
Effects of Light Spectral Quality on Photosynthetic Activity, Biomass Production, and Carbon Isotope Fractionation in Lettuce, Lactuca sativa L., Plants


 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Cultivation Conditions
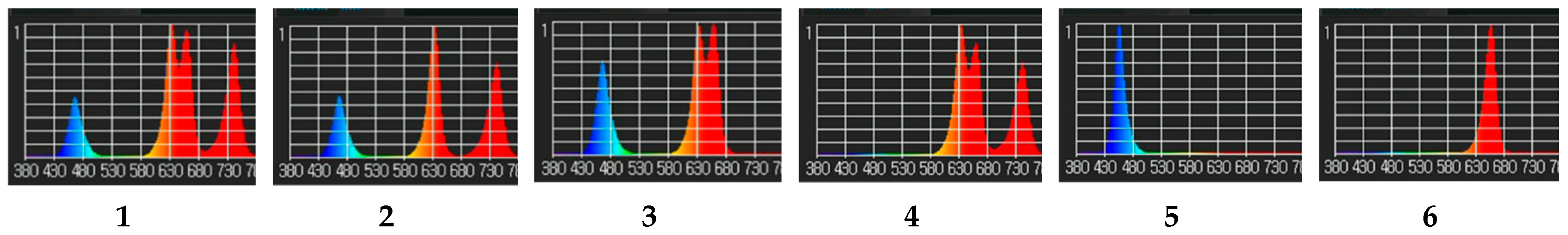
2.3. Light Treatments
2.4. Plant Growth Parameters Analyses
2.5. Photosynthesis and Transpiration
2.6. Chlorophyll a Fluorescence Determination
2.7. Carbon Isotope Discrimination (Δ) Measurements
2.8. Statistical Analysis of Experimental Data
3. Results
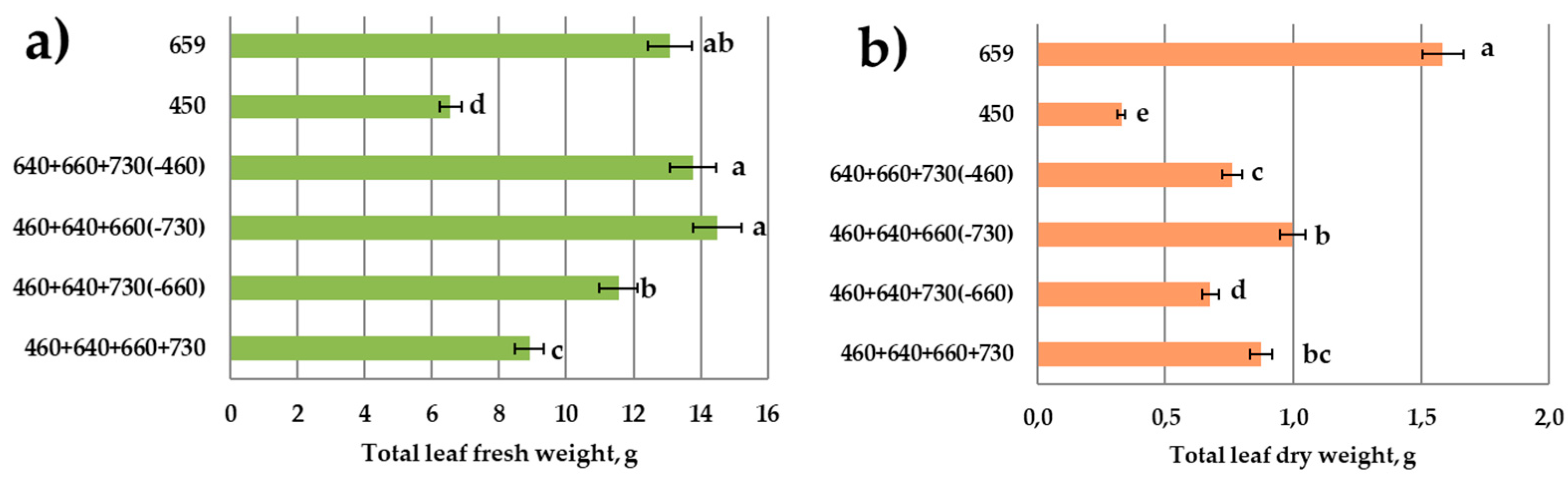
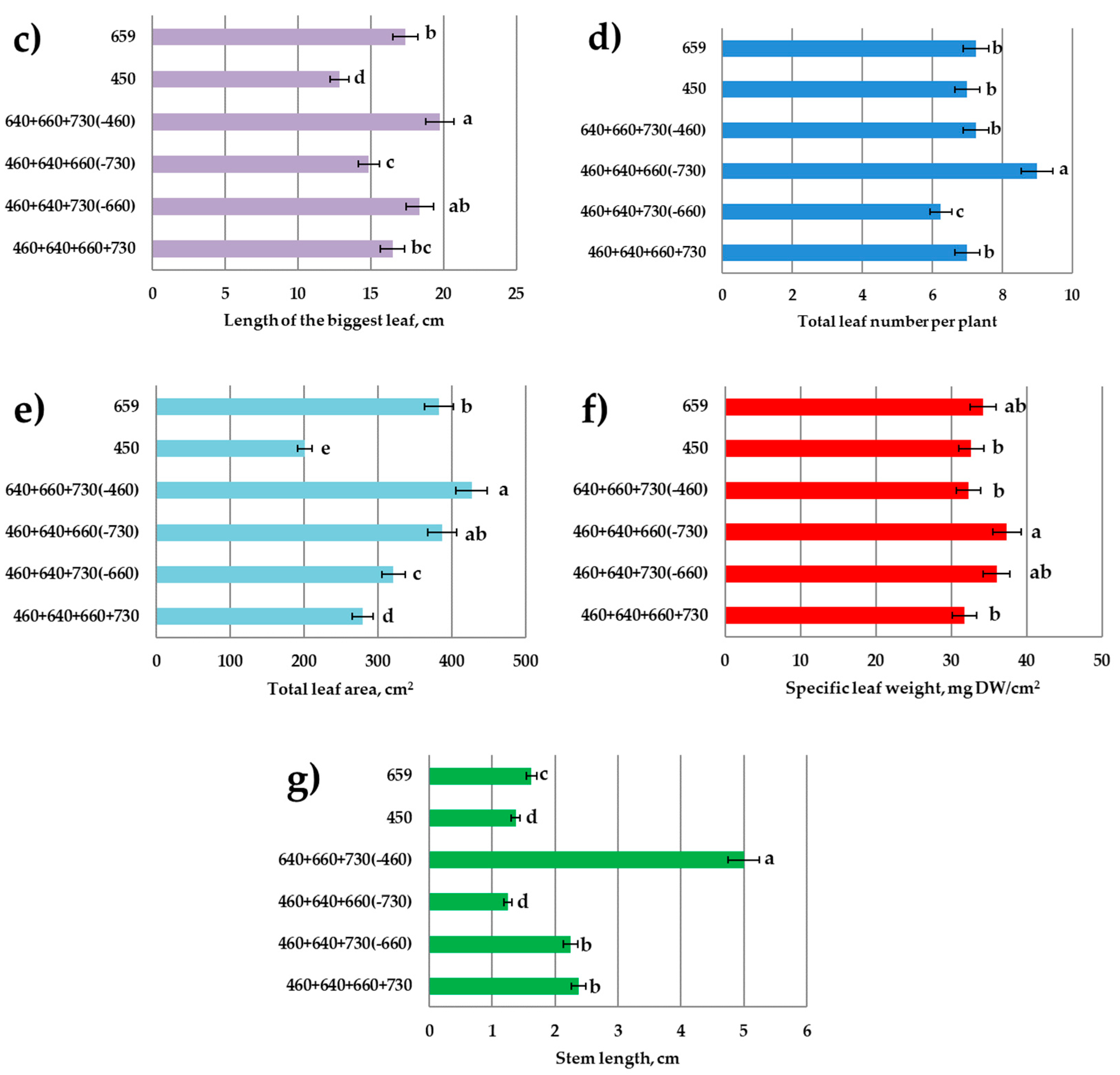
3.1. Growth Responses
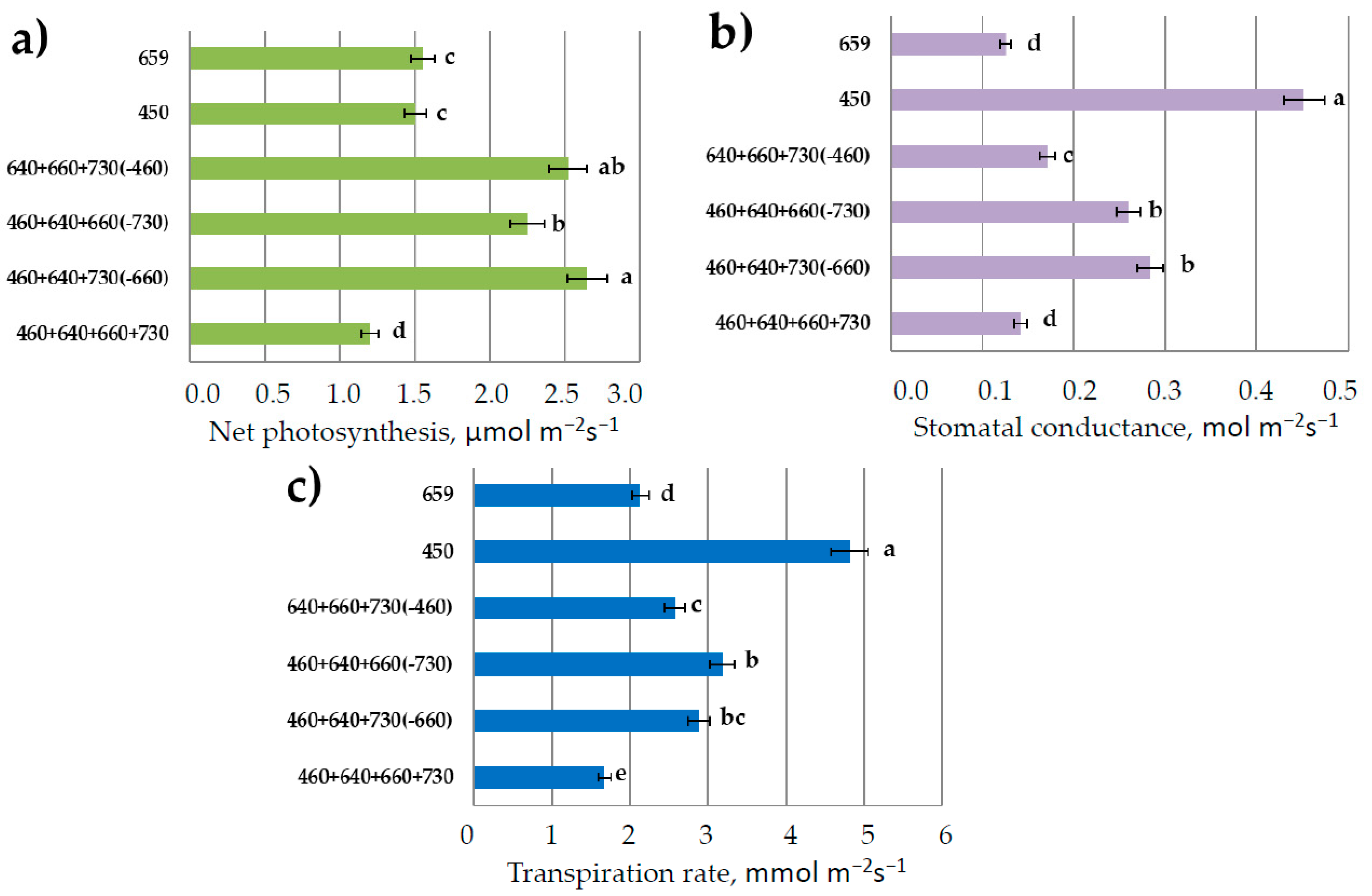
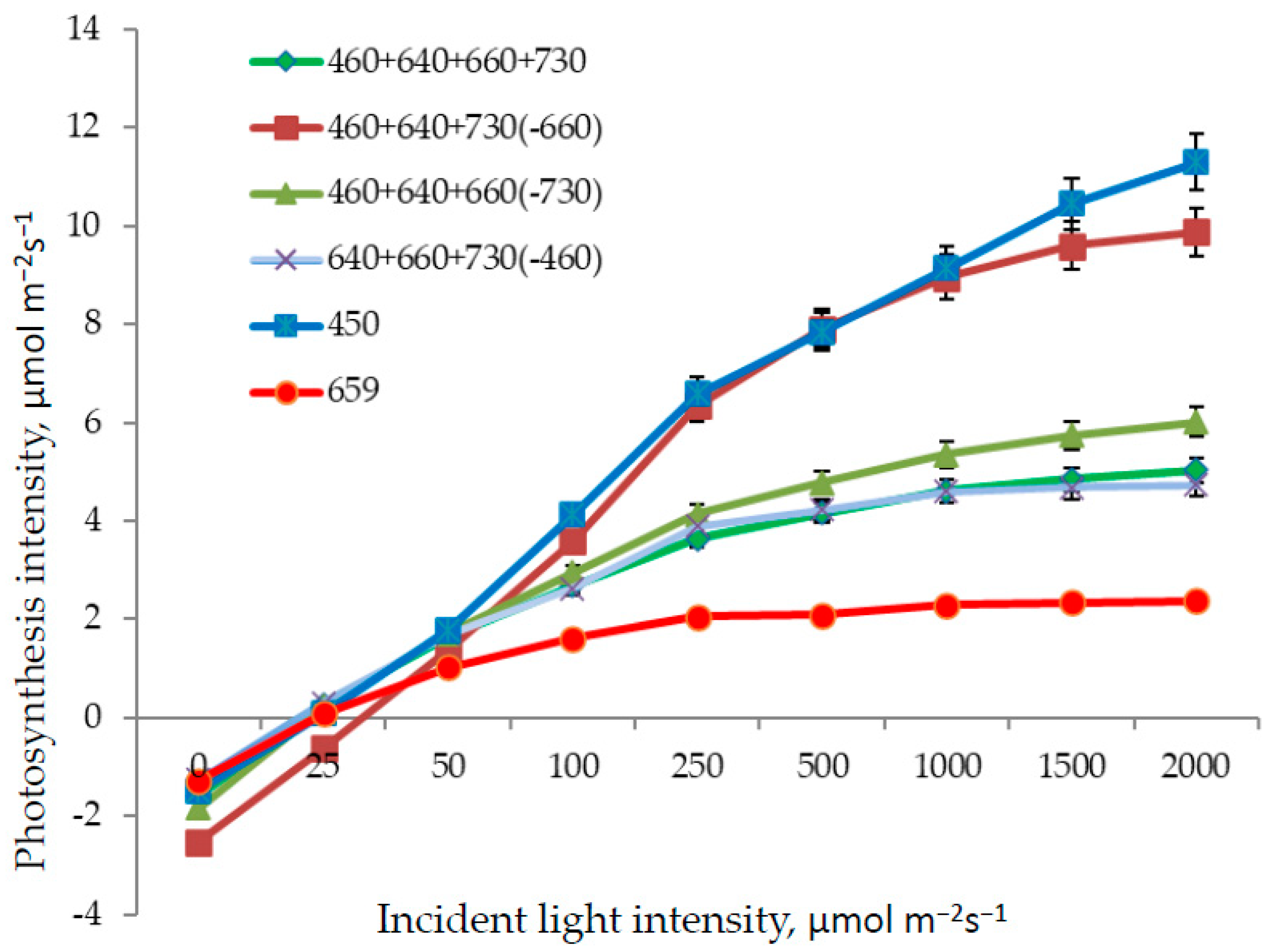
3.2. Photosynthesis and Transpiration
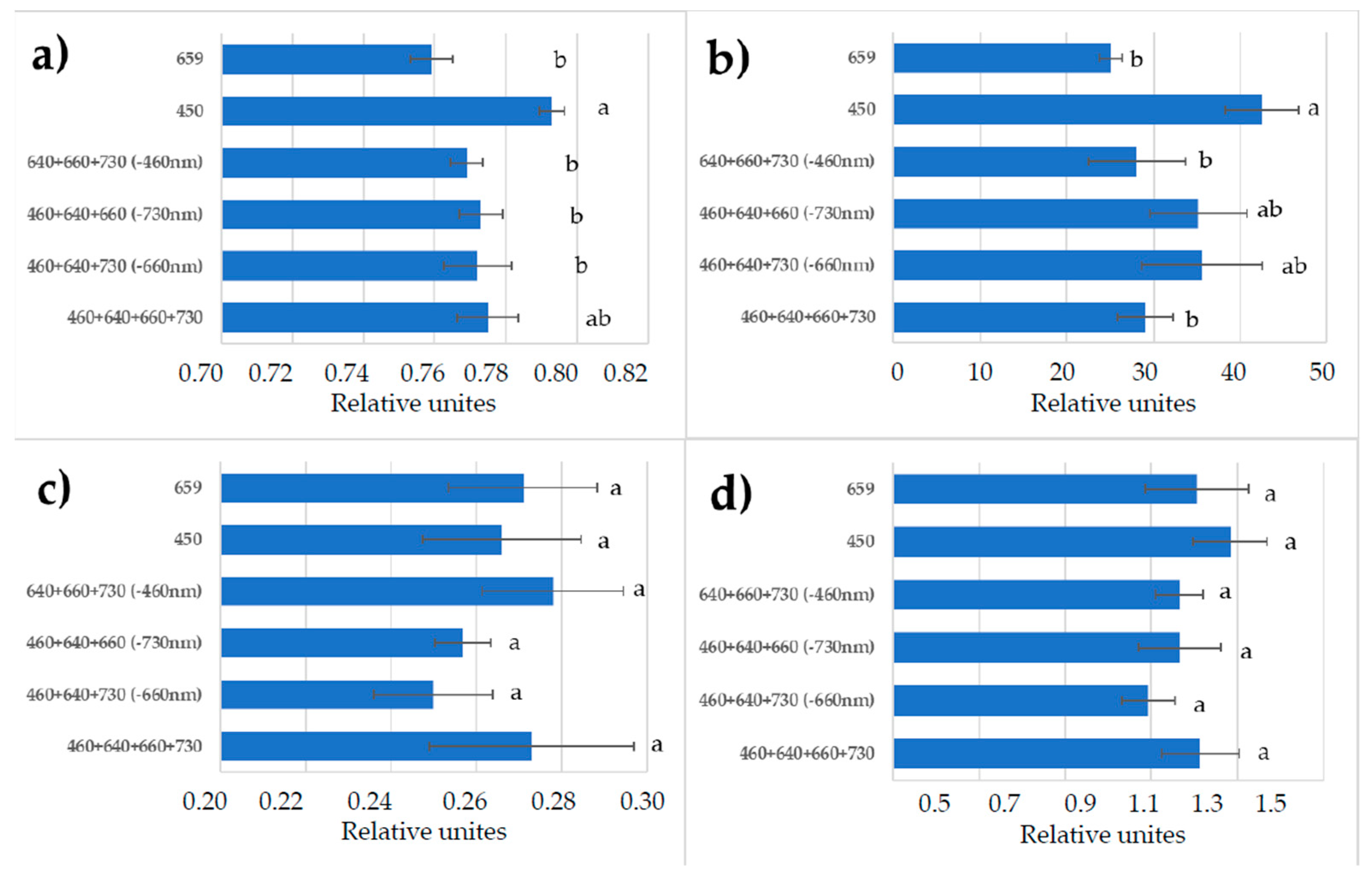
3.3. Chlorophyll a Fluorescence
3.4. Carbon Isotopes Discrimination
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bian, Z.; Jiang, N.; Grundy, S.; Lu, C. Uncovering LED light effects on plant growth: New angles and perspectives-LED light for improving plant growth, nutrition and energy-use efficiency. Acta Hortic. 2017, 1227, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Loconsole, D.; Cocetta, G.; Santoro, P.; Ferrante, A. Optimization of LED Lighting and Quality Evaluation of Romaine Lettuce Grown in an Indoor Cultivation System. Sustainability 2019, 11, 841. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.L.; Yang, Q.C.; Song, W.P.; Wang, L.C.; Guo, W.Z.; Xue, X.Z. Growth and nutritional properties of lettuce affected by different alternating intervals of red and blue LED irradiation. Sci. Hortic. 2017, 223, 44–52. [Google Scholar] [CrossRef]
- Lee, M.J.; Son, K.H.; Oh, M.M. Increase in biomass and bioactive compounds in lettuce under various ratios of red to far-red LED light supplemented with blue LED light. Hortic. Environ. Biotechnol. 2016, 57, 139–147. [Google Scholar] [CrossRef]
- Fankhauser, C.; Chory, J. Light control of plant development. Annu. Rev. Cell Dev. Biol. 1997, 13, 203–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olle, M.; Viršile, A. The effects of light-emitting diode lighting on greenhouse plant growth and quality. Agric. Food Sci. 2013, 22, 223–234. [Google Scholar] [CrossRef]
- Ouzounis, T.; Rosenqvist, E.; Ottosen, C.O. Spectral Effects of Artificial Light on Plant Physiology and Secondary Metabolism: A Review. Hort. Sci. 2015, 50, 1128–1135. [Google Scholar] [CrossRef] [Green Version]
- Berkovich, Y.A.; Konovalova, I.O.; Smolyanina, S.O.; Erokhin, A.N.; Avercheva, O.V.; Bassarskaya, E.M.; Kochetova, G.V.; Zhigalova, T.V.; Yakovleva, O.S.; Tarakanov, I.G. LED crop illumination inside space greenhouses. Reach. Rev. Hum. Space Explor. 2017, 6, 11–24. [Google Scholar] [CrossRef]
- Whitelam, G.; Halliday, K. Light and Plant Development; Blackwell: Oxford, UK, 2007. [Google Scholar]
- Matsuda, R.; Ohashi-Kaneko, K.; Fujiwara, K.; Kurata, K. Effects of blue light deficiency on acclimation of light energy partitioning in PSII and CO2 assimilation capacity to high irradiance in spinach leaves. Plant Cell Physiol. 2008, 49, 664–670. [Google Scholar] [CrossRef] [Green Version]
- Yorio, N.C.; Goins, G.D.; Kagie, H.R.; Wheeler, R.M.; Sager, J.C. Improving spinach, radish, and lettuce growth under red light-emitting diodes (LEDs) with blue light supplementation. Hort. Sci. 2001, 36, 380–383. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, R.; Ohashi-Kaneko, K.; Fujiwara, K.; Goto, E.; Kurata, K. Photosynthetic characteristics of rice leaves grown under red light with or without supplemental blue light. Plant Cell Physiol. 2004, 45, 1870–1874. [Google Scholar] [CrossRef]
- Terashima, I.; Fujita, T.; Inoue, T.; Chow, W.S.; Oguchi, R. Green light drives leaf photosynthesis more efficiently than red light in strong white light: Revisiting the enigmatic question of why leaves are green. Plant Cell Physiol. 2009, 50, 684–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folta, K.M.; Maruhnich, S.A. Green light: A signal to slow down or stop. J. Exp. Bot. 2007, 58, 3099–3111. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.X.; Xu, Z.G.; Liu, X.Y.; Tang, C.M.; Wang, L.W.; Han, X.L. Effect of light intensity on the growth and leaf development of young tomato plants grown under a combination of red and blue light. Sci. Hort. 2013, 153, 50–55. [Google Scholar] [CrossRef]
- Lin, K.H.; Huang, M.Y.; Huang, W.D.; Hsu, M.H.; Yang, Z.W.; Yang, C.M. The effects of red blue and white light-emitting diodes on the growth development and edible quality of hydroponically grown lettuce (Lactuca sativa L Var Capitata). Sci. Hort. 2013, 150, 86–91. [Google Scholar] [CrossRef]
- Li, Q.; Kubota, C. Effects of supplemental light quality on growth and phytochemicals of baby leaf lettuce. Environ. Exp. Bot. 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Camejo, D.; Frutos, A.; Mestre, T.; Carmen Pinero, M.; Rivero, R.M.; Martinez, V. Artifcial light impacts the physical and nutritional quality of lettuce plants. Hortic. Environ. Biotechnol. 2020, 61, 69–82. [Google Scholar] [CrossRef]
- Kim, H.H.; Wheeler, R.; Sager, J.; Norikane, J. Photosynthesis of lettuce exposed to different short-term light qualities. Environ. Control Biol. 2005, 43, 113–119. [Google Scholar] [CrossRef] [Green Version]
- Kochetova, G.V.; Belyaeva, O.B.; Gorshkova, D.S.; Vlasova, T.A.; Bassarskaya, E.M.; Zhigalova, T.V.; Avercheva, O.V. Long-term acclimation of barley photosynthetic apparatus to narrow-band red and blue light. Photosythetica 2018, 56, 851–860. [Google Scholar] [CrossRef]
- Liere, K.; Weihe, A.; Börner, T. The transcription machineries of plant mitochondria and chloroplasts: Composition, function, and regulation. J. Plant Physiol. 2011, 168, 1345–1360. [Google Scholar] [CrossRef]
- Goins, G.D.; Yorio, N.C.; Sanwo, M.M.; Brown, C.S. Photomorphogenesis, photosynthesis, and seed yield of wheat plants grown under red light-emitting diodes (LEDs) with and without supplemental blue lighting. J. Exp. Bot. 1997, 48, 1407–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-H.; Tewari, R.K.; Hahn, E.-J.; Paek, K.-Y. Photon flux density and light quality induce changes in growth, stomatal development, photosynthesis and transpiration of Withania somnifera (L.) Dunal. plantlets. Plant Cell Tissue Organ Cult. 2007, 90, 141–151. [Google Scholar] [CrossRef]
- Shin, K.S.; Murthy, H.N.; Heo, J.W.; Hahn, E.J.; Paek, K.Y. The effect of light quality on the growth and development of in vitro cultured Doritaenopsis plants. Acta Physiol. Plant 2008, 30, 339–343. [Google Scholar] [CrossRef]
- Prikupets, L.B.; Boos, G.V.; Terekhov, V.G.; Tarakanov, I.G. Research into influence from different ranges of PAR radiation on efficiency and biochemical composition of green salad foliage biomass. Light Eng. 2018, 26, 38–47. [Google Scholar] [CrossRef]
- Prikupets, L.B.; Boos, G.V.; Terekhov, V.G.; Tarakanov, I.G. Optimization of lighting parameters of irradiation in light culture of lettuce plants using LED emitters. Light Eng. 2019, 27, 43–54. [Google Scholar] [CrossRef]
- Kreslavski, V.D.; Carpentier, R.; Klimov, V.V.; Allakhverdiev, S.I. Transduction mechanisms of photoreceptor signals in plant cells. J. Photochem. Photobiol. C Photochem. Rev. 2009, 10, 63–80. [Google Scholar] [CrossRef]
- Sipos, L.; Boros, L.F.; Csambalik, L.; Szekely, G.; Jung, A.; Balazs, L. Horticultural lighting system optimization: A review. Sci. Hortic. 2020, 273, 109631. [Google Scholar] [CrossRef]
- Litvin, A.G.; Currey, C.J.; Wilson, L.F. Effects of supplemental light source on basil, dill, and parsley growth, morphology, aroma, and flavor. J. Am. Soc. Hort. Sci. 2020, 145, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Shulgina, A.A.; Kalashnikova, E.A.; Tarakanov, I.G.; Kirakosyan, R.N.; Cherednichenko, M.Y.; Polivanova, O.B.; Baranova, E.N.; Khaliluev, M.R. Influence of Light Conditions and Medium Composition on Morphophysiological Characteristics of Stevia rebaudiana Bertoni In Vitro and In Vivo. Horticulturae 2021, 7, 195. [Google Scholar] [CrossRef]
- Voronin, V.; Ivlev, A.A.; Oskolkov, V.; Boettger, T. Intra-seasonal dynamics in metabolic processes of 13C/12C and 18O/16O in components of Scots pine twigs from southern Siberia interpreted with a conceptual framework based on the Carbon Metabolism Oscillatory Model. BMC Plant Biol. 2012, 30, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivlev, A.A.; Igamberdiev, A.Y.; Dubinsky, A.Yu. Isotopic composition of carbon metabolites and metabolic oscillations in the course of photosynthesis. Biophysics. 2010, 49 (Suppl. 1), 3–16. [Google Scholar]
- Igamberdiev, A.U.; Ivlev, A.A.; Bykova, N.V.; Threlkeld, Ch.; Lea, P.J.; Gardestrom, P. Decarboxylation of glycine contributes to carbon isotope fractionation in photosythetic organisms. Photosynth. Res. 2001, 67, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Roussel, M.R.; Ivlev, A.A.; Igamberdiev, A.Y. Oscillations of the internal CO2 concentration in tobacco leaves transferred to low CO2. J. Plant Physiol. 2007, 164, 1188–1196. [Google Scholar] [CrossRef] [PubMed]
- Ivlev, A.A. Oscillatory nature of metabolism and carbon isotope distribution in photosynthesizing cells. In Advances in Photosynthesis-Fundamental Aspects; Najafpour, M.M., Ed.; Intech Publishers: Balkan, Croatia, 2012; pp. 341–366. [Google Scholar]
- Mampholo, B.M.; Maboko, M.M.; Soundy, P.; Sivakumar, D. Phytochemicals and overall quality of leafy lettuce (Lactuca sativa L.) varieties grown in closed hydroponic system. J Food Qual. 2016, 39, 805–815. [Google Scholar] [CrossRef]
- Goltsev, V.N.; Kalaji, H.M.; Paunov, M.; Bąba, W.; Horaczek, T.; Mojski, J.; Kociel, H.; Allakhverdiev, S.I. Variable chlorophyll fluorescence and its use for assessing physiological condition of plant photosynthetic apparatus. Russ. J. Plant Physiol. 2016, 63, 869–893. [Google Scholar] [CrossRef]
- Shimazaki, K.; Doi, M.; Assmann, S.M.; Kinoshita, T. Light regulation of stomatal movement. Annu. Rev. Plant Biol. 2007, 58, 219–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gessler, A.; Tcherkez, G.; Peuke, A.D.; Ghashghaie, J.; Farquhar, G. Experimental evidence for diel variations of the carbon isotope composition in leaf, stem and phloem sap organic matter in Ricinus communis. Plant Cell Environ. 2008, 31, 941–953. [Google Scholar] [CrossRef] [PubMed]
- McCree, K.J. The action spectrum, absorptance and quantum yield of photosynthesis in crop plants. Agric. Meteorol. 1972, 9, 191–216. [Google Scholar] [CrossRef]
- Inada, K. Action spectra for photosynthesis in higher plants. Plant Cell Physiol. 1976, 17, 355–365. [Google Scholar]
- Evans, J.R. The dependence of quantum yield on wavelength and growth irradiance. Aust. J. Plant Physiol. 1987, 14, 69–79. [Google Scholar] [CrossRef]
- Walters, R.G. Towards an understanding of photosynthetic acclimation. J. Exp. Bot. 2005, 411, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Son, K.H.; Oh, M.M. Leaf shape, growth, and antioxidant phenolic compounds of two lettuce cultivars grown under various combinations of blue and red light-emitting diodes. Hort. Sci. 2013, 48, 988–995. [Google Scholar] [CrossRef]
- Wollaeger, H.M.; Runkle, E.S. Growth of impatiens, petunia, salvia, and tomato seedlings under blue, green, and red lightemitting diodes. Hort. Sci. 2014, 49, 734–740. [Google Scholar]
- Chen, X.L.; Li, Y.; Wang, L.C.; Guo, W.Z. Red and blue wavelengths affect the morphology, energy use efficiency and nutritional content of lettuce (Lactuca sativa L.). Sci. Rep. 2021, 11, 8374. [Google Scholar] [CrossRef] [PubMed]
- Lanoue, J.; Leonardos, E.D.; Grodzinski, B. Effects of light quality and intensity on diurnal patterns and rates of photo-assimilate translocation and transpiration in tomato leaves. Front. Plant Sci. 2018, 9, 756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivlev, A.A. Carbon isotope effect (13C/ 13C) in biological systems. Sep. Sci. Technol. 2001, 36, 1815–1910. [Google Scholar] [CrossRef]
- Ivlev, A.A. On the descretness of CO2 assimilation by C-plants in the light. Biofizika 1989, 34, 887–901. [Google Scholar]
- Sanchez-Bragado, R.; Serret, M.D.; Marimon, R.M.; Bort, J.; Araus, J.L. The Hydrogen Isotope Composition δ2H Reflects Plant Performance. Plant Physiol. 2019, 180, 793–812. [Google Scholar] [CrossRef] [Green Version]
- Voskresenskaya, N.P. Regulatory Role of Blue Light in Photosynthesis; Nauka: Moscow, Russia, 1982; pp. 203–220. [Google Scholar]
- Ivlev, A.A. On the flows of light and heavy carbon during photosynthesis and photorespiration coupling. Russ. J. Plant Physiol. 1993, 40, 752–758. [Google Scholar]
- Igamberdiev, A.U.; Mikkelsen, T.N.; Ambus, P.; Bauwe, H.; Lea, P.J.; Gardeström, P. Photorespiration contributes to stomatal regulation and carbon isotope fractionation: A study with barley, potato and Arabidopsis plants deficient in glycine decarboxylase. Photosynth. Res. 2004, 81, 139–152. [Google Scholar] [CrossRef]
- Ivlev, A.A. Carbon Isotope Heterogeneity in Photosynthesizing Biomass and Perspectives of Its Application in Biological Studies. In Photosynthesis: Functional Genomics, Physiological Processes and Environmental Benefits; Khan, N., Ed.; Nova Science Publishers: New York, NY, USA, 2015; pp. 73–104. [Google Scholar]
- Schmidt, H.-L.; Kexel, H.; Butzenlechner, M.; Schwarz, S.; Gleixner, G.; Thimet, S.; Werner, R.A.; Gensler, M. 2 Non-statistical isotope distribution in natural compounds: Mirror of their biosynthesis and key for their origin assignment. In Stable Isotopes in the Biosphere; Wada, E., Yoneyama, T., Minagawa, M., Ando, T., Fry, B.D., Eds.; Kyoto University Press: Kyoto, Japan, 1995; pp. 17–35. [Google Scholar]
- Dubinsky, A.Y.; Ivlev, A.A. Computational analysis of the possibility of the oscillatory dynamics in the processes of CO2 assimilation and photorespiration. BioSystems 2011, 103, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Ivlev, A.A.; Pichouzkin, V.I.; Tarakanov, I.G. Soil salinity effect on carbon isotope composition of plant biomass. Adv. Stud. Biol. 2013, 5, 223–234. [Google Scholar] [CrossRef]
- Cernusak, L.A.; Tcherkez, G.; Keitel, C.; Cornwell, W.K.; Santiago, L.S.; Knohl, A.; Barbour, M.M.; Williams, D.G.; Reich, P.B.; Ellsworth, D.S.; et al. Why are non-photosynthetic tissues generally 13C enriched compared with leaves in C3 plants? Review and synthesis of current hypotheses. Funct. Plant Biol. 2009, 36, 199–213. [Google Scholar] [CrossRef]
- Tarakanov, I.G.; Kosobryukhov, A.A.; Tovstyko, D.A.; Anisimov, A.A.; Shulgina, A.A.; Sleptsov, N.N.; Kalashnikova, E.A.; Vassilev, A.V.; Kirakosyan, R.N. Effects of Light Spectral Quality on the Micropropagated Raspberry Plants during Ex Vitro Adaptation. Plants 2021, 10, 2071. [Google Scholar] [CrossRef] [PubMed]
- Wingler, A.; Lea, P.J.; Quick, W.P.; Leegood, R.C. Photorespiration: Metabolic pathways and their role in stress protection. Philos. Trans. R. Soc. Lond B Biol. Sci. 2000, 355, 1517–1529. [Google Scholar] [CrossRef] [Green Version]
- Fernie, A.R.; Bauwe, H.; Eisenhut, M.; Florian, A.; Hanson, D.T.; Hagemann, M.; Keech, O.; Mielewczik, M.; Nikoloski, Z.; Peterhänsel, C.; et al. Perspectives on plant photorespiratory metabolism. Plant Biol. 2013, 15, 748–753. [Google Scholar] [CrossRef] [PubMed]
- Biel, K.J.; Fomina, I.R.; Yensen, R.G. Complex Biological Systems: Adaptation and Tolerance to Extreme Environments; Gorod: Krasnoyarsk, Russia, 1982; p. 334. [Google Scholar]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarakanov, I.G.; Tovstyko, D.A.; Lomakin, M.P.; Shmakov, A.S.; Sleptsov, N.N.; Shmarev, A.N.; Litvinskiy, V.A.; Ivlev, A.A. Effects of Light Spectral Quality on Photosynthetic Activity, Biomass Production, and Carbon Isotope Fractionation in Lettuce, Lactuca sativa L., Plants. Plants 2022, 11, 441. https://doi.org/10.3390/plants11030441
Tarakanov IG, Tovstyko DA, Lomakin MP, Shmakov AS, Sleptsov NN, Shmarev AN, Litvinskiy VA, Ivlev AA. Effects of Light Spectral Quality on Photosynthetic Activity, Biomass Production, and Carbon Isotope Fractionation in Lettuce, Lactuca sativa L., Plants. Plants. 2022; 11(3):441. https://doi.org/10.3390/plants11030441
Chicago/Turabian StyleTarakanov, Ivan G., Daria A. Tovstyko, Maxim P. Lomakin, Alexander S. Shmakov, Nikolay N. Sleptsov, Alexander N. Shmarev, Vladimir A. Litvinskiy, and Alexander A. Ivlev. 2022. "Effects of Light Spectral Quality on Photosynthetic Activity, Biomass Production, and Carbon Isotope Fractionation in Lettuce, Lactuca sativa L., Plants" Plants 11, no. 3: 441. https://doi.org/10.3390/plants11030441