
In Vitro Technologies for American Chestnut (Castanea dentata (Marshall) Borkh) Conservation


Abstract
:1. Introduction
2. Results
2.1. Micropropagation
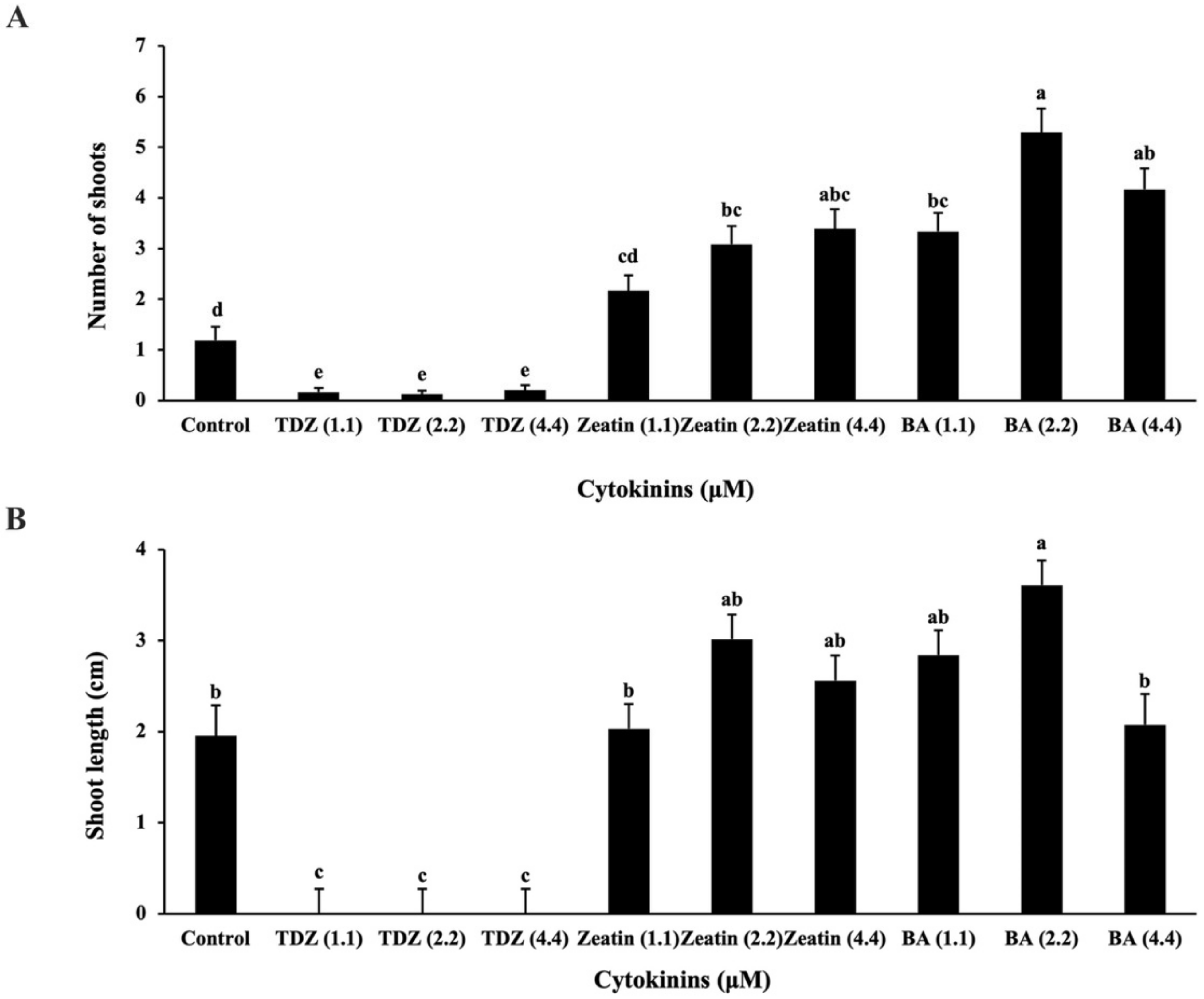
2.1.1. Establishment of Shoot Cultures
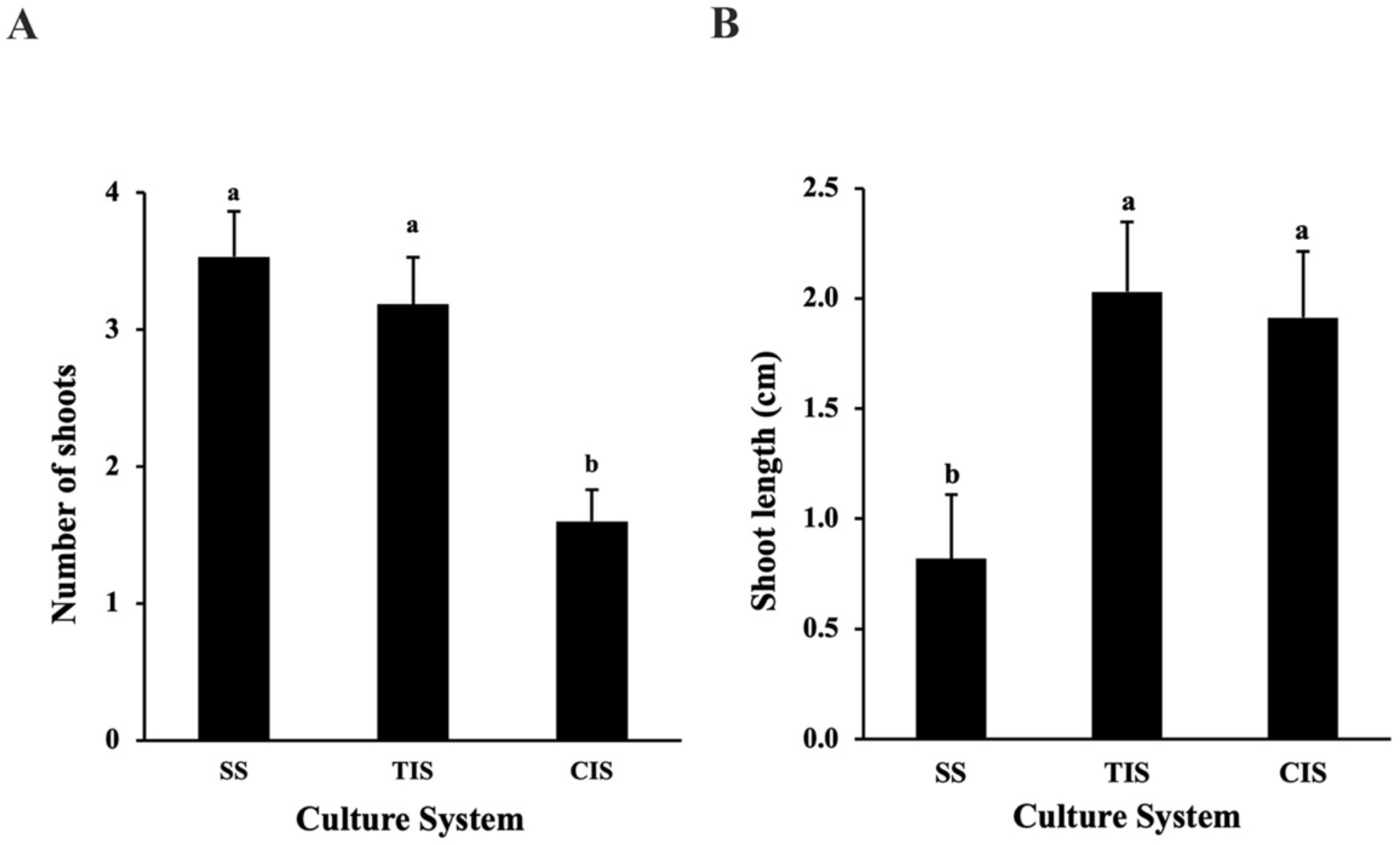
2.1.2. Comparison of Semi-Solid System (SS), Temporary Immersion System (TIS), and Continuous Immersion System (CIS) for Shoot Multiplication and Growth
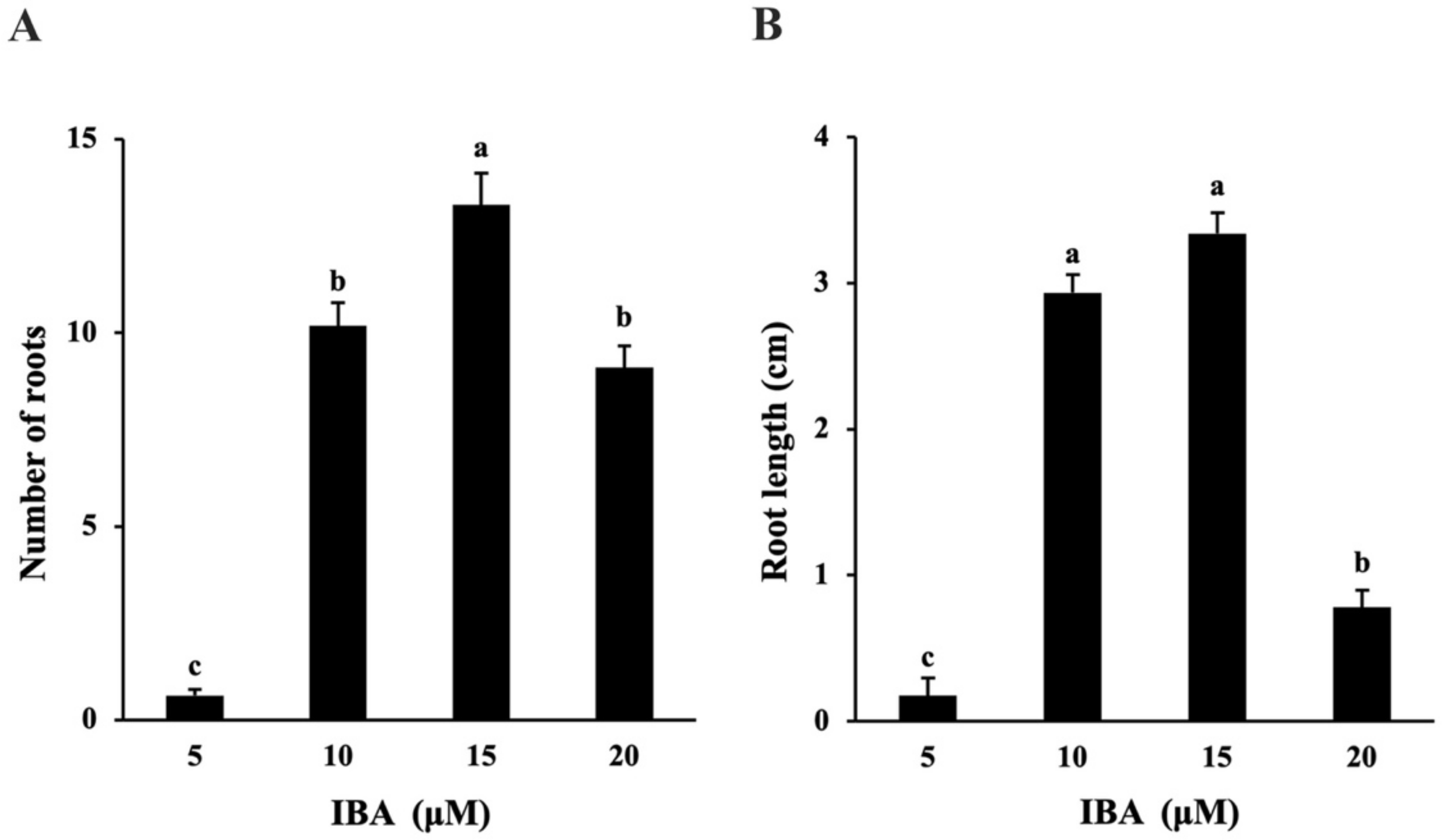
2.1.3. In Vitro Rooting and Acclimatization
2.2. Cryopreservation
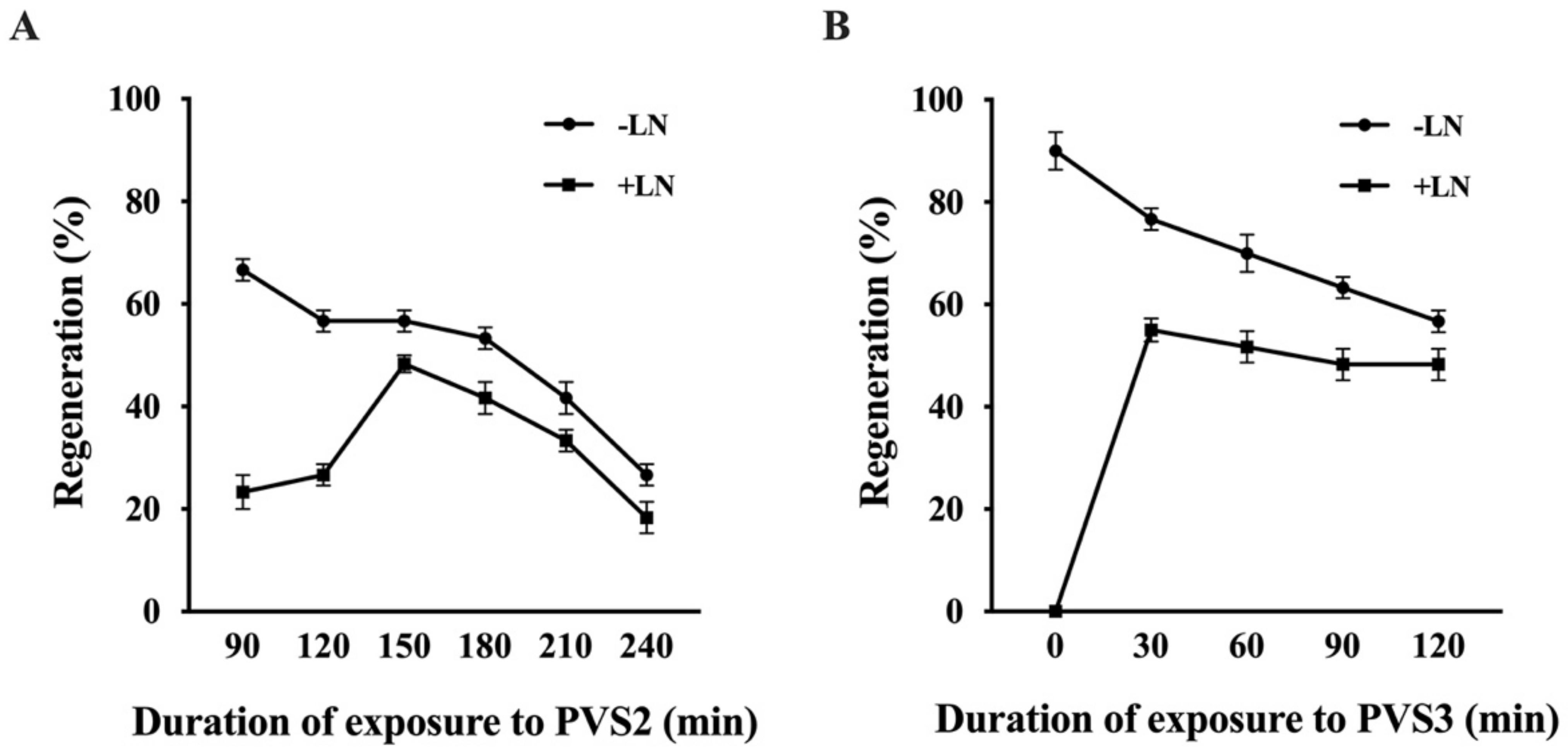
2.2.1. Preculture
2.2.2. Dehydration
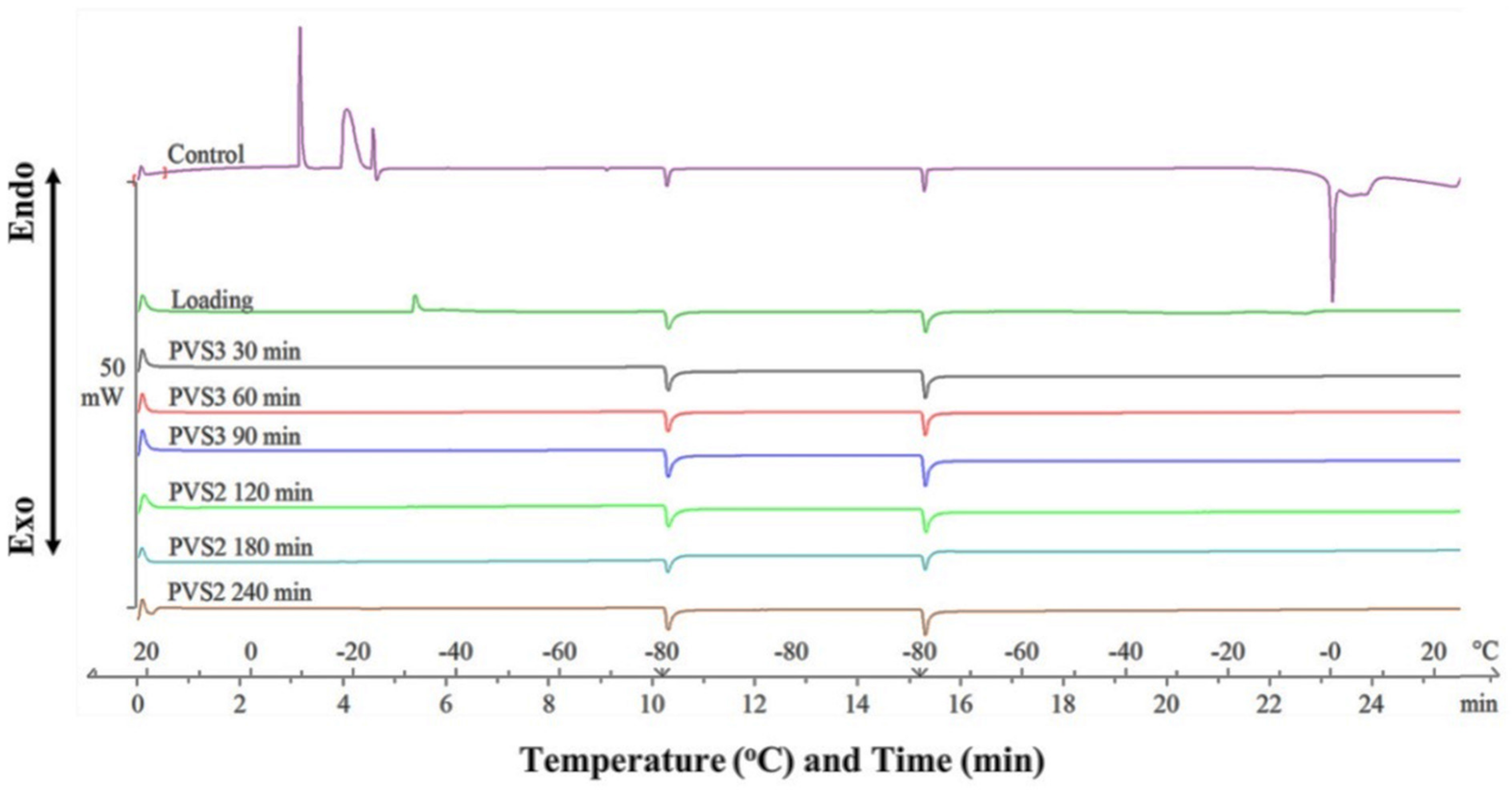
2.2.3. Differential Scanning Calorimetry Analysis
3. Discussion
4. Materials and Methods
4.1. Micropropagation
4.1.1. Initiation and Establishment of Shoot Cultures
4.1.2. Shoot Proliferation
4.1.3. Comparison of Semi-Solid System (SS), Continuous Immersion System (CIS), and Temporary Immersion System (TIS) for In vitro Growth
4.1.4. Rooting and Plantlet Development
4.1.5. Greenhouse Acclimatization
4.2. Cryopreservation
Differential Scanning Calorimetry Analysis
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saucier, J.R. American chestnut: An American wood. USDA For. Serv. 1973, FS-230, 230. [Google Scholar]
- COSEWIC Assessment and update status report on the American chestnut Castanea dentata in Canada. Available online: https://www.canada.ca/en/environment-climate-change/services/species-risk-public-registry/cosewic-assessments-status-reports/american-chestnut.html (accessed on 12 December 2021).
- Van Drunen, S.G.; Schutten, K.; Bowen, C.; Boland, G.J.; Husband, B.C. Population dynamics and the influence of blight on American chestnut at its northern range limit: Lessons for conservation. For. Ecol. Manag. 2017, 400, 375–383. [Google Scholar] [CrossRef]
- Boland, G.J.; Ambrose, J.; Husband, B.; Elliott, K.A.; Melzer, M.S. Recovery Strategy for the American Chestnut (Castanea Dentata) in Ontario; Ontario Recovery Strategy Series; Ontario Ministry of Natural Resources: Peterborough, ON, Canada, 2012; p. vi–43. [Google Scholar]
- Saxena, A.; Shukla, M.; Saxena, P. Synthetic Seeds: Relevance to endangered germplasm conservation in vitro. In Synthetic Seeds; Springer: Cham, Switzerland, 2019; pp. 21–60. [Google Scholar]
- Bi, W.; Saxena, A.; Ayyanath, M.-M.; Harpur, C.; Shukla, M.R.; Saxena, P.K. conservation, propagation, and redistribution (CPR) of hill’s thistle: Paradigm for plant species at risk. Plant Cell Tissue Organ Cult. 2021, 145, 75–88. [Google Scholar] [CrossRef]
- Saxena, A.; Bi, W.-L.; Shukla, M.R.; Cannings, S.; Bennett, B.; Saxena, P.K. Micropropagation and cryopreservation of Yukon Draba (Draba yukonensis), a Special Concern Plant Species Endemic to Yukon Territory, Canada. Plants 2021, 10, 2093. [Google Scholar] [CrossRef] [PubMed]
- Vieitez, E. Vegetative propagation of chestnut. NZJ For. Sci. 1974, 4, 242–252. [Google Scholar]
- Keys, R.N. Prospects for Vegetative Propagation in the Genus Castanea. In Proceedings of the American Chestnut Symposium, Morgantown, WV, USA, 4–5 January 1978; MacDonald, W.L., Cech, F.C., Luchok, J., Smith, C., Eds.; West Virginia University: Morgantown, WV, USA, 1978. [Google Scholar]
- Hussain, A.; Qarshi, I.A.; Nazir, H.; Ullah, I. Plant tissue culture: Current status and opportunities. Recent Adv. Plant Vitr. Culture 2012, 6, 1–28. [Google Scholar]
- Yang, G.; Lu, Z.; Asante, T.; Read, P. In vitro responses of American chestnut to plant growth regulators in culture medium. Acta Hortic. 2009, 844, 229–234. [Google Scholar] [CrossRef]
- Xing, Z.; Satchwell, M.F.; Powell, W.A.; Maynard, C.A. Micropropagation of American chestnut: Increasing rooting rate and preventing shoot-tip necrosis. Vitr. Cell. Dev. Biol. Plant 1997, 33, 43–48. [Google Scholar] [CrossRef]
- Oakes, A.D. An Investigation of Micropropagation Techniques for American Chestnut; State University of New York College of Environmental Science and Forestry: Syracuse, NY, USA, 2015; ISBN 1-339-48292-4. [Google Scholar]
- Oakes, A.D.; Powell, W.A.; Maynard, C.A. Doubling acclimatization survival of micropropagated American chestnuts with darkness and shortened rooting induction time. J. Environ. Hortic. 2013, 31, 77–83. [Google Scholar] [CrossRef]
- Kong, L.; Holtz, C.T.; Nairn, C.J.; Houke, H.; Powell, W.A.; Baier, K.; Merkle, S.A. Application of airlift bioreactors to accelerate genetic transformation in American chestnut. Plant Cell Tiss Organ Cult. 2014, 117, 39–50. [Google Scholar] [CrossRef]
- Johnson, S.B.; Le, H.T.; Kormanik, T.L.; Merkle, S.A. Advances in American chestnut somatic seedling production. In Proceedings of the 29th Southern Forest Tree Improvement Conference Tree Improvement in North America: Past, Present, and Future, Galveston, TX, USA, 19–22 June 2007; pp. 85–95. [Google Scholar]
- Andrade, G.M.; Merkle, S.A. Enhancement of American chestnut somatic seedling production. Plant Cell Rep. 2005, 24, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Xing, Z.; Powell, W.A.; Maynard, C.A. Development and germination of American chestnut somatic embryos. Plant Cell Tissue Organ Cult. 1999, 57, 47–55. [Google Scholar] [CrossRef]
- Robichaud, R.L.; Lessard, V.C.; Merkle, S.A. Treatments affecting maturation and germination of American chestnut somatic embryos. J. Plant Physiol. 2004, 161, 957–969. [Google Scholar] [CrossRef]
- Carraway, D.T.; Merkle, S.A. Plantlet regeneration from somatic embryos of American chestnut. Can. J. For. Res. 1997, 27, 1805–1812. [Google Scholar] [CrossRef]
- Rothrock, R.E.; Polin-McGuigan, L.D.; Newhouse, A.E.; Powell, W.A.; Maynard, C.A. Plate flooding as an alternative agrobacterium-mediated transformation method for American chestnut somatic embryos. Plant Cell Tissue Organ Cult. 2007, 88, 93–99. [Google Scholar] [CrossRef]
- Nagori, R.; Rathore, P.; Vyas, S.; Joshi, N.; Rao, M.S.; Suthar, R.; Mathur, P.; Purohit, S.D. Liquid Culture System Stimulates In Vitro Growth and Shoot Multiplication in Some Medicinally Important Plant Species of Aravallis in Rajasthan; Professor HC Arya Commemoration Volume; IK International Publishers: New Delhi, India, 2009; pp. 395–405. [Google Scholar]
- Mbiyu, M.; Muthoni, J.; Kabira, J.; Muchira, C.; Pwaipwai, P.; Ngaruiya, J.; Onditi, J.; Otieno, S. Comparing liquid and solid media on the growth of plantlets from three Kenyan potato cultivars. J. Exp. Agric. Int. 2012, 2, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Pati, P.K.; Kaur, J.; Singh, P. A Liquid culture system for shoot proliferation and analysis of pharmaceutically active constituents of Catharanthus roseus (L.) G. Don. Plant Cell Tissue Organ Cult. 2011, 105, 299–307. [Google Scholar] [CrossRef]
- Berthouly, M.; Etienne, H. Temporary Immersion System: A new concept for use liquid medium in mass propagation. In Liquid Culture Systems For In Vitro Plant Propagation; Springer: Berlin/Heidelberg, Germany, 2005; pp. 165–195. [Google Scholar]
- Etienne, H.; Berthouly, M. Temporary immersion systems in plant micropropagation. Plant Cell Tissue Organ Cult. 2002, 69, 215–231. [Google Scholar] [CrossRef]
- Arencibia, A.D.; Bernal, A.; Yang, L.; Cortegaza, L.; Carmona, E.R.; Pérez, A.; Hu, C.-J.; Li, Y.-R.; Zayas, C.M.; Santana, I. New role of phenylpropanoid compounds during sugarcane micropropagation in temporary immersion bioreactors (TIBs). Plant Sci. 2008, 175, 487–496. [Google Scholar] [CrossRef]
- Georgiev, V.; Schumann, A.; Pavlov, A.; Bley, T. Temporary immersion systems in plant biotechnology. Eng. Life Sci. 2014, 14, 607–621. [Google Scholar] [CrossRef]
- Akula, A.; Becker, D.; Bateson, M. High-yielding repetitive somatic embryogenesis and plant recovery in a selected tea clone,‘TRI-2025’, by temporary immersion. Plant Cell Rep. 2000, 19, 1140–1145. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Liu, C.; Romero, R.M.; Saxena, P.K. In vitro culture and temporary immersion bioreactor production of Crescentia cujete. Plant Cell Tissue Organ Cult. 2004, 78, 63–68. [Google Scholar] [CrossRef]
- Posada-Pérez, L.; Montesinos, Y.P.; Guerra, D.G.; Daniels, D.; Gómez-Kosky, R. Complete germination of papaya (Carica papaya L. cv. maradol roja) somatic embryos using temporary immersion system type RITA® and phloroglucinol in semi-solid culture medium. Vitr. Cell. Dev. Biol. Plant 2017, 53, 505–513. [Google Scholar] [CrossRef]
- Shandil, A.S.; Tuia, V.S. Micropropagation of breadfruit (A. altilis) enhanced using a bioreactor system. Acta Hortic. 2015, 1101, 159–164. [Google Scholar] [CrossRef]
- Chakrabarty, D.; Dewir, Y.H.; Dewir, E.J.; Hahn, S.K.; Paek, K.Y. The dynamics of nutrient utilization and growth of apple root stock ‘M9 EMLA’ in temporary versus continuous immersion bioreactors. Plant Growth Regul. 2007, 51, 11–19. [Google Scholar] [CrossRef]
- Latawa, J.; Shukla, M.R.; Saxena, P.K. An efficient temporary immersion system for micropropagation of hybrid hazelnut. Botany 2016, 94, 1–8. [Google Scholar] [CrossRef]
- Nicholson, J.; Shukla, M.R.; Saxena, P.K. In vitro rooting of hybrid hazelnuts (Corylus avellana × Corylus americana) in a temporary immersion system. Botany 2020, 98, 343–352. [Google Scholar] [CrossRef]
- Troch, V.; Sapeta, H.; Werbrouck, S.; Geelen, D.; Van Labeke, M.-C. In vitro culture of chestnut (Castanea sativa Mill.) using temporary immersion bioreactors. Acta Hortic. 2010, 885, 383–389. [Google Scholar] [CrossRef]
- Vidal, N.; Blanco, B.; Cuenca, B. A Temporary immersion system for micropropagation of axillary shoots of hybrid chestnut. Plant Cell, Tissue Organ Cult. 2015, 123, 229–243. [Google Scholar] [CrossRef]
- Vidal, N.; Correa, B.; Rial, E.; Regueira, M.; Sánchez, C.; Cuenca, B. Comparison of temporary and continuous immersion systems for micropropagation of axillary shoots of chestnut and willow. Acta Hortic. 2015, 1083, 227–233. [Google Scholar] [CrossRef]
- Engelmann, F. Plant cryopreservation: Progress and prospects. Vitr. Cell. Dev. Biol. Plant 2004, 40, 427–433. [Google Scholar] [CrossRef]
- Benson, E.E. Cryopreservation of phytodiversity: A critical appraisal of theory & practice. Crit. Rev. Plant Sci. 2008, 27, 141–219. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, Z.; Yin, Z.; Feng, C.; Wang, Q. Novel and potential application of cryopreservation to plant genetic transformation. Biotechnol. Adv. 2012, 30, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Harding, K. Genetic Integrity of Cryopreserved Plant Cells: A Review. CryoLetters 2004, 25, 3–22. [Google Scholar] [PubMed]
- Kim, H.-H.; Cho, E.-G.; Baek, H.-J.; Kim, C.-Y.; Keller, E.R.J.; Engelmann, F. Cryopreservation of garlic shoot tips by vitrification: Effects of dehydration, rewarming, unloading and regrowth conditions. CryoLetters 2004, 25, 59–70. [Google Scholar] [PubMed]
- Jorquera, L.; Vidal, N.; Sánchez, C.; Vieitez, A.M. optimizing conditions for successful plant regeneration from cryopreserved Castanea sativa shoot tips. Acta Hortic. 2005, 693, 511–518. [Google Scholar] [CrossRef]
- San-José, M.C.; Jorquera, L.; Vidal, N.; Corredoira, E.; Sánchez, C. Cryopreservation of European chestnut germplasm. Acta Hortic. 2005, 693, 225–232. [Google Scholar] [CrossRef]
- Vidal, N.; Sánchez, C.; Jorquera, L.; Ballester, A.; Vieitez, A.M. Cryopreservation of chestnut by vitrification of in vitro-grown shoot tips. Vitr. Cell. Dev. Biol. Plant 2005, 41, 63–68. [Google Scholar] [CrossRef]
- Sakai, A.; Engelmann, F. Vitrification, encapsulation-vitrification and droplet-vitrification: A review. CryoLetters 2007, 28, 151–172. [Google Scholar] [PubMed]
- Halmagyi, A.; Deliu, C.; Isac, V. Cryopreservation of Malus cultivars: Comparison of two droplet protocols. Sci. Hortic 2010, 124, 387–392. [Google Scholar] [CrossRef]
- Folgado, R.; Panis, B. Cryopreservation of Ashe magnolia shoot-tips by droplet-vitrification. Acta Hortic. 2019, 1234, 233–240. [Google Scholar] [CrossRef]
- Sant, R.; Panis, B.; Taylor, M.; Tyagi, A. Cryopreservation of shoot-tips by droplet vitrification applicable to all taro (Colocasia esculenta var. esculenta) accessions. Plant Cell Tissue Organ Cult. 2008, 92, 107–111. [Google Scholar] [CrossRef]
- Park, S.-U.; Kim, H.-H. Cryopreservation of sweet potato shoot tips using a droplet-vitrification procedure. CryoLetters 2015, 36, 344–352. [Google Scholar]
- Gallard, A.; Panis, B.; Dorion, N.; Swennen, R.; Grapin, A. Cryopreservation of Pelargonium apices by droplet-vitrification. CryoLetters 2008, 29, 243–251. [Google Scholar] [PubMed]
- Yin, Z.-F.; Bi, W.-L.; Chen, L.; Zhao, B.; Volk, G.M.; Wang, Q.-C. An efficient, widely applicable cryopreservation of Lilium shoot tips by droplet vitrification. Acta Physiol. Plant. 2014, 36, 1683–1692. [Google Scholar] [CrossRef]
- Zhang, J.-M.; Huang, B.; Zhang, X.-N.; Volk, G.M.; Zhou, Y.-C.; Chen, X.-L. Identification of a highly successful cryopreservation method (droplet-vitrification) for petunia. Vitr. Cell. Dev. Biol. Plant 2015, 51, 445–451. [Google Scholar] [CrossRef]
- Rathwell, R.; Shukla, M.R.; Jones, A.M.P.; Saxena, P.K. In vitro propagation of cherry birch (Betula lenta L.). Can. J. Plant Sci. 2016, 96, 571–578. [Google Scholar] [CrossRef] [Green Version]
- Salama, A.; Popova, E.; Jones, M.P.; Shukla, M.R.; Fisk, N.S.; Saxena, P.K. cryopreservation of the critically endangered golden paintbrush (Castilleja levisecta Greenm.): From nature to cryobank to nature. Vitr. Cell. Dev. Biol. Plant 2018, 54, 69–78. [Google Scholar] [CrossRef]
- Sheikholeslami, B.; Shukla, M.; Turi, C.; Harpur, C.; Saxena, P.K. Saving threatened plant species: Reintroduction of hill’s thistle (Cirsium hillii. (Canby) Fernald) to its natural habitat. PLoS ONE 2020, 15, e0231741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popova, E.V.; Shukla, M.R.; McIntosh, T.; Saxena, P.K. In vitro and cryobiotechnology approaches to safeguard Lupinus rivularis Douglas ex Lindl., an endangered plant in Canada. Agronomy 2021, 11, 37. [Google Scholar] [CrossRef]
- Yang, X.; Popova, E.; Shukla, M.R.; Saxena, P.K. Root cryopreservation to biobank medicinal plants: A case study for Hypericum perforatum L. Vitr. Cell. Dev. Biol. Plant 2019, 55, 392–402. [Google Scholar] [CrossRef]
- Identifying Your Chestnut Tree. Available online: https://acf.org/ma-ri/the-project/tree-identification (accessed on 15 January 2022).
- Barnhill-Dilling, S.K.; Rivers, L.; Delborne, J.A. Rooted in recognition: Indigenous environmental justice and the genetically engineered American chestnut tree. Soc. Nat. Resour. 2020, 33, 83–100. [Google Scholar] [CrossRef]
- Carvalho, L.S.O.; Ozudogru, E.A.; Lambardi, M.; Paiva, L.V. Temporary immersion system for micropropagation of tree species: A Bibliographic and Systematic Review. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 47, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Welander, M.; Persson, J.; Asp, H.; Zhu, L.H. Evaluation of a new vessel system based on temporary immersion system for micropropagation. Sci. Hortic 2014, 179, 227–232. [Google Scholar] [CrossRef]
- Businge, E.; Trifonova, A.; Schneider, C.; Rödel, P.; Egertsdotter, U. evaluation of a new temporary immersion bioreactor system for micropropagation of cultivars of eucalyptus, birch and fir. Forests 2017, 8, 196. [Google Scholar] [CrossRef]
- Shukla, M.R.; Singh, A.S.; Piunno, K.; Saxena, P.K.; Jones, A.M.P. Application of 3D printing to prototype and develop novel plant tissue culture systems. Plant Methods 2017, 13, 6. [Google Scholar] [CrossRef] [Green Version]
- Capuana, M.; Di Lonardo, S. In vitro Conservation of chestnut (Castanea sativa) by slow growth. Vitr. Cell. Dev. Biol. Plant 2013, 49, 605–610. [Google Scholar] [CrossRef]
- Tafazoli, M.; Nasr, S.M.H.; Jalilvand, H.; Bayat, D. Plant regeneration through indirect organogenesis of chestnut (Castanea sativa Mill.). Afr. J. Biotechnol. 2013, 12, 7063–7069. [Google Scholar]
- Vieitez, A.M.; Vieitez, M.L. Castanea sativa plantlets proliferated from axillary buds cultivated in vitro. Sci. Hortic. 1983, 18, 343–351. [Google Scholar] [CrossRef]
- Roussos, P.A.; Archimandriti, A.; Beldekou, I. Improving in vitro multiplication of juvenile european chestnut (Castanea sativa Mill) explants by the use of growth retardants. Sci. Hortic. 2016, 198, 254–256. [Google Scholar] [CrossRef]
- San-Jose, M.C.; Ballester, A.; Vieite, A. Effect of thidiazuron on multiple shoot induction and plant regeneration from cotyledonary nodes of chestnut. J. Hortic. Sci. Biotechnol. 2001, 76, 588–595. [Google Scholar] [CrossRef]
- Soylu, A.; Ertürk, Ü. Researches on micropropagation of chestnut. Acta Hortic. 1999, 494, 247–254. [Google Scholar] [CrossRef]
- Tetsumura, T.; Yamashita, K. Micropropagation of japanese chestnut (Castanea crenata Sieb. et Zucc.) Seedlings. HortSci. 2004, 39, 1684–1687. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Guo, S.; Wang, G. Effects of in vitro subculture on the physiological characteristics of adventitious root formation in microshoots of Castanea mollissima cv. ‘Yanshanhong.’ J. For. Res. 2010, 21, 155–160. [Google Scholar] [CrossRef]
- Garoosi, G.; Delijam, M.; Nezami-Alanagh, E.; Hosseini, R. Improving Pistacia vera Micropropagation: With emphasis on the efficiency of minerals, vitamins and plant growth regulators. J. Plant Mol. Breed. 2016, 4, 43–54. [Google Scholar] [CrossRef]
- George, E.F.; Hall, M.A.; de Klerk, G.-J. Plant Propagation by Tissue Culture, 3rd ed.; Springer: Dordrecht, The Netherlands, 2008; ISBN 978-1-4020-5004-6. [Google Scholar]
- Fernández-Lorenzo, J.L.; Ballester, A.; Rigueiro, A. Phenolic content of microcuttings of adult chestnut along rooting induction. Plant Cell Tissue Organ Cult. 2005, 83, 153–159. [Google Scholar] [CrossRef]
- Gonçalves, J.C.; Diogo, G.; Coelho, M.T.; Amâncio, S. Effect of rooting conditions on survival and growth during acclimatization of micropropagated chestnut plants (Castanea sativa × C. crenata). Acta Hortic. 1999, 494, 235–242. [Google Scholar] [CrossRef]
- Sato, T. Basic studies of organ and callus culture in woody plants. Bull. For. For. Prod. Res. Inst. Jpn. 1990, 360, 35–119. [Google Scholar]
- Davies, P.J. The Plant Hormones: Their Nature, Occurrence, and Functions; Davies, P.J., Ed.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013; ISBN 978-94-011-0473-9. [Google Scholar]
- Mert, C.; Soylu, A. Shoot apex culture of dormant buds in chestnut. Acta Hortic. 2009, 844, 145–152. [Google Scholar] [CrossRef]
- Algül, B.E.; Dalkılıç, G.G. Micropropagation of chestnut (Castanea sativa Mill.) in semi-solid culture. Acta Hortic. 2014, 1043, 205–209. [Google Scholar] [CrossRef]
- Chauvin, J.E.; Salesses, G. Advances in chestnut micropropagation (Castanea sp.). Acta Hortic. 1988, 227, 340–345. [Google Scholar] [CrossRef]
- Sanchez, M.C.; San-Jose, M.C.; Ferro, E.; Ballester, A.; Vieitez, A.M. Improving micropropagation conditions for adult-phase shoots of chestnut. J. Hortic. Sci. Biotechnol. 1997, 72, 433–443. [Google Scholar] [CrossRef]
- Ballester, A.; Bourrain, L.; Corredoira, E.; Gonçalves, J.C.; Lê, C.; Miranda -Fontaíña, M.E.; San-José, M.D.C.; Sauer, U.; Vieitez, A.M.; Wilhelm, E. Improving chestnut micropropagation through axillary shoot development and somatic embryogenesis. For. Snow Landsc. Res 2001, 76, 460–467. [Google Scholar]
- Vieitez, A.M.; Sänchez, M.C.; García-Nimo, M.L.; Ballester, A. Protocol for micropropagation of Castanea sativa. In Protocols for Micropropagation of Woody Trees and Fruits; Jain, S.M., Häggman, H., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 229–312. ISBN 978-1-4020-6352-7. [Google Scholar]
- Giovannelli, A.; Giannini, R. Reinvigoration of mature chestnut (Castanea sativa) by repeated graftings and micropropagation. Tree Physiol. 2000, 20, 1243–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, J.C.; Diogo, G.; Amâncio, S. In vitro propagation of chestnut (Castanea sativa × C. crenata): Effects of rooting treatments on plant survival, peroxidase activity and anatomical changes during adventitious root formation. Sci. Hortic. 1998, 72, 265–275. [Google Scholar] [CrossRef]
- Gupta, S.D.; Prasad, V.S.S. Matrix-supported liquid culture systems for efficient micropropagation of floricultural plants. In Floriculture, ornamental and plant biotechnology: Advances and topical issues, volume II; Global Science Books: East Sussex, UK, 2006; pp. 487–495. [Google Scholar]
- Naylor Adelberg, J.A.; Adelberg, J.W.; Rapaka, V.K. A Rooting matrix and tissue culture vessel system yielded larger, faster growing Echeveria plants. Acta Hortic. 2016, 1113, 53–58. [Google Scholar] [CrossRef]
- Hazarika, B.N. Acclimatization of tissue-cultured plants. Curr. Sci. 2003, 85, 1704–1712. [Google Scholar]
- Panis, B.; Piette, B.; Swennen, R. Droplet vitrification of apical meristems: A cryopreservation protocol applicable to all Musaceae. Plant Sci. 2005, 168, 45–55. [Google Scholar] [CrossRef]
- Halmagyi, A.; Pinker, I. Plant regeneration from Rosa shoot tips cryopreserved by a combined droplet vitrification method. Plant Cell Tissue Organ Cult. 2006, 84, 145–153. [Google Scholar] [CrossRef]
- Condello, E.; Caboni, E.; Andrè, E.; Piette, B.; Druart, P.; Swennen, R.; Panis, B. Cryopreservation of apple in vitro axillary buds using droplet-vitrification. CryoLetters 2011, 32, 175–185. [Google Scholar]
- Vujović, T.; Sylvestre, I.; Ružić, D.; Engelmann, F. Droplet-Vitrification of apical shoot tips of Rubus fruticosus L. and Prunus cerasifera Ehrh. Sci. Hortic. 2011, 130, 222–228. [Google Scholar] [CrossRef]
- Wang, L.-Y.; Li, Y.-D.; Sun, H.-Y.; Liu, H.-G.; Tang, X.-D.; Wang, Q.-C.; Zhang, Z.-D. An Efficient droplet-vitrification cryopreservation for valuable blueberry germplasm. Sci. Hortic. 2017, 219, 60–69. [Google Scholar] [CrossRef]
- Wang, M.-R.; Zhang, Z.; Haugslien, S.; Sivertsen, A.; Rasmussen, M.; Wang, Q.-C.; Blystad, D.-R. Cryopreservation of shallot (Allium cepa var. aggregatum) shoot tips by droplet-vitrification. Acta Hortic. 2019, 1234, 241–248. [Google Scholar] [CrossRef]
- Vidal, N.; Vieitez, A.M.; Fernández, M.R.; Cuenca, B.; Ballester, A. Establishment of cryopreserved gene banks of European chestnut and cork oak. Eur. J. For. Res. 2010, 129, 635–643. [Google Scholar] [CrossRef]
- Touchell, D.; Dixon, K.; Tan, B. Cryopreservation of shoot-tips of Grevillea scapigera (Proteaceae): A rare and endangered plant from western Australia. Aust. J. Bot. 1992, 40, 305–310. [Google Scholar] [CrossRef]
- Vandenbussche, B.; Weyens, G.; De Proft, M. Cryopreservation of in vitro sugar beet (Beta vulgaris L.) shoot tips by a vitrification technique. Plant Cell Rep. 2000, 19, 1064–1068. [Google Scholar] [CrossRef]
- Verleysen, H.; Fernandes, P.; Pinto, I.S.; Bockstaele, E.V.; Debergh, P. Cryopreservation of Robinia pseudoacacia. Plant Cell Tissue Organ Cult. 2005, 81, 193–202. [Google Scholar] [CrossRef]
- Yoon, J.-W.; Kim, H.-H.; Ko, H.-C.; Hwang, H.-S.; Hong, E.-S.; Cho, E.-G.; Engelmann, F. Cryopreservation of cultivated and wild potato varieties by droplet vitrification: Effect of subculture of mother-plants and of preculture of shoot tips. CryoLetters 2006, 27, 211–222. [Google Scholar]
- Pathirana, R.; McLachlan, A.; Hedderley, D.; Panis, B.; Carimi, F. Pre-treatment with salicylic acid improves plant regeneration after cryopreservation of grapevine (Vitis spp.) by droplet vitrification. Acta Physiol. Plant. 2015, 38, 12. [Google Scholar] [CrossRef]
- Sakai, A.; Hirai, D.; Niino, T. Development of pvs-based vitrification and encapsulation–vitrification protocols. In Plant Cryopreservation: A Practical Guide; Reed, B.M., Ed.; Springer New York: New York, NY, USA, 2008; pp. 33–57. ISBN 978-0-387-72276-4. [Google Scholar]
- Nishizawa, S.; Sakai, A.; Amano, Y.; Matsuzawa, T. Cryopreservation of asparagus (Asparagus officinalis L.) Embryogenic suspension cells and subsequent plant regeneration by vitrification. Plant Sci. 1993, 91, 67–73. [Google Scholar] [CrossRef]
- Matsumoto, T.; Sakai, A.; Yamada, K. Cryopreservation of in vitro-grown apical meristems of wasabi (Wasabia japonica) by vitrification and subsequent high plant regeneration. Plant Cell Rep. 1994, 13, 442–446. [Google Scholar] [CrossRef]
- Volk, G.M.; Walters, C. Plant vitrification solution 2 lowers water content and alters freezing behavior in shoot tips during cryoprotection. Cryobiology 2006, 52, 48–61. [Google Scholar] [CrossRef]
- Sakai, A.; Kobayashi, S.; Oiyama, I. Survival by vitrification of nucellar cells of navel orange (Citrus sinensis var. Brasiliensis Tanaka) cooled to -196 °C. J. Plant Physiol. 1991, 137, 465–470. [Google Scholar] [CrossRef]
- Wu, Y.; Zhao, Y.; Engelmann, F.; Zhou, M.; Zhang, D.; Chen, S. Cryopreservation of apple dormant buds and shoot tips. CryoLetters 2001, 22, 375–380. [Google Scholar] [PubMed]
- Martà nez-Montero, M.E.; Martà nez, J.; Engelmann, F.; Gonzalez-Arnao, M.T. Cryopreservation of pineapple (Ananas comosus (L.) Merr) apices and calluses. Acta Hortic. 2005, 666, 127–131. [Google Scholar] [CrossRef]
- Kim, H.-H.; Lee, Y.-G.; Shin, D.-J.; Ko, H.-C.; Gwag, J.-G.; Cho, E.-G.; Engelmann, F. Development of alternative plant vitrification solutions in droplet-vitrification procedures. CryoLetters 2009, 30, 320–334. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, P.; Batuman, Ö.; Gafny, R.; Mawassi, M. Effect of benzyladenine on recovery of cryopreserved shoot tips of grapevine and citrus cultured in vitro. CryoLetters 2003, 24, 293–302. [Google Scholar] [PubMed]
- Gresshoff, P.M.; Doy, C.H. Development and differentiation of haploid Lycopersicon esculentum (tomato). Planta 1972, 107, 161–170. [Google Scholar] [CrossRef]
- Menon, A.; Funnekotter, B.; Kaczmarczyk, A.; Bunn, E.; Turner, S.; Mancera, R.L. Cryopreservation of Lomandra sonderi (Asparagaceae) shoot tips using droplet-vitrification. CryoLetters 2012, 33, 259–270. [Google Scholar]
- Kim, H.-H.; Popova, E.V.; Shin, D.-J.; Bae, C.-H.; Baek, H.-J.; Park, S.-U.; Engelmann, F. Development of a droplet-vitrification protocol for cryopreservation of Rubia akane (Nakai) hairy roots using a systematic approach. CryoLetters 2012, 33, 506–517. [Google Scholar]
- Zhang, J.; Huang, B.; Lu, X.; Volk, G.M.; Xin, X.; Yin, G.; He, J.; Chen, X. Cryopreservation of in vitro-grown shoot tips of chinese medicinal plant Atractylodes macrocephala Koidz. using a droplet-vitrification method. CryoLetters 2015, 36, 195–204. [Google Scholar] [PubMed]
- Driver, J.A. In vitro Propagation of paradox walnut rootstock. HortScience 1984, 19, 507–709. [Google Scholar]
- Murashige, T.; Skoog, F. A Revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Lloyd, G. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. Comb. Proc. Int. Plant Prop. Soc. 1980, 30, 421–427. [Google Scholar]
- Sakai, A.; Kobayashi, S.; Oiyama, I. Cryopreservation of nucellar cells of navel orange (Citrus sinensis Osb. var. brasiliensis Tanaka) by vitrification. Plant Cell Rep. 1990, 9, 30–33. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Continuous Preculture | +LN (%) | −LN (%) |
|---|---|---|
| 0.5 M (24 h) | 00.0 ± 4.6 c | 86.7 ± 8.3 |
| 0.5 M (48 h) | 38.3 ± 4.6 ab | 80.0 ± 8.3 |
| 0.5 M (72 h) | 40.0 ± 4.6 a | 76.7 ± 8.3 |
| 0.5 M (96 h) | 20.0 ± 4.6 b | 70.0 ± 8.3 |
| Stepwise Preculture | ||
| 0.25–1.0 M (96 h) | 55.0 ± 4.6 a | 76.7 ± 8.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Bi, W.-L.; Shukla, M.R.; Saxena, P.K. In Vitro Technologies for American Chestnut (Castanea dentata (Marshall) Borkh) Conservation. Plants 2022, 11, 464. https://doi.org/10.3390/plants11030464
Liu Z, Bi W-L, Shukla MR, Saxena PK. In Vitro Technologies for American Chestnut (Castanea dentata (Marshall) Borkh) Conservation. Plants. 2022; 11(3):464. https://doi.org/10.3390/plants11030464
Chicago/Turabian StyleLiu, Zhuoya, Wen-Lu Bi, Mukund R. Shukla, and Praveen K. Saxena. 2022. "In Vitro Technologies for American Chestnut (Castanea dentata (Marshall) Borkh) Conservation" Plants 11, no. 3: 464. https://doi.org/10.3390/plants11030464