
Irrigation Effect on Yield, Skin Blemishes, Phellem Formation, and Total Phenolics of Red Potatoes

 ,
,
Abstract
:1. Introduction
2. Results
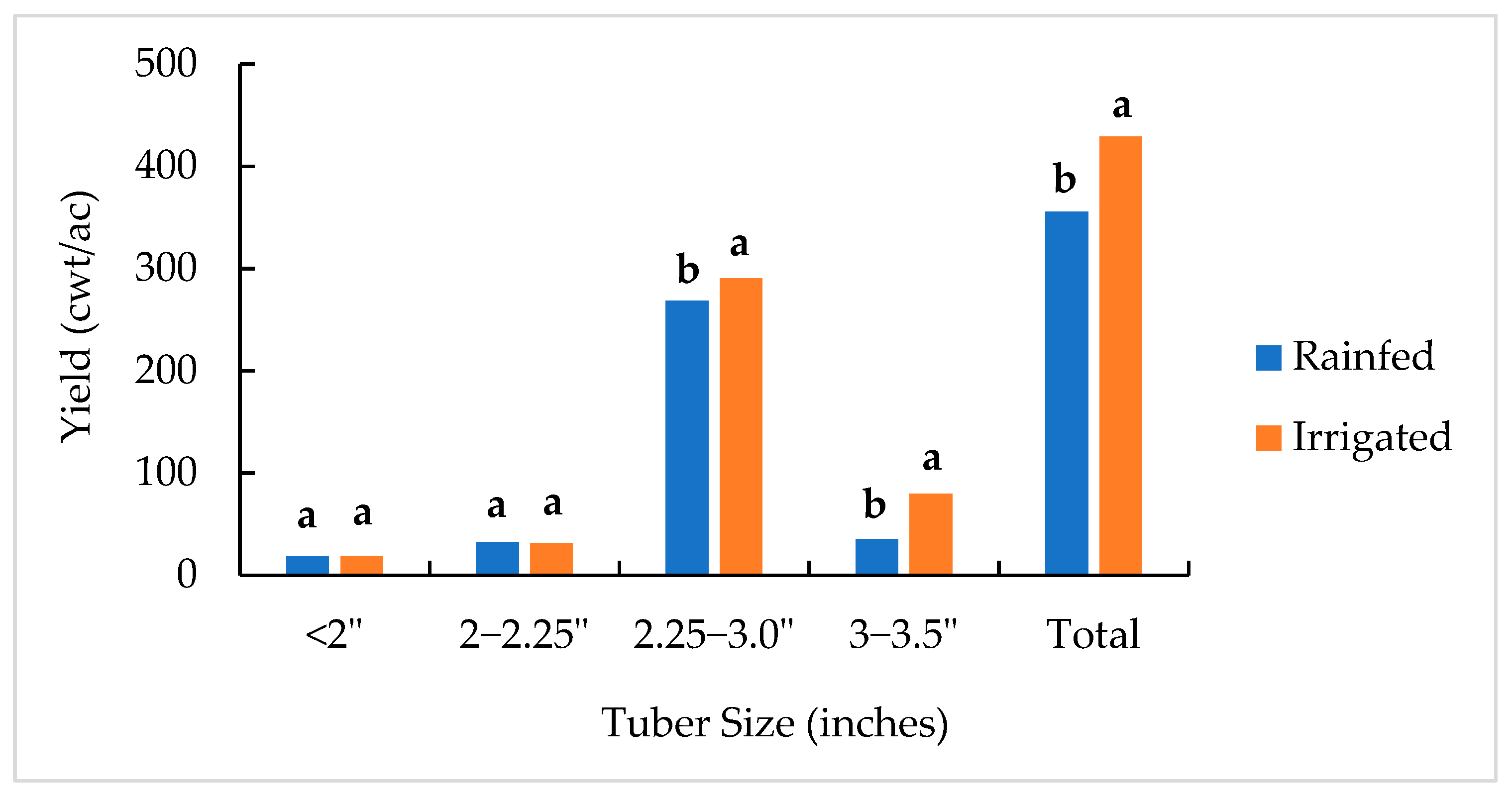
2.1. Total Yield
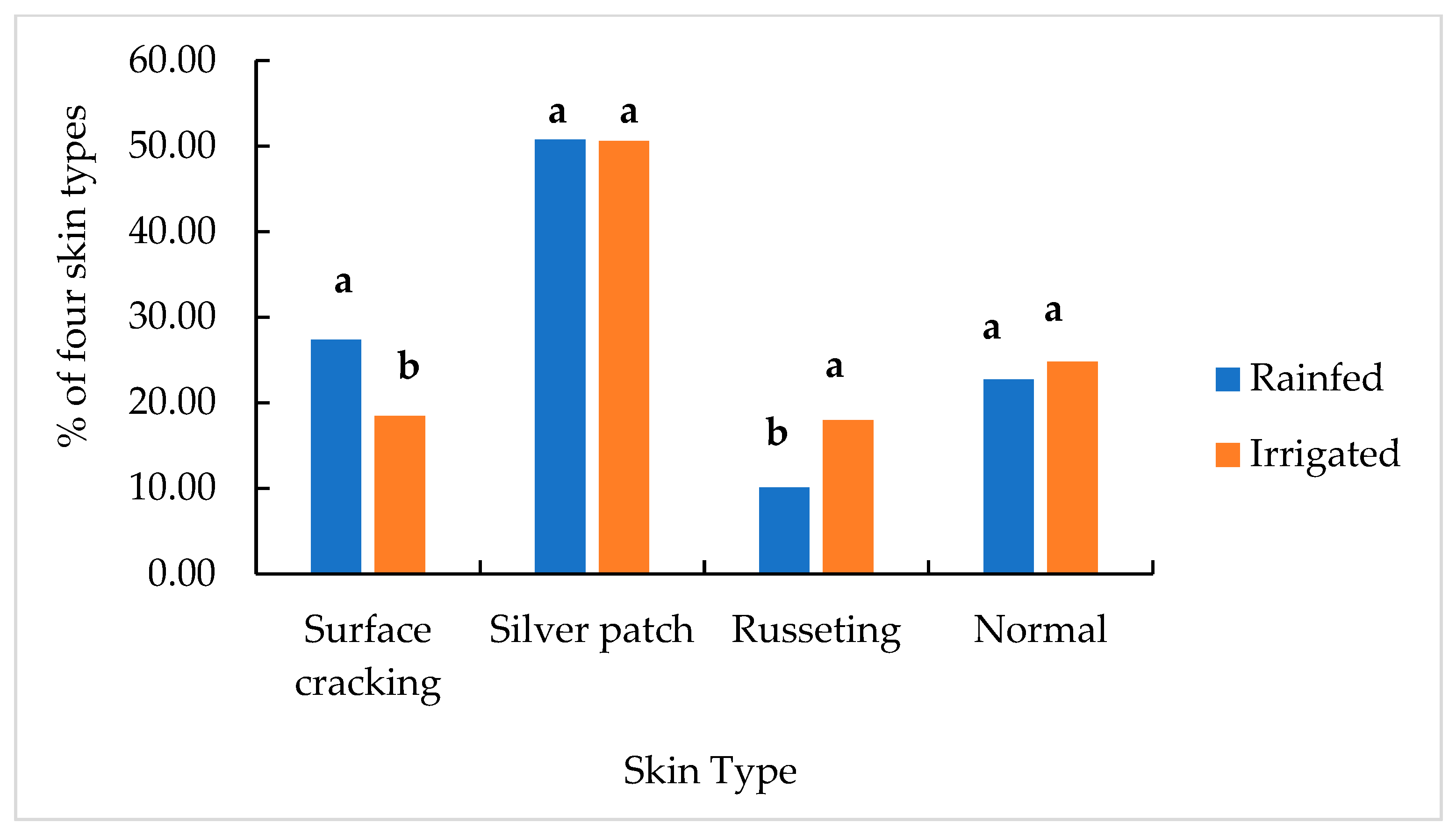
2.2. Skin Blemishes
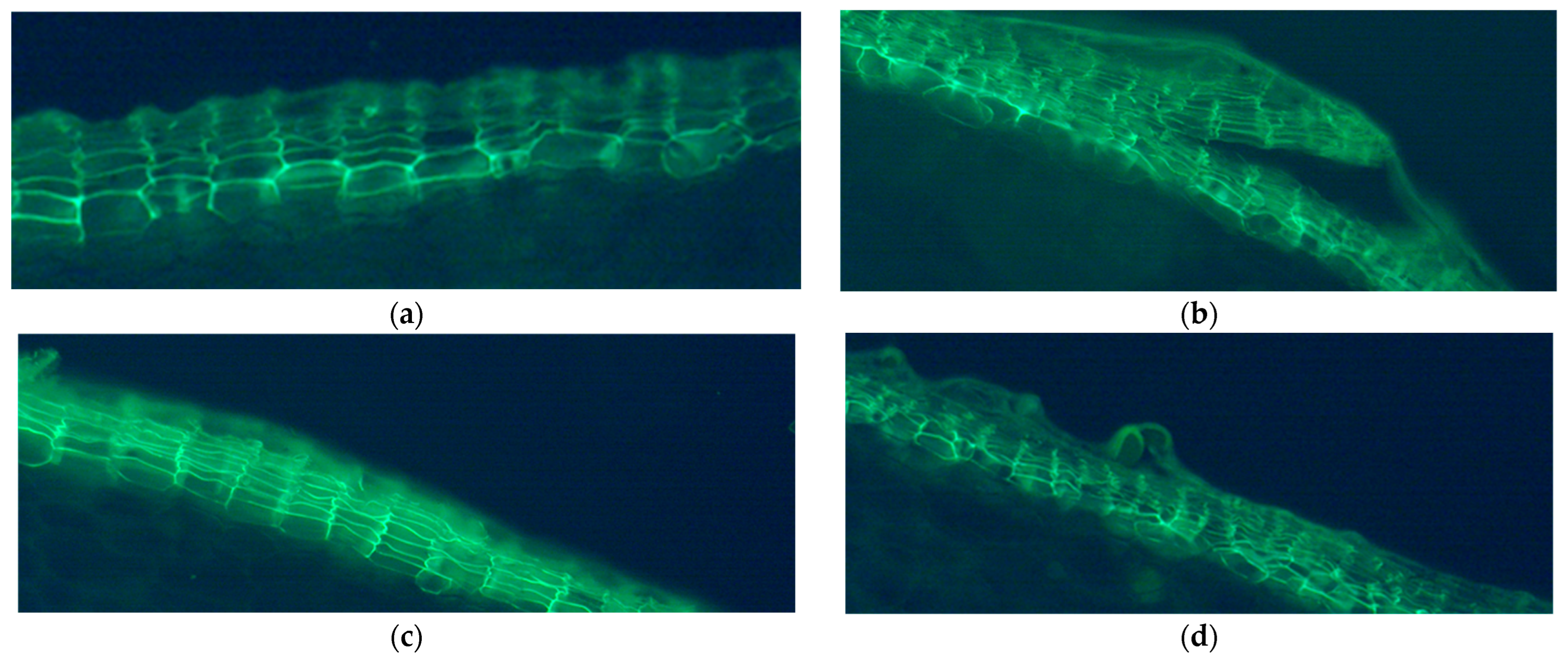
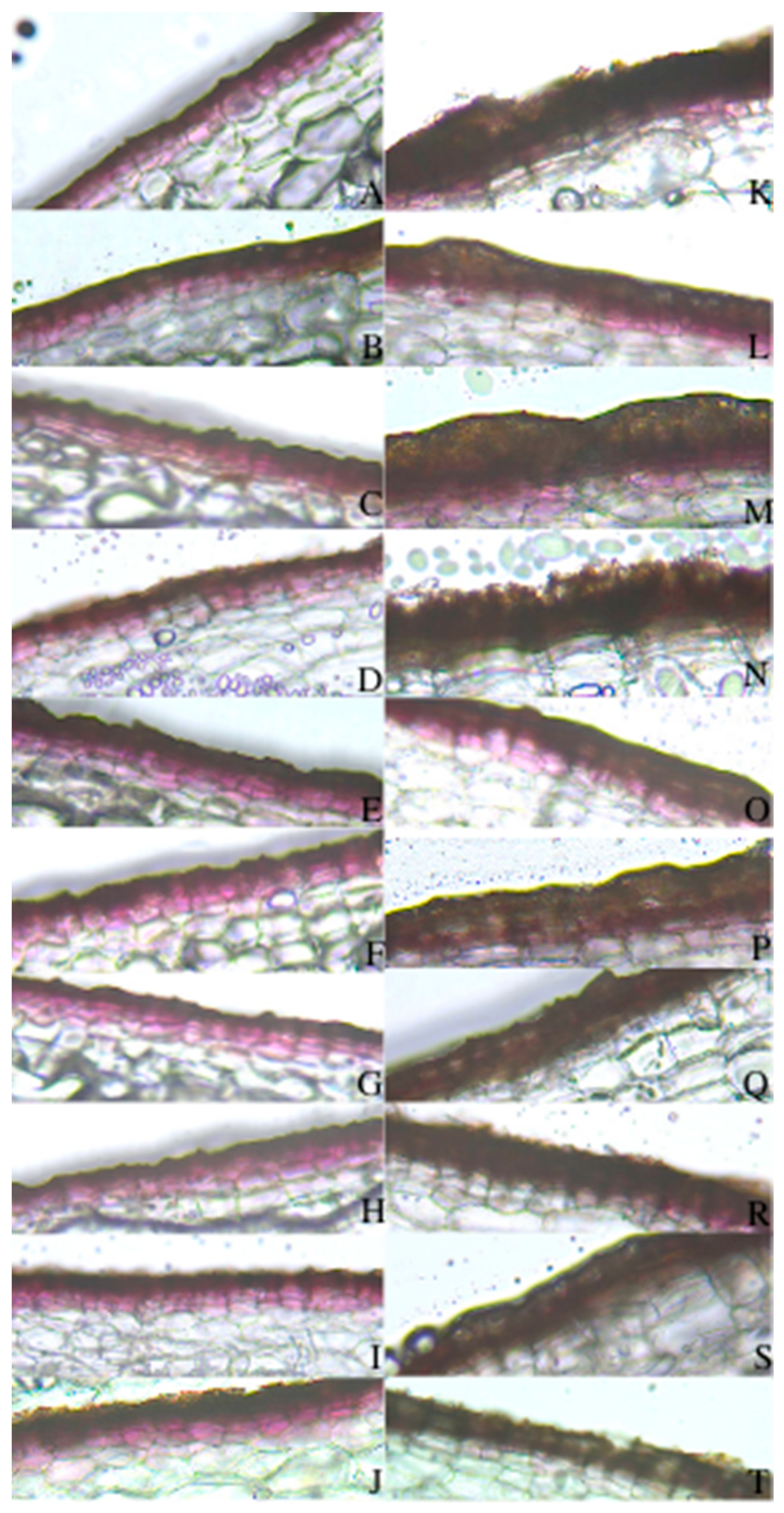
2.3. Suberized Cell Layer
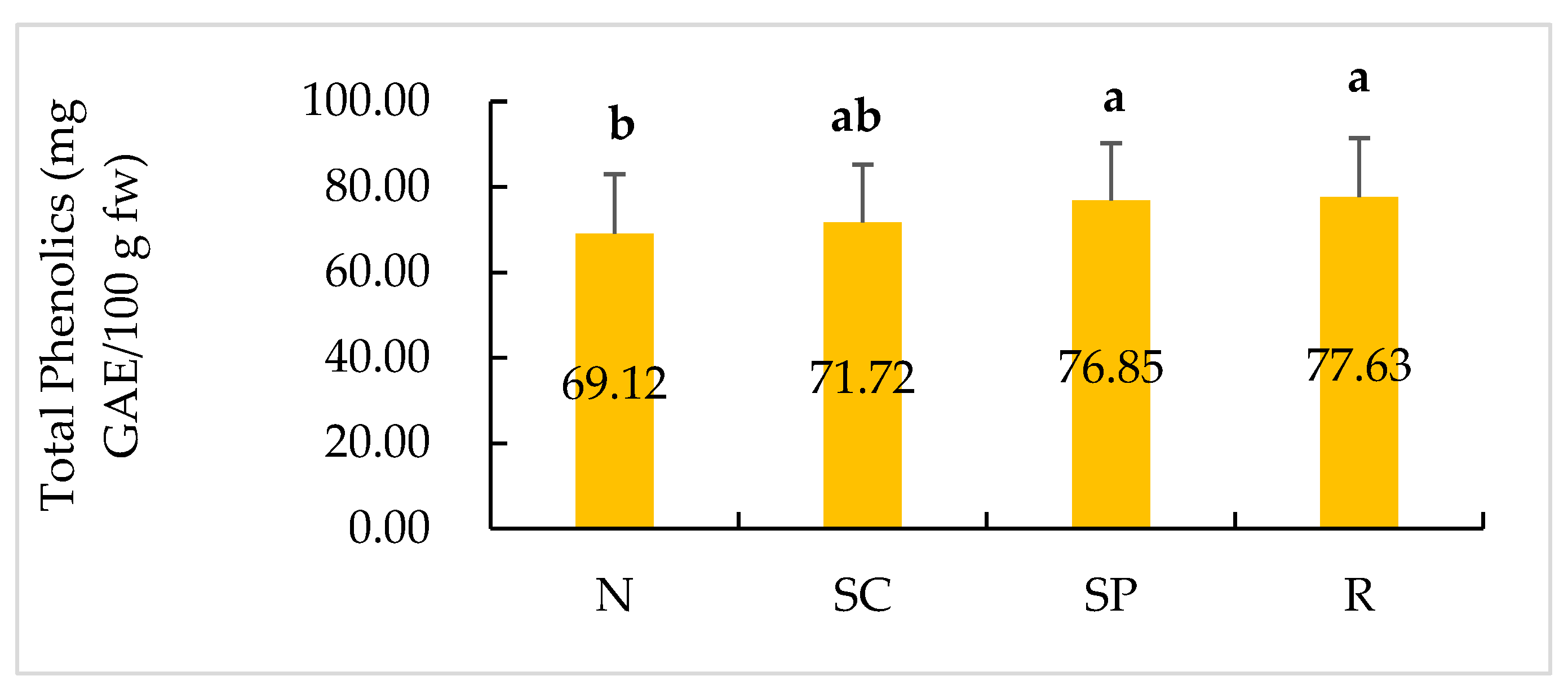
2.4. Total Phenolic Content
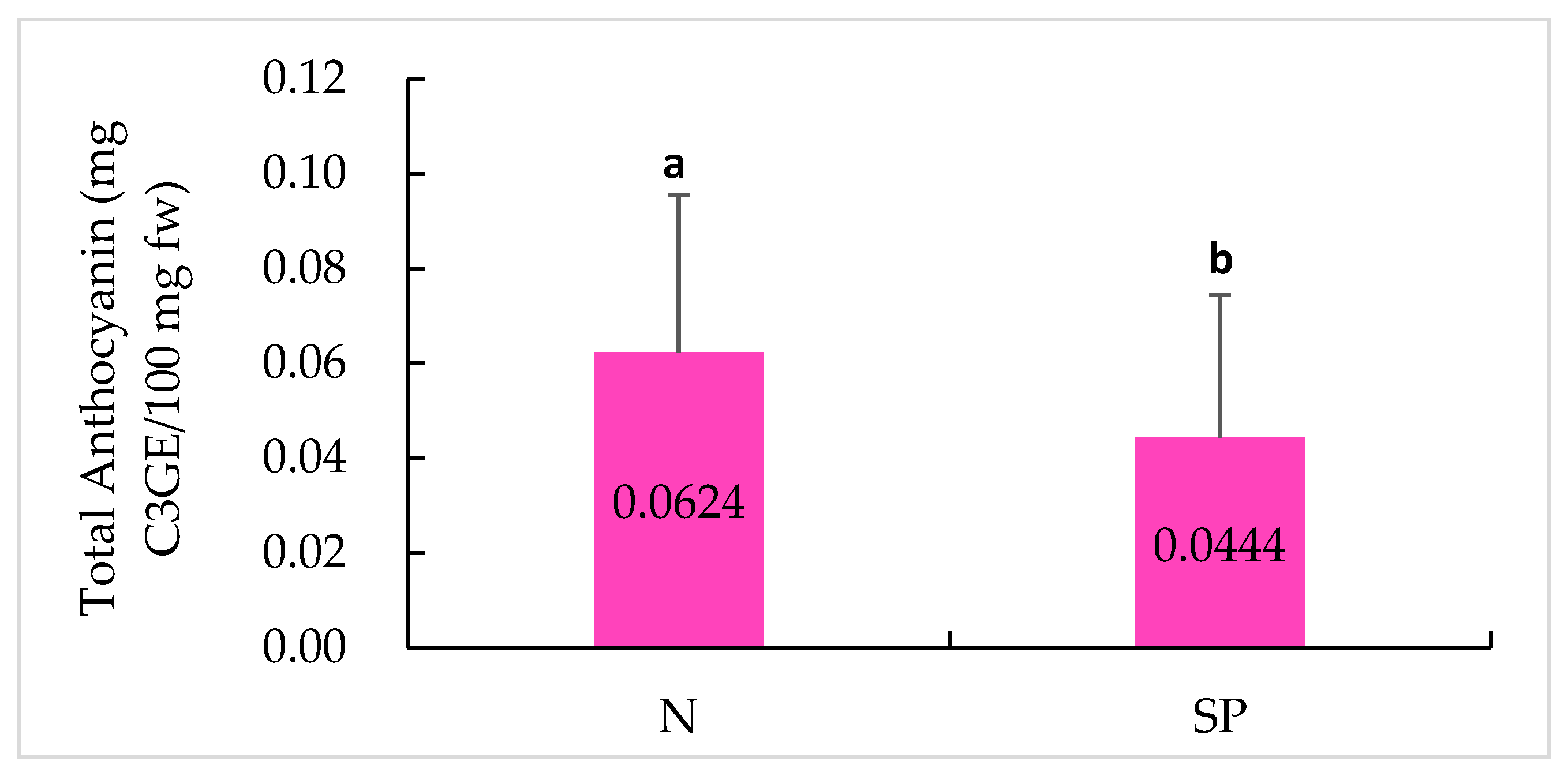
2.5. Anthocyanin Content
3. Materials and Methods
3.1. Three-Year Field Trial
3.2. Sampling
3.2.1. Sampling for Suberin Analysis
3.2.2. Sampling for Total Phenolics and Anthocyanin Analyses
3.3. Evaluation of Suberized Cell Layer
3.4. Determination of Total Phenolics
3.5. Determination of Total Anthocyanin
3.6. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stewart, D.; McDougall, G. Potato; A Nutritious, Tasty but Often Maligned Staple Food. Available online: https://www.hutton.ac.uk/webfm_send/743 (accessed on 10 December 2022).
- Asim, A.; Öztürk Gökçe, Z.N.; Bakhsh, A.; Tindaş Çayli, İ.; Aksoy, E.; Çalişkan, S.; Çalişkan, M.E.; DemiRel, U. Individual and Combined Effect of Drought and Heat Stresses in Contrasting Potato Cultivars Overexpressing MiR172b-3p. Turk. J. Agric. For. 2021, 45, 651–668. [Google Scholar] [CrossRef]
- Hossain, M.J.; Aksoy, E.; Öztürk Gökçe, N.Z.; Joyia, F.A.; Khan, M.S.; Bakhsh, A. Rapid and Efficient in Vitro Regeneration of Transplastomic Potato (Solanum tuberosum L.) Plants after Particle Bombardment. Turk. J. Agric. For. 2021, 45, 313–323. [Google Scholar] [CrossRef]
- Agriculture and Agri-Food Canada. Potato Market Information Review, 2020–2021. Available online: https://agriculture.canada.ca/en/agriculture-and-agri-food-canada/canadas-agriculture-sectors/horticulture/horticulture-sector-reports/potato-market-information-review-2020-2021 (accessed on 5 December 2022).
- Vulavala, V.K.R.; Fogelman, E.; Faigenboim, A.; Shoseyov, O.; Ginzberg, I. The Transcriptome of Potato Tuber Phellogen Reveals Cellular Functions of Cork Cambium and Genes Involved in Periderm Formation and Maturation. Sci. Rep. 2019, 9, 10216. [Google Scholar] [CrossRef] [Green Version]
- Keren-Keiserman, A.; Baghel, R.S.; Fogelman, E.; Faingold, I.; Zig, U.; Yermiyahu, U.; Ginzberg, I. Effects of Polyhalite Fertilization on Skin Quality of Potato Tuber. Front. Plant Sci. 2019, 10, 1379. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Bhardwaj, V.; Kaur, K.; Kukreja, S.; Goutam, U. Potato Periderm Is the First Layer of Defence against Biotic and Abiotic Stresses: A Review. Potato Res. 2021, 64, 131–146. [Google Scholar] [CrossRef]
- André, C.M.; Schafleitner, R.; Legay, S.; Lefèvre, I.; Aliaga, C.A.A.; Nomberto, G.; Hoffmann, L.; Hausman, J.-F.; Larondelle, Y.; Evers, D. Gene Expression Changes Related to the Production of Phenolic Compounds in Potato Tubers Grown under Drought Stress. Phytochemistry 2009, 70, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Seitz, H.U.; Hinderer, W. CHAPTER 3—Anthocyanins. In Phytochemicals in Plant Cell Cultures; Constabel, F., Vasil, I.K., Eds.; Academic Press: Cambridge, MA, USA, 1988; pp. 49–76. ISBN 978-0-12-715005-5. [Google Scholar]
- Strygina, K.V.; Kochetov, A.V.; Khlestkina, E.K. Genetic Control of Anthocyanin Pigmentation of Potato Tissues. BMC Genet. 2019, 20, 27. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.M.; Visser, R.G.F.; Bovy, A. Anthocyanin Biosynthesis and Degradation Mechanisms in Solanaceous Vegetables: A Review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef] [PubMed]
- Mikitzel, L. Tuber Physiological Disorders. In The Potato: Botany, Production and Uses; Botany, Production and Uses; CABI: Oxford, UK, 2014; pp. 237–254. ISBN 978-1-78064-280-2. [Google Scholar]
- Ginzberg, I.; Minz, D.; Faingold, I.; Soriano, S.; Mints, M.; Fogelman, E.; Warshavsky, S.; Zig, U.; Yermiyahu, U. Calcium Mitigated Potato Skin Physiological Disorder. Am. J. Pot. Res. 2012, 89, 351–362. [Google Scholar] [CrossRef]
- Lynch, D.R.; Foroud, N.; Kozub, G.C.; Fames, B.C. The Effect of Moisture Stress at Three Growth Stages on the Yield, Components of Yield and Processing Quality of Eight Potato Varieties. Am. Potato J. 1995, 72, 375–385. [Google Scholar] [CrossRef]
- van Loon, C.D. The Effect of Water Stress on Potato Growth, Development, and Yield. Am. Potato J. 1981, 58, 51–69. [Google Scholar] [CrossRef]
- Onder, S.; Caliskan, M.E.; Onder, D.; Caliskan, S. Different Irrigation Methods and Water Stress Effects on Potato Yield and Yield Components. Agric. Water Manag. 2005, 73, 73–86. [Google Scholar] [CrossRef]
- Karafyllidis, D.I.; Stavropoulos, N.; Georgakis, D. The Effect of Water Stress on the Yielding Capacity of Potato Crops and Subsequent Performance of Seed Tubers. Potato Res. 1996, 39, 153–163. [Google Scholar] [CrossRef]
- Miller, D.E.; Martin, M.W. Effect of Declining or Interrupted Irrigation on Yield and Quality of Three Potato Cultivars Grown on Sandy Soil. Am. Potato J. 1987, 64, 109–117. [Google Scholar] [CrossRef]
- Hill, D.; Nelson, D.; Hammond, J.; Bell, L. Morphophysiology of Potato (Solanum tuberosum) in Response to Drought Stress: Paving the Way Forward. Front. Plant Sci. 2021, 11, 597554. [Google Scholar] [CrossRef]
- Zhang, Z.; Huang, Z.; Lu, H.; Wang-Pruski, G. A Rapid and Effective Method for Observation of Suberized Cell Layers in Potato Tuber Skin. Sci. Hortic. 2017, 224, 215–218. [Google Scholar] [CrossRef]
- Škerget, M.; Kotnik, P.; Hadolin, M.; Hraš, A.R.; Simonič, M.; Knez, Ž. Phenols, Proanthocyanidins, Flavones and Flavonols in Some Plant Materials and Their Antioxidant Activities. Food Chem. 2005, 89, 191–198. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Characterization and Measurement of Anthocyanins by UV-Visible Spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, F1.2.1–F1.2.13. [Google Scholar] [CrossRef]
- Francis, F.J. Food Colorants: Anthocyanins. Crit. Rev. Food Sci. Nutr. 1989, 28, 273–314. [Google Scholar] [CrossRef]
- Ávila-Valdés, A.; Quinet, M.; Lutts, S.; Martínez, J.P.; Lizana, X.C. Tuber Yield and Quality Responses of Potato to Moderate Temperature Increase during Tuber Bulking under Two Water Availability Scenarios. Field Crops Res. 2020, 251, 107786. [Google Scholar] [CrossRef]
- Ojala, J.C.; Stark, J.C.; Kleinkopf, G.E. Influence of Irrigation and Nitrogen Management on Potato Yield and Quality. Am. Potato J. 1990, 67, 29–43. [Google Scholar] [CrossRef]
- Wagg, C.; Hann, S.; Kupriyanovich, Y.; Li, S. Timing of Short Period Water Stress Determines Potato Plant Growth, Yield and Tuber Quality. Agric. Water Manag. 2021, 247, 106731. [Google Scholar] [CrossRef]
- Kumar, P.; Ginzberg, I. Potato Periderm Development and Tuber Skin Quality. Plants 2022, 11, 2099. [Google Scholar] [CrossRef]
- Barel, G.; Ginzberg, I. Potato Skin Proteome Is Enriched with Plant Defence Components. J. Exp. Bot. 2008, 59, 3347–3357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woolfson, K.N.; Esfandiari, M.; Bernards, M.A. Suberin Biosynthesis, Assembly, and Regulation. Plants 2022, 11, 555. [Google Scholar] [CrossRef] [PubMed]
- Ginzberg, I.; Barel, G.; Ophir, R.; Tzin, E.; Tanami, Z.; Muddarangappa, T.; de Jong, W.; Fogelman, E. Transcriptomic Profiling of Heat-Stress Response in Potato Periderm. J. Exp. Bot. 2009, 60, 4411–4421. [Google Scholar] [CrossRef]
- Tanios, S.; Thangavel, T.; Eyles, A.; Tegg, R.S.; Nichols, D.S.; Corkrey, R.; Wilson, C.R. Suberin Deposition in Potato Periderm: A Novel Resistance Mechanism against Tuber Greening. New Phytol. 2020, 225, 1273–1284. [Google Scholar] [CrossRef] [PubMed]
- Vulavala, V.K.R.; Fogelman, E.; Rozental, L.; Faigenboim, A.; Tanami, Z.; Shoseyov, O.; Ginzberg, I. Identification of Genes Related to Skin Development in Potato. Plant Mol. Biol. 2017, 94, 481–494. [Google Scholar] [CrossRef]
- Neubauer, J.D.; Lulai, E.C.; Thompson, A.L.; Suttle, J.C.; Bolton, M.D. Wounding Coordinately Induces Cell Wall Protein, Cell Cycle and Pectin Methyl Esterase Genes Involved in Tuber Closing Layer and Wound Periderm Development. J. Plant Physiol. 2012, 169, 586–595. [Google Scholar] [CrossRef]
- Serra, O.; Soler, M.; Hohn, C.; Sauveplane, V.; Pinot, F.; Franke, R.; Schreiber, L.; Prat, S.; Molinas, M.; Figueras, M. CYP86A33 -Targeted Gene Silencing in Potato Tuber Alters Suberin Composition, Distorts Suberin Lamellae, and Impairs the Periderm’s Water Barrier Function. Plant Physiol. 2009, 149, 1050–1060. [Google Scholar] [CrossRef]
- Bjelica, A.; Haggitt, M.L.; Woolfson, K.N.; Lee, D.P.N.; Makhzoum, A.B.; Bernards, M.A. Fatty Acid ω-Hydroxylases from Solanum tuberosum. Plant Cell. Rep. 2016, 35, 2435–2448. [Google Scholar] [CrossRef] [PubMed]
- Haggitt, M. Role of Fatty Acid Omega-Hydroxylase 1 and Abscisic Acid in Potato Tuber Suberin Formation. Available online: https://ir.lib.uwo.ca/cgi/viewcontent.cgi?article=5355&context=etd (accessed on 10 December 2022).
- Serra, O.; Hohn, C.; Franke, R.; Prat, S.; Molinas, M.; Figueras, M. A Feruloyl Transferase Involved in the Biosynthesis of Suberin and Suberin-Associated Wax Is Required for Maturation and Sealing Properties of Potato Periderm: FHT Function in Potato Periderm. Plant J. 2010, 62, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Rykaczewska, K. The Effect of High Temperature Occurring in Subsequent Stages of Plant Development on Potato Yield and Tuber Physiological Defects. Am. J. Potato Res. 2015, 92, 339–349. [Google Scholar] [CrossRef] [Green Version]
- Malejane, D.N.; Tinyani, P.; Soundy, P.; Sultanbawa, Y.; Sivakumar, D. Deficit Irrigation Improves Phenolic Content and Antioxidant Activity in Leafy Lettuce Varieties. Food Sci. Nutr. 2017, 6, 334–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payyavula, R.S.; Navarre, D.A.; Kuhl, J.C.; Pantoja, A.; Pillai, S.S. Differential Effects of Environment on Potato Phenylpropanoid and Carotenoid Expression. BMC Plant Biol. 2012, 12, 39. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Álvarez, E.P.; Intrigliolo, D.S.; Almajano, M.P.; Rubio-Bretón, P.; Garde-Cerdán, T. Effects of Water Deficit Irrigation on Phenolic Composition and Antioxidant Activity of Monastrell Grapes under Semiarid Conditions. Antioxidants 2021, 10, 1301. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, V.; Alooparampil, S.; Pandya, R.V.; Tank, J.G.; Chowdhary, V.; Alooparampil, S.; Pandya, R.V.; Tank, J.G. Physiological Function of Phenolic Compounds in Plant Defense System; IntechOpen: London, UK, 2021; ISBN 978-1-83969-347-2. [Google Scholar]
- Kałużewicz, A.; Lisiecka, J.; Gąsecka, M.; Krzesiński, W.; Spiżewski, T.; Zaworska, A.; Frąszczak, B. The Effects of Plant Density and Irrigation on Phenolic Content in Cauliflower. Hortic. Sci. 2017, 44, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Hura, T.; Hura, K.; Grzesiak, S. Contents of Total Phenolics and Ferulic Acid, and PAL Activity during Water Potential Changes in Leaves of Maize Single-Cross Hybrids of Different Drought Tolerance. J. Agron. Crop Sci. 2008, 194, 104–112. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drought Stress Enhances Nutritional and Bioactive Compounds, Phenolic Acids and Antioxidant Capacity of Amaranthus Leafy Vegetable. BMC Plant Biol. 2018, 18, 258. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Rodríguez, E.; Ruiz, J.M.; Ferreres, F.; Moreno, D.A. Phenolic Profiles of Cherry Tomatoes as Influenced by Hydric Stress and Rootstock Technique. Food Chem. 2012, 134, 775–782. [Google Scholar] [CrossRef]
- Robbins, R.J.; Keck, A.-S.; Banuelos, G.; Finley, J.W. Cultivation Conditions and Selenium Fertilization Alter the Phenolic Profile, Glucosinolate, and Sulforaphane Content of Broccoli. J. Med. Food 2005, 8, 204–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cogo, S.L.P.; Chaves, F.C.; Schirmer, M.A.; Zambiazi, R.C.; Nora, L.; Silva, J.A.; Rombaldi, C.V. Low Soil Water Content during Growth Contributes to Preservation of Green Colour and Bioactive Compounds of Cold-Stored Broccoli (Brassica oleraceae L.) Florets. Postharvest Biol. Technol. 2011, 60, 158–163. [Google Scholar] [CrossRef] [Green Version]
- Fortier, E.; Desjardins, Y.; Tremblay, N.; Bélec, C.; Côté, M. Influence of Irrigation and Nitrogen Fertilization on Broccoli Polyphenolics Concentration. Acta Hortic. 2010, 856, 55–62. [Google Scholar] [CrossRef]
- Mao, L.; Jett, L.E.; Story, R.N.; Hammond, A.M.; Peterson, J.K.; Labonte, D.R. Influence of Drought Stress on Sweetpotato Resistance to Sweetpotato Weevil, Cylas Formicarius (Coleoptera: Apoinidae), and Storage Root Chemistry. Fla. Entomol. 2004, 87, 261–267. [Google Scholar] [CrossRef]
- The Effect of Water Deficit Stress on the Composition of Phenolic Compounds in Medicinal Plants. South Afr. J. Bot. 2020, 131, 12–17. [CrossRef]
- Andersen, A.W.; Tong, C.B.S.; Krueger, D.E. Comparison of Periderm Color and Anthocyanins of Four Red Potato Varieties. Am. J. Potato Res 2002, 79, 249–253. [Google Scholar] [CrossRef]
- Jiang, Y. Role of Anthocyanins, Polyphenol Oxidase and Phenols in Lychee Pericarp Browning. J. Sci. Food Agric. 2000, 80, 305–310. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Regime | 2019 | 2020 | 2021 |
|---|---|---|---|
| Rainfed | 7.327 a | 6.730 a | 7.110 a |
| Irrigated | 6.675 b | 6.370 b | 6.735 b |
| Skin Type | Treatment | |||||||
|---|---|---|---|---|---|---|---|---|
| Rainfed | Irrigated | |||||||
| No Stress | Tuber Initiation | Tuber Bulking | Tuber Skinset | No Stress | Tuber Initiation | Tuber Bulking | Tuber Skinset | |
| N | 7.000 c | 6.900 b | 6.959 b | 7.358 b | 6.752 c | 6.385 c | 6.600 c | 6.611 b |
| SC | 7.856 b | 7.970 a | 7.805 a | 8.330 a | 8.329 a | 8.170 a | 8.333 a | 8.031 a |
| SP | 8.495 a | 8.000 a | 7.740 a | 8.086 a | 7.835 b | 7.950 a | 7.710 b | 8.119 a |
| R | 6.464 c | 6.282 c | 6.860 b | 6.700 c | 7.000 c | 7.044 b | 6.810 c | 6.214 b |
| Year | Date of Irrigation | Amount Applied (Inches) |
|---|---|---|
| 2019 | 3 June | 0.50 |
| 5 June | 0.50 | |
| 12 June | 0.50 | |
| 27 June | 0.50 | |
| 3 July | 0.50 | |
| 18 July | 0.50 | |
| 24 July | 0.50 | |
| 29 July | 0.50 | |
| 5 August | 0.75 | |
| 7 August | 0.50 | |
| 13 August | 0.75 | |
| 2020 | 4 June | 0.75 |
| 17 June | 0.75 | |
| 24 June | 0.50 | |
| 29 June | 0.50 | |
| 21 July | 0.75 | |
| 29 July | 1.20 | |
| 2021 | 24 June | 1.00 |
| 30 June | 0.75 | |
| 6 July | 0.50 | |
| 9 July | 0.50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, M.; Shinners-Carnelley, T.; Gibson, D.; Jones, D.; Joshi, J.; Wang-Pruski, G. Irrigation Effect on Yield, Skin Blemishes, Phellem Formation, and Total Phenolics of Red Potatoes. Plants 2022, 11, 3523. https://doi.org/10.3390/plants11243523
Jiang M, Shinners-Carnelley T, Gibson D, Jones D, Joshi J, Wang-Pruski G. Irrigation Effect on Yield, Skin Blemishes, Phellem Formation, and Total Phenolics of Red Potatoes. Plants. 2022; 11(24):3523. https://doi.org/10.3390/plants11243523
Chicago/Turabian StyleJiang, Manlin, Tracy Shinners-Carnelley, Darin Gibson, Debbie Jones, Jyoti Joshi, and Gefu Wang-Pruski. 2022. "Irrigation Effect on Yield, Skin Blemishes, Phellem Formation, and Total Phenolics of Red Potatoes" Plants 11, no. 24: 3523. https://doi.org/10.3390/plants11243523