
Growth and Biochemical Composition of Microgreens Grown in Different Formulated Soilless Media
 and
and
Abstract
:1. Introduction
2. Results
2.1. Growing Media Properties
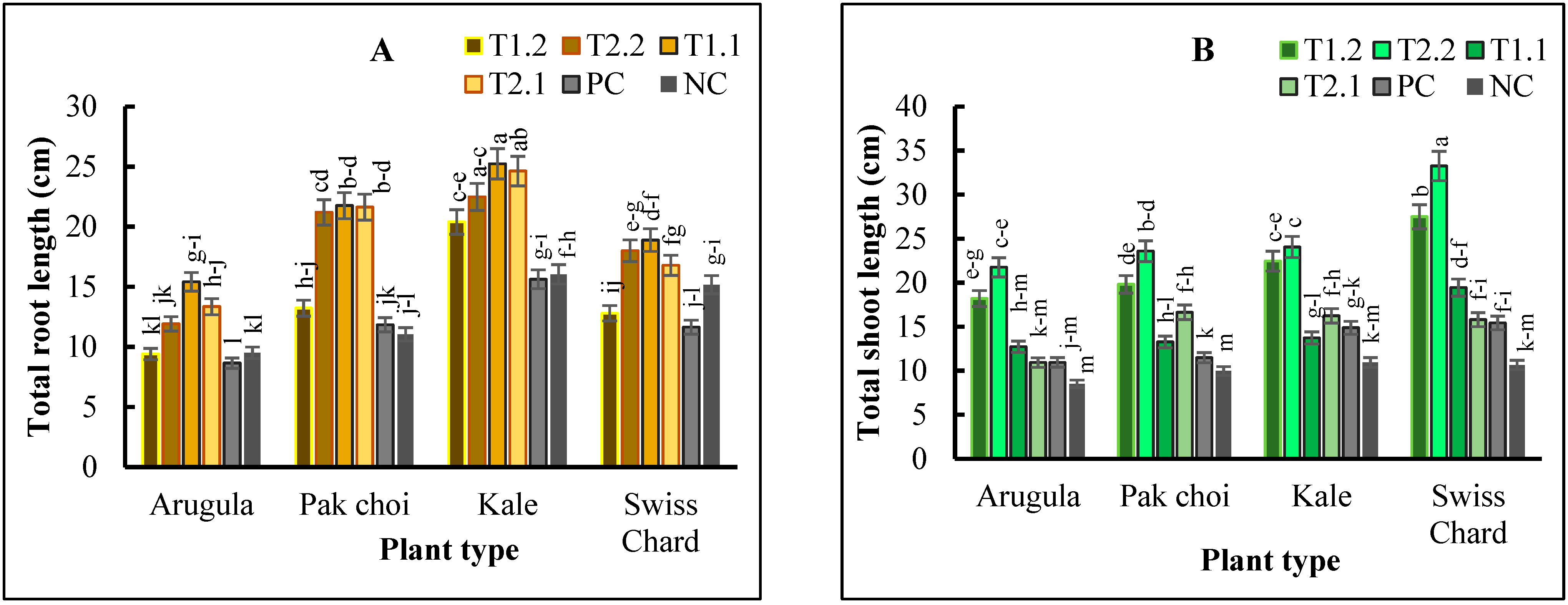
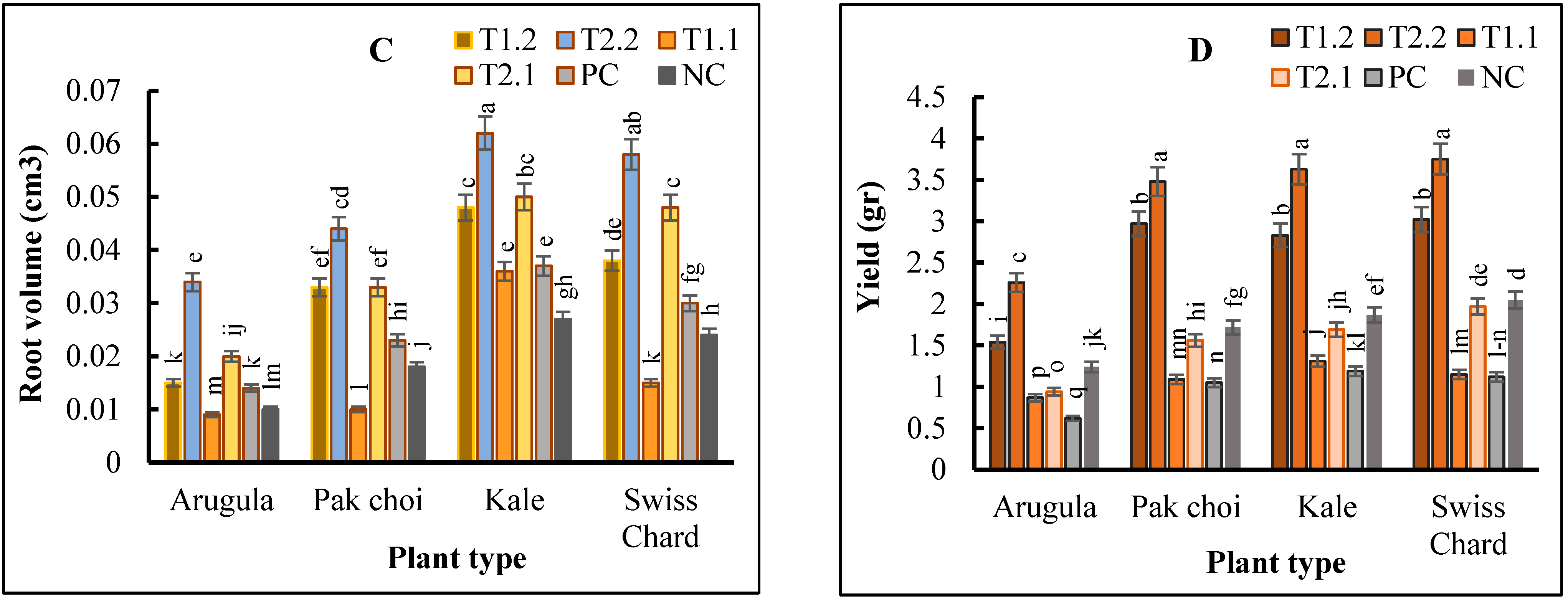
2.2. Plant Growth and Yield
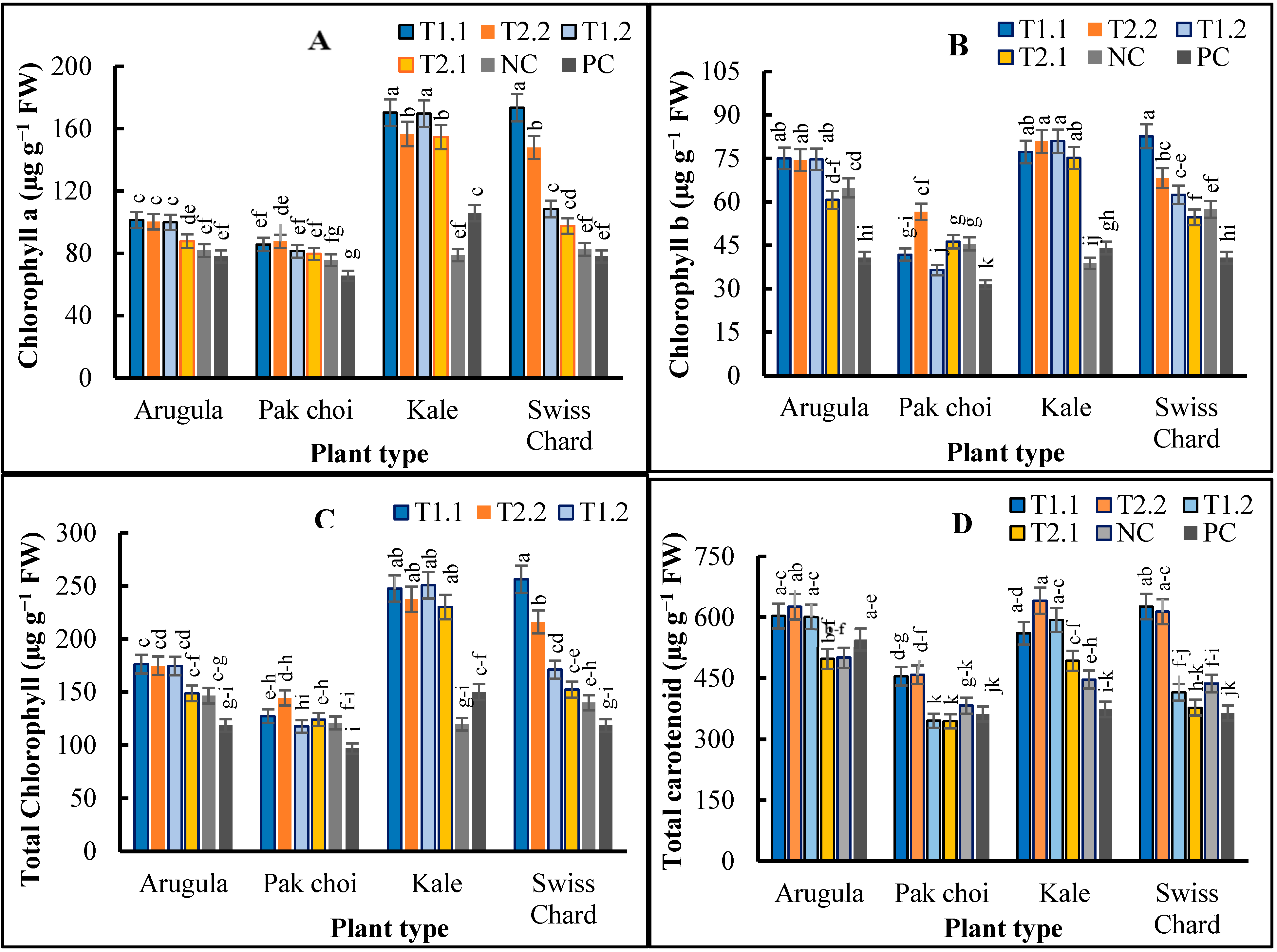
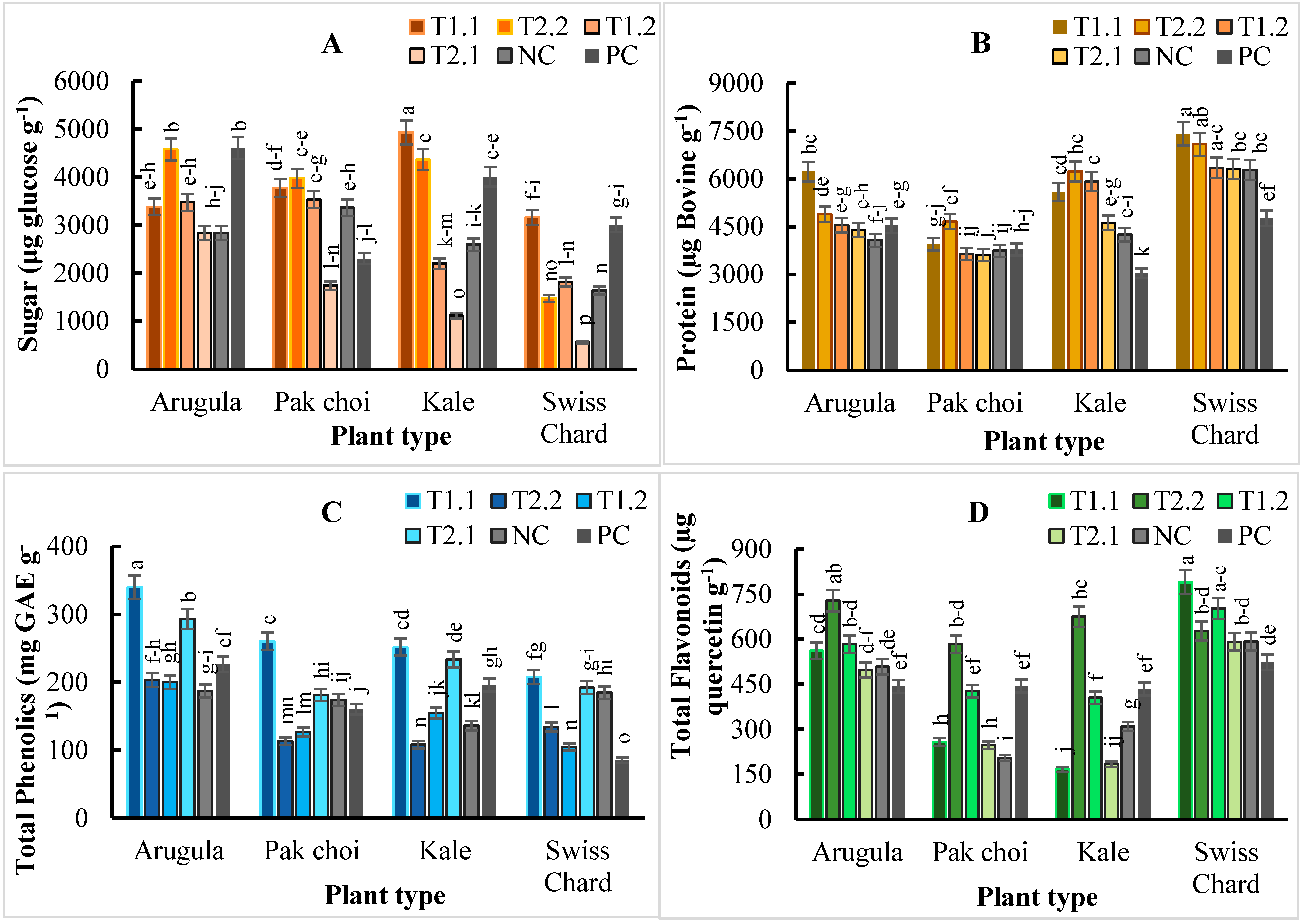
2.3. Microgreens Biochemical Composition
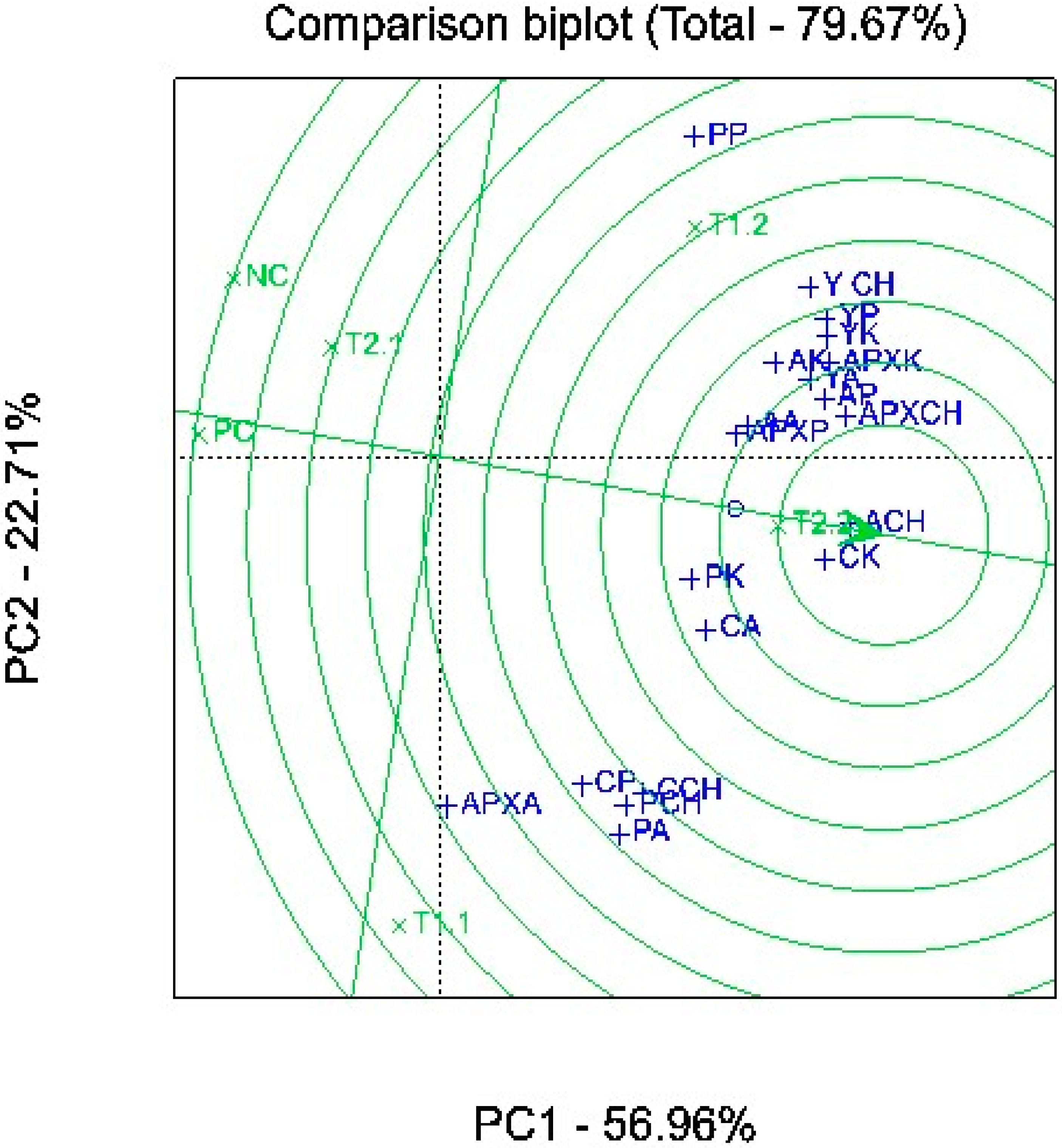
2.4. Association among Media, Plants, and Biochemical Composition
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growing Condition
4.2. Experimental Treatment and Design
4.3. Growing Media Physicochemical Properties
4.4. Plant Growth and Yield Components
4.5. Microgreen Quality and Phytochemical Analysis
4.5.1. Chlorophylls a and b, Total Chlorophyll, and Total Carotenoid
4.5.2. Total Sugar
4.5.3. Total Protein
4.5.4. Total Phenolics
4.5.5. Total Flavonoids
4.5.6. Total Ascorbate
4.5.7. Antioxidant Enzyme Activity
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bulgari, R.; Negri, M.; Santoro, P.; Ferrante, A. Quality evaluation of indoor-grown microgreens cultivated on three different substrates. Horticulturae 2021, 7, 96. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; El-Nakhel, C.; Graziani, G.; Pannico, A.; Soteriou, G.A.; Giordano, M.; Rouphael, Y. Functional quality in novel food sources: Genotypic variation in the nutritive and phytochemical composition of thirteen microgreens species. Food Chem. 2019, 277, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, S.; Khan, A. Antioxidants and diabetes. Indian J. Endocrinol. Metab. 2012, 16 (Suppl. 2), S267. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, M.C.; Rouphael, Y.; Di Gioia, F.; Kyratzis, A.; Serio, F.; Renna, M.; Santamaria, P. Micro-scale vegetable production and the rise of microgreens. Trends Food Sci. Technol. 2016, 57, 103–115. [Google Scholar] [CrossRef]
- Pinto, E.; Almeida, A.A.; Aguiar, A.A.; Ferreira, I.M. Comparison between the mineral profile and nitrate content of microgreens and mature lettuces. J. Food Compos. Anal. 2015, 37, 38–43. [Google Scholar] [CrossRef]
- Di Noia, J. Peer Reviewed: Defining powerhouse fruits and vegetables: A nutrient density approach. Prev. Chronic Dis. 2014, 11, 57–76. [Google Scholar] [CrossRef] [Green Version]
- Aftab, T. A review of medicinal and aromatic plants and their secondary metabolites status under abiotic stress. J. Med. Plants 2019, 7, 99–106. [Google Scholar]
- Nguyen, D.T.; Lu, N.; Kagawa, N.; Takagaki, M. Optimization of Photosynthetic Photon Flux Density and Root-zone Temperature for Enhancing Secondary Metabolite Accumulation and Production of Coriander in Plant Factory. Agronomy 2019, 9, 224. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Tiyip, T.; Ding, J.L.; He, Q.S. The effects of the chemical components of soil salinity on electrical conductivity in the region of the delta oasis of Weigan and Kuqa Rivers, China. Agric. Sci. China 2009, 8, 985–993. [Google Scholar] [CrossRef]
- Zhang, B.; Wijesundara, N.M.; Abbey, L.; Rupasinghe, H.V. Growing medium amendments effect on growth, secondary metabolites, and anti-streptococcal activity of two species of Plectranthus. J. Appl. Res. Med. Aromat. Plants 2017, 5, 53–59. [Google Scholar] [CrossRef]
- Abbott, L.K.; Macdonald, L.M.; Wong, M.T.F.; Webb, M.J.; Jenkins, S.N.; Farrell, M. Potential roles of biological amendments for profitable grain production—A review. Agric. Ecosyst. Environ. 2018, 256, 34–50. [Google Scholar] [CrossRef]
- da Costa Jaeggi, M.E.P.; Rodrigues, R.R.; Pereira, I.M.; do Carmo Parajara, M.; Rocha, R.S.; da Cruz, D.P.; de Araújo Capetini, S. Vegetative Development of Radish Seedlings in Different Organic Substrates. J. Exp. Agric. Int. 2019, 10, 1–8. [Google Scholar] [CrossRef]
- Treutter, D. Managing phenol contents in crop plants by phytochemical farming and breeding—Visions and constraints. Int. J. Mol. Sci. 2010, 11, 807–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbey, L.; Pham, T.H.; Annan, N.; Leke-Aladekoba, A.; Thomas, R.H. Chemical composition of kale as influenced by dry vermicast, potassium humate and volcanic minerals. Food Res. Int. 2018, 107, 726–737. [Google Scholar] [CrossRef]
- Vahid Afagh, H.; Saadatmand, S.; Riahi, H.; Khavari-Nejad, R.A. Influence of spent mushroom compost (SMC) as an organic fertilizer on nutrient, growth, yield, and essential oil composition of German chamomile (Matricaria recutita L.). Commun. Soil Sci. Plant Anal. 2019, 50, 538–548. [Google Scholar] [CrossRef]
- Abbey, L.; Udenigwe, C.; Mohan, A.; Anom, E. Microwave irradiation effects on vermicasts potency, and plant growth and antioxidant activity in seedlings of Chinese cabbage (Brassica rapa subsp. pekinensis). J. Radiat. Res. Appl. Sci. 2017, 10, 110–116. [Google Scholar] [CrossRef] [Green Version]
- Iheshiulo, E.M.A.; Abbey, L.; Asiedu, S.K. Response of kale to single-dose application of K humate, dry vermicasts, and volcanic minerals. Int. J. Veg. Sci. 2007, 23, 135–144. [Google Scholar] [CrossRef]
- Karthikeyan, M.; Hussain, N.; Gajalakshmi, S.; Abbasi, S.A. Effect of vermicast generated from an allelopathic weed lantana (Lantana camara) on seed germination, plant growth, and yield of cluster bean (Cyamopsis tetragonoloba). Environ. Sci. Pollut. Res. 2014, 21, 12539–12548. [Google Scholar] [CrossRef]
- Shiri, M.A.; Ghasemnezhad, M.; Bakhshi, D.; Saadatian, M. Effects of ascorbic acid on phenolic compounds and antioxidant activity of packaged fresh cut table grapes. Electron. J.Environ. Agric. Food Chem. 2011, 10, 27–39. [Google Scholar]
- Trivedi, P.; Singh, K.; Pankaj, U.; Verma, S.K.; Verma, R.K.; Patra, D.D. Effect of organic amendments and microbial application on sodic soil properties and growth of an aromatic crop. Ecol. Eng. 2017, 102, 127–136. [Google Scholar] [CrossRef]
- Maharani, R.; Yutaka, T.; Yajima, T.; Minoru, T. Scrutiny on physical properties of sawdust from tropical commercial wood species: Effects of different mills and sawdust’s particle size. Indones. J. For. Res. 2010, 7, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Agboola, O.O.; Oseni, O.M.; Adewale, O.M.; Shonubi, O. Effect of the use of sawdust as a growing medium on the growth and yield of tomato. Ann. West Univ. Timisoara. Ser. Biol. 2018, 21, 67–74. [Google Scholar]
- Lin, S. Combined Effect of Vermicast-Trichoderma-Sawdust on Kale, Swiss Chard, and Pak Choi Growth. Master’s Thesis, Faculty of Agriculture, Department of Plant, Food, and Environmental Sciences, Dalhousie University, Halifax, NS, Canada, 2020. Available online: http://hdl.handle.net/10222/79587 (accessed on 2 October 2022).
- Mohan, G.; Johnson, R.L.; Yu, J. Conversion of Pine Sawdust into Polyhydroxyalkanoate Bioplastics. ACS Sustain. Chem. Eng. 2021, 9, 8383–8392. [Google Scholar]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 3–7. [Google Scholar]
- Mahboub-Khomami, A.; Padasht, M.N.; Ajili Lahiji, A.; Shirinfekr, A. The effect of sawdust vermicompost extract on Syngonium podophyllum growth and nutrition. J. Plant Nutr. 2019, 42, 410–416. [Google Scholar] [CrossRef]
- Hernández, D.; Ros, M.; Carmona, F.; Saez-Tovar, J.A.; Pascual, J.A. Composting Spent Mushroom Substrate from Agaricus bisporus and Pleurotus ostreatus Production as a Growing Media Component for Baby Leaf Lettuce Cultivation under Pythium irregulare Biotic Stress. Horticulturae 2021, 7, 13. [Google Scholar] [CrossRef]
- Renaldo, B.; Gaius, E.; Gregory, G.; Paul, V. Effects of organic amendments on early plant growth. In Proceedings of the Conference: 20th World Congress of Soil Science, Jeju, Korea, 8–13 June 2014; Available online: https://www.researchgate.net/publication/263656746_Effects_of_organic_amendments_on_early_plant_growth (accessed on 2 October 2022).
- Demir, H. The effects of spent mushroom compost on growth and nutrient contents of pepper seedlings. Mediterr. Agric. Sci. 2017, 30, 91–96. [Google Scholar]
- Emami, H.; Astaraei, A.R. Effect of organic and inorganic amendments on parameters of water retention curve, bulk density, and aggregate diameter of a saline-sodic soil. J. Agric. Sci. Technol. 2012, 14, 1625–1636. [Google Scholar]
- Chen, Z.; Han, Y.; Ning, K.; Luo, C.; Sheng, W.; Wang, S.; Wang, Q. Assessing the performance of different irrigation systems on lettuce (Lactuca sativa L.) in the greenhouse. PLoS ONE 2019, 14, e0209329. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, D.P.; Kubota, C.; Miller, S.A. Effects of low pH of hydroponic nutrient solution on plant growth, nutrient uptake, and root rot disease incidence of basil (Ocimum basilicum L.). HortScience 2020, 55, 1251–1258. [Google Scholar] [CrossRef]
- Ur Rahman, S.; Xuebin, Q.; Riaz, L.; Yasin, G.; Noor Shah, A.; Shahzad, U.; Du, Z. The interactive effect of pH variation and cadmium stress on wheat (Triticum aestivum L.) growth, physiological and biochemical parameters. PLoS ONE 2021, 16, e0253798. [Google Scholar] [CrossRef] [PubMed]
- Vidal, N.P.; Pham, H.T.; Manful, C.; Pumphrey, R.; Nadeem, M.; Cheema, M.; Thomas, R. The use of natural media amendments to produce kale enhanced with functional lipids in controlled environment production system. Sci. Rep. 2018, 8, 14771. [Google Scholar] [CrossRef]
- Warrence, N.J.; Bauder, J.W.; Pearson, K.E. Basics of salinity and solidity effects on soil physical properties. Land Resour. Environ. Sci. 2002, 7, 129. [Google Scholar]
- Lee, Y.H.; Ahn, B.K.; Sonn, Y.K. Effects of electrical conductivity on the soil microbial community in a controlled horticultural land for strawberry cultivation. Korean J. Soil Sci. Fertil. 2011, 44, 830–835. [Google Scholar] [CrossRef] [Green Version]
- Meyer, M.H.; Cunliffe, B.A. Effects of media porosity and container size on overwintering and growth of ornamental grasses. HortScience 2004, 39, 248–250. [Google Scholar] [CrossRef] [Green Version]
- Machado, R.M.A.; Serralheiro, R.P. Soil salinity: Effect on vegetable crop growth. Management practices to prevent and mitigate soil salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Shannon, M.C.; Grieve, C.M.; Lesch, S.M.; Draper, J.H. Analysis of salt tolerance in nine leafy vegetables irrigated with saline drainage water. J. Am. Soc. Hortic. Sci. 2000, 125, 658–664. [Google Scholar] [CrossRef] [Green Version]
- Gonani, Z.; Riahi, H.; Sharifi, K. Impact of using leached spent mushroom compost as a partial growing media for horticultural plants. J. Plant Nutr. 2011, 34, 337–344. [Google Scholar] [CrossRef]
- Zietz, M.; Weckmuller, A.; Schmidt, S.; Rohn, S.; Schreiner, M.; Krumbein, A.; Kroh, L.W. Genotypic and climatic influence on the antioxidant activity of flavonoids in kale (Brassica oleracea var. sabellica). J. Agric. Food Chem. 2010, 58, 2123–2130. [Google Scholar] [CrossRef]
- Sharp, R.G. A review of the applications of chitin and its derivatives in agriculture to modify plant-microbial interactions and improve crop yields. Agronomy 2013, 3, 757–793. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yoshida, S.; Mitani, N.; Egusa, M.; Takagi, M.; Izawa, H.; Ifuku, S. Disease resistance and growth promotion activities of chitin/cellulose nanofiber from spent mushroom substrate to plant. Carbohydr. Polym. 2022, 284, 119–233. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, J.; Vig, A.P. Earthworm as ecological engineers to change the physico-chemical properties of soil: Soil vs vermicast. Ecol. Eng. 2016, 90, 1–5. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Jaafar, H.Z.; Rahmat, A.; Rahman, Z.A. The relationship between phenolics and flavonoids production with total non-structural carbohydrate and photosynthetic rate in Labisia pumila Benth. under high CO2 and nitrogen fertilization. Molecules 2011, 16, 162–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arumugam, G.; Swamy, M.; Sinniah, U. Plectranthus amboinicus (Lour.) Spreng: Botanical, phytochemical, pharmacological and nutritional significance. Molecules 2016, 21, 369. [Google Scholar] [CrossRef] [Green Version]
- Armah, A. The Use of Rock Dust as a Natural Media Amendment for the Production of Horticultural Crops in Controlled Environments. Master’s Thesis, Memorial University, Grenfell Campus, Corner Brook, NL, Canada, 2021. [Google Scholar]
- Lightenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Mohammadkhani, N.; Heidari, R. Drought-induced accumulation of soluble sugars and proline in two maize varieties. World Appl. Sci. J. 2008, 3, 448–453. [Google Scholar]
- Hammond, J.B.; Kruger, N.J. The Bradford method for protein quantitation. In New Protein Techniques; Humana Press: Clifton, NJ, USA, 1988; pp. 25–32. [Google Scholar]
- Alothman, M.; Bhat, R.; Karim, A.A. Antioxidant capacity and phenolic content of selected tropical fruits from Malaysia, extracted with different solvents. Food Chem. 2009, 115, 785–788. [Google Scholar]
- Ma, Y.H.; Ma, F.W.; Zhang, J.K.; Li, M.J.; Wang, Y.H.; Liang, D. Effects of high temperature on activities and gene expression of enzymes involved in ascorbate–glutathione cycle in apple leaves. Plant Sci. 2008, 175, 761–766. [Google Scholar] [CrossRef]
- Peterson, J.M. Soils-Part 2: Physical Properties of Soil and Soil Water. 1999. Available online: https://passel2.unl.edu/view/lesson/0cff7943f577 (accessed on 2 October 2022).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Bulk Density (g/cm3) | Porosity (%) | Field Capacity (%) | pH | Salinity (mg/L) | Electric Conductivity (μS/cm) | Total Dissolved Solids (mg/L) |
|---|---|---|---|---|---|---|---|
| T1.1 | 0.07 b | 31.3 c | 29.2 c | 5.7 b | 1299.7 c | 2260.0 c | 1719.6 c |
| T1.2 | 0.12 a | 35.0 b | 34.2 a | 6.4 a | 1689.7 a | 2570.0 b | 2139.7 b |
| T2.1 | 0.07 b | 26.6 d | 25.5 d | 5.8 ab | 1319.7 c | 1233.0 e | 1709.6 c |
| T2.2 | 0.10 ab | 35.7 b | 33.2 ab | 6.3 ab | 1494.9 b | 2205.5 c | 2028.4 b |
| PC | 0.10 ab | 37.8 a | 30.2 bc | 6.1 ab | 802.9 d | 1486.0 d | 1233.5 d |
| NC | 0.09 ab | 27.6 d | 24.5 d | 5.9 ab | 1861.2 a | 3412.5 a | 2479.7 a |
| p-value | 0.015 | 0.000 | 0.000 | 0.029 | 0.000 | 0.001 | 0.001 |
| Treatment | Total Ascorbate (μmol g−1 FW) | Peroxidase Activity (Unit mg−1 FW) | Ascorbate Peroxidase Activity (Unit mg−1 FW) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Arugula | Pak Choi | Swiss Chard | Kale | Arugula | Pak Choi | Swiss Chard | Kale | Arugula | Pak Choi | Swiss Chard | Kale | |
| T1.1 | 24.0 de | 20.4 fgh | 25.4 cd | 23.6 de | 0.94 bc | 0.50 ij | 0.96 bc | 0.56 gh | 0.23 a | 0.08 fg | 0.07 fg | 0.06 g |
| T2.2 | 24.2 cde | 28.0 bc | 32.0 a | 29.7 ab | 0.67 ef | 1.05 b | 1.02 b | 0.54 hij | 0.15 cd | 0.07 g | 0.19 ab | 0.19 ab |
| T1.2 | 32.2 a | 29.5 ab | 30.0 ab | 32.0 a | 0.60 fgh | 1.25 a | 0.50 ij | 0.64 f | 0.11 e | 0.11 e | 0.16 bcd | 0.17 bcd |
| T2.1 | 21.5 efg | 22.4 ef | 21.9 ef | 28.9 b | 0.50 ij | 0.94 bc | 0.40 k | 0.48 j | 0.18 abc | 0.04 j | 0.11 e | 0.05 hi |
| NC | 17.9 hij | 16.0 j | 18.6 ghij | 19.8 fgh | 0.29 k | 0.75 de | 0.51 ij | 0.17 m | 0.06 g | 0.06 gh | 0.07 j | 0.08 f |
| PC | 20.6 fgh | 18.1 hij | 17.4 ij | 21.4 efg | 0.50 ij | 0.87 cd | 0.62 fg | 0.50 ij | 0.15 d | 0.06 gh | 0.05 de | 0.07 fg |
| p value G | 0.001 | 0.000 | 0.000 | 0.000 | 0.001 | 0.001 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| P | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.001 | 0.001 | 0.000 | 0.000 | 0.000 | 0.000 |
| G × P | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| Treatment | Formulation |
|---|---|
| T1.1 | 30% vermicast + 30% sawdust + 10% perlite + 30% PittMoss (PM) |
| T1.2 | 30% vermicast + 30% sawdust + 10% perlite + 30% mushroom compost (MC) |
| T2.1 | 30% vermicast + 20% sawdust + 20% perlite + 30% PittMoss (PM) |
| T2.2 | 30% vermicast + 20% sawdust + 20% perlite + 30% mushroom compost (MC) |
| NC | 60% sawdust + 40% PittMoss |
| PC | Pro-mix BX™ potting medium alone |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saleh, R.; Gunupuru, L.R.; Lada, R.; Nams, V.; Thomas, R.H.; Abbey, L. Growth and Biochemical Composition of Microgreens Grown in Different Formulated Soilless Media. Plants 2022, 11, 3546. https://doi.org/10.3390/plants11243546
Saleh R, Gunupuru LR, Lada R, Nams V, Thomas RH, Abbey L. Growth and Biochemical Composition of Microgreens Grown in Different Formulated Soilless Media. Plants. 2022; 11(24):3546. https://doi.org/10.3390/plants11243546
Chicago/Turabian StyleSaleh, Roksana, Lokanadha R. Gunupuru, Rajasekaran Lada, Vilis Nams, Raymond H. Thomas, and Lord Abbey. 2022. "Growth and Biochemical Composition of Microgreens Grown in Different Formulated Soilless Media" Plants 11, no. 24: 3546. https://doi.org/10.3390/plants11243546