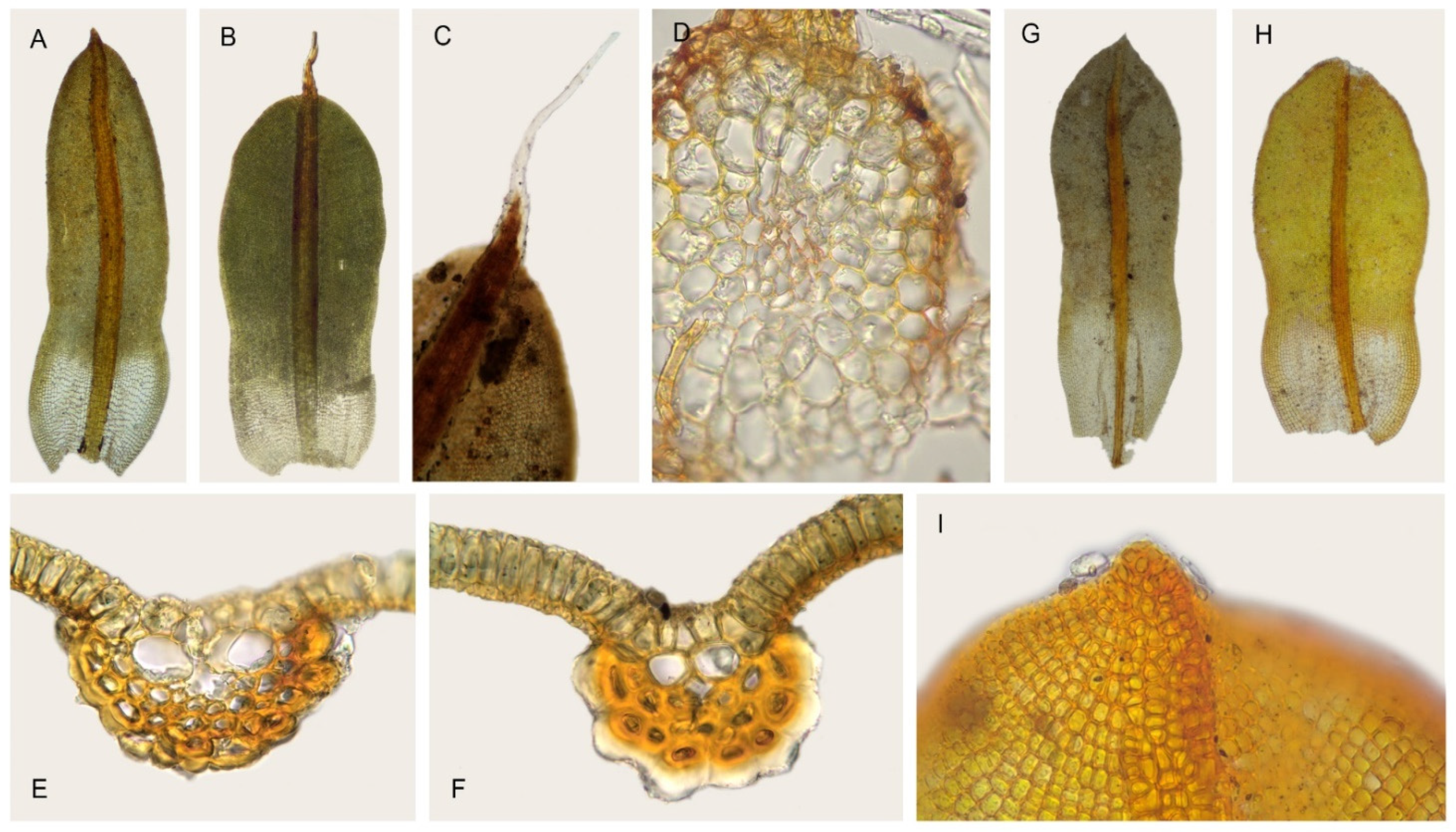
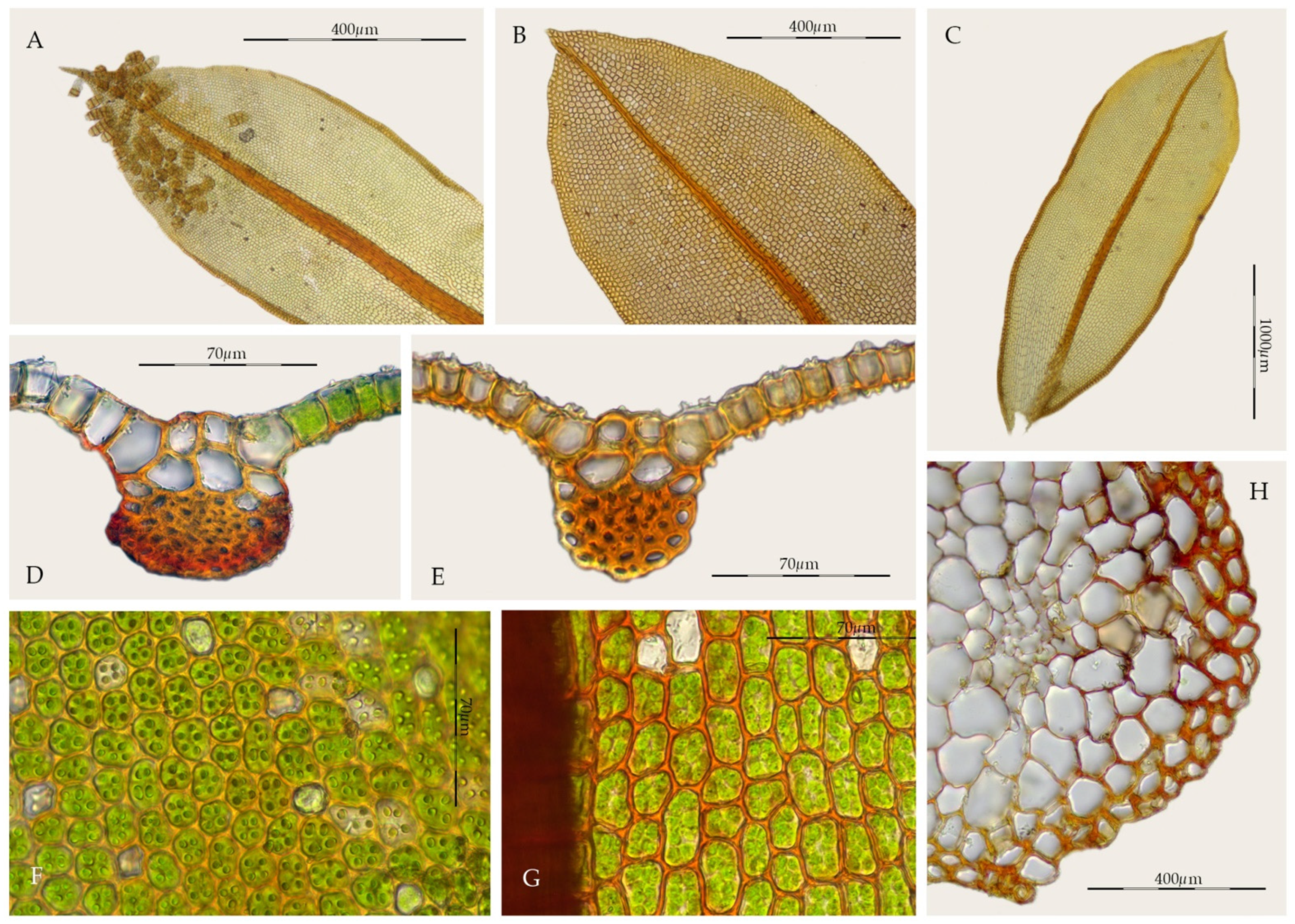
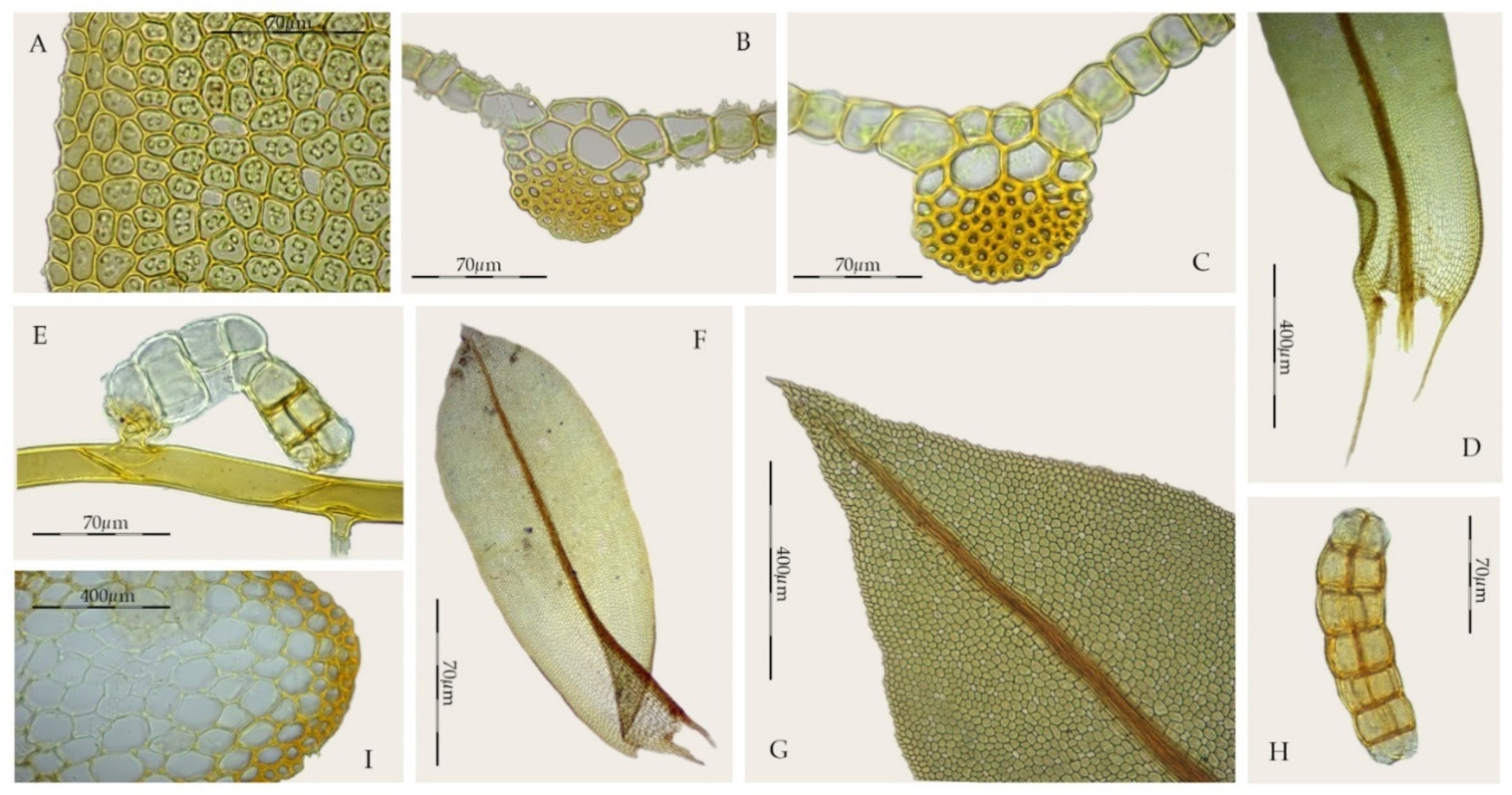
2.1. Morphological Study
2.2. Molecular Taxon Sampling
2.3. DNA Extraction, Amplification, and Sequencing
2.4. Phylogenetic Analysis Sequencing
3. Results
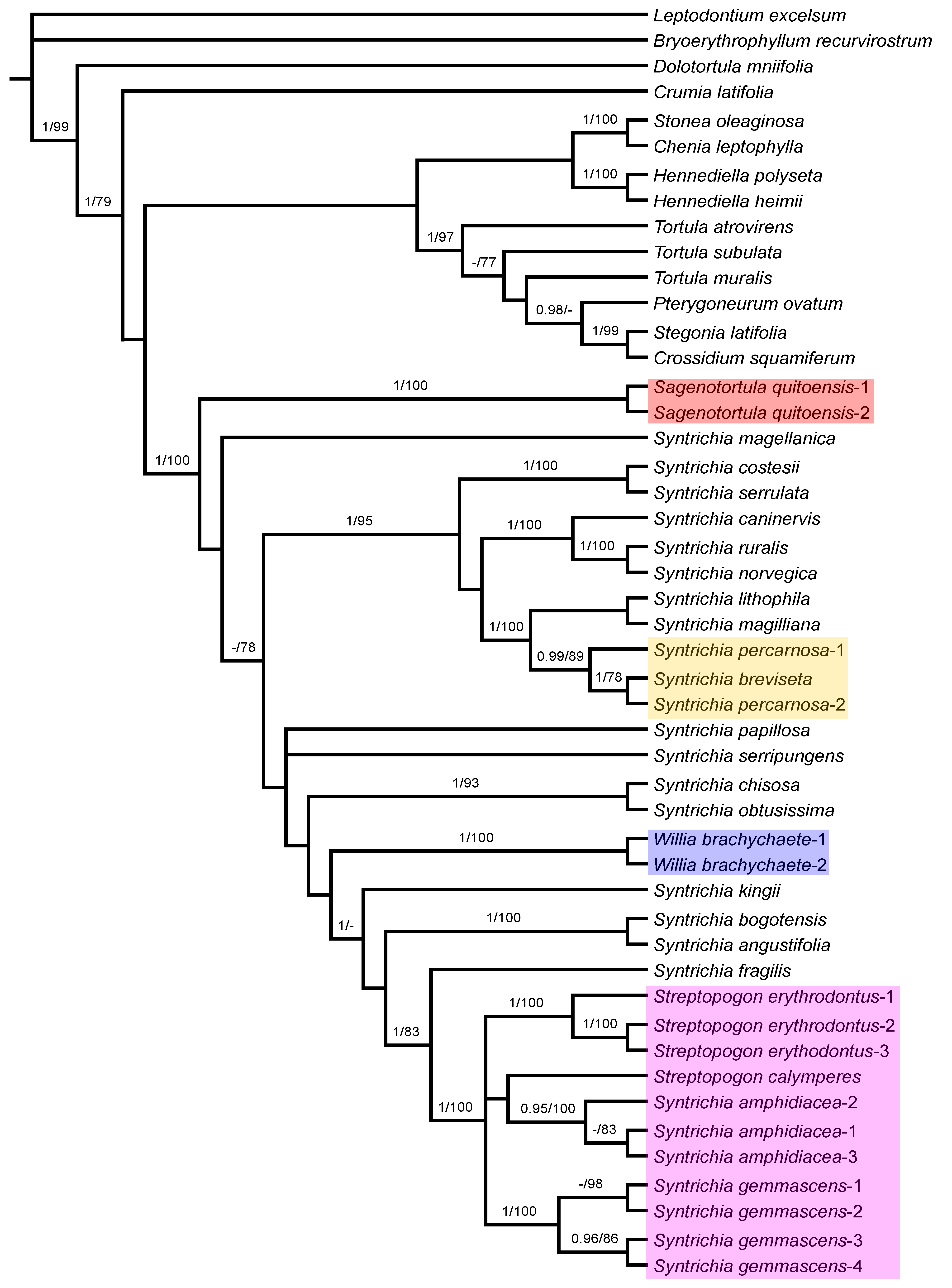
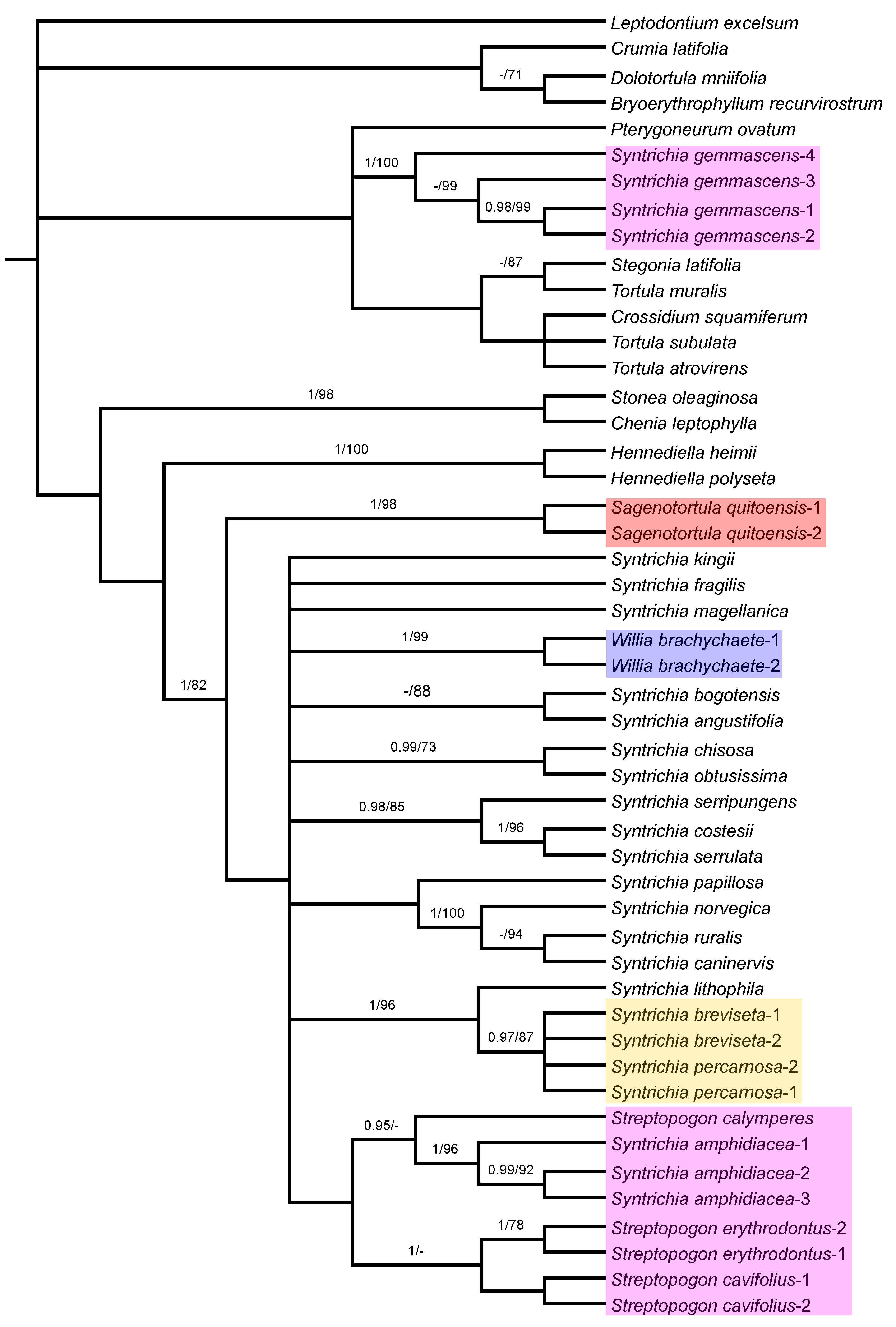
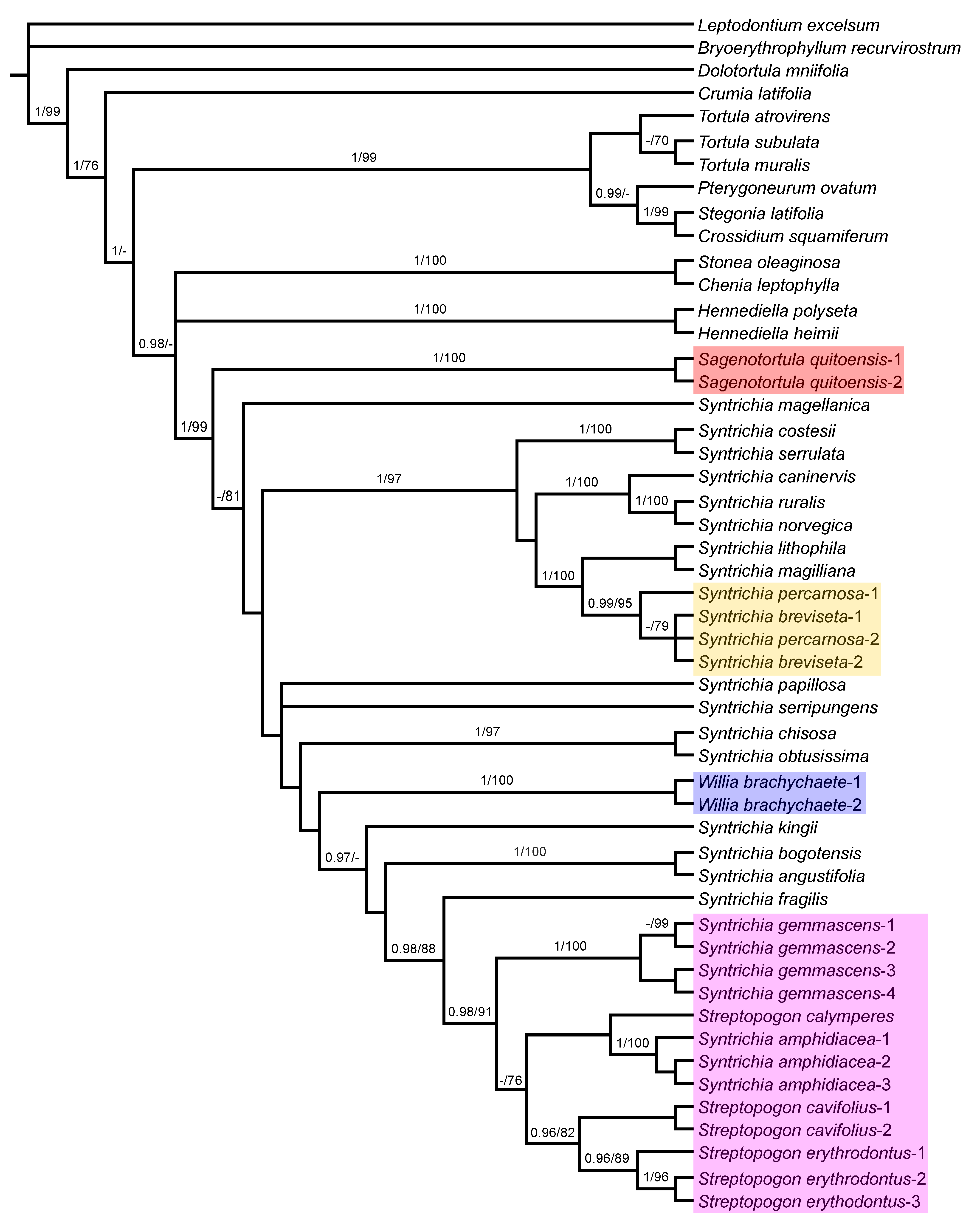
3.1. Phylogenetic Analyses
3.2. Analysis of ITS Sequences
3.3. Analysis of trnG–trnL-F Sequences
3.4. Analysis of Combined Plastid and ITS Sequences
4. Discussion and Conclusions
5. Taxonomic Changes
5.1. New Combinations in Streptopogon
5.2. New Synonyms for Syntrichia
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Zander, R.H. The Pottiaceae s. str. as an evolutionary Lazarus taxon. J. Hattori Bot. Lab. 2006, 100, 581–602. [Google Scholar]
- Frey, W.; Stech, M. Marchantiophyta, Bryophyta, Anthocerotophyta. In Syllabus of Plant Families, A. Engler’s Syllabus der Pflanzenfamilien, 13th ed.; Part 3 Bryophytes and Seedless Vascular Plants; Frey, W., Ed.; Gebr. Borntraeger: Stuttgart, Germany, 2009; pp. 1–263. [Google Scholar]
- Zander, R.H. Genera of the Pottiaceae: Mosses of harsh environments. Bull. Buffalo Soc. Nat. Sci. 1993, 32, 1–378. [Google Scholar]
- Goffinet, B.; Buck, W.R.; Shaw, A.J. Morphology, anatomy, and classification of the Bryophyta. In Bryophyte Biology, 2nd ed.; Goffinet, B., Shaw, A.J., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 55–138. [Google Scholar]
- Saito, K. A monograph of Japanese Pottiaceae (Musci). J. Hattori Bot. Lab. 1975, 39, 373–537. [Google Scholar]
- Cano, M.J.; Jiménez, J.A.; Gallego, M.T.; Guerra, J. A molecular approach to the phylogeny of the moss genus Pseudocrossidium (Pottiaceae, Bryopsida) and its taxonomic implications. J. Syst. Evol. 2021. [CrossRef]
- Gallego, M.T.; Hugonnot, V.; Cano, M.J. Taxonomic resurrection of an awnless variety of Syntrichia ruralis and comparison with other European muticous taxa in this genus. J. Bryol. 2018, 40, 244–250. [Google Scholar] [CrossRef]
- Gallego, M.T.; Cano, M.J. Syntrichia splendida M.T.Gallego & M.J.Cano (Pottiaceae), a new moss species from northern Chile. J. Bryol. 2021, 43, 277–282. [Google Scholar] [CrossRef]
- Zander, R.H. Seven new genera in Pottiaceae (Musci) and a lectotype for Syntrichia. Phytologia 1989, 65, 424–436. [Google Scholar]
- Ochyra, R. New combinations in Syntrichia and Warnstorfia (Musci). Fragm. Florist. Geobot. 1992, 37, 211–214. [Google Scholar]
- Spagnuolo, V.; Caputo, P.; Cozzolino, R.; Castaldo, R.; De Luca, P. Patterns of relationships in Trichostomoideae (Pottiaceae, Musci). Plant Syst. Evol. 1999, 216, 69–79. [Google Scholar] [CrossRef]
- Werner, O.; Ros, R.M.; Cano, M.J.; Guerra, J. Tortula and some related genera (Pottiaceae, Musci): Phylogenetic relationships based on chloroplast rps4 sequences. Pl. Syst. Evol. 2002, 235, 197–207. [Google Scholar] [CrossRef]
- Afonina, O.M.; Ignatova, E.A.; Fedosov, V.E.; Kuznetsova, O.I. Toward a new understanding of Syntrichia submontana (Pottiaceae, Bryophyta). Arctoa 2014, 23, 11–24. [Google Scholar] [CrossRef]
- Gallego, M.T.; Cano, M.J.; Jiménez, J.F.; Jiménez, J.A.; Guerra, J. Morphological and molecular data support a new combination in the Neotropical complex of cucullate-leaved species of Syntrichia (Pottiaceae). Syst. Bot. 2014, 39, 361–368. [Google Scholar] [CrossRef]
- Hedenäs, L.; Heinrichs, J.; Gallego, M.T. The Scandinavian Syntrichia ruralis complex (Musci, Pottiaceae): A chaos of diversification. Plant Syst. Evol. 2019, 305, 639–661. [Google Scholar] [CrossRef] [Green Version]
- Ochyra, R.; Zander, R.H. Is Tortula lithophila conspecific with Sarconeurum glaciale (Bryopsida: Pottiaceae)? Frag. Florist. Geobot. Polon. 2007, 14, 209–212. [Google Scholar]
- Cano, M.J. Typification and taxonomical identity of some infraspecific name related to Tortula subulata complex (Pottiaceae, Bryophyta). Taxon 2007, 56, 949–952. [Google Scholar] [CrossRef] [Green Version]
- Cano, M.J. Taxonomic revision of Hennediella Paris (Pottiaceae, Bryophyta). Nova Hedwig. 2008, 87, 3–4. [Google Scholar]
- Cano, M.J.; Gallego, M.T. The genus Tortula (Pottiaceae, Bryophyta) in South America. Bot. J. Linn. Soc. 2008, 156, 173–220. [Google Scholar] [CrossRef]
- Matteri, C.M. New combination and new synonyms in Fuegian mosses. Lindbergia 2003, 28, 80–82. [Google Scholar] [CrossRef]
- Gallego, M.T.; Cano, M.J.; Guerra, J. New synonymy in Syntrichia (Pottiaceae) in the Neotropics. Bryologist 2009, 112, 173–177. [Google Scholar] [CrossRef]
- Gallego, M.T.; Cano, M.J.; Guerra, J. New records, synonyms and one combination in the genus Syntrichia (Pottiaceae) from South America. Bryologist 2011, 114, 556–562. [Google Scholar] [CrossRef]
- Brinda, J.C.; Jáuregui-Lazo, J.A.; Oliver, M.J.; Mishler, B.D. Notes on the genus Syntrichia with a revised infrageneric classification and the recognition of a new genus Syntrichiadelphus (Bryophyta, Pottiaceae). Phytologia 2021, 103, 90–103. [Google Scholar]
- Zander, R.H. Macroevolutionary versus molecular analysis: Systematics of the Didymodon segregates Aithobryum, Exobryum and Fuscobryum (Pottiaceae, Bryophyta). Hattoria 2019, 10, 1–38. [Google Scholar] [CrossRef]
- Pottiaceae, Integrated Taxonomic Information System. Available online: http://www.pottiaceae.com/index.php?mod=field_trips (accessed on 9 January 2022).
- Casado, C.M. A Taxonomic Revision of Streptopogon Wils. (Pottiaceae), Thesis; University of Missouri: Columbia, MO, USA, 2000. [Google Scholar]
- Costa, D.P. Validation of the new species of Streptopogon (Pottiaceae, Bryophyta) and a synopsis of the genus for Brazil. Syst. Bot. 2012, 37, 583–586. [Google Scholar] [CrossRef]
- Salmon, E.S. A monograph of the genus Streptopogon, Wils. Ann. Bot. 1903, 17, 107–150. [Google Scholar] [CrossRef]
- Jiménez, J.A.; Cano, M.J.; Guerra, J. A multilocus phylogeny of the moss genus Didymodon and allied genera (Pottiaceae): Generic delimitations and their implications for systematics. J. Syst. Evol. 2021. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Suzuki, T.; Inoue, Y.; Tsuba, H.; Iwatsuki, Z. Notes on Aptychella (Sematophyllaceae, Bryopsida): Yakushimabryum longissimum, syn. nov. Hattoria 2013, 4, 107–118. [Google Scholar]
- Cano, M.J.; Jiménez, J.F.; Gallego, M.T.; Jiménez, J.A.; Guerra, J. Phylogenetic relationships in the genus Hennediella (Pottiaceae, Bryophyta) inferred from nrITS sequence data. Pl. Syst. Evol. 2009, 281, 209–216. [Google Scholar] [CrossRef]
- Pacak, A.; Szweykowska-Kulińska, Z. Molecular data concerning the allopolyploid character and the origin of chloroplast and mithochondrial genomes in the liverwort species Pellia borealis. J. Pl. Biotech. 2000, 2, 101–108. [Google Scholar]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Pl. Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef]
- Stech, M.; Frahm, J.P. The status of Platyhypnidium mutatum Ochyra & Vanderpoorten and the systematic value of Donrichardsiaceae based on molecular data. J. Bryol. 1999, 21, 191–195. [Google Scholar] [CrossRef]
- Sawicki, J.; Szczecińska, M. A comparison of PCR-based markers for molecular identification of Sphagnum species of the section Acutifolia. Acta Soc. Bot. Pol. 2011, 80, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Olsson, S.; Buchbender, V.; Enroth, J.; Hedenäs, L.; Huttunen, S.; Quandt, D. Phylogenetic analyses reveal high levels of polyphyly among pleurocarpous lineages as well as novel clades. Bryologist 2009, 112, 447–466. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data (version 9.1.8). Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucl. Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Müller, K. SeqState: Primer design and sequence statistics for phylogenetic DNA data sets. Appl. Bioinform. 2005, 4, 65–69. [Google Scholar] [CrossRef]
- Simmons, M.P.; Ochoterena, H. Gaps as characters in sequence based phylogenetic analyses. Syst. Biol. 2000, 49, 369–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol Evol. 1993, 10, 512–526. [Google Scholar]
- Stamatakis, A. Raxml version 8: A tool for phylogenetic analysis andpost-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. raxmlGUI 2.0 beta: A graphical interface and toolkit for phylogenetic analyses using RAxML. bioRxiv 2020, 12, 373–377. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES science gateway: Enabling high-impact science for phylogenetics researchers with limited resources. In Proceedings of the Conference of the Extreme Science and Engineering Discovery Environment: Bridging from the Extreme to the Campus and Beyond, Association for Computing Machinery, Atlanta, GA, USA, 16 July 2012; pp. 1–8. [Google Scholar]
- Rambaut, A. FigTree, Version 1.4.4. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 13 November 2021).
- Mishler, B.D. Tortula. In The Moss Flora of Mexico; Sharp, A.J., Crum, H., Eckel, P.M., Eds.; The New York Botanical Garden: New York, NY, USA, 1994; Volume 1, pp. 319–350. [Google Scholar]
- Montagne, C. Cinquième centurie de plantes cellulaires exotiques nouvelles. Ann. Sci. Nat. Bot. Sér. 3 1845, 4, 86–123. [Google Scholar]
- Ochyra, R.; Lewis Smith, R.I.; Bednarek-Ochyra, H. The Illustrated Moss Flora of Antarctica; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Müller, C. Prodromus bryologiae Argentinicae. I. Linnaea 1879, 42, 217–486. [Google Scholar]
- Brotherus, V.F. Bryales. In Die Natürlichen Pflanzenfamilien; Engler, H.G.A., Prantl, K.A.E., Eds.; Verlag von Wilhelm Engelmann: Leipzig, Germany, 1902; Volume 1, pp. 385–432. [Google Scholar]
- Anderson, L.E. Syntrichia magilliana (Pottiaceae), a new species from South Africa. J. Hattori Bot. Lab. 1997, 82, 15–18. [Google Scholar]
- Van Rooy, J.; Perold, S.M. Bryophyta. In A Checklist of South African plants, Germishuizen, G.; Meyer, N.L., Steenkamp, Y., Keith, M., Eds.; South African Botanical Diversity, Network Report; South African National Biodiversity Institute: Pretoria, South Africa, 2006; Volume 41, pp. 1–52. [Google Scholar]
- Crum, H.A. Two new species of Mexican mosses collected by Aaron, J. Sharp. Bryologist 1952, 55, 51–58. [Google Scholar] [CrossRef]
- Müller, F. An updated checklist of the mosses of Chile. Arch. Bryol. 2009, 58, 1–124. [Google Scholar]
- Maddison, W.P. Gene trees in species trees. Syst. Biol. 1997, 46, 523–536. [Google Scholar] [CrossRef]
- Košnar, J.; Herbstová, M.; Kolář, F.; Koutecký, P.; Kučera, J. A case of intragenomic ITS variation in bryophytes: Assessment of gene flow and role of plyploidy in the origin of European taxa of the Tortula muralis (Musci: Pottiaceae) complex. Taxon 2012, 61, 709–720. [Google Scholar] [CrossRef]
- Afonina, O.M.; Ignatova, E.A. Syntrichia amphidiacea (Pottiaceae), a new species for the moss flora of Russia. Bot. Zhur. (St. Petersburg) 2009, 94, 439–443. [Google Scholar]
- Andrews, A.L. Tortula caroliniana, new species. Bryologist 1920, 5, 72–76. [Google Scholar] [CrossRef]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.H.; Li, D.Z.; Marhold, K.; et al. International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017, Regnum Veg.; Koeltz Botanical Books: Glashütten, Germany, 2018; Volume 159. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Number of Specimens | New Generated Sequences | Sequence Length | Parsimony-Informative Characters |
|---|---|---|---|---|
| ITS | 48 | 35 | 1908 | 802 (42.03%) |
| trnG | 43 | 22 | 647 | 91 (14%) |
| trnL-F | 48 | 25 | 459 | 62 (13.50%) |
| Nuclear + plastid | 51 | 3014 | 1213 (36.8%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallego, M.T.; Cano, M.J.; Jiménez, J.A.; Guerra, J. Circumscription and Phylogenetic Position of Two Propagulose Species of Syntrichia (Pottiaceae, Bryophyta) Reveals Minor Realignments within the Tribe Syntricheae. Plants 2022, 11, 626. https://doi.org/10.3390/plants11050626
Gallego MT, Cano MJ, Jiménez JA, Guerra J. Circumscription and Phylogenetic Position of Two Propagulose Species of Syntrichia (Pottiaceae, Bryophyta) Reveals Minor Realignments within the Tribe Syntricheae. Plants. 2022; 11(5):626. https://doi.org/10.3390/plants11050626
Chicago/Turabian StyleGallego, M. Teresa, María J. Cano, Juan A. Jiménez, and Juan Guerra. 2022. "Circumscription and Phylogenetic Position of Two Propagulose Species of Syntrichia (Pottiaceae, Bryophyta) Reveals Minor Realignments within the Tribe Syntricheae" Plants 11, no. 5: 626. https://doi.org/10.3390/plants11050626