
Antagonistic Activity of Fungal Strains against Fusarium Crown Rot

Abstract
:1. Introduction
2. Results
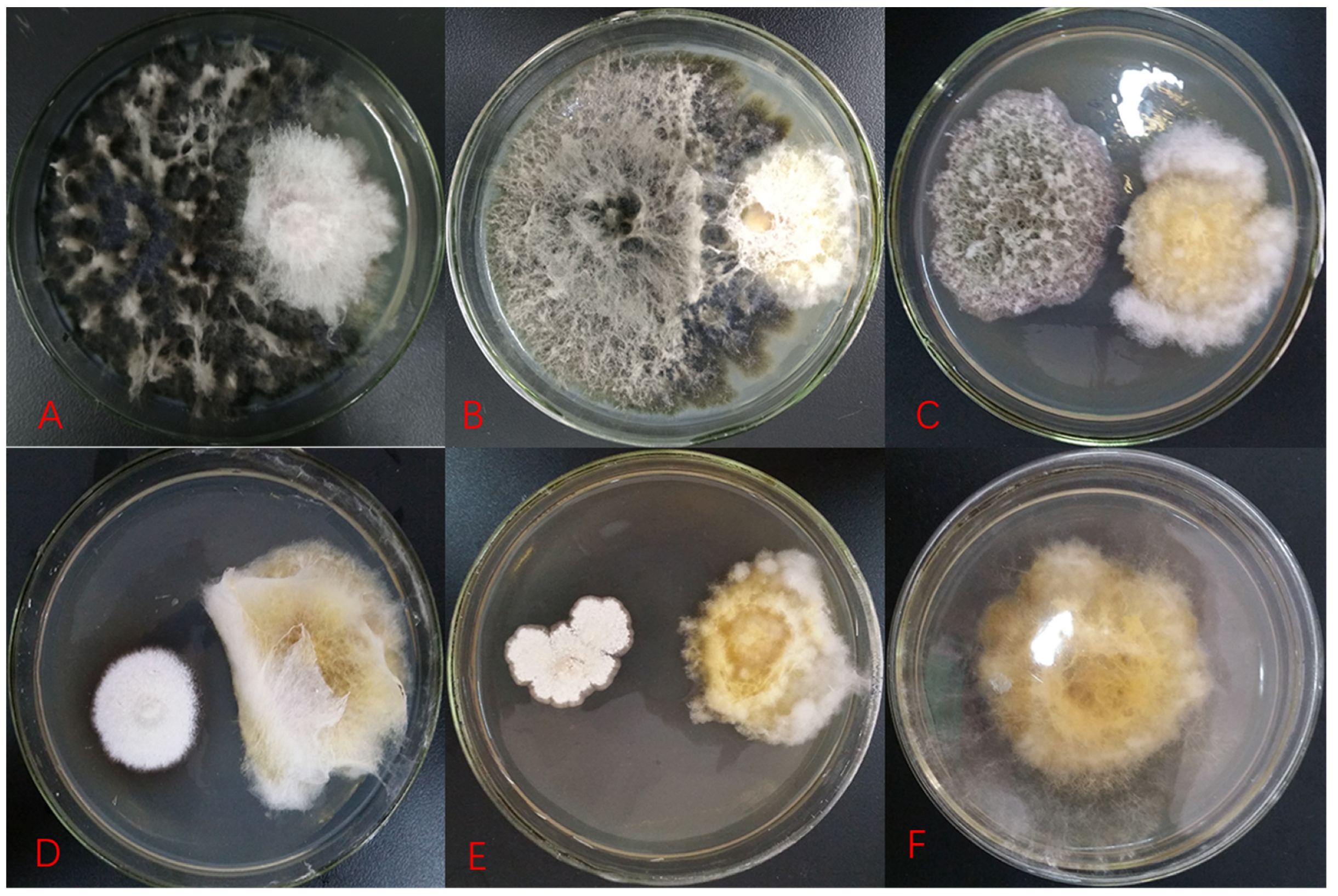
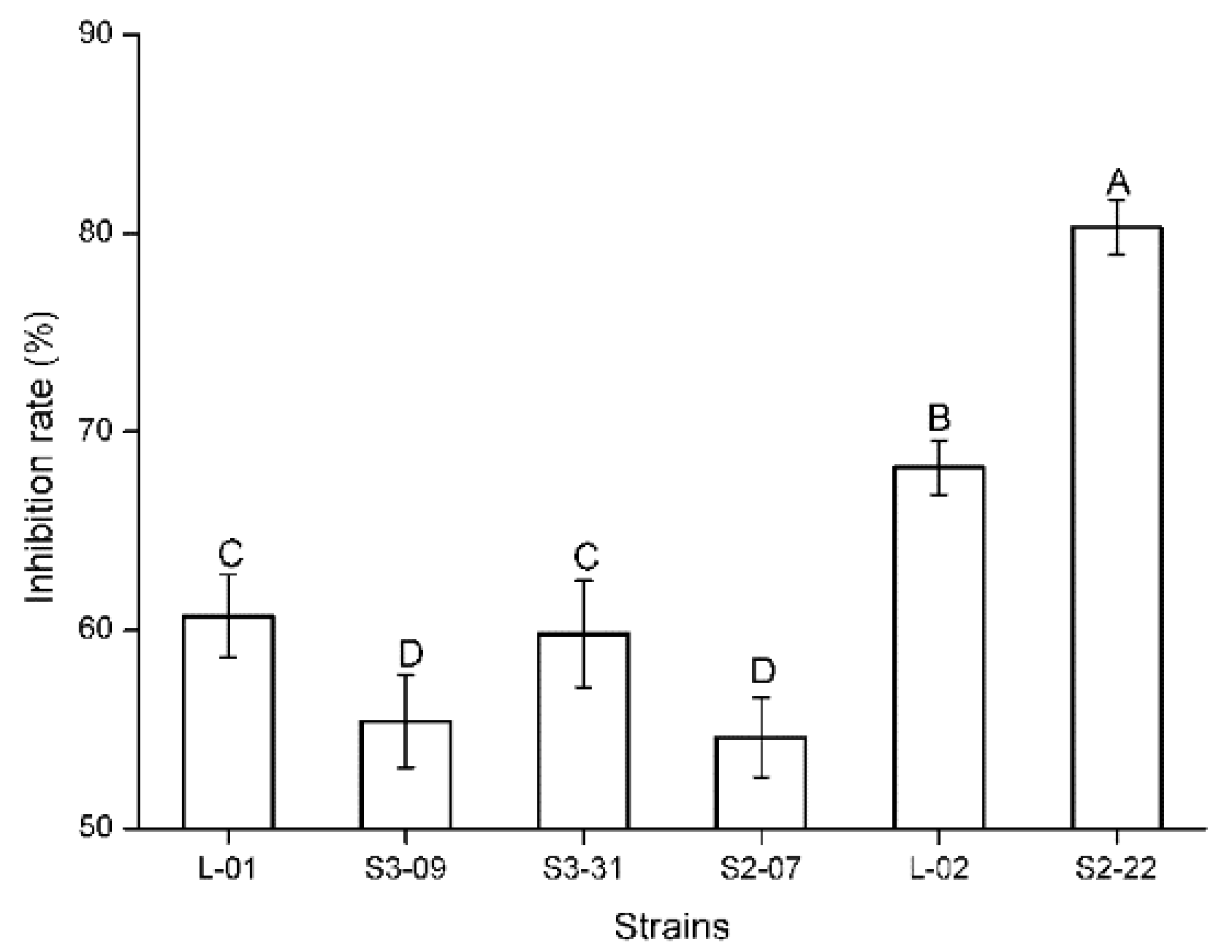
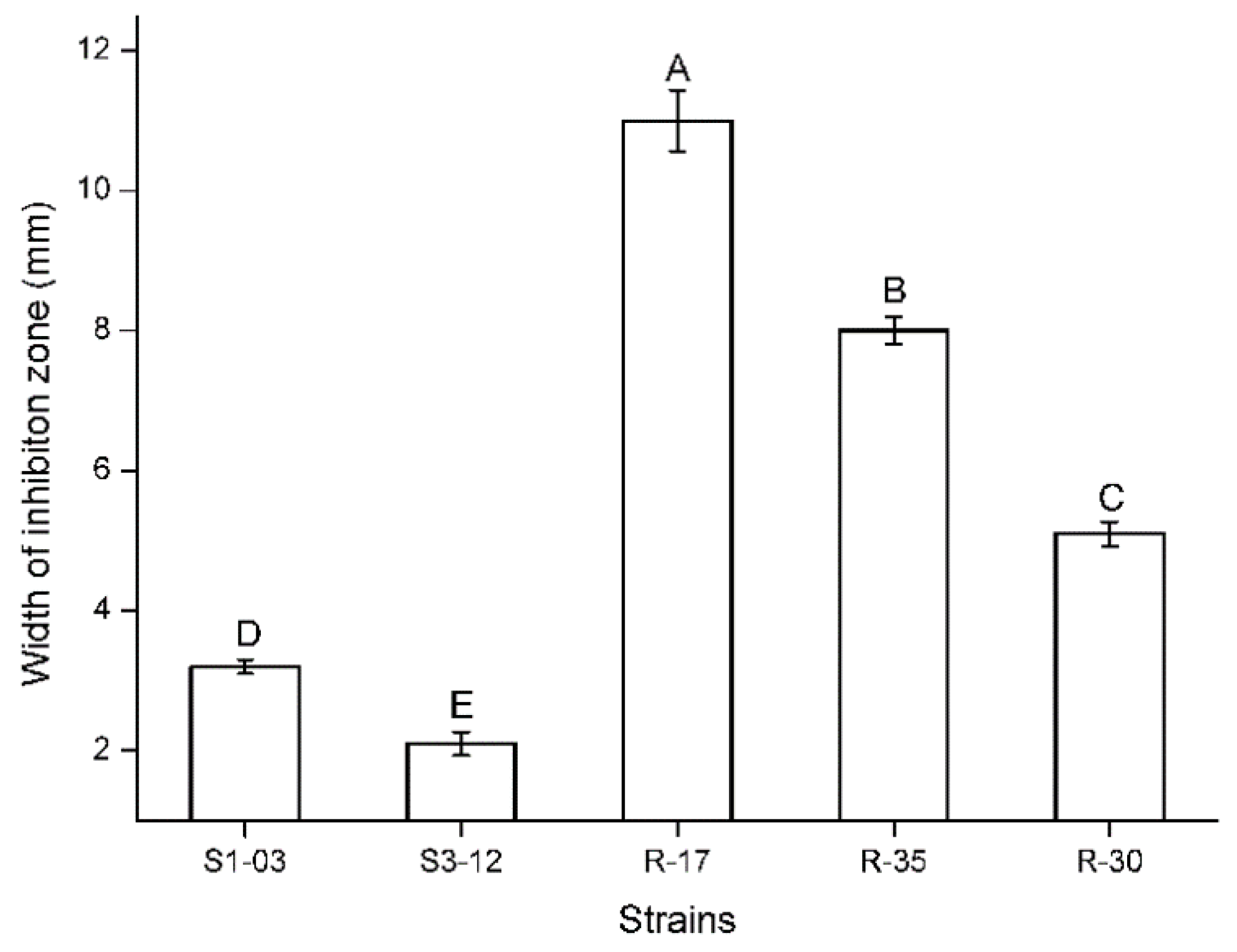
2.1. Antifungal Activity of Testing Fungal Isolates
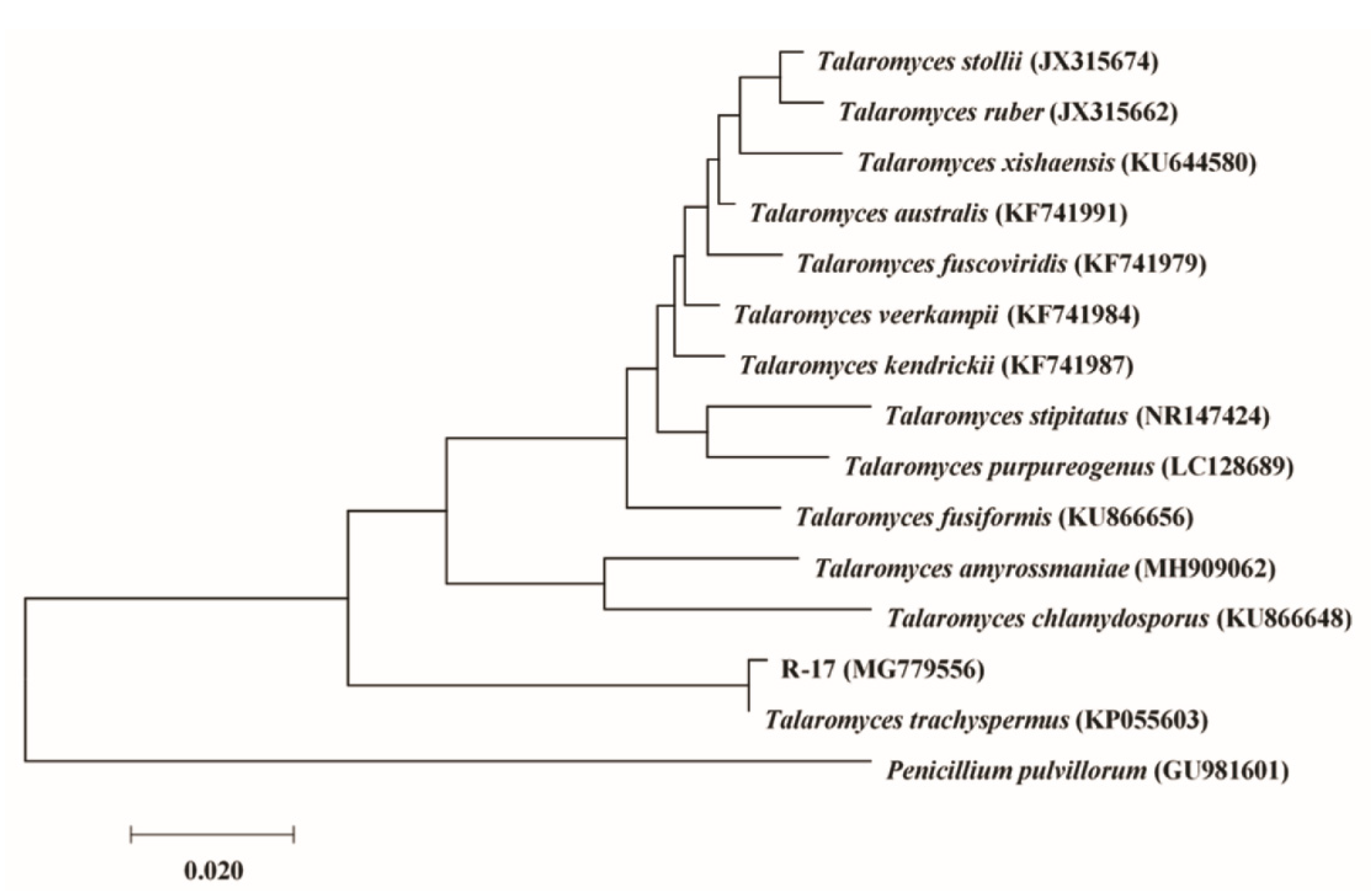
2.2. Identification of Isolates with Antifungal Activity
2.3. Detection of PKS-I, PKS-II, and NRPS Gene Sequences
3. Discussion
4. Materials and Methods
4.1. Strains
4.2. Determination of Antifungal Activity
4.3. Morphological Identification of Fungi with Antifungal Activity
4.4. DNA Extraction, PCR Amplification and ITS Sequencing
4.5. Detection of PKS-I, PKS-II and NRPS Gene Sequences
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, F.; Yang, G.; Wang, J.; Song, Y.; Liu, L.; Zhao, K.; Li, Y.; Han, Z. Spatial distribution of root and crown rot fungi associated with winter wheat in the North China Plain and its relationship with climate variables. Front. Microbiol. 2018, 9, 1054. [Google Scholar] [CrossRef]
- Kim, Y.T.; Monkhung, S.; Lee, Y.S.; Kim, K.Y. Effects of Lysobacter antibioticus HS124, an effective biocontrol agent against Fusarium graminearum, on crown rot disease and growth promotion of wheat. Can. J. Microbiol. 2019, 65, 904–912. [Google Scholar] [CrossRef]
- Li, H.L.; Yuan, H.X.; Fu, B.; Xing, X.P.; Sun, B.J.; Tang, W.H. First report of Fusarium pseudograminearum causing crown rot of wheat in Henan, China. Plant Dis. 2012, 96, 1065. [Google Scholar] [CrossRef]
- Moya-Elizondo, E.A. Fusarium crown rot disease: Biology, interactions, management and function as a possible sensor of global climate change. Cienc. E Investig. Agrar. 2013, 40, 235–252. [Google Scholar] [CrossRef]
- Dyer, A.T.; Johnston, R.H.; Hogg, A.C.; Johnston, J.A. Comparison of pathogenicity of the Fusarium crown rot (FCR) complex (F. culmorum, F. pseudograminearum and F. graminearum) on hard red spring and durum wheat. Eur. J. Plant Pathol. 2009, 125, 387–395. [Google Scholar] [CrossRef]
- Backhouse, D.; Abubakar, A.A.; Burgess, L.W.; Dennis, J.I.; Hollaway, G.J.; Wildermuth, G.B.; Wallwork, H.; Henry, F.J. Survey of Fusarium species associated with crown rot of wheat and barley in eastern Australia. Australas. Plant Path. 2004, 33, 255–261. [Google Scholar] [CrossRef] [Green Version]
- Poole, G.J.; Smiley, R.W.; Walker, C.; Huggins, D.; Rupp, R.; Abatzoglou, J.; Garland-Campbell, K.; Paulitz, T.C. Effect of climate on the distribution of Fusarium spp. causing crown rot of wheat in the Pacific Northwest of the United States. Phytopathology 2013, 103, 1130–1140. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; He, X.; Wang, S.; Ma, Q.; Sun, B.; Ding, S.; Chen, L.; Zhang, M.; Li, H. Diversity of the Fusarium pathogens associated with crown rot in the Huanghuai wheat-growing region of China. Environ. Microbiol. 2019, 21, 2740–2754. [Google Scholar] [CrossRef]
- Hollaway, G.J.; Evans, M.L.; Wallwork, H.; Dyson, C.B.; McKay, A.C. Yield loss in cereals, caused by Fusarium culmorum and F. pseudograminearum, is related to fungal DNA in soil prior to planting, rainfall, and cereal type. Plant Dis. 2013, 97, 977–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium toxins in cereals: Occurrence, legislation, factors promoting the appearance and their management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef] [Green Version]
- Daoubi, M.; Hernandez-Galan, R.; Benharref, A.; Collado, I.G. Screening study of lead compounds for natural product-based fungicides: Antifungal activity and biotransformation of 6alpha, 7alpha-dihydroxy-beta-himachalene by Botrytis cinerea. J. Agr. Food Chem. 2005, 53, 6673–6677. [Google Scholar] [CrossRef]
- Monkhung, S.; Kim, Y.; Lee, Y.; Cho, J.; Moon, J.; Kim, K. Isolation of antifungal compound and biocontrol potential of Lysobacter antibioticus HS124 against Fusarium crown rot of wheat. Korean J. Soil Sci. Fert. 2016, 49, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.X.; Sun, H.Y.; Shen, C.M.; Li, W.; Yu, H.S.; Chen, H.G. Survey of Fusarium spp. causing wheat crown rot in major winter wheat growing regions of China. Plant Dis. 2015, 99, 1610–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakhesar, D.P.S.; Backhouse, D.; Kristiansen, P. Nutritional constraints on displacement of Fusarium pseudograminearum from cereal straw by antagonists. Biol. Control 2010, 55, 241–247. [Google Scholar] [CrossRef]
- Moya-Elizondo, E.A.; Jacobsen, B.J. Integrated management of Fusarium crown rot of wheat using fungicide seed treatment, cultivar resistance, and induction of systemic acquired resistance (SAR). Biol. Control 2016, 92, 153–163. [Google Scholar] [CrossRef]
- Nosanchuk, J.D.; Casadevall, A. The contribution of melanin to microbial pathogenesis. Cell. Microbiol. 2003, 5, 203–223. [Google Scholar] [CrossRef] [PubMed]
- Calvo, A.M.; Wilson, R.A.; Bok, J.W.; Keller, N.P. Relationship between secondary metabolism and fungal development. Microbiol. Mol. Biol. Rev. 2002, 66, 447–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polak, A. Melanin as a virulence factor in pathogenic fungi. Mycoses 1990, 33, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, C. Ascocarps of Aspergillus and Penicillium. Mycologia 1955, 47, 669–687. [Google Scholar] [CrossRef]
- Samson, R.A.; Yilmaz, N.; Houbraken, J.; Spierenburg, H.; Seifert, K.A.; Peterson, S.W.; Varga, J.; Frisvad, J.C. Phylogeny and nomenclature of the genus Talaromyces and taxa accommodated in Penicillium subgenus Biverticillium. Stud. Mycol. 2011, 70, 159–183. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, N.; López-Quintero, C.A.; Vasco-Palacios, A.M.; Frisvad, J.C.; Theelen, B.; Boekhout, T.; Samson, R.A.; Houbraken, J. Four novel Talaromyces species isolated from leaf litter from Colombian Amazon rain forests. Mycol. Prog. 2016, 15, 1041–1056. [Google Scholar] [CrossRef] [Green Version]
- Su, L.; Niu, Y. Multilocus phylogenetic analysis of Talaromyces species isolated from cucurbit plants in China and description of two new species, T. cucurbitiradicus and T. endophyticus. Mycologia 2018, 110, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Komai, S.; Hosoe, T.; Itabashi, T.; Nozawa, K.; Yaguchi, T.; Fukushima, K.; Kawai, K. New penicillide derivatives isolated from Penicillium simplicissimum. J. Nat. Med. 2006, 60, 185–190. [Google Scholar] [CrossRef]
- Shiozawa, H.; Takahashi, M.; Takatsu, T.; Kinoshita, T.; Tanzawa, K.; Hosoya, T.; Furuya, K.; Takahashi, S.; Furihata, K.; Seto, H. Trachyspic acid, a new metabolite produced by Talaromyces trachyspermus, that inhibits tumor cell heparanase: Taxonomy of the producing strain, fermentation, isolation, structural elucidation, and biological activity. J. Antibiot. 1995, 48, 357–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dethoup, T.; Kumla, D.; Kijjoa, A. Mycocidal activity of crude extracts of marine-derived beneficial fungi against plant pathogenic fungi. J. Biopest. 2015, 8, 107–115. [Google Scholar]
- Hemmati, R.; Gholizadeh, R. Talaromyces trachyspermus a potential biocontrol agent for branched broom rape (Orobanche ramosa). Australas. Plant Path. 2019, 48, 217–219. [Google Scholar] [CrossRef]
- Sahu, S.; Prakash, A.; Shende, K. Talaromyces trachyspermus, an endophyte from Withania somnifera with plant growth promoting attributes. Environ. Sustain. 2019, 2, 13–21. [Google Scholar] [CrossRef]
- Kampapongsa, D.; Kaewkla, O. Biodiversity of endophytic actinobacteria from jasmine rice (Oryza sativa L. KDML 105) grown in Roi-Et Province, Thailand and their antimicrobial activity against rice pathogens. Ann. Microbiol. 2016, 66, 587–595. [Google Scholar] [CrossRef]
- Qin, S.; Li, J.; Chen, H.; Zhao, G.; Zhu, W.; Jiang, C.; Xu, L.; Li, W. Isolation, diversity, and antimicrobial activity of rare actinobacteria from medicinal plants of tropical rain forests in Xishuangbanna, China. Appl. Environ. Microb. 2009, 75, 6176–6186. [Google Scholar] [CrossRef] [Green Version]
- Ansari, M.Z.; Yadav, G.; Gokhale, R.S.; Mohanty, D. NRPS-PKS: A knowledge-based resource for analysis of NRPS/PKS megasynthases. Nucleic Acids Res. 2004, 32, W405–W413. [Google Scholar] [CrossRef]
- Zhao, K.; Penttinen, P.; Guan, T.; Xiao, J.; Chen, Q.; Xu, J.; Lindström, K.; Zhang, L.; Zhang, X.; Strobel, G.A. The diversity and anti-microbial activity of endophytic actinomycetes isolated from medicinal plants in Panxi Plateau, China. Curr. Microbiol. 2011, 62, 182–190. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Haydock, S.F.; Mironenko, T.; Spiteller, D.; Li, Y.; Spencer, J.B. The neomycin biosynthetic gene cluster of Streptomyces fradiae NCIMB 8233: Characterisation of an aminotransferase involved in the formation of 2-deoxystreptamine. Org. Biomol. Chem. 2005, 3, 1410–1419. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, J.; Yang, L.; Zhang, L.; Jiang, D.; Chen, W.; Li, G. Diversity and biocontrol potential of endophytic fungi in Brassica napus. Biol. Control 2014, 72, 98–108. [Google Scholar] [CrossRef]
- Wei, J.C. Handbook for Fungal Identification; Shanghai Science and Technology Press: Shanghai, China, 1979. [Google Scholar]
- Kim, W.K.; Mauthe, W.; Hausner, G.; Klassen, G.R. Isolation of high molecular weight DNA and double-stranded RNAs from fungi. Can. J. Plant Sci. 1990, 68, 1898–1902. [Google Scholar] [CrossRef]
- Silva-Hughes, A.F.; Wedge, D.E.; Cantrell, C.L.; Carvalho, C.R.; Pan, Z.; Moraes, R.M.; Madoxx, V.L.; Rosa, L.H. Diversity and antifungal activity of the endophytic fungi associated with the native medicinal cactus Opuntia humifusa (Cactaceae) from the United States. Microbiol. Res. 2015, 175, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Kusari, P.; Kusari, S.; Spiteller, M.; Kayser, O. Endophytic fungi harbored in Cannabis sativa L.: Diversity and potential as biocontrol agents against host plant-specific phytopathogens. Fungal Divers. 2013, 60, 137–151. [Google Scholar] [CrossRef]
- Yuan, Z.L.; Zhang, C.L.; Lin, F.C.; Kubicek, C.P. Identity, diversity, and molecular phylogeny of the endophytic mycobiota in the roots of rare wild rice (Oryza granulate) from a nature reserve in Yunnan, China. Appl. Environ. Microb. 2010, 76, 1642–1652. [Google Scholar] [CrossRef] [Green Version]
- Ayuso-Sacido, A.; Genilloud, O. New PCR primers for the screening of NRPS and PKS-I systems in actinomycetes: Detection and distribution of these biosynthetic gene sequences in major taxonomic groups. Microb. Ecol. 2005, 49, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Metsa-Ketela, M.; Salo, V.; Halo, L.; Hautala, A.; Hakala, J.; Mantsala, P.; Ylihonko, K. An efficient approach for screening minimal PKS genes from Streptomyces. Fems. Microbiol. Lett. 1999, 180, 1–6. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Number | Antagonism | Most Closely Related Strain (Accession Number) | GenBank Accession Number | Maximum Identity % | Presence of Gene PKS-I PKS-II NRPS | ||
|---|---|---|---|---|---|---|---|
| L-01 | MC(60.7) | Alternaria alternata (MT134991) | MG779581 | 100 | − | + | − |
| S3-09 | MC(55.4) | Alternaria alternata (KJ526174) | MG779560 | 100 | − | + | − |
| S3-31 | MC(59.8) | Alternaria alternata (MG214868) | MG779590 | 100 | − | + | − |
| S2-07 | MC(54.6) | Alternaria tenuissima (MH790256) | MG779609 | 99 | + | + | − |
| L-02 | MC(68.2) | Botryosphaeria dothidea (KY788303) | MG779587 | 99 | − | + | + |
| S2-22 | MC(80.3) | Botryosphaeria dothidea (KF293775) | MG779603 | 99 | − | + | + |
| S1-03 | AB (3.2) | Botryosphaeria dothidea (KP183180) | MG779610 | 99 | − | + | + |
| S3-12 | AB (2.1) | Phoma moricola (KF293794) | MG779600 | 99 | − | + | − |
| R-17 | AB (11.0) | Talaromyces trachyspermus (KP055603) | MG779556 | 99 | − | − | − |
| R-35 | AB (8.0) | Talaromyces trachyspermus (KJ482651) | MG779553 | 99 | − | − | − |
| R-30 | AB (5.1) | Talaromyces trachyspermus (MK271298) | MG779552 | 99 | − | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Hou, D.; Xu, J.; Wang, K.; Hu, Z. Antagonistic Activity of Fungal Strains against Fusarium Crown Rot. Plants 2022, 11, 255. https://doi.org/10.3390/plants11030255
Zhao X, Hou D, Xu J, Wang K, Hu Z. Antagonistic Activity of Fungal Strains against Fusarium Crown Rot. Plants. 2022; 11(3):255. https://doi.org/10.3390/plants11030255
Chicago/Turabian StyleZhao, Xingli, Dianyun Hou, Jiaqi Xu, Kaixuan Wang, and Zhenjie Hu. 2022. "Antagonistic Activity of Fungal Strains against Fusarium Crown Rot" Plants 11, no. 3: 255. https://doi.org/10.3390/plants11030255