
Trade-Off between Facilitation and Interference of Allelopathic Compounds in Vegetation Recovery: The Case of Rosmarinus officinalis in Degraded Gypsum Habitats



Abstract
:1. Introduction
2. Results
2.1. Effects of Treatments on Seedling Emergence
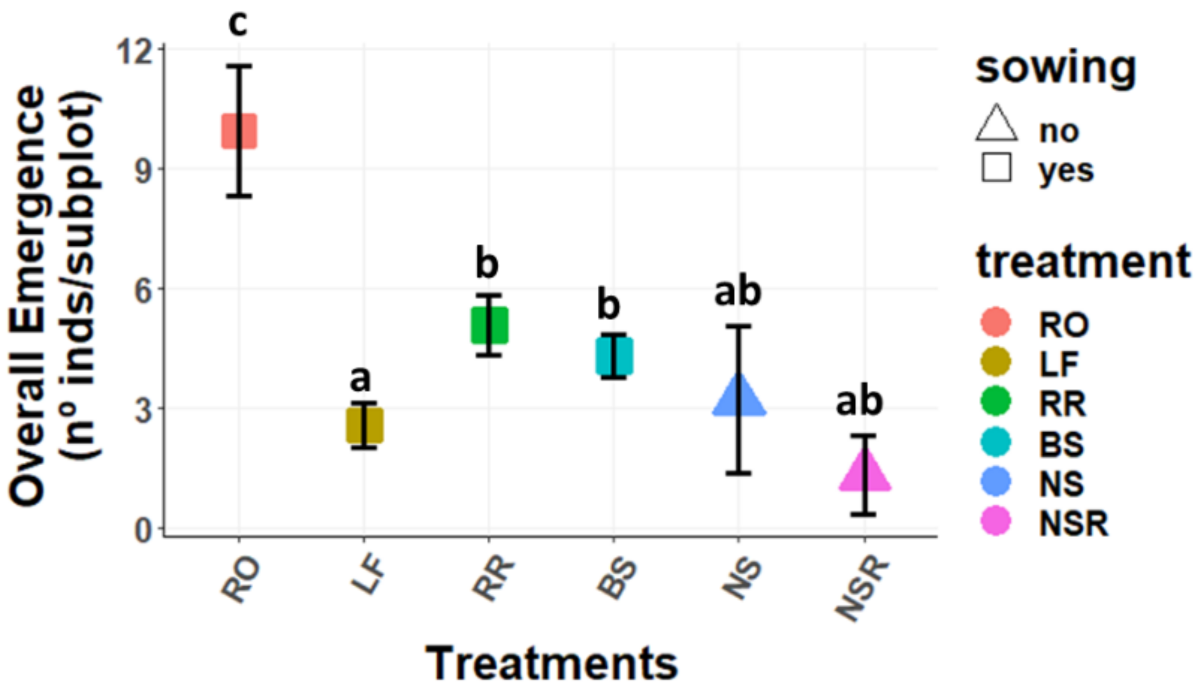
2.1.1. Overall Emergence
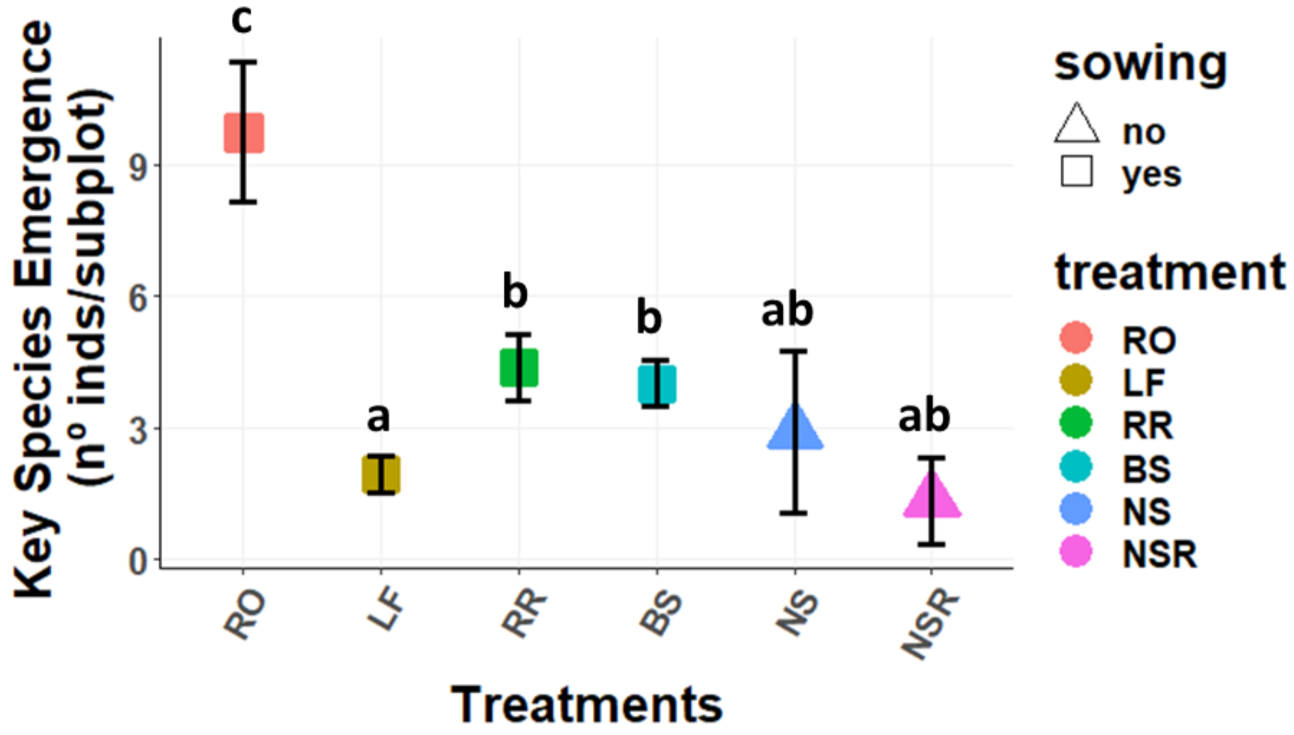
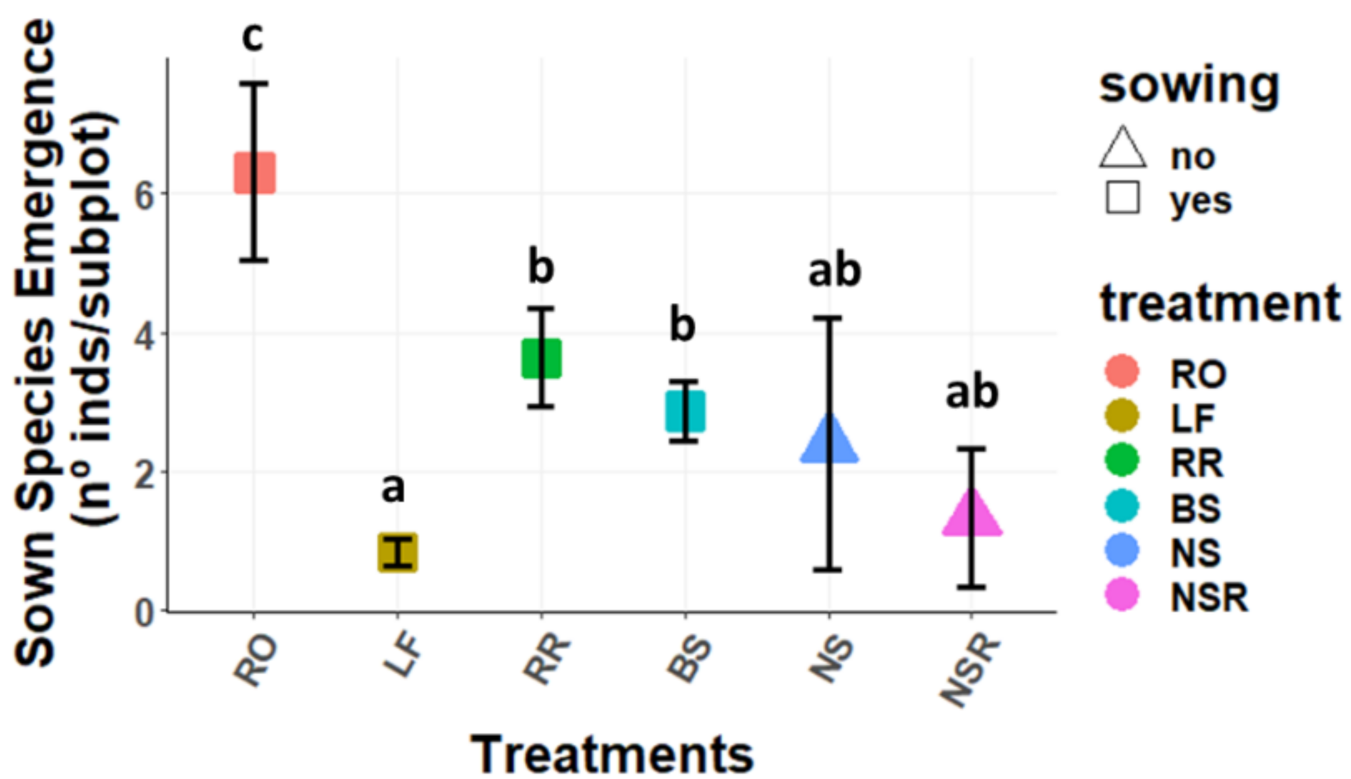
2.1.2. Emergence Per Species
2.2. Essential Oils in Soils and Leaves
3. Discussion
4. Materials and Methods
4.1. Study Area
4.2. Experimental Design
4.3. Sampling
4.3.1. Seedling Emergence
4.3.2. Terpene Content
4.4. Laboratory Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Boyadgiev, T.G.; Verheye, W.H. Contribution to a utilitarian classification of gypsiferous soil. Geoderma 1996, 74, 321–338. [Google Scholar] [CrossRef]
- Riba, O.; Macau, F. Situación, Características y Extension de los Terrenos Yesíferos en España; Servicio Geológico de Obras Públicas: Madrid, Spain, 1962. [Google Scholar]
- Rivas Goday, S.; Borja Carbonell, J.; Monasterio Fernández, A.; Fernández-Galiano, E.; Rivas Martínez, S. Aportaciones a la Fitosociología hispánica (Proyectos de comunidades hispánicas). An. Jard. Bot. Madr. 1956, 13, 333–422. Available online: http://www.rjb.csic.es/jardinbotanico/ficheros/documentos/pdf/anales/1957/Anales_14(1)_433_500a.pdf (accessed on 17 January 2022).
- Loidi, J.; Costa, M. Sintaxonomía de los matorrales gipsícolas españoles. Fitosociología 1957, 32, 221–227. Available online: https://jolube.files.wordpress.com/2008/04/loidi_costa_1997.pdf (accessed on 17 January 2022).
- Meyer, S. The ecology of gypsophile endemism in the Eastern Mojave Desert. Ecology 1986, 67, 1303–1313. [Google Scholar] [CrossRef]
- Commission of the European Communities. Council Directive 92/43/EEC of 21st May 1992 on the Conservation of Natural Habitats and of Wild Fauna and Flora; European Commission’s Directorate-General for the Environment: Brussels, Belgium, 1992; Available online: https://ec.europa.eu/environment/nature/legislation/habitatsdirective/index_en.htm (accessed on 16 October 2021).
- Mota, J.F.; Sola, A.J.; Jimenez-Sanchez, M.L.; Perez-García, F.J.; Merlo, M.E. Gypsicolous flora, conservation and restoration of quarries in the southeast of the Iberian Peninsula. Biodivers. Conserv. 2004, 13, 1797–1808. [Google Scholar] [CrossRef]
- Ballesteros, M.; Cañadas, E.M.; Foronda, A.; Peñas, J.; Lorite, J. Vegetation recovery of gypsum quarries: Short-term sowing response to different soil treatments. Appl. Veg. Sci. 2012, 15, 187–197. [Google Scholar] [CrossRef]
- Ballesteros, M.; Cañadas, E.M.; Foronda, A.; Peñas, J.; Lorite, J. Conservation status of the narrow endemic gypsophile Ononis tridentata subsp. crassifolia in southern Spain: Effects of habitat disturbance. Fauna Flora Int. Oryx 2013, 47, 199–202. [Google Scholar] [CrossRef] [Green Version]
- Mota, J.F.; Martínez-Hernandez, F.; Guirado, J.S. Diversidad Vegetal de las Yeseras Ibericas. El reto de los Archipielagos Edaficos Para la Biología de la Conservacion; ADIF-Mediterraneo Asesores Consultores: Almería, Spain, 2011; Available online: https://jolube.wordpress.com/2015/10/21/libro-diversidad-vegetal-de-las-yeseras-ibericas-en-pdf (accessed on 17 January 2022).
- Craig, J.; Vaughan, D.J.; Skinner, B.J. Recursos de la Tierra: Origen, uso e Impacto Ambiental, 3rd ed.; Pearson Prentice Hall: Madrid, Spain, 2007. [Google Scholar]
- Dana, E.D.; Mota, J.F. Vegetation and soil recovery on gypsum outcrops in semi-arid Spain. J. Arid Environ. 2006, 65, 444–459. [Google Scholar] [CrossRef]
- Mota, J.F.; Sola, A.J.; Dana, D.E.; Jimenez-Sanchez, M.L. Plant succession in abandoned gypsum quarries. Phytocoenologia 2003, 33, 13–28. [Google Scholar] [CrossRef]
- Sharma, K.D.; Kumar, S.; Gough, L.P. Rehabilitation of gypsum mined-lands in the Indian Desert. Arid Land Res. Manag. 2001, 15, 61–76. [Google Scholar] [CrossRef]
- Blignaut, A.; Milton, S.J. Effects of multispecies clumping on survival of three succulent plant species translocated onto mine spoil in the Succulent Karoo Desert, South Africa. Restor. Ecol. 2005, 13, 15–19. [Google Scholar] [CrossRef]
- Matesanz, S.; Valladares, F. Improving revegetation of gypsum slopes is not a simple matter of adding autochthonous species: Insights from a multispecies experiment. Ecol. Eng. 2007, 30, 67–77. [Google Scholar] [CrossRef]
- Ballesteros, M.; Cañadas, E.M.; Marrs, R.H.; Foronda, A.; Martín-Peinado, F.J.; Lorite, J. Restoration of Gypsicolous Vegetation on Quarry Slopes: Guidance for Hydroseeding Under Contrasting Inclination and Aspect. Land Degrad. Dev. 2017, 28, 2146–2154. [Google Scholar] [CrossRef]
- Lorite, J.; Ballesteros, M.; García-Robles, H.; Cañadas, E.M. Economic evaluation of ecological restoration options in gypsum habitats after mining. J. Nat. Conserv. 2021, 59, 125935. [Google Scholar] [CrossRef]
- Ballesteros-Jiménez, M. Restoration of Gypsum Habitats Affected by Quarrying: Guidance for Assisting Vegetation Recovery. Ph.D. Thesis, Universidad de Granada, Granada, Spain, 2018. Available online: http://digibug.ugr.es/handle/10481/49583 (accessed on 17 January 2022).
- De Mastro, G.; El Mahdi, J.; Ruta, C. Bioherbicidal Potential of the Essential Oils from Mediterranean Lamiaceae for Weed Control in Organic Farming. Plants 2021, 10, 818. [Google Scholar] [CrossRef] [PubMed]
- Angelini, L.G.; Carpanese, G.; Cioni, P.L.; Morelli, I.; Macchia, M.; Flamini, G. Essential Oils from Mediterranean Lamiaceae as Weed Germination Inhibitors. J. Agric. Food Chem. 2003, 51, 6158–6164. [Google Scholar] [CrossRef]
- Zimdahl, R.L. Chapter 9-Allelopathy. In Fundamentals of Weed Science, 5th ed.; Zimdahl, R.L., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 253–270. ISBN 9780128111437. [Google Scholar] [CrossRef]
- Muller, C.H. Allelopathy as a factor in ecological process. Vegetation 1969, 18, 348–357. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: New York, NY, USA, 1984. [Google Scholar]
- Rizvi, S.J.H.; Rizvi, V. Allelopathy: Basic and Applied Aspects; Chapman and Hall: London, UK, 1992. [Google Scholar]
- Chou, C.H. Roles of allelopathy in plant biodiversity and sustainable agriculture. Crit. Rev. Plant Sci. 1999, 18, 609–636. [Google Scholar] [CrossRef]
- Scognamiglio, M.; D’Abrosca, B.; Esposito, A.; Pacifico, S.; Monaco, P.; Fiorentino, A. Plant growth inhibitors: Allelopathic role or phytotoxic effects? Focus on Mediterranean biomes. Phytochem. Rev. 2013, 12, 803–830. [Google Scholar] [CrossRef]
- Wardle, D.A.; Nicholson, K.S.; Rahman, A. Use of a comparative approach to identify allelopathic potential and relationship between allelopathy bioassays and competition experiments for ten grassland and plant species. J. Chem. Ecol. 1996, 22, 933–948. [Google Scholar] [CrossRef]
- Ben-Kaab, S.; Rebey, I.B.; Hanafi, M.; Berhal, C.; Fauconnier, M.L.; de-Clerck, C.; Ksouri, R.; Jijakli, H. Rosmarinus officinalis essential oil as an effective antifungal and herbicidal agent. Span. J. Agric. Res. 2019, 17, e1006. [Google Scholar] [CrossRef] [Green Version]
- Ramezani, S.; Saharkhiz, M.J.; Ramezani, F.; Fotokian, M.H. Use of Essential Oils as Bioherbicides. J. Essent. Oil-Bear. Plants 2008, 11, 319–327. [Google Scholar] [CrossRef]
- Eslahi, H.; Fahimi, N.; Sardarian, A.R. Chemical composition of essential oils. In Essential Oils in Food Processing: Chemistry, Safety and Applications; Hashemi, S.M.B., Khaneghah, A.M., de Souza Sant’ Ana, A., Eds.; IFT Press; Wiley Blackwell: Hoboken, NJ, USA, 2017; pp. 119–171. [Google Scholar]
- Arminante, F.; De Falco, E.; De Feo, V.; De Martino, L.; Mancini, E.; Quaranta, E. Allelopathic activity of essential oils from Mediterranean Labiatae. Acta Hortic. 2006, 723, 347–356. [Google Scholar] [CrossRef]
- Fisher, N.H. The function of mono and sesquiterpenes as plant germination and growth regulators. In The Science of Allelopathy; Putnam, A.R., Tang, C.S., Eds.; Wiley: New York, NY, USA, 1986; pp. 203–218. [Google Scholar]
- Vaughn, S.; Spencer, G. Volatile Monoterpenes as Potential Parent Structures for New Herbicides. Weed Sci. 1993, 41, 114–119. [Google Scholar] [CrossRef] [Green Version]
- Flamini, G. Natural Herbicides as a Safer and More Environmentally Friendly Approach to Weed Control: A Review of the Literature Since 2000. Stud. Nat. Prod. Chem. Bioact. Nat. Prod. 2012, 38, 353–396. [Google Scholar] [CrossRef]
- Chaves-Lobón, N.; Sosa-Díaz, T.; Valares-Masa, C.; Alías-Gallego, J.C. Chapter 10: Terpenoids. In Research Methods in Plant Science; Narwal, S.S., Szajdak, L., Sampietro, D.A., Eds.; Soil Allelochemicals; Studium Press, LLC: Houston, TX, USA, 2010; Volume 1, pp. 223–238. [Google Scholar]
- Scrivanti, L.R.; Zunino, M.P.; Zygadlo, J.A. Tagetes minuta and Schinus areira essential oils as allelopathic agents. Biochem. Syst. Ecol. 2003, 31, 563–572. [Google Scholar] [CrossRef]
- Ormeño, E.; Baldy, V.; Ballini, C.; Fernandez, C. Production and diversity of volatile terpenes from plants on calcareous and siliceous soils: Effect of soil nutrients. Springer Science + Business Media, LLC 2008. J. Chem. Ecol. 2008, 34, 1219–1229. [Google Scholar] [CrossRef]
- Alipour, M.; Saharkhiz, M.J.; Niakousari, M.; Seidi Damyeh, M. Phytotoxicity of encapsulated essential oil of rosemary on germination and morphophysiological features of amaranth and radish seedlings. Sci. Hortic. 2019, 243, 131–139. [Google Scholar] [CrossRef]
- Alias-Gallego, J.C.; González-Caro, J.; Hinojal-Campos, V.; Chaves-Lobón, N. Effect of leaf litter from Cistus ladanifer L. on the germination and growth of accompanying shrubland species. Plants 2020, 9, 593. [Google Scholar] [CrossRef]
- Soliveres, S.; Maestre, F.T. Plant-plant interactions, environmental gradients and plant diversity: A global synthesis of community-level studies. Perspect. Plant Ecol. Evol. Syst. 2014, 16, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Siles, G.; Rey, P.J.; Alcántara, J.M.; Ramírez, J.M. Assessing the long-term contribution of nurse plants to restoration of Mediterranean forests through Markovian models. J. Appl. Ecol. 2008, 45, 1790–1798. [Google Scholar] [CrossRef]
- Pugnaire, F.I.; Luque, M.T. Changes in plant interactions along a gradient of environmental stress. Oikos 2001, 93, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Tormo-Blanes, J.; Amat-Martínez, B.; Cortina, J. Litter as a filter for germination in semi-arid Stipa tenacissima steppes. J. Arid Environ. 2020, 183, 104258. [Google Scholar] [CrossRef]
- Hassiotis, C.N.; Lazari, D.M. Decomposition process in the Mediterranean region. Chemical compounds and essential oil degradation from Myrtus communis. Int. Biodeterior. Biodegrad. 2010, 64, 356–362. [Google Scholar] [CrossRef]
- Cañadas, E.M.; Valle, F.; Ballesteros, M.; Lorite, J. Does gypsum influence seed germination? Turk. J. Bot. 2014, 38, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Cañadas, E.M.; Ballesteros, M.; Foronda, A.; Navarro, F.B.; Jiménez, M.N.; Lorite, J. Enhancing seedling production of native species to restore gypsum habitats. J. Environ. Manag. 2015, 163, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Escudero, A.; Iriondo, J.M.; Olano, J.M.; Rubio, A.; Somolinos, R.C. Factors affecting establishment of a gypsophite: The case of Lepidium subulatum (Brassicaceae). Am. J. Bot. 2000, 87, 861–871. [Google Scholar] [CrossRef] [Green Version]
- Escudero, A.; Palacio, S.; Maestre, F.T.; Luzuriaga, A.L. Plant life on gypsum: A review of its multiple facets. Biol. Rev. 2014, 90, 1–18. [Google Scholar] [CrossRef]
- Escudero, A.; Somolinos, R.; Olano, J.M.; Rubio, A. Factors controlling the establishment of Helianthemum squamatum, an endemic gypsophile of semi-arid Spain. J. Ecol. 1999, 87, 290–302. Available online: http://www.jstor.org/stable/2648320 (accessed on 17 January 2022). [CrossRef]
- Foronda, A.; Pueyo, Y.; Arroyo, A.I.; Saiz, H.; Giner, M.L.; Alados, C.L. The role of nurse shrubs on the spatial patterning of plant establishment in semi-arid gypsum plant communities. J. Arid Environ. 2019, 160, 82–90. [Google Scholar] [CrossRef]
- García-Fayos, P.; Gasque, M. Seed vs. microsite limitation for seedling emergence in the perennial grass Stipa tenacissima L. (Poaceae). Acta Oecologica 2006, 30, 276–282. [Google Scholar] [CrossRef]
- Olano, J.M.; Eugenio, M.; Escudero, A. Site effect is stronger than species identity in driving demographic responses of Helianthemum (Cistaceae) shrubs in gypsum environments. Am. J. Bot. 2011, 98, 1016–1023. [Google Scholar] [CrossRef]
- Yakoubi, S.; Cherrat, A.; Diouri, M.; Hilali, F.E.; Zair, T. Chemical composition and antibacterial activity of Thymus zygis subsp. gracilis (Boiss.) R. Morales essential oils from Morocco. Mediterr. J. Chem. 2014, 3, 746–758. [Google Scholar] [CrossRef]
- Hierro, J.L.; Callaway, R.M. The Ecological Importance of Allelopathy. Annu. Rev. Ecol. Evol. 2021, 52, 25–45. [Google Scholar] [CrossRef]
- Linhart, Y.B.; Gauthier, P.; Keefover-Ring, K.; Thompson, J.D. Variable Phytotoxic Effects of Thymus vulgaris (Lamiaceae) Terpenes on Associated Species. Int. J. Plant Sci. 2015, 176, 20–30. [Google Scholar] [CrossRef]
- Inderjit; del Moral, R. Is separating resource competition from allelopathy realistic? Bot. Rev. 1997, 63, 221–230. [Google Scholar] [CrossRef]
- Azirak, S.; Karaman, S. Allelopathic effect of some essential oils and components on germination of weed species. Acta Agric. Scand.-B Soil Plant Sci. 2008, 58, 88–92. [Google Scholar] [CrossRef]
- Ren, H.; Yang, L.; Liu, N. Nurse plant theory and its application in ecological restoration in lower subtropics of China. Prog. Nat. Sci. 2008, 18, 137–142. [Google Scholar] [CrossRef]
- Aldaya, F.; Delgado, F.; Diaz de Federico, A.; Fontboté, J.M.; Garcia-Duedas, V.; Ramón-Lluch, R. Mapa y Memoria Explicativa de la Hoja 1011 (Guadix) del Mapa Geológico National a Escala 150 000; IGME: Madrid, Spain, 1980. [Google Scholar]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, Update 2015. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2014; 192p. [Google Scholar]
- Marchal, F.M.; Lendínez, M.I.; Salazar, C.; Torres, J.A. Aportaciones al conocimiento de la vegetación gipsícola en el occidente de la provincia de Granada (Sur de España). Lazaroa 2008, 29, 95–100. [Google Scholar]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 6212, Chloroform. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Chloroform (accessed on 18 October 2021).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012; Available online: https://www.scirp.org/(S(vtj3fa45qm1ean45vvffcz55))/reference/ReferencesPapers.aspx?ReferenceID=1462563 (accessed on 17 January 2022).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.scirp.org/(S(351jmbntvnsjt1aadkposzje))/reference/ReferencesPapers.aspx?ReferenceID=2144573 (accessed on 17 January 2022).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Wickham, H. The split-apply-combine strategy for data analysis. J. Stat. Softw. 2011, 40, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5–7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 17 January 2022).
- Wheeler, R.E.; Torchiano, M. Permutation Tests for Linear Models in R. The Comprehensive R Archive Network, 1. 2016. Available online: https://cran.r-project.org/web/packages/lmPerm/vignettes/lmPerm.pdf (accessed on 17 January 2022).




{kind=link}
{kind=link}
{kind=link}
| Key Species | Emergence (Mean + SE) Per Treatment | |||
|---|---|---|---|---|
| RO | LF | RR | BS | |
| O. tridentata subsp. crassifolia | 0.13 ± 0.10 a | 0.13 ± 0.08 a | 0.07 ± 0.04 a | 0.13 ± 0.06 a |
| H. squamatum | 6.03 ± 1.25 d | 0.67 ± 0.19 a | 3.50 ± 0.71 c | 2.23 ± 0.39 b |
| L. subulatum | 0.13 ± 0.08 ab | 0.03 ± 0.03 a | 0.07 ± 0.05 a | 0.50 ± 0.20 b |
| R. officinalis | 1.4 ± 0.89 b | 0.13 ± 0.08 a | 0.10 ± 0.07 a | 0.10 ± 0.06 a |
| H. syriacum | 0.17 ± 0.11 ab | 0.00 ± 0.00 a | 0.03 ± 0.03 a | 0.23 ± 0.08 b |
| T. zygis subsp. gracilis | 2.03 ± 0.79 b | 0.97 ± 0.29 a | 0.53 ± 0.24 a | 1.03 ± 0.20 a |
| S. tenacissima | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.10 ± 0.07 a | 0.00 ± 0.00 a |
| N | Terpene Compound | Retention Time (Seconds) | Key Terpene | Sample Types | |||
|---|---|---|---|---|---|---|---|
| Leaves | Soils | ||||||
| GL | FL | BS | RS | ||||
| 1 | tricyclene | 7.34 | No | X | X | ||
| 2 | thujene | 7.46 | No | X | |||
| 3 | α-pinene | 7.59–7.78 | Yes | X | X | X | |
| 4 | camphene | 8.11–8.27 | Yes | X | X | X | |
| 5 | verbenene | 8.22 | No | X | |||
| 6 | β-pinene | 9.03–9.19 | Yes | X | X | X | |
| 7 | 1-octen-3-ol | 9.21–9.27 | No | X | |||
| 8 | 3- octanone | 9.34–9.45 | No | X | |||
| 9 | β-myrcene | 9.51–9.65 | No | X | X | X | |
| 10 | 3- heptanol | 9.86 | No | X | |||
| 11 | α-phellandrene | 10.07–10.18 | Yes | X | X | ||
| 12 | artemisatriene | 10.45–10.56 | No | X | |||
| 13 | p-cymene | 10.74–10.84 | Yes | X | X | X | |
| 14 | limonene | 10.94–11.05 | Yes | X | X | X | |
| 15 | eucalyptol (1’8 cineole) | 11.04–11.16 | Yes | X | X | X | |
| 16 | ɤ_terpinene | 11.97–12.06 | No | X | X | ||
| 17 | terpinolene | 12.97–13.04 | No | X | |||
| 18 | linalool | 13.51–13.61 | No | X | X | X | |
| 19 | 3.5 heptadien-2-ol-2.6 | 14.26–14.33 | No | X | X | X | |
| 20 | camphenol | 14.45–14.53 | No | X | |||
| 21 | pinocarveol | 14.99–15.13 | No | X | |||
| 22 | camphor | 15.17–15.36 | Yes | X | X | X | |
| 23 | iso-pinocamphone | 15.66–15.73 | No | X | |||
| 24 | borneol | 16.10–16.21 | Yes | X | X | X | |
| 25 | terpinen-4-ol | 16.41–16.49 | No | X | X | X | |
| 26 | α-terpineol | 16.97–17.02 | No | X | X | X | |
| 27 | α-santolin-alcohol | 17.30 | No | X | |||
| 28 | verbenone | 17.42 | No | X | X | X | |
| 29 | bornyl acetate | 20.04 | No | X | |||
| 30 | carvacrol | 20.3 | Yes | X | |||
| 31 | copaene | 23.11 | No | X | |||
| 32 | caryophyllene | 24.52–24.61 | No | X | X | X | |
| 33 | aromadendrene | 25.63 | No | X | X | ||
| 34 | cis-α-bisabolene | 25.715 | No | X | X | X | |
| 35 | methyl 8′11′14′17-eicosatetraenoate | 27.544 | No | X | |||
| 36 | caryophyllene oxide | 29.42 | No | X | X | X | |
| 37 | farnesol | 29.51–30.00 | No | X | |||
| 38 | tridecan | 31.05 | No | X | |||
| 39 | ledene oxyde | 31.58 | No | X | |||
| Pairs | Df | SumsOfSqs | F.Model | R2 | p. Value | p. Adjusted | Sig |
|---|---|---|---|---|---|---|---|
| FL vs. GL | 1.00 | 0.27 | 4.92 | 0.38 | 0.01 | 0.02 | * |
| FL vs. RS | 1.00 | 0.13 | 1.17 | 0.13 | 0.32 | 0.97 | |
| GL vs. RS | 1.00 | 0.33 | 3.55 | 0.31 | 0.01 | 0.02 | * |
| Terpenes | Sample Types | |||
|---|---|---|---|---|
| Leaves | Soils | |||
| GL | FL | BS | RS | |
| α-pinene | b | b | a | b |
| camphene | b | b | a | b |
| β-pinene | c | ab | a | bc |
| α-phellandrene | a | a | a | a |
| p-cymene | ab | b | a | ab |
| limonene | b | ab | a | ab |
| eucalyptol | b | b | a | b |
| camphor | b | b | a | b |
| borneol | b | b | a | b |
| carvacrol | a | a | a | a |
| Treatments | Rosemary Plant Presence | Rosemary Leaves Presence | Allelopathic Compounds | Facilitation Effect | Effect over Emergence |
|---|---|---|---|---|---|
| RO | Yes | Yes | Yes | Yes | +++ |
| LF | Removal | Yes | Yes | No | -- |
| RR | Removal | Removal | Residual | No | + |
| BS | Never | Never | No | No | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Robles, H.; Cañadas, E.M.; Lorite, J.; Fernández-Ondoño, E. Trade-Off between Facilitation and Interference of Allelopathic Compounds in Vegetation Recovery: The Case of Rosmarinus officinalis in Degraded Gypsum Habitats. Plants 2022, 11, 459. https://doi.org/10.3390/plants11030459
García-Robles H, Cañadas EM, Lorite J, Fernández-Ondoño E. Trade-Off between Facilitation and Interference of Allelopathic Compounds in Vegetation Recovery: The Case of Rosmarinus officinalis in Degraded Gypsum Habitats. Plants. 2022; 11(3):459. https://doi.org/10.3390/plants11030459
Chicago/Turabian StyleGarcía-Robles, Helena, Eva María Cañadas, Juan Lorite, and Emilia Fernández-Ondoño. 2022. "Trade-Off between Facilitation and Interference of Allelopathic Compounds in Vegetation Recovery: The Case of Rosmarinus officinalis in Degraded Gypsum Habitats" Plants 11, no. 3: 459. https://doi.org/10.3390/plants11030459