
Transcriptome Analysis Revealed a Positive Role of Ethephon on Chlorophyll Metabolism of Zoysia japonica under Cold Stress

 , and
, and
Abstract
:1. Introduction
2. Results
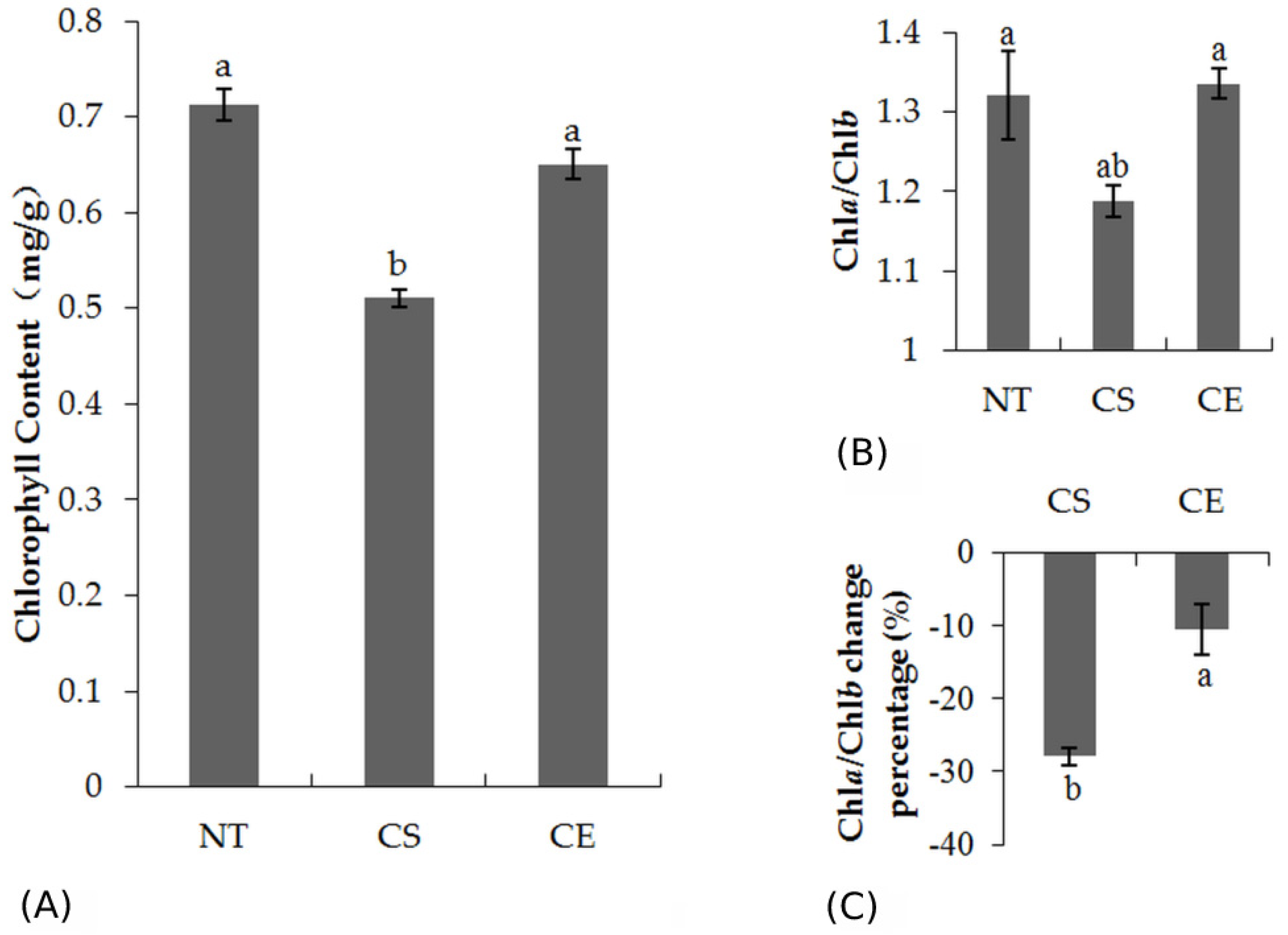
2.1. Chlorophyll Content in Leaves
2.2. Sequence Assembly
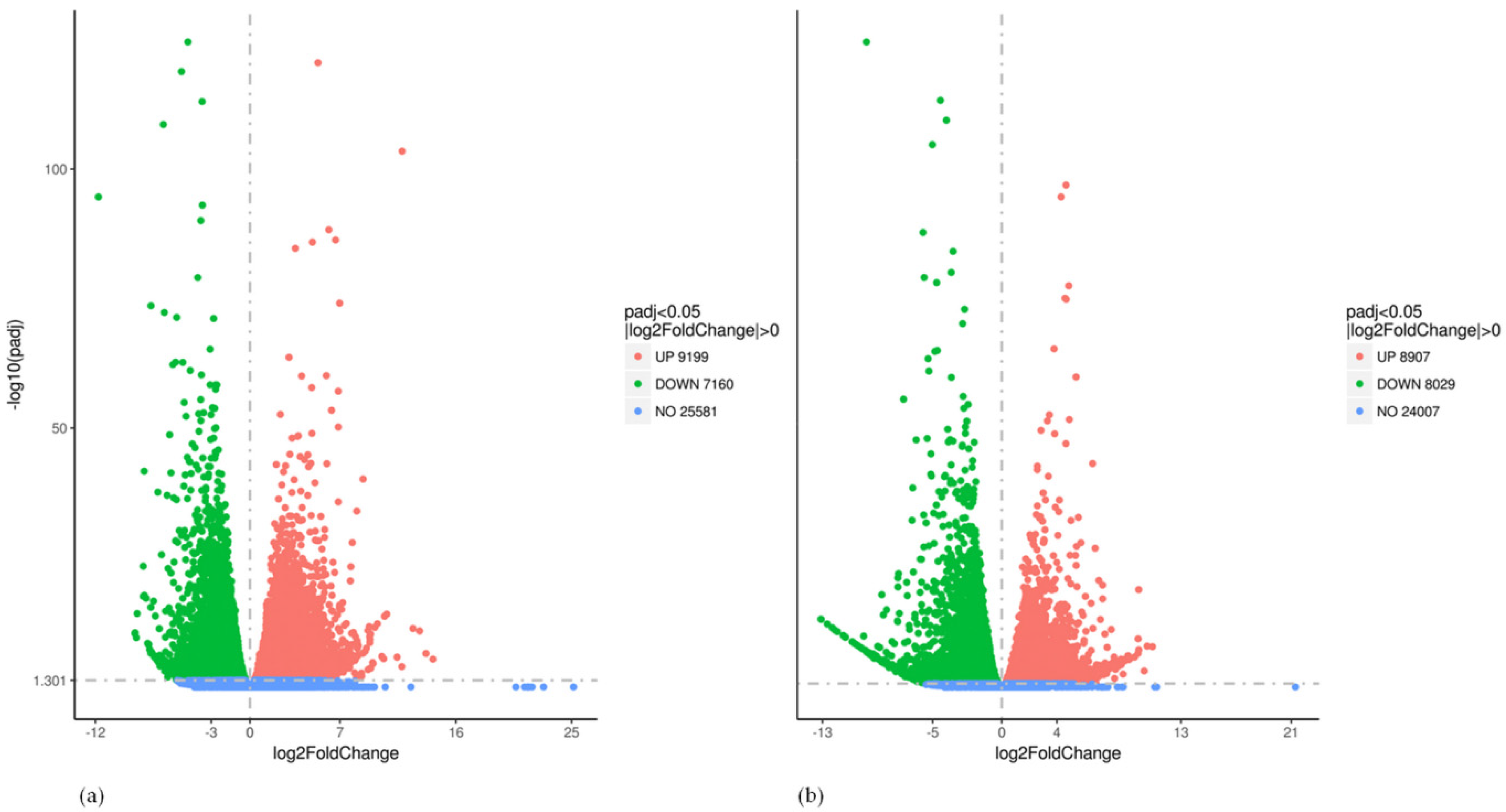
2.3. Global Gene Expression Analysis
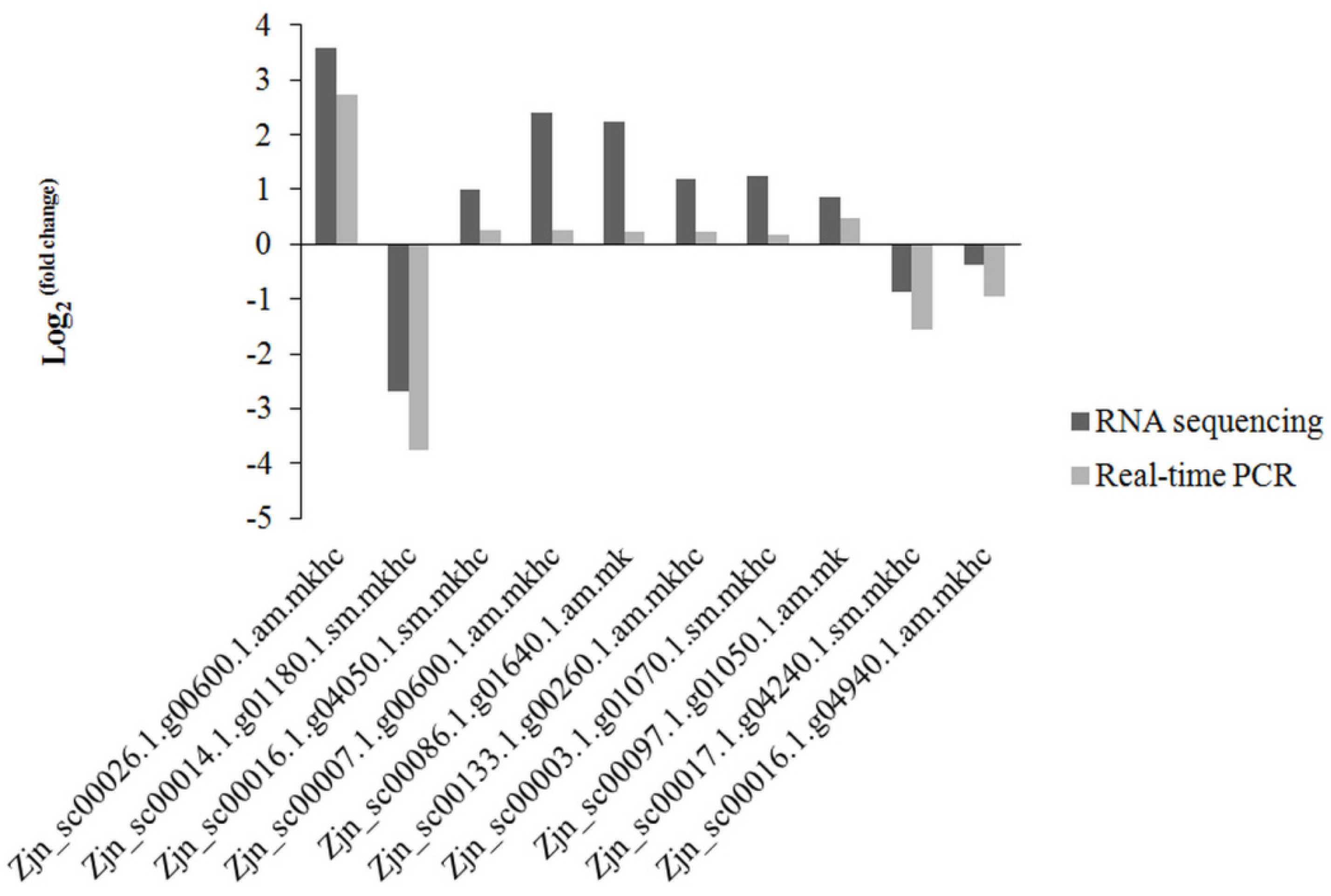
2.4. qRT-PCR Confirmation
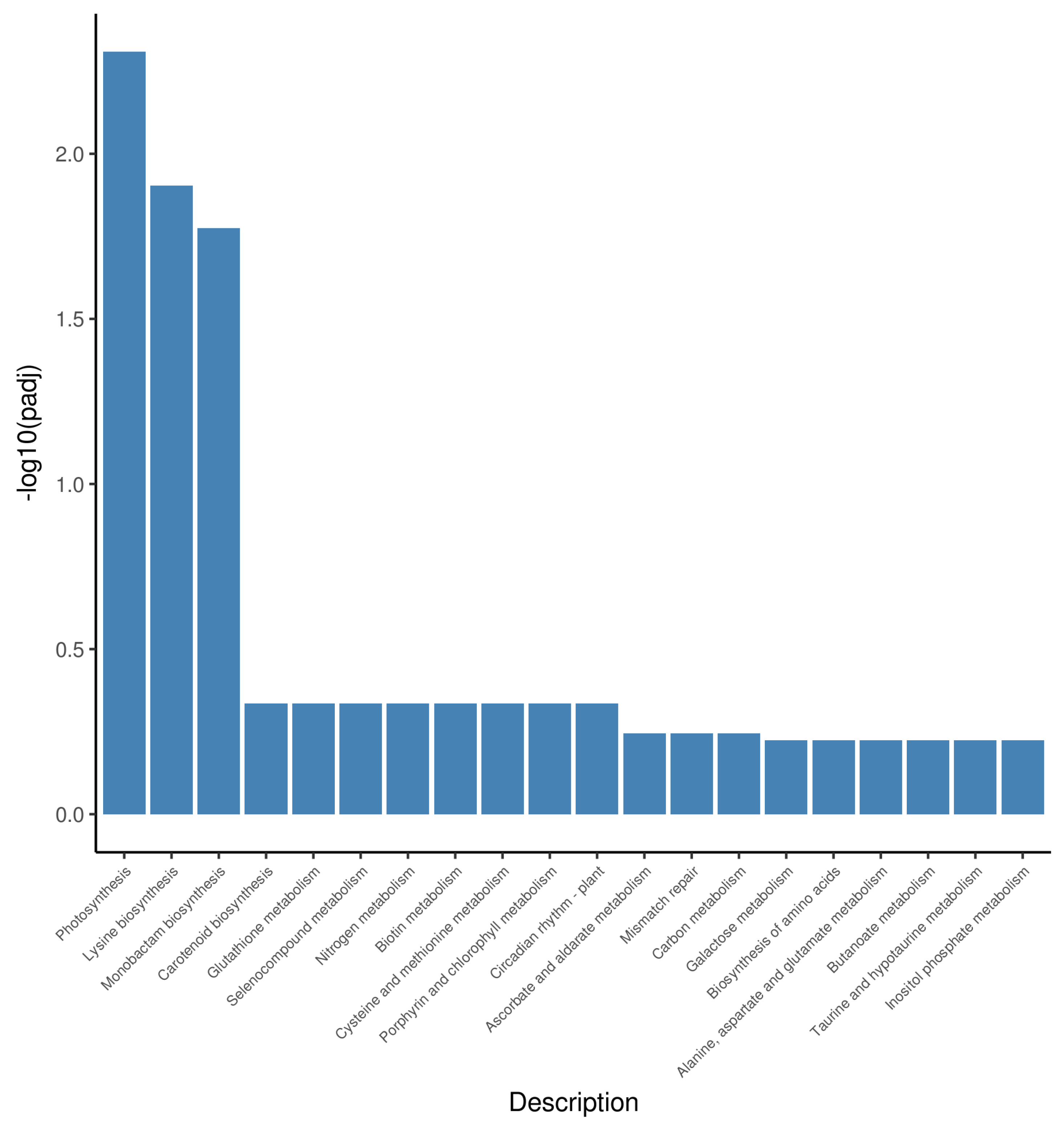
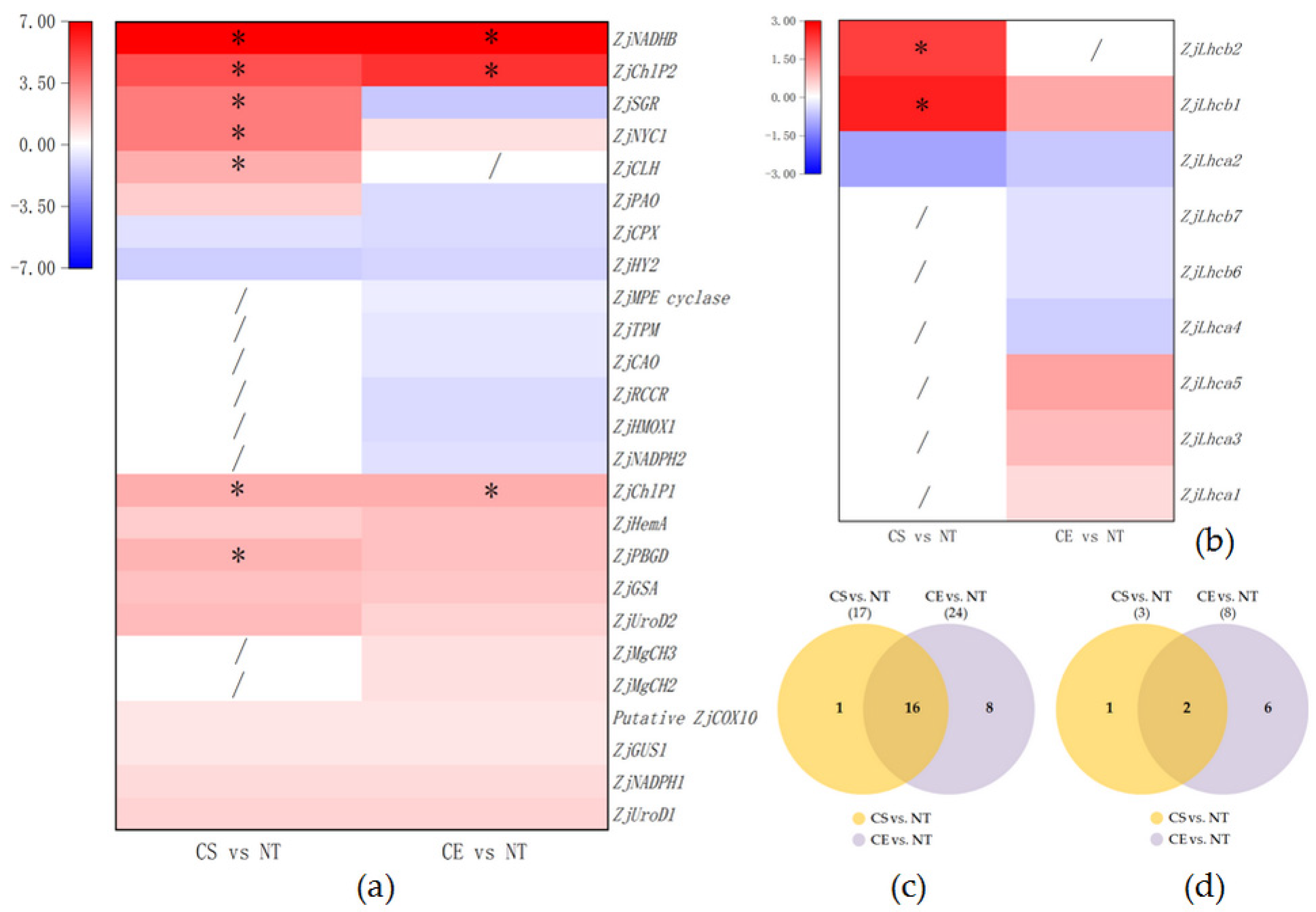
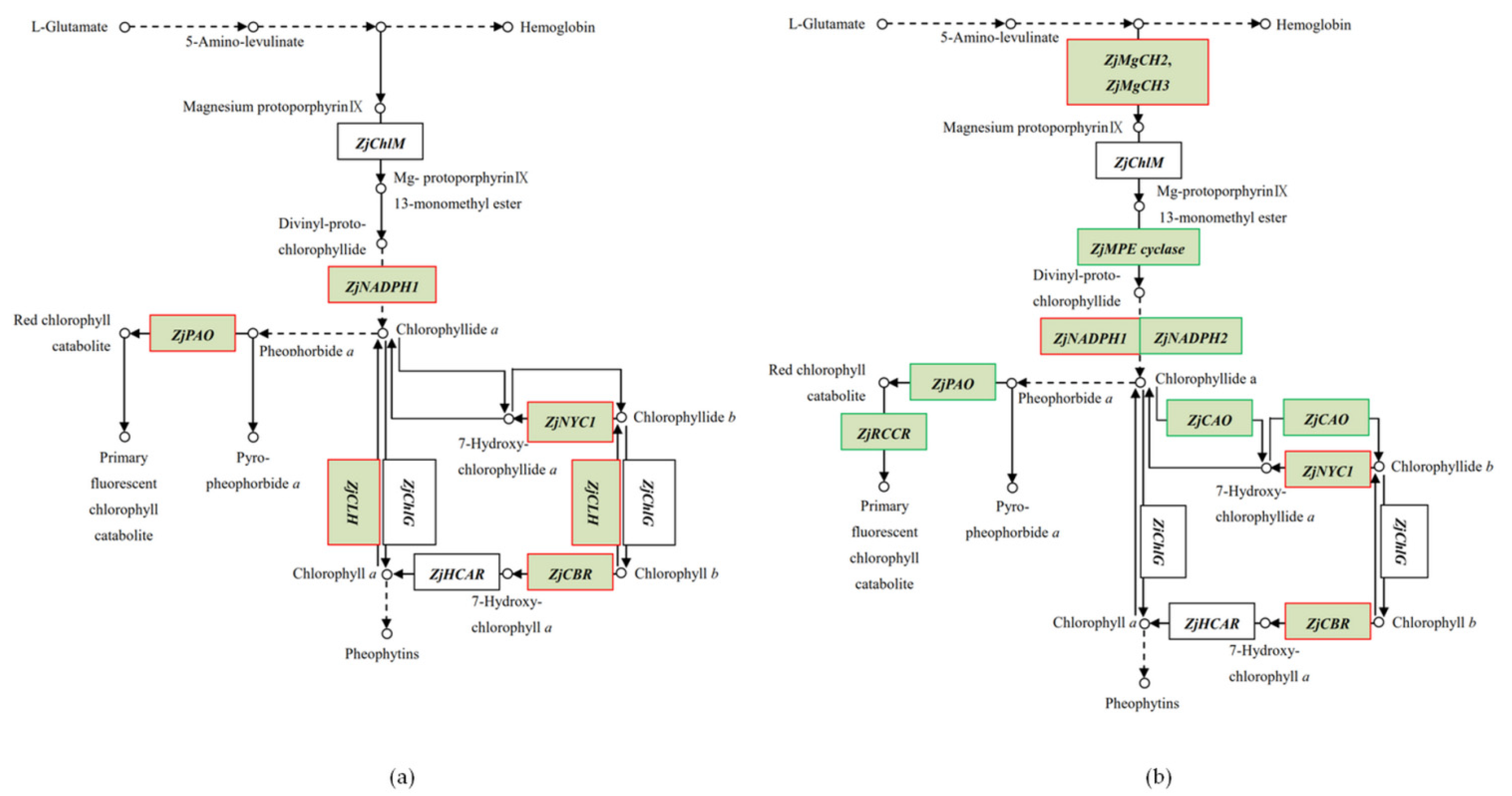
2.5. Differential Gene Expression Related to Photosynthesis-Antenna Proteins and Porphyrin and Chlorophyll Metabolism
3. Discussion
3.1. Gene Expression Analysis of Porphyrin and Chl Metabolism
3.2. Effect of Ethephon Application on Chl Content under Cold Stress
3.3. Gene Expression Analysis of Photosynthesis-Antenna Proteins
4. Materials and Methods
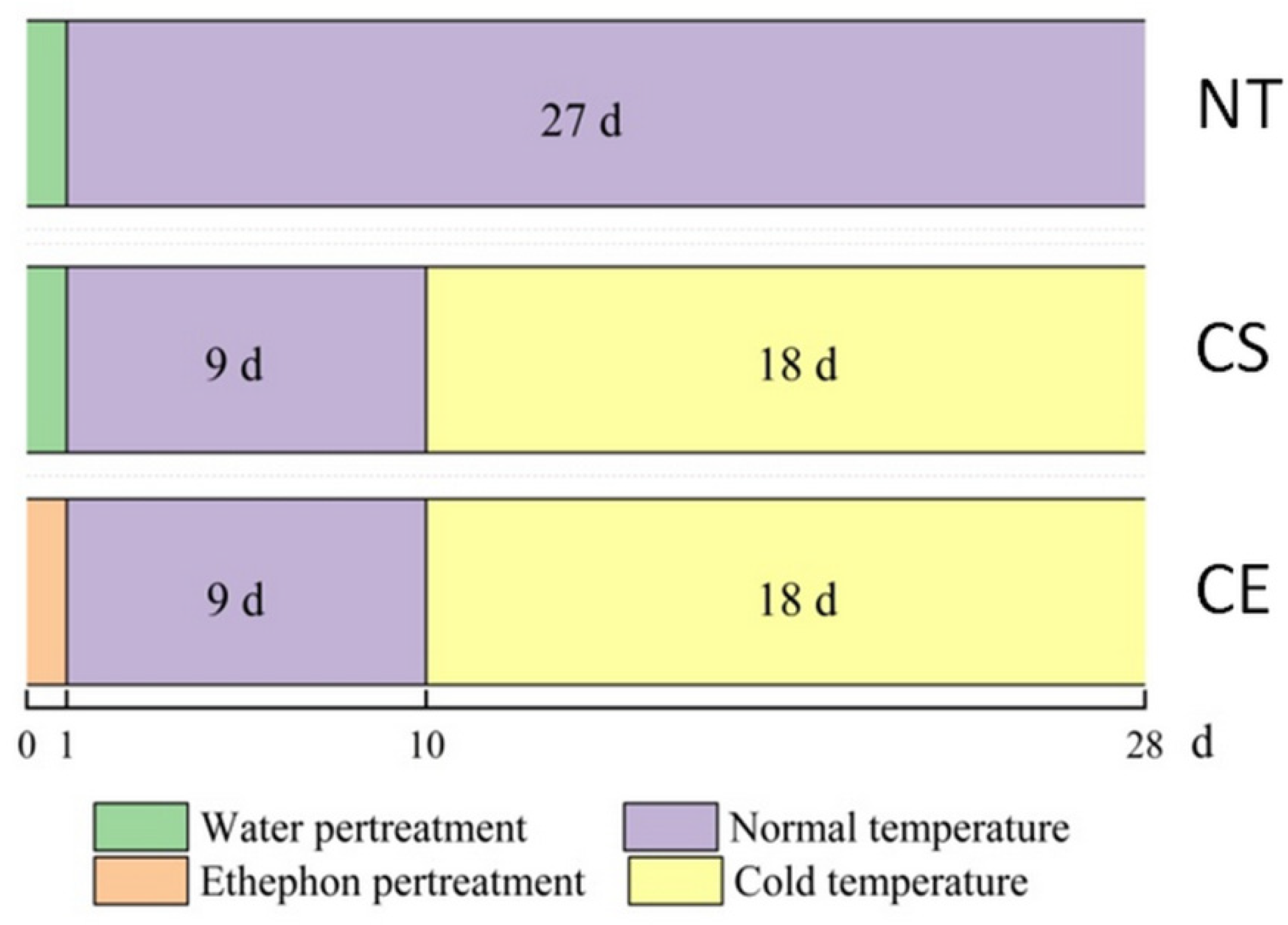
4.1. Plant Material and Growth Conditions
4.2. Determination of Chl Content in Leaves
4.3. Total RNA Extraction, RNA-Seq Library Construction, and RNA-Seq
4.4. RNA-Seq Data Processing and Assembly
4.5. DEG Analysis
4.6. Annotation Analysis
4.7. qRT-PCR Confirmation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, Y.; Zong, J.; Tan, Z.; Li, L.; Hu, B.; Chen, C.; Chen, J.; Liu, J. Systematic mining of salt-tolerant genes in halophyte-Zoysia matrella through cDNA expression library screening. Plant. Physiol. Biochem. 2015, 89, 44–52. [Google Scholar] [CrossRef]
- Teng, K.; Tan, P.; Xiao, G.; Han, L.; Chang, Z.; Chao, Y. Heterologous expression of a novel Zoysia japonica salt-induced glycine-rich RNA-binding protein gene, ZjGRP, caused salt sensitivity in Arabidopsis. Plant. Cell Rep. 2017, 36, 179–191. [Google Scholar] [CrossRef]
- Du, Y.; Yu, L.; Sun, J.; Lu, W.; Han, L. Study on Cold-resistance and Its Mechanism of Different Cultivars of Zoysia japonica Steud. Acta Agrestia Sin. 2008, 4, 347–352. [Google Scholar] [CrossRef]
- Teng, K.; Chang, Z.; Li, X.; Sun, X.; Liang, X.; Xu, L.; Chao, Y.; Han, L. Functional and RNA-Sequencing Analysis Revealed Expression of a Novel Stay-Green Gene from Zoysia japonica (ZjSGR) Caused Chlorophyll Degradation and Accelerated Senescence in Arabidopsis. Front. Plant. Sci. 2016, 7, 1894–1907. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Wu, Y.; Zeng, J.; He, L.; Zeng, Q. Chill-induced inhibition of photosynthesis was alleviated by 24-epibrassinolide pretreatment in cucumber during chilling and subsequent recovery. Photosynthetica 2010, 48, 537–544. [Google Scholar] [CrossRef]
- Palta, J.P.; Whitaker, B.D.; Weiss, L.S. Plasma membrane lipids associated with genetic variability in freezing tolerance and cold acclimation of solanum species. Plant. Physiol. 1993, 103, 793–803. [Google Scholar] [CrossRef] [Green Version]
- Orvar, B.L.; Sangwan, V.; Omann, F.; Dhindsa, R.S. Early steps in cold sensing by plant cells: The role of actin cytoskeleton and membrane fluidity. Plant. J. 2000, 23, 785–794. [Google Scholar] [CrossRef]
- Nobuhiro, S.; Ron, M. Reactive oxygen species and temperature stresses: A delicate balance between signaling and destruction. Physiol. Plant. 2006, 126, 45–51. [Google Scholar] [CrossRef]
- Hughes, M.A.; Dunn, M.A. The Effect of Temperature on Plant Growth and Development. Biotechnol. Genet. Eng. Rev. 2013, 8, 161–188. [Google Scholar] [CrossRef]
- Mohabbati, F.; Paknejad, F.; Vazan, S.; Habibi, D.; Tookallo, M.R.; Moradi, F. Protective effect of exogenous PGPRs on chlorophyll florescence and membrane integrity of rice seedlings under chilling stress. Res. J. Appl. Sci. Eng. Technol. 2013, 5, 146–153. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, K.; Ervin, E.H.; Waltz, C.; Murphy, T. Metabolic changes during cold acclimation and deacclimation in five bermudagrass varieties. I. Proline, total amino acid, protein, and dehydrin expression. Crop. Sci. 2011, 51, 847–853. [Google Scholar] [CrossRef]
- Ryoji, O.; Seiji, K. Reduction of chlorophyll fluorescence in zoysiagrasses at chilling and high temperatures with moderate light. Jpn. J. Grassl. Sci. 1995, 41, 31–36. [Google Scholar] [CrossRef]
- Ryoji, O.; Seiji, K. Changes in photosynthetic oxygen evolution and chlorophyll fluorescence in some cool season grasses and zoysiagrasses (Zoysia spp.) from autumn to winter. Soil Sci. Plant. Nutr. 1995, 41, 801–806. [Google Scholar] [CrossRef]
- Wei, S.; Du, Z.; Gao, F.; Ke, X.; Li, J.; Liu, J.; Zhou, Y. Global Transcriptome Profiles of ‘Meyer’ Zoysiagrass in Response to Cold Stress. PLoS ONE 2015, 10, e0131153. [Google Scholar] [CrossRef]
- He, J.; Liu, H.; Wang, Y.; Guo, J. Low Temperature and Photosynthesis of Plants. Plant. Physiol. Commun. 1986, 2, 1–6. [Google Scholar] [CrossRef]
- Ma, D.; Pang, J.; Huo, Z. Research Progress on low temperature tolerance of Cucumber. Tianjin Agric. Sci. 1997, 4, 3–10. [Google Scholar]
- Nagata, N.; Tanaka, R.; Tanaka, A. The major route for chlorophyll synthesis includes [3,8-divinyl]-chlorophyllide a reduction in Arabidopsis thaliana. Plant. Cell Physiol. 2007, 48, 1803–1808. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Han, Q.; Ding, C.; Huang, Y.; Liao, J.; Chen, T.; Feng, S.; Zhou, L.; Zhang, Z.; Chen, Y.; et al. Effect of Low Temperature on Chlorophyll Biosynthesis and Chloroplast Biogenesis of Rice Seedlings during Greening. Int. J. Mol. Sci. 2020, 21, 1390. [Google Scholar] [CrossRef] [Green Version]
- Pruzinská, A.; Tanner, G.; Anders, I.; Roca, M.; Hörtensteiner, S. Chlorophyll breakdown: Pheophorbide a oxygenase is a Rieske-type iron-sulfur protein, encoded by the accelerated cell death 1 gene. Proc. Natl. Acad. Sci. USA 2003, 100, 15259–15264. [Google Scholar] [CrossRef] [Green Version]
- Tanimoto, M.; Roberts, K.; Dolan, L. Ethylene is a positive regulator of root hair development in Arabidopsis thaliana. Plant. J. 1995, 8, 943–948. [Google Scholar] [CrossRef]
- Lynch, J.; Brown, K.M. Ethylene and plant responses to nutritional stress. Physiol. Plant 1997, 100, 613–619. [Google Scholar] [CrossRef]
- Sharp, R.E.; LeNoble, M.E. ABA, ethylene and the control of shoot and root growth under water stress. J. Exp. Bot. 2002, 53, 33–37. [Google Scholar] [CrossRef]
- Maruthasalam, S.; Ling, Y.S.; Murugan, L.; Wei, C.L.; Yi, L.L.; Ching, M.S.; Chih, W.Y.; Shu, H.H.; Yeong, K.; Chin, H.L. Forced flowering of pineapple (Ananas comosus cv. Tainon 17) in response to cold stress, ethephon and calcium carbide with or without activated charcoal. Plant Growth Regul. 2009, 60, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Han, L.; Xu, L. Effects of Ethephon Treatment on Plant Drought Tolerance. Acta Agrestia Sin. 2013, 21, 631–636. [Google Scholar] [CrossRef]
- Wang, W.; Wang, Z.; Ping, J.; Zhang, Y. Effect of Ethephon on Several Cold Hardiness Indexes of Carbernet Sauvignon. Sino-Overseas Grapevine Wine 2005, 5, 14–15. [Google Scholar] [CrossRef]
- Niu, H. Effects of DPCA, Paclobutrazol and ethephon on resistance of Larch Seedlings. Yunnan For. Sci. Technol. 1998, 3, 3–5. [Google Scholar] [CrossRef]
- Wei, D.; Li, Y.; Di, N.; Bi, Z.; Bu, Z. Effect of Ethephon on Cold Resistance of Banana (Musa AAA Cavendish subgroup) Seedlings. Chin. J. Trop. Crops 2009, 30, 1789–1792. [Google Scholar]
- Sone, C.; Sakagami, J.I. Physiological mechanism of chlorophyll breakdown for leaves under complete submergence in rice. Crop. Sci. 2017, 57, 2729–2738. [Google Scholar] [CrossRef]
- Liu, W. Application of Ethephon Impact Cold Tolerance of Zoysiagrass; Beijing Forestry University: Beijing, China, 2019. [Google Scholar]
- Zhang, Z. Effects of Ethephon on Chlorophyll Metabolic Pathway of Zoysia Japonica under Low Temperature Stress; Beijing Forestry University: Beijing, China, 2020. [Google Scholar]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Wilhelm, B.T.; Landry, J.R. RNA-Seq—Quantitative measurement of expression through massively parallel RNA-sequencing. Methods 2009, 48, 249–257. [Google Scholar] [CrossRef]
- Hu, X.; Makita, S.; Schelbert, S.; Sano, S.; Ochiai, M.; Tsuchiya, T.; Hasegawa, S.F.; Hörtensteiner, S.; Tanaka, A.; Tanaka, R. Reexamination of chlorophyllase function implies its involvement in defense against chewing herbivores. Plant. Physiol. 2015, 167, 660–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bushman, B.S.; Amundsen, K.L.; Warnke, S.E.; Robins, J.G.; Johnson, P.G. Transcriptome profiling of Kentucky bluegrass (Poa pratensis L.) accessions in response to salt stress. BMC Genom. 2016, 17, 48–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Shukla, V.; Merewitz, E.B. Transcriptome analysis of creeping bentgrass exposed to drought stress and polyamine treatment. PLoS ONE 2017, 12, e175848. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; An, C.; Guo, H.; Yang, X.; Chen, J.; Zong, J.; Li, J.; Liu, J. Physiological and transcriptomic analyses reveal the mechanisms underlying the salt tolerance of Zoysia japonica Steud. BMC Plant. Bio. 2020, 20, 114. [Google Scholar] [CrossRef]
- Voitsekhovskaja, O.V.; Tyutereva, E.V. Chlorophyll b in angiosperms: Functions in photosynthesis, signaling and ontogenetic regulation. J. Plant. Physiol. 2015, 189, 51–64. [Google Scholar] [CrossRef]
- Wang, G.; Chen, Z.; Zhang, Y.; Gong, G.; Wen, P. Effects of three plant growth substances on Chilling Resistance of potato. J. Huizhou Univ. 2018, 38, 21–28. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, W. Effects of chilling stress on contents of chlorophyll and malondialdehyde in tomato seedlings. Acta Agric. Shanghai 2012, 28, 74–77. [Google Scholar]
- Stenbaek, A.; Jensen, P.E. Redox Regulation of Chlorophyll Biosynthesis. Phytochemistry 2010, 71, 853–859. [Google Scholar] [CrossRef]
- Pontier, D.; Albrieux, C.; Joyard, J.; Lagrange, T.; Block, M.A. Knock-out of the Magnesium Protoporphyrin IX Methyltransferase Gene in Arabidopsis: Effects on chloroplast development and on chloroplast-to-nucleus signaling. J. Biol. Chem. 2007, 282, 2297–2304. [Google Scholar] [CrossRef] [Green Version]
- Shioi, Y.; Takamiya, K. Monovinyl and divinyl protochlorophyllide pools in etiolated tissues of higher plants. Plant Physiol. 1992, 100, 1291–1295. [Google Scholar] [CrossRef] [Green Version]
- Reinbothe, S.; Reinbothe, C.; Apel, K.; Lebedev, N. Evolution of Chlorophyll Biosynthesis -The Challenge to Survive Photooxidation. Cell 1996, 86, 703–705. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, A.; Ito, H.; Tanaka, R.; Tanaka, N.K.; Yoshida, K.; Okada, K. Chlorophyll a oxygenase (CAO) is involved in chlorophyll b formation from chlorophyll a. Proc. Natl. Acad. Sci. USA 1998, 95, 12719–12723. [Google Scholar] [CrossRef] [Green Version]
- Dinc, E.; Ceppi, M.G.; Toth, S.Z.; Bottka, S.; Schansker, G. The chla fluorescence intensity is remarkably insensitive to changes in the chlorophyll content of the leaf as long as the chla/b ratio remains unaffected. Biochim. Biophys. Acta 2012, 1817, 770–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.; Li, H.; Chen, L.; Lou, Y.; Amombo, E.; Fu, J. RNA-seq for gene identification and transcript profiling in relation to root growth of bermudagrass (Cynodon dactylon) under salinity stress. BMC Genom. 2015, 16, 575–586. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Li, M.; Chen, Y.; Wu, P.; Wu, G.; Jiang, H. Knockdown of OsPAO and OsRCCR1 cause different plant death phenotypes in rice. J. Plant. Physiol. 2011, 168, 1952–1959. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Guo, F.Q. Nitric oxide deficiency accelerates chlorophyll breakdown and stability loss of thylakoid membranes during dark-induced leaf senescence in Arabidopsis. PLoS ONE 2013, 8, e56345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.Y.; Yu, J.W.; Park, J.S.; Li, J.; Yoo, S.C.; Lee, N.Y.; Lee, S.K.; Jeong, S.W.; Seo, H.S.; Koh, H.J.; et al. The senescence-induced staygreen protein regulates chlorophyll degradation. Plant Cell 2007, 19, 1649–1664. [Google Scholar] [CrossRef] [Green Version]
- Armstead, I.; Donnison, I.; Aubry, S.; Harper, J.; Hörtensteiner, S.; James, C.; Mani, J.; Moffet, M.; Ougham, H.; Roberts, L.; et al. Cross-species identification of Mendel’s I locus. Science 2007, 315, 73. [Google Scholar] [CrossRef] [Green Version]
- María, E.G.; Sofía, A.M.; Pedro, M.C.; Gustavo, A.M. Expression of Stay-Green encoding gene (BoSGR) during postharvest senescence of broccoli. Postharvest Biol. Technol. 2014, 95, 88–94. [Google Scholar] [CrossRef]
- Caudle, K.L.; Johnson, L.C.; Baer, S.G.; Maricle, B.R. A comparison of seasonal foliar chlorophyll change among ecotypes and cultivars of Andropogon gerardii (Poaceae) by using nondestructive and destructive methods. Photosynthetica 2014, 52, 511–518. [Google Scholar] [CrossRef]
- Damkjær, J.T.; Kereïche, S.; Johnson, M.P.; Kovacs, L.; Kiss, A.Z.; Boekema, E.J.; Ruban, A.V.; Horton, P.; Jansson, S. The photosystem II Light-Harvesting protein lhcb3 affects the macrostructure of photosystem II and the rate of state transitions in arabidopsis. Plant Cell 2009, 21, 3245–3256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansson, S. A guide to the Lhc genes and their relatives in Arabidopsis. Trends Plant. Sci. 1999, 4, 236–240. [Google Scholar] [CrossRef]
- Klimmek, F.; Sjödin, A.; Noutsos, C.; Leister, D.; Jansson, S. Abundantly and rarely expressed Lhc protein genes exhibit distinct regulation patterns in plants. Plant. Physiol. 2006, 140, 793–804. [Google Scholar] [CrossRef] [Green Version]
- Goussias, C.; Boussac, A.; Rutherford, A.W. Photosystem II and photosynthetic oxidation of water: An overview. Philos. Trans. R. Soc. B: Biol. Sci. 2002, 357, 1369–1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawakami, K.; Shen, J.R. Purification of fully active and crystallizable photosystem II from thermophilic cyanobacteria. Methods Enzym. 2018, 613, 1–16. [Google Scholar] [CrossRef]
- Barber, J.; Andersson, B. Too much of a good thing: Light can be bad for photosynthesis. Trends Biochem Sci. 1992, 17, 61–66. [Google Scholar] [CrossRef]
- Chan, T.; Shimizu, Y.; Pospíšil, P.; Nijo, N.; Fujiwara, A.; Taninaka, Y.; Ishikawa, T.; Hori, H.; Nanba, D.; Imai, A.; et al. Quality control of photosystem II: Lipid peroxidation accelerates photoinhibition under excessive illumination. PLoS ONE 2012, 7, e52100. [Google Scholar] [CrossRef] [Green Version]
- Dekker, J.P.; Boekema, E.J. Supramolecular organization of thylakoid membrane proteins in green plants. Biochim. Biophys. Acta 2005, 1706, 12–39. [Google Scholar] [CrossRef] [Green Version]
- Drop, B.; Webber-Birungi, M.; Yadav, S.K.; Filipowicz-Szymanska, A.; Fusetti, F.; Boekema, E.J.; Croce, R. Light-harvesting complex II (LHCII) and its supramolecular organization in Chlamydomonas reinhardtii. Biochim. Biophys. Acta 2014, 1837, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Croce, R.; Canino, G.; Ros, F.; Bassi, R. Chromophore organization in the higher-plant photosystem II antenna protein CP26. Biochem 2002, 41, 7334–7343. [Google Scholar] [CrossRef]
- Passarini, F.; Wientjes, E.; Hienerwadel, R.; Croce, R. Molecular basis of light harvesting and photoprotection in CP24: Unique features of the most recent antenna complex. J. Biol. Chem. 2009, 284, 29536–29546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, X.; Li, M.; Wan, T.; Wang, L.; Jia, C.; Hou, Z.; Zhao, X.; Zhang, J.; Chang, W. Structural insights into energy regulation of light-harvesting complex CP29 from spinach. Nat. Struct. Mol. Biol. 2011, 18, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Deng, X.; Tang, X.; Tan, T.; Zhang, D.; Liu, B.; Lin, H. Involvement of Lhcb6 and Lhcb5 in Photosynthesis Regulation in Physcomitrella patens Response to Abiotic Stress. Int. J. Mol. Sci. 2019, 20, 3665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peers, G.; Niyogi, K.K. Pond scum genomics: The genomes of chlamydomonas and ostreococcus. Plant Cell 2008, 20, 502–507. [Google Scholar] [CrossRef] [Green Version]
- Nelson, N. Plant photosystem I-the most efficient Nano-Photochemical machine. J. Nanosci. Nanotechnol. 2009, 9, 1709–1713. [Google Scholar] [CrossRef]
- Croce, R.; van Amerongen, H. Light-harvesting in photosystem I. Photosynth Res. 2013, 116, 153–166. [Google Scholar] [CrossRef] [Green Version]
- Wientjes, E.; van Stokkum, I.H.; van Amerongen, H.; Croce, R. The role of the individual Lhcas in photosystem I excitation energy trapping. Biophys. J. 2011, 101, 745–754. [Google Scholar] [CrossRef] [Green Version]
- Bellafiore, S.; Barneche, F.; Peltier, G.; Rochaix, J. State transitions and light adaptation require chloroplast thylakoid protein kinase STN7. Nature 2005, 433, 892–895. [Google Scholar] [CrossRef]
- Allen, J.F. BOTANY: State transitions--a question of balance. Science 2003, 299, 1530–1532. [Google Scholar] [CrossRef]
- Kanervo, E.; Suorsa, M.; Aro, E.M. Functional flexibility and acclimation of the thylakoid membrane. Photochem. Photobiol. Sci. 2005, 4, 1072–1080. [Google Scholar] [CrossRef]
- Haldrup, A.; Jensen, P.E.; Lunde, C.; Scheller, H.V. Balance of power: A view of the mechanism of photosynthetic state transitions. Trends Plant. Sci. 2001, 6, 301–305. [Google Scholar] [CrossRef]
- Nawrocki, W.J.; Santabarbara, S.; Mosebach, L.; Wollman, F.A.; Rappaport, F. State transitions redistribute rather than dissipate energy between the two photosystems in Chlamydomonas. Nature Plants. 2016, 2, 16031. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.F. Protein phosphorylation in regulation of photosynthesis. Biochim. Biophys. Acta 1992, 1098, 275–335. [Google Scholar] [CrossRef]
- Sartory, D.P.; Grobbelaar, J.U. Extraction of chlorophyll a from freshwater phytoplankton for spectrophotometric analysis. Hydrobiologia 1984, 114, 177–187. [Google Scholar] [CrossRef]
- Chi, Y.; Ren, T.; Shi, X.; Jin, X.; Jin, P. Mechanism of nutrient removal enhancement in low carbon/nitrogen wastewater by a novel high-frequency micro-aeration/anoxic (HMOA) mode. Chemosphere 2021, 263, 128003. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323–338. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550–570. [Google Scholar] [CrossRef] [Green Version]
- Hadadi, N.; MohammadiPeyhani, H.; Miskovic, L.; Seijo, M.; Hatzimanikatis, V. Enzyme annotation for orphan and novel reactions using knowledge of substrate reactive sites. Proc. Natl. Acad. Sci. USA 2019, 116, 7298–7307. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Total Reads | Clean Reads | Clean Bases | Q30 | GC Content | Total Map |
|---|---|---|---|---|---|---|---|
| (Gbp) | (%) | (%) | (%) | ||||
| CE1 1 | 51696468 | 50530932 | 50530932 | 7.58 | 90.95 | 51.3 | 91.45 |
| CE2 | 67066970 | 65714084 | 65714084 | 9.86 | 92.08 | 51.23 | 92.35 |
| CE3 | 65392538 | 64083968 | 64083968 | 9.61 | 92.1 | 52.98 | 93.06 |
| CS1 2 | 61806546 | 60439330 | 60439330 | 9.07 | 90.91 | 53.48 | 92.11 |
| CS2 | 59147316 | 58002528 | 58002528 | 8.7 | 90.51 | 53.46 | 92.19 |
| CS3 | 61224860 | 60038194 | 60038194 | 9.01 | 91.67 | 52.75 | 92.79 |
| NT1 3 | 50204448 | 48478352 | 48478352 | 7.27 | 92.51 | 49.73 | 88.08 |
| NT2 | 46169382 | 44677484 | 44677484 | 6.7 | 92.1 | 50.06 | 88.44 |
| NT3 | 49753502 | 48029758 | 48029758 | 7.2 | 92.21 | 49.75 | 87.66 |
| Gene ID | Primer Sequence |
|---|---|
| Actin | F:5′-GGTGTTATGGTTGGGATGG-3′ |
| R:5′-CAGTGAGCAGGACAGGGTG-3′ | |
| Zjn_sc00026.1.g00600.1.am.mkhc | F:5′-GCAGCAAGAACGAATGAT-3′ |
| R:5′-CTGAAGAGTGGAAGGAGAA-3′ | |
| Zjn_sc00014.1.g01180.1.sm.mkhc | F: GATGACAGAGATGCCAAT |
| R: CGATGAATACACCAGACA | |
| Zjn_sc00016.1.g04050.1.sm.mkhc | F: GGCAAGTGGTATTAGTGAA |
| R: CAGTATGTGTTCCGTTGT | |
| Zjn_sc00007.1.g00600.1.am.mkhc | F:GGACCTTGGACAGCATCTT |
| R:CGGCGACGAAGTAGAGAAT | |
| Zjn_sc00086.1.g01640.1.am.mk | F:CACGGACCAAGGACTCAAG |
| R:CCAGCGTCAGTCACAAGA | |
| Zjn_sc00133.1.g00260.1.am.mkhc | F:5′-GAAGGACACAGGAGTTGATG-3′ |
| R:5′-CCATTACCAAGGCGTCTC-3′ | |
| Zjn_sc00003.1.g01070.1.sm.mkhc | F:5′-ATCCTTACACCACTTCCT-3′ |
| R:5′-CTCATCTCGCAACACATT-3′ | |
| Zjn_sc00097.1.g01050.1.am.mk | F:5′-CTACCACGCTCAATCCTAT-3′ |
| R:5′-GTCATCCTCCTCTTCATCTT-3′ | |
| Zjn_sc00017.1.g04240.1.sm.mkhc | F: GGTGGTCATTGTGGATAA |
| R: GGAGTCAGGTTCAGATAAG | |
| Zjn_sc00016.1.g04940.1.am.mkhc | F: GCAAGAATGGAACCTGTG |
| R: TCAGCAGCAATCTCATCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Zhang, Z.; Liu, W.; Li, L.; Han, L.; Xu, L.; Zhao, Y. Transcriptome Analysis Revealed a Positive Role of Ethephon on Chlorophyll Metabolism of Zoysia japonica under Cold Stress. Plants 2022, 11, 442. https://doi.org/10.3390/plants11030442
Zhang J, Zhang Z, Liu W, Li L, Han L, Xu L, Zhao Y. Transcriptome Analysis Revealed a Positive Role of Ethephon on Chlorophyll Metabolism of Zoysia japonica under Cold Stress. Plants. 2022; 11(3):442. https://doi.org/10.3390/plants11030442
Chicago/Turabian StyleZhang, Jiahang, Zhiwei Zhang, Wen Liu, Lijing Li, Liebao Han, Lixin Xu, and Yuhong Zhao. 2022. "Transcriptome Analysis Revealed a Positive Role of Ethephon on Chlorophyll Metabolism of Zoysia japonica under Cold Stress" Plants 11, no. 3: 442. https://doi.org/10.3390/plants11030442