
Isolation of the Novel Strain Bacillus amyloliquefaciens F9 and Identification of Lipopeptide Extract Components Responsible for Activity against Xanthomonas citri subsp. citri


Abstract
:1. Introduction
2. Results
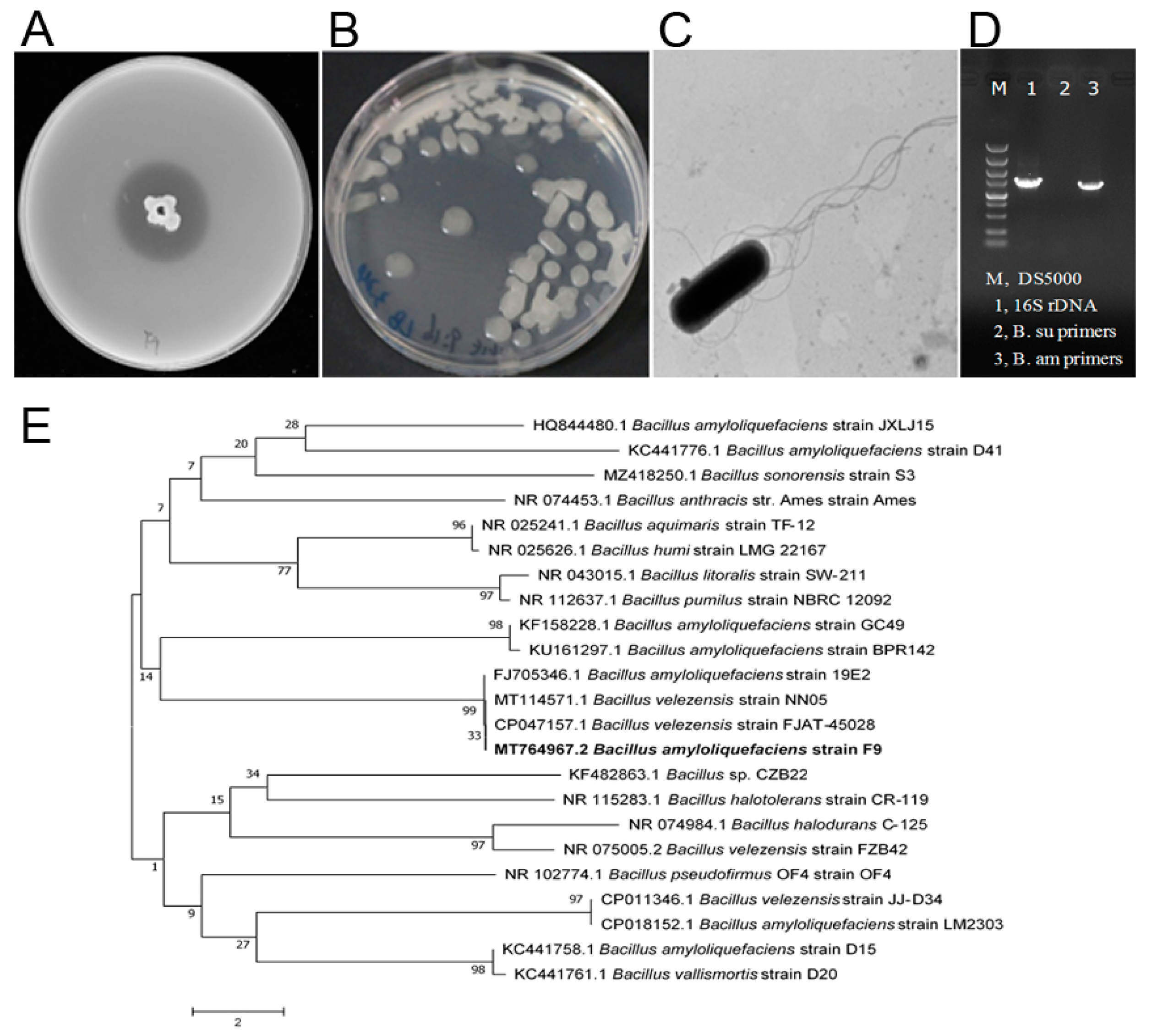
2.1. Isolation and Screening of Antagonistic Bacteria of Xcc
2.2. Characterization of a New Antagonistic Strain Bacillus amyloliquefaciens F9 against Xcc
2.3. Biochemical Characteristics of Strain F9 Lipopeptide Extract (F9LE)
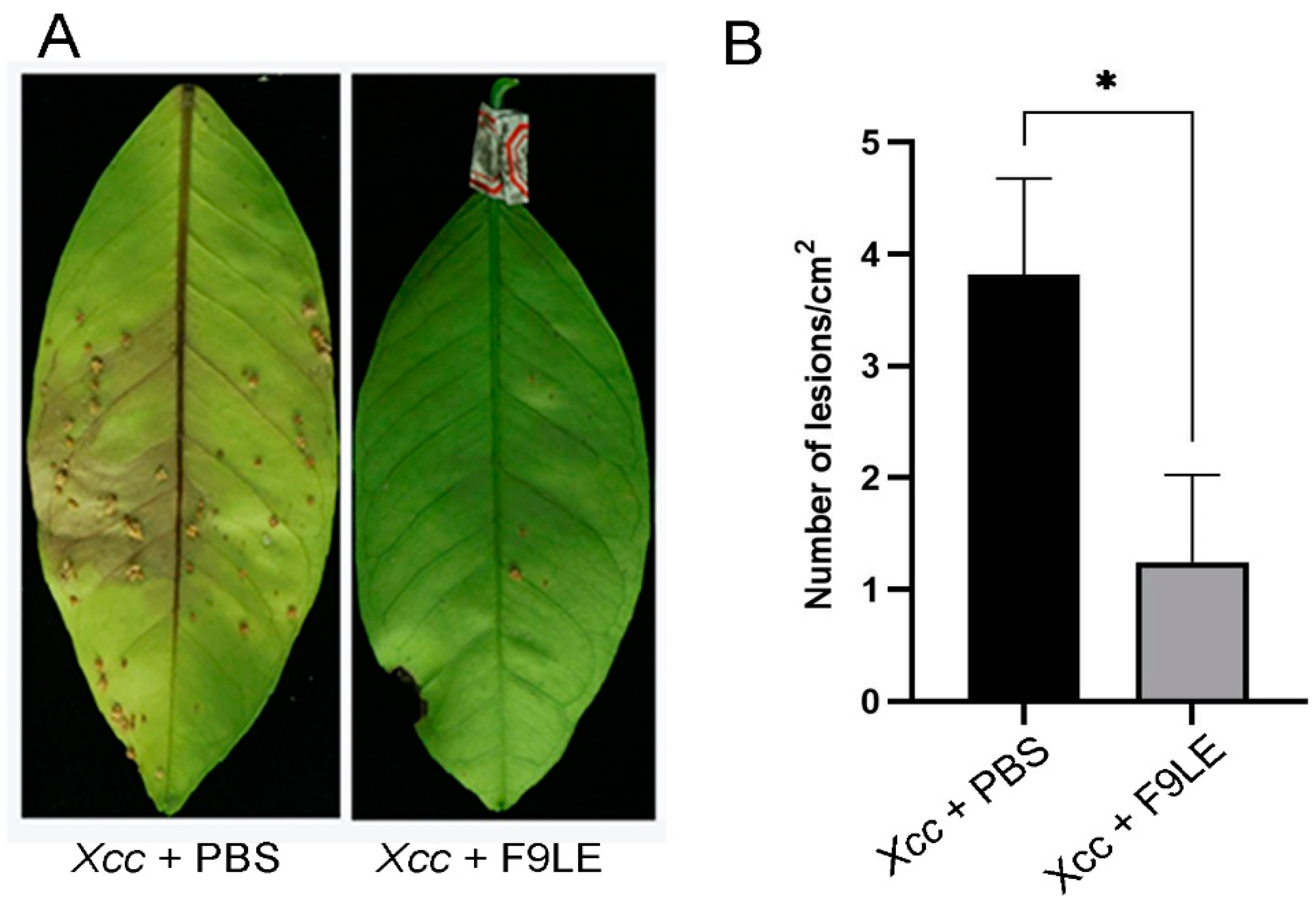
2.4. Biological Control Efficiency of F9LE on Xcc jx-6 under Greenhouse Conditions
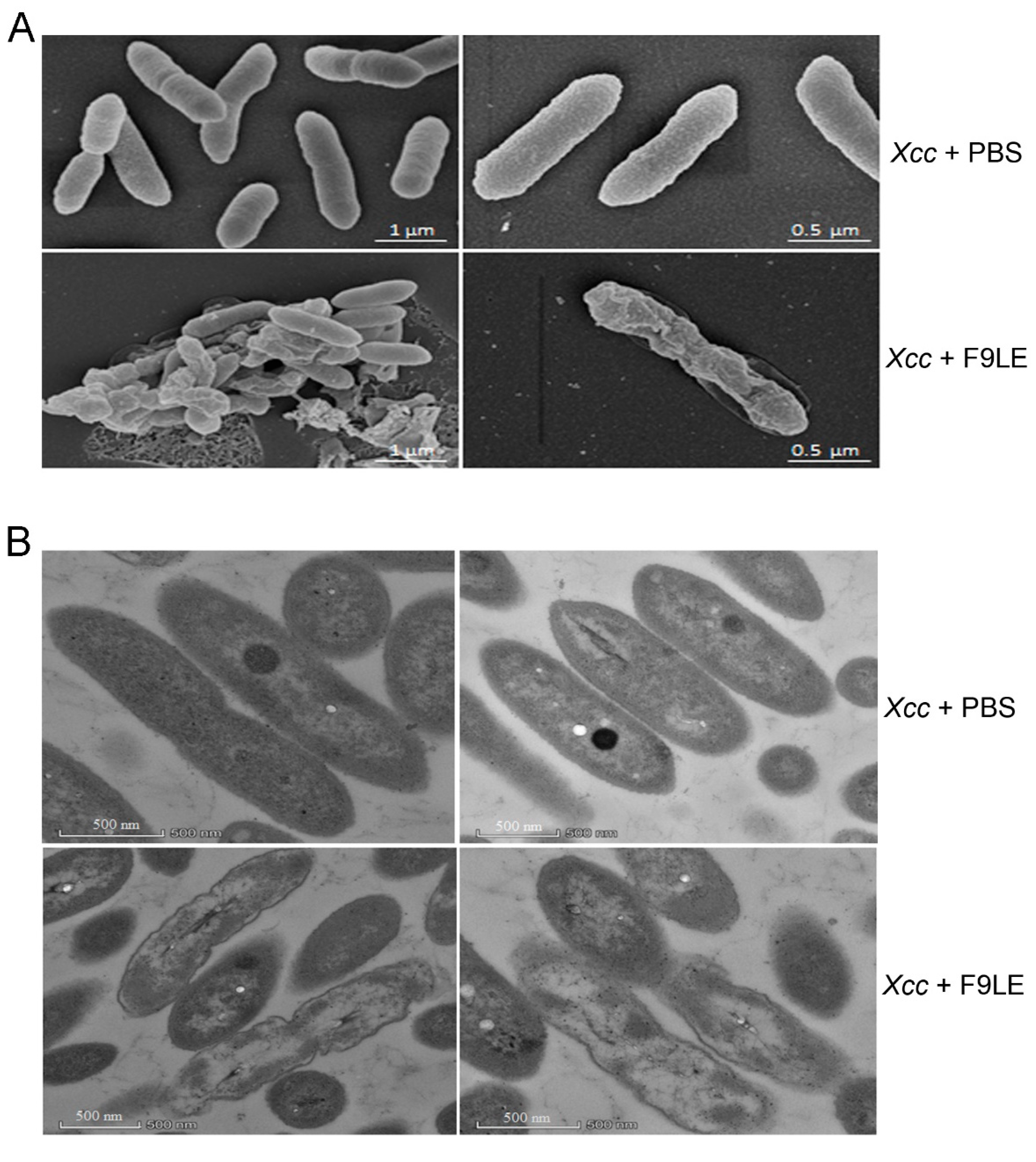
2.5. The Effect of F9LE on the Morphology and Ultrastructure of Xcc jx-6 Cells
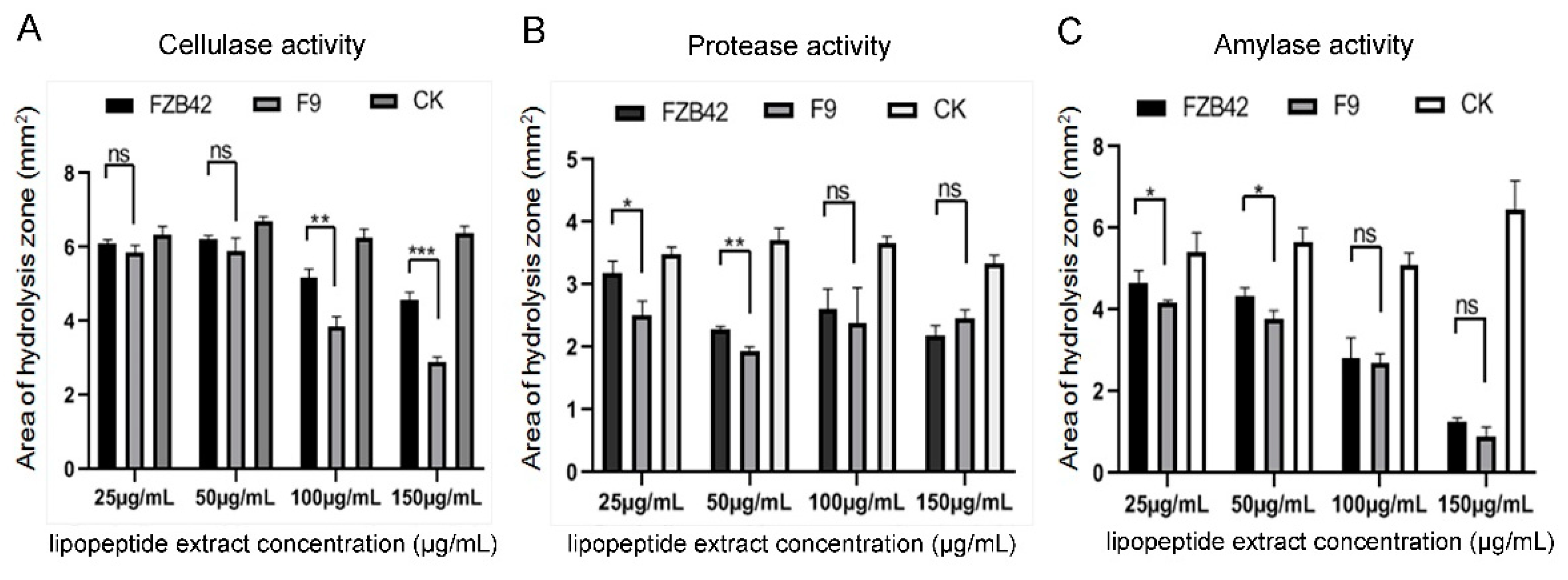
2.6. Inhibitory Effect of F9LE on Extracellular Enzyme Activity of Xcc jx-6
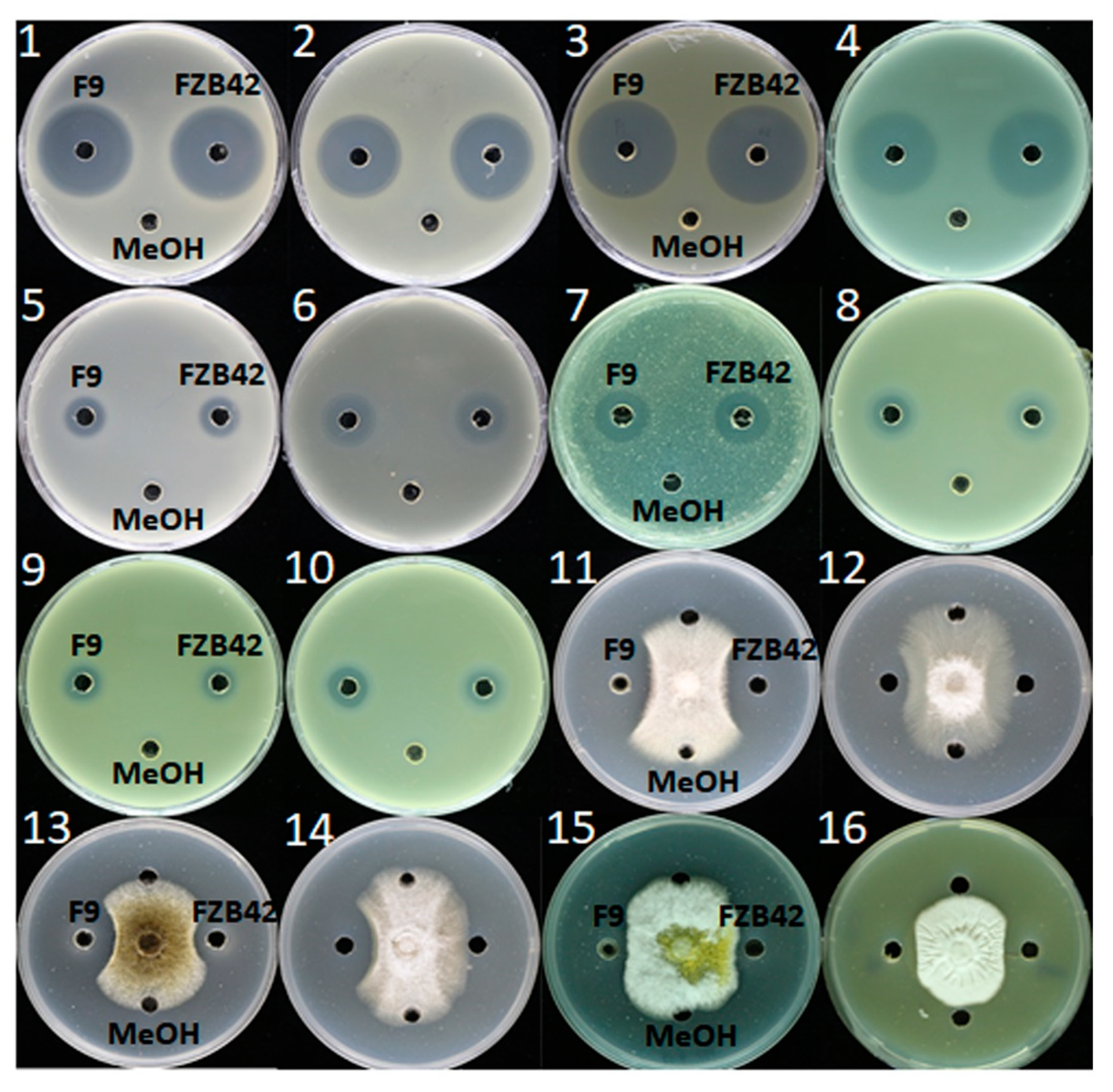
2.7. Determination of Antimicrobial Spectrum of F9LE
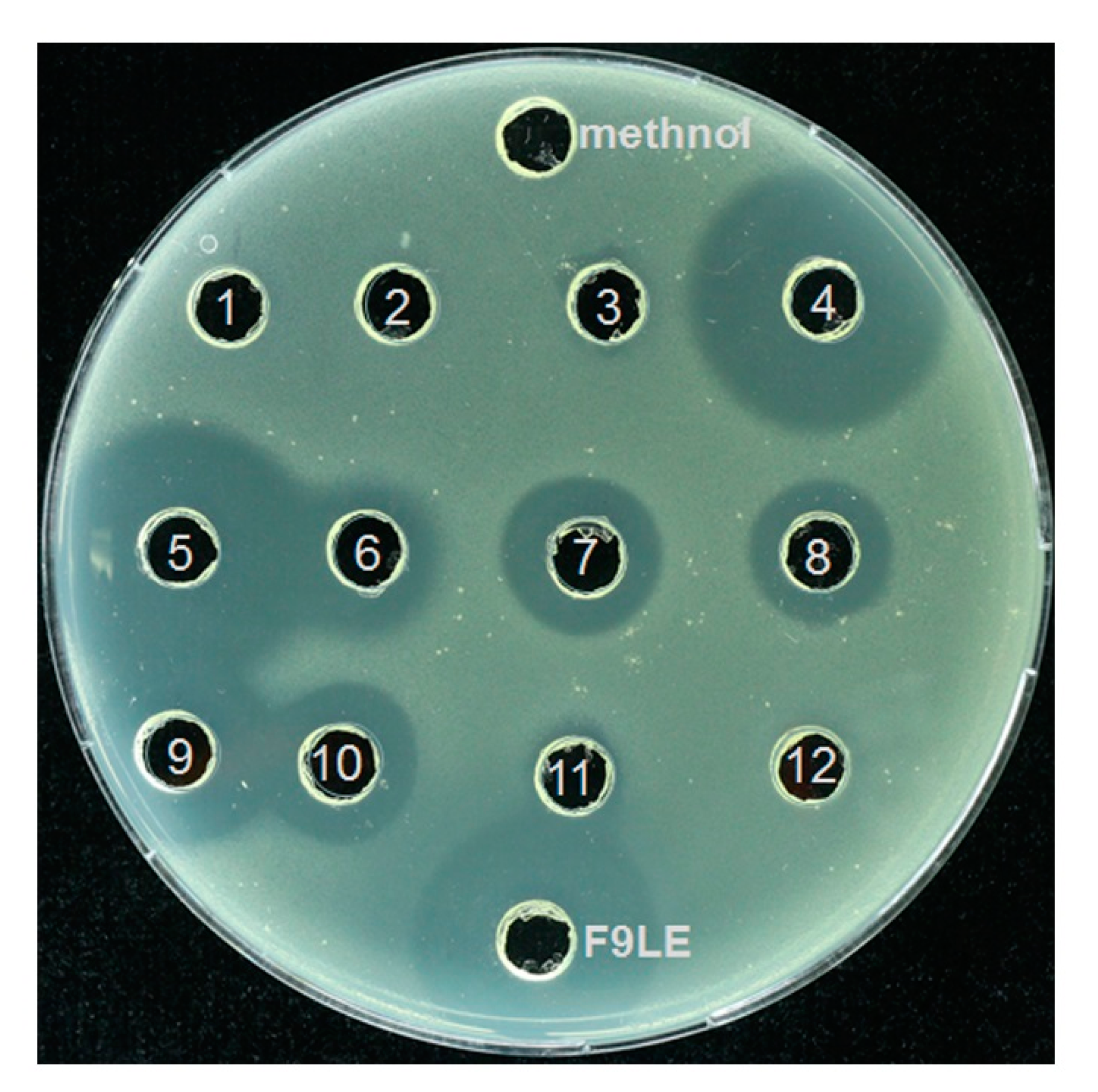
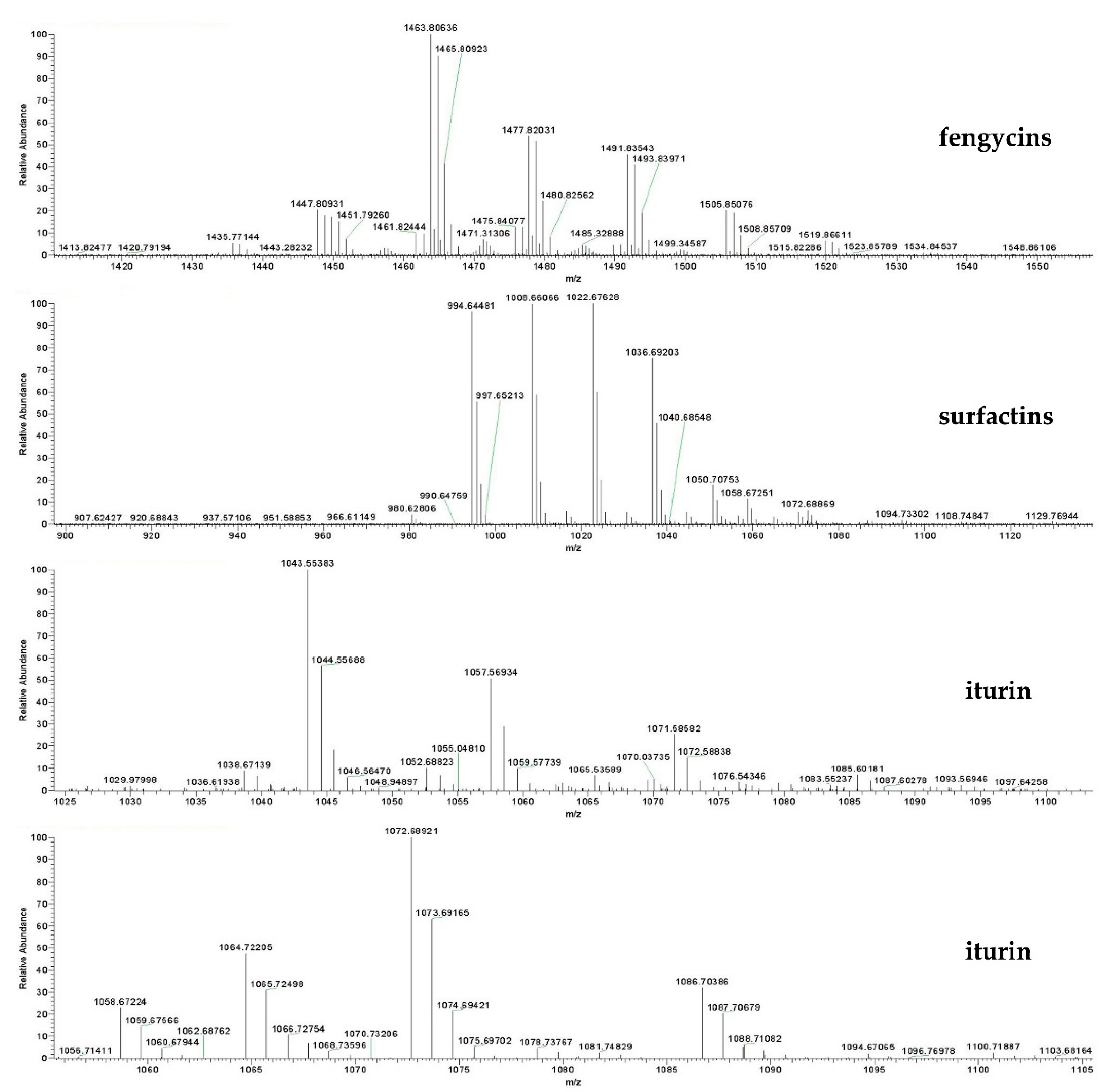
2.8. Identification of the Bioactive Compounds of F9LE by HPLC and LC–ESI–MS
3. Discussion
4. Materials and Methods
4.1. Pathogenic Strains and Cultural Conditions
4.2. Soil Sample Collection from Citrus Rhizosphere Soil
4.3. Isolation of Antagonistic Bacteria of Xcc
4.4. Characterization of the New Antagonistic Isolate B. amyloliquefaciens F9
4.5. Determination of Antibacterial Activity by Agar Diffusion Assay
4.6. Analysis of Antibacterial Stability of Culture Supernatant
4.7. Preparation of Crude Lipopeptide Extract and Its Stability Analysis
4.8. Pathogenicity Assays under Greenhouse Conditions
4.9. Determination of Extracellular Enzymes’ Activities
4.10. Scanning Electron Microscopy (SEM) and Transmission Electron Microscopy (TEM) Studies
4.11. Determination of Antimicrobial Spectrum of F9LE
4.12. HPLC Purification of Putative Bioactive Compounds
4.13. LC–ESI–MS Analysis of Bioactive Compounds of F9LE
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- An, S.Q.; Potnis, N.; Dow, M.; Vorholter, F.J.; He, Y.Q.; Becker, A.; Teper, D.; Li, Y.; Wang, N.; Bleris, L.; et al. Mechanistic insights into host adaptation, virulence and epidemiology of the phytopathogen Xanthomonas. FEMS Microbiol. Rev. 2020, 44, 1–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalan, N.; Kumar, D.; Yu, F.; Jones, J.B.; Graham, J.H.; Wang, N. Complete Genome Sequence of Xanthomonas citri subsp. citri Strain Aw12879, a Restricted-Host-Range Citrus Canker-Causing Bacterium. Genome Announc. 2013, 1, e00235-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, A.K. Citrus canker: A review. J. Appl. Hortic. 2003, 5, 9. [Google Scholar] [CrossRef]
- Gottwald, T.R.; Sun, X.; Riley, T.; Graham, J.H.; Ferrandino, F.; Taylor, E.L. Geo-referenced spatiotemporal analysis of the urban citrus canker epidemic in Florida. Phytopathology 2002, 92, 361–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.N.; Ali, M.S.; Choi, S.J.; Hyun, J.W.; Baek, K.H. Biocontrol of Citrus Canker Disease Caused by Xanthomonas citri subsp. citri Using an Endophytic Bacillus thuringiensis. Plant Pathol. J. 2019, 35, 486–497. [Google Scholar] [CrossRef]
- Behlau, F.; Canteros, B.I.; Minsavage, G.V.; Jones, J.B.; Graham, J.H. Molecular characterization of copper resistance genes from Xanthomonas citri subsp. citri and Xanthomonas alfalfae subsp. citrumelonis. Appl. Environ. Microbiol. 2011, 77, 4089–4096. [Google Scholar] [CrossRef] [Green Version]
- Graham, J.H.; Leite, R.P.; Yonce, H.D.; Myers, M. Streptomycin controls citrus canker on sweet orange in Brazil and reduces risk of copper burn on grapefruit in Florida. Proc. Fl. State Hortic. Soc. 2008, 121, 6. [Google Scholar]
- Hyun, J.W.; Kim, H.J.; Yi, P.H.; Hwang, R.Y.; Park, E.W. Mode of Action of Streptomycin Resistance in the Citrus Canker Pathogen (Xanthomonas smithii subsp. citri) in Jeju Island. Plant Pathol. J. 2012, 28, 5. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, D.B.; Tounsi, S.; Gharsallah, H.; Hammami, A.; Frikha-Gargouri, O. Lipopeptides from Bacillus amyloliquefaciens strain 32a as promising biocontrol compounds against the plant pathogen Agrobacterium tumefaciens. Environ. Sci. Pollut. Res. Int. 2018, 25, 36518–36529. [Google Scholar] [CrossRef]
- Cao, Y.; Pi, H.; Chandrangsu, P.; Li, Y.; Wang, Y.; Zhou, H.; Xiong, H.; Helmann, J.D.; Cai, Y. Antagonism of Two Plant-Growth Promoting Bacillus velezensis Isolates Against Ralstonia solanacearum and Fusarium oxysporum. Sci. Rep. 2018, 8, 4360. [Google Scholar] [CrossRef]
- Hanif, A.; Zhang, F.; Li, P.; Li, C.; Xu, Y.; Zubair, M.; Zhang, M.; Jia, D.; Zhao, X.; Liang, J.; et al. Fengycin Produced by Bacillus amyloliquefaciens FZB42 Inhibits Fusarium graminearum Growth and Mycotoxins Biosynthesis. Toxins 2019, 11, 295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poveda, J.; Roeschlin, R.A.; Marano, M.R.; Favaro, M.A. Microorganisms as biocontrol agents against bacterial citrus diseases. Biol. Control. 2021, 158, 104602. [Google Scholar] [CrossRef]
- McSpadden Gardener, B.B.; Driks, A. Overview of the Nature and Application of Biocontrol Microbes: Bacillus spp. Phytopathology 2004, 94, 1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ongena, M.; Jourdan, E.; Adam, A.; Paquot, M.; Brans, A.; Joris, B.; Arpigny, J.L.; Thonart, P. Surfactin and fengycin lipopeptides of Bacillus subtilis as elicitors of induced systemic resistance in plants. Environ. Microbiol. 2007, 9, 1084–1090. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, J.D.; Baker, J.L.; Mahoney, N.E. Isolation of bacterial antagonists of Aspergillus flavus from almonds. Microb. Ecol. 2006, 52, 45–52. [Google Scholar] [CrossRef]
- Chen, X.H.; Scholz, R.; Borriss, M.; Junge, H.; Mogel, G.; Kunz, S.; Borriss, R. Difficidin and bacilysin produced by plant-associated Bacillus amyloliquefaciens are efficient in controlling fire blight disease. J. Biotechnol. 2009, 140, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Zeriouh, H.; Romero, D.; Garcia-Gutierrez, L.; Cazorla, F.M.; de Vicente, A.; Perez-Garcia, A. The iturin-like lipopeptides are essential components in the biological control arsenal of Bacillus subtilis against bacterial diseases of cucurbits. Mol. Plant Microbe Interact. 2011, 24, 1540–1552. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Wu, H.; Chen, L.; Yu, X.; Borriss, R.; Gao, X. Difficidin and bacilysin from Bacillus amyloliquefaciens FZB42 have antibacterial activity against Xanthomonas oryzae rice pathogens. Sci. Rep. 2015, 5, 12975. [Google Scholar] [CrossRef]
- Poveda, J.; Gonzalez-Andres, F. Bacillus as a source of phytohormones for use in agriculture. Appl. Microbiol. Biotechnol. 2021, 105, 8629–8645. [Google Scholar] [CrossRef]
- Ngalimat, M.S.; Yahaya, R.S.R.; Baharudin, M.M.A.; Yaminudin, S.M.; Karim, M.; Ahmad, S.A.; Sabri, S. A Review on the Biotechnological Applications of the Operational Group Bacillus amyloliquefaciens. Microorganisms 2021, 9, 614. [Google Scholar] [CrossRef]
- Preecha, C.; Sadowsky, M.J.; Prathuangwong, S. Lipopeptide Surfactin Produced by Bacillus amyloliquefaciens KPS46 is Required for Biocontrol Efficacy Against Xanthomonas axonopodis pv. glycines. Kasetsart J. 2010, 44, 16. [Google Scholar]
- Grady, E.N.; MacDonald, J.; Ho, M.T.; Weselowski, B.; McDowell, T.; Solomon, O.; Renaud, J.; Yuan, Z.C. Characterization and complete genome analysis of the surfactin-producing, plant-protecting bacterium Bacillus velezensis 9D-6. BMC Microbiol. 2019, 19, 5. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Shui-Lin, S.; Xiao-Li, L.; Ming-Xia, Y.; Ling-Ling, G.; University, J.A. Screening, identification of bio-control endophytic bacterium against citrus canker and stability of its bioactive metabolites. Acta Agric. Zhejiangensis 2015, 27, 2152–2156. [Google Scholar]
- Rabbee, M.F.; Baek, K.H. Endophyte bacillus velezensis isolated from citrus spp. controls streptomycin-resistant xanthomonas citri subsp. citri that causes citrus bacterial canker. Agronomy 2019, 9, 470. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.P.; Tzeng, D.D.; Wong, A.C.; Chen, C.H.; Lu, K.M.; Lee, Y.H.; Huang, W.D.; Hwang, B.F.; Tzeng, K.C. DNA polymorphisms and biocontrol of Bacillus antagonistic to citrus bacterial canker with indication of the interference of phyllosphere biofilms. PLoS ONE 2012, 7, e42124. [Google Scholar] [CrossRef] [PubMed]
- Das, R.; Mondal, B.; Mondal, P.; Khatua, D.C.; Mukherjee, N. Biological management of citrus canker on acid lime through Bacillus subtilis (S-12) in West Bengal, India. J. Biopest. 2014, 7, 38. [Google Scholar]
- Ibrahim, Y.E.; Saleh, A.A.; El_Komy, M.H.; Al-Saleh, M.A. Bacillus subtilis qst 713, copper hydroxide, and their tank mixes for control of bacterial citrus canker in saudi arabia. J. Citrus Pathol. 2016, 3, 30994. [Google Scholar] [CrossRef]
- Daungfu, O.; Youpensuk, S.; Lumyong, S. Endophytic Bacteria Isolated from Citrus Plants for Biological Control of Citrus Canker in Lime Plants. Trop. Life Sci. Res. 2019, 30, 73–88. [Google Scholar] [CrossRef]
- Wang, Y.; Li, L.; Li, B.; Wu, G.; Tang, Q.; Ibrahim, M.; Li, H.; Xie, G.; Sun, G. Action of chitosan against Xanthomonas pathogenic bacteria isolated from Euphorbia pulcherrima. Molecules 2012, 17, 7028–7041. [Google Scholar] [CrossRef]
- Tahir, H.A.; Gu, Q.; Wu, H.; Niu, Y.; Huo, R.; Gao, X. Bacillus volatiles adversely affect the physiology and ultra-structure of Ralstonia solanacearum and induce systemic resistance in tobacco against bacterial wilt. Sci. Rep. 2017, 7, 40481. [Google Scholar] [CrossRef]
- Penha, R.O.; Vandenberghe, L.; Faulds, C.; Soccol, V.T.; Soccol, C.R. Bacillus lipopeptides as powerful pest control agents for a more sustainable and healthy agriculture: Recent studies and innovations. Planta 2020, 251, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihalache, G.; Balaes, T.; Gostin, I.; Stefan, M.; Coutte, F.; Krier, F. Lipopeptides produced by Bacillus subtilis as new biocontrol products against fusariosis in ornamental plants. Environ. Sci. Pollut. Res. Int. 2018, 25, 29784–29793. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.C.; Wang, J.P.; Zhu, Y.J.; Liu, B.; Yang, W.J.; Ruan, C.Q. Antibacterial activity against Ralstonia solanacearum of the lipopeptides secreted from the Bacillus amyloliquefaciens strain FJAT-2349. J. Appl. Microbiol. 2019, 126, 1519–1529. [Google Scholar] [CrossRef] [PubMed]
- Sharga, B.M.; Lyon, G.D. Bacillus subtilis BS 107 as an antagonist of potato blackleg and soft rot bacteria. Can. J. Microbiol. 1998, 44, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Purwanto; Maria, G.M. The Role and Efficiency of Ammonium Sulphate Precipitation in Purification Process of Papain Crude Extract. Procedia Chem. 2016, 18, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.M.; Yoon, M.Y.; Choi, G.J.; Choi, Y.H.; Jang, K.S.; Shin, T.S.; Park, H.W.; Yu, N.H.; Kim, Y.H.; Kim, J.C. Diffusible and Volatile Antifungal Compounds Produced by an Antagonistic Bacillus velezensis G341 against Various Phytopathogenic Fungi. Plant Pathol. J. 2017, 33, 488–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.X.; Zhang, Y.; Shan, H.H.; Tong, Y.H.; Chen, X.J.; Liu, F.Q. Isolation and identification of antifungal peptides from Bacillus amyloliquefaciens W10. Environ. Sci. Pollut. Res. Int. 2017, 24, 25000–25009. [Google Scholar] [CrossRef]
- Radovanovic, N.; Milutinovic, M.; Mihajlovski, K.; Jovic, J.; Nastasijevic, B.; Rajilic-Stojanovic, M.; Dimitrijevic-Brankovic, S. Biocontrol and plant stimulating potential of novel strain Bacillus sp. PPM3 isolated from marine sediment. Microb. Pathog. 2018, 120, 71–78. [Google Scholar] [CrossRef]
- Song, S.; Fu, S.; Sun, X.; Li, P.; Wu, J.; Dong, T.; He, F.; Deng, Y. Identification of Cyclic Dipeptides from Escherichia coli as New Antimicrobial Agents against Ralstonia Solanacearum. Molecules 2018, 23, 214. [Google Scholar] [CrossRef] [Green Version]
- Redburn, A.C.; Patel, B.K. Phylogenetic analysis of Desulfotomaculum thermobenzoicum using polymerase chain reaction-amplified 16S rRNA-specific DNA. FEMS Microbiol. Lett. 1993, 113, 81–86. [Google Scholar] [CrossRef]
- Liu, Y.; Li, H.; Cheng, C. Cloning and Homology Analysis of Beta-mannanase Gene from the Collection of Bacillus subtilis Group. Genom. Appl. Biol. 2009, 28, 845–850. [Google Scholar]
- Zhang, Q.X.; Zhang, Y.; He, L.L.; Ji, Z.L.; Tong, Y.H. Identification of a small antimycotic peptide produced by Bacillus amyloliquefaciens 6256. Pestic. Biochem. Physiol. 2018, 150, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Perez, K.J.; Viana, J.D.; Lopes, F.C.; Pereira, J.Q.; Dos Santos, D.M.; Oliveira, J.S.; Velho, R.V.; Crispim, S.M.; Nicoli, J.R.; Brandelli, A.; et al. Bacillus spp. Isolated from Puba as a Source of Biosurfactants and Antimicrobial Lipopeptides. Front. Microbiol. 2017, 8, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Armstrong, C.M.; Zhou, M.; Duan, Y. Bismerthiazol Inhibits Xanthomonas citri subsp. citri Growth and Induces Differential Expression of Citrus Defense-Related Genes. Phytopathology 2016, 106, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.L.; Liu, Y.N.; Barber, C.E.; Dow, J.M.; Wootton, J.C.; Daniels, M.J. Genetic and molecular analysis of a cluster of rpf genes involved in positive regulation of synthesis of extracellular enzymes and polysaccharide in Xanthomonas campestris pathovar campestris. Mol. Gen. Genet. 1991, 226, 409–417. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Result | Characteristic | Result |
|---|---|---|---|
| Catalase | + | Phenylalanine deaminase | − |
| Oxidase | − | 2% Salt tolerance test with 2% NaCl | + |
| Gelatin hydrolysis | + | Salt tolerance test with 5% NaCl | + |
| V-P determination | + | Salt tolerance test with 7% NaCl | + |
| Propionate | − | Salt tolerance test with 10% NaCl | − |
| Urease | − | Amylolysis | + |
| Anaerobic culture | + | Glucose to acid | + |
| Nitrate reduction | + | Citrate | + |
| Lecithin enzyme | − | Indole test | − |
| Tyrosine hydrolysis | − | Casein hydrolysis | + |
| Broth at pH 5.7 | + | Broth at pH 6.8 | + |
| Treatment Conditions | Diameter of Inhibition Zones (mm) | Relative Percentage of Control (%) |
|---|---|---|
| Temperature (℃) | ||
| 25.0 (control) | 20.00 ± 1.47 | 100.0 |
| 45.9 | 19.83 ± 1.25 | 99.2 |
| 70.2 | 19.83 ± 0.62 | 99.2 |
| 81.6 | 16.50 ± 0.71 | 82.5 |
| 85.5 | 16.33 ± 0.85 | 81.7 |
| 91.0 | 16.17 ± 1.03 | 80.9 |
| 95.4 | 14.17 ± 1.25 * | 70.9 |
| 100.0 | 13.50 ± 1.87 * | 67.5 |
| Proteinase K (mg/mL) | ||
| 0.0 (control) | 14.33 ± 0.24 | 100.0 |
| 1.0 | 12.50 ± 0.71 | 87.2 |
| 2.0 | 12.50 ± 1.08 | 87.2 |
| 3.0 | 13.00 ± 1.08 | 90.7 |
| 4.0 | 13.17 ± 0.94 | 91.9 |
| Lipopeptide Isoforms | Fatty Acid Chain | Calculated (m/z) | ||
|---|---|---|---|---|
| [M + H]+ | [M + Na]+ | [M + K]+ | ||
| Iturin A/Mycosubtilin | C14 | 1043.5538 | 1065.5538 | 1081.5538 |
| C15 | 1057.5693 | 1079.5693 | 1095.5693 | |
| C16 | 1071.5858 | 1093.5858 | 1109.5858 | |
| C17 | 1085. 6018 | 1107.6018 | 1123.6018 | |
| Iturin B | C15 | 1058.6722 | 1080.6722 | 1096.6722 |
| C16 | 1072.6892 | 1094.6892 | 1110.6892 | |
| C17 | 1086.7038 | 1108.7038 | 1127.7038 | |
| C18 | 1100.7188 | 1122.7188 | 1138.7188 | |
| Surfactin | C13 | 994.6448 | 1016.6448 | 1032.6448 |
| C13, C14 | 1008.6606 | 1030.6606 | 1046.6606 | |
| C14, C15 | 1022.6762 | 1044.6762 | 1060.6762 | |
| C15, C16 | 1036.6920 | 1058.6920 | 1074.6920 | |
| C16 | 1050.7075 | 1072.7075 | 1088.7075 | |
| Fengycin | C14 | 1435.7714 | 1457.7714 | 1473.7714 |
| C15 | 1449.7926 | 1471.7926 | 1487.7926 | |
| C16, C14 | 1463.8063 | 1485.8063 | 1501.8063 | |
| C17, C15 | 1477.8203 | 1499.8203 | 1515.8203 | |
| C18, C16 | 1491.8354 | 1513.8354 | 1529.8354 | |
| C17 | 1505.8507 | 1527.8507 | 1543.8507 | |
| C18 | 1519.8661 | 1541.8661 | 1557.8661 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Liang, L.; Shao, H.; Ye, X.; Yang, X.; Chen, X.; Shi, Y.; Zhang, L.; Xu, L.; Wang, J. Isolation of the Novel Strain Bacillus amyloliquefaciens F9 and Identification of Lipopeptide Extract Components Responsible for Activity against Xanthomonas citri subsp. citri. Plants 2022, 11, 457. https://doi.org/10.3390/plants11030457
Wang X, Liang L, Shao H, Ye X, Yang X, Chen X, Shi Y, Zhang L, Xu L, Wang J. Isolation of the Novel Strain Bacillus amyloliquefaciens F9 and Identification of Lipopeptide Extract Components Responsible for Activity against Xanthomonas citri subsp. citri. Plants. 2022; 11(3):457. https://doi.org/10.3390/plants11030457
Chicago/Turabian StyleWang, Xin, Liqiong Liang, Hang Shao, Xiaoxin Ye, Xiaobei Yang, Xiaoyun Chen, Yu Shi, Lianhui Zhang, Linghui Xu, and Junxia Wang. 2022. "Isolation of the Novel Strain Bacillus amyloliquefaciens F9 and Identification of Lipopeptide Extract Components Responsible for Activity against Xanthomonas citri subsp. citri" Plants 11, no. 3: 457. https://doi.org/10.3390/plants11030457