
Genome-Wide Identification and Expression Profiling of Aconitase Gene Family Members Reveals Their Roles in Plant Development and Adaptation to Diverse Stress in Triticum aestivum L.

 , ,
, , 
Abstract
:1. Introduction
2. Results
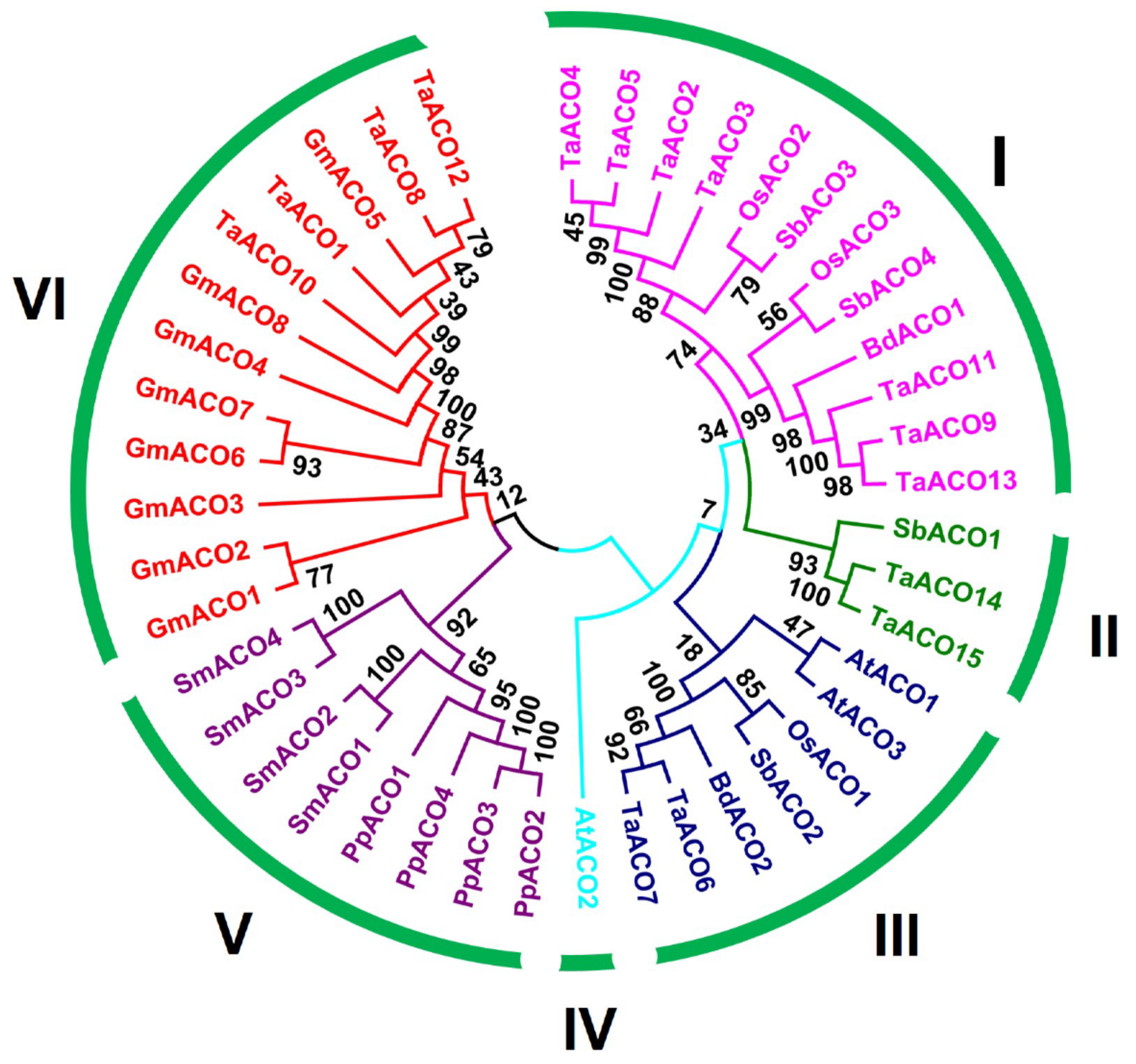
2.1. Genome-Wide Characterization and Evolutionary Analysis of TaACO Genes
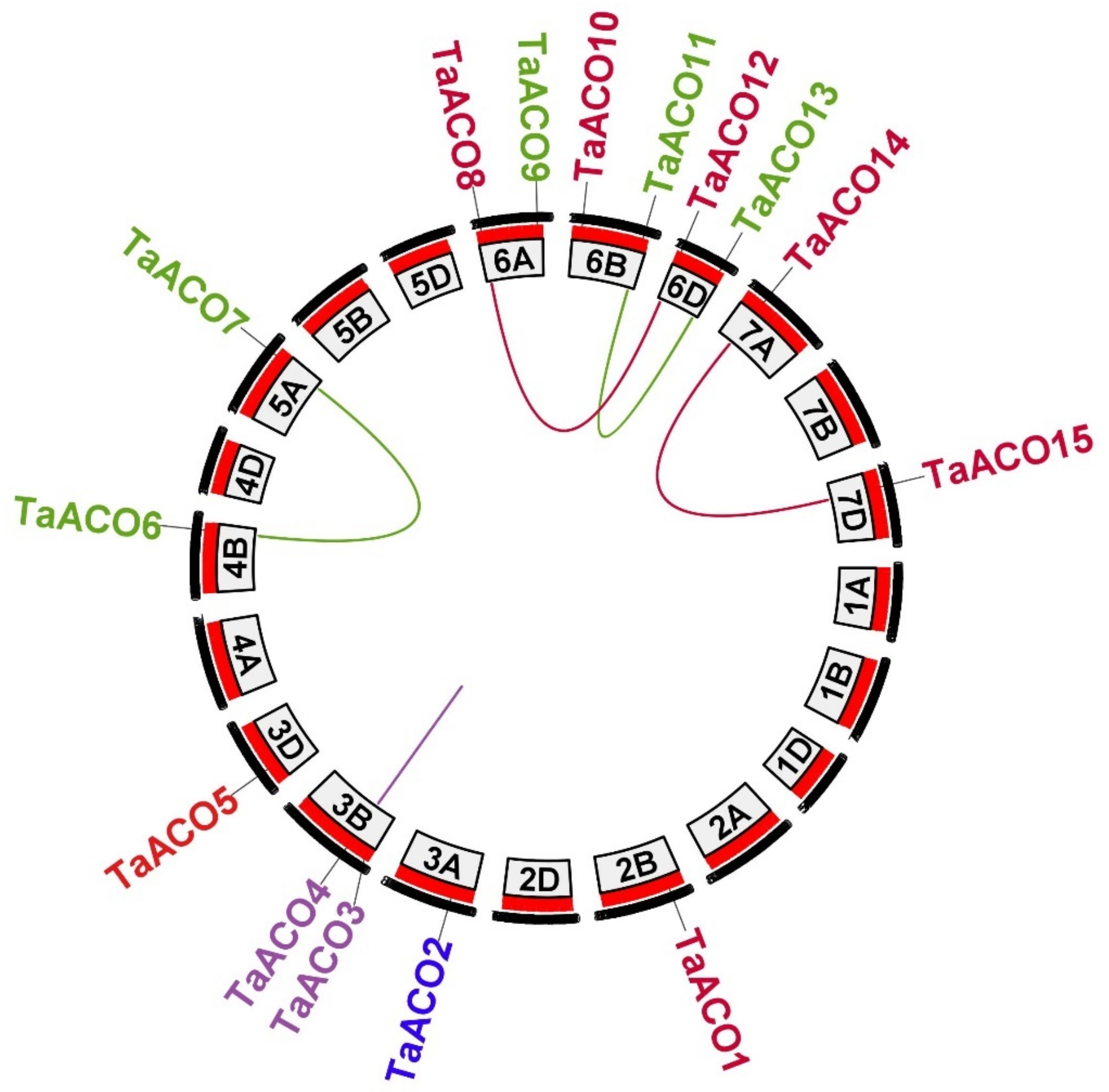
2.2. Chromosomal Location, Gene Duplication and Synteny Relationship of TaACO Genes
2.3. Gene Structure and Conserved Motif and Protein Tertiary Structure Analysis of TaACO Genes
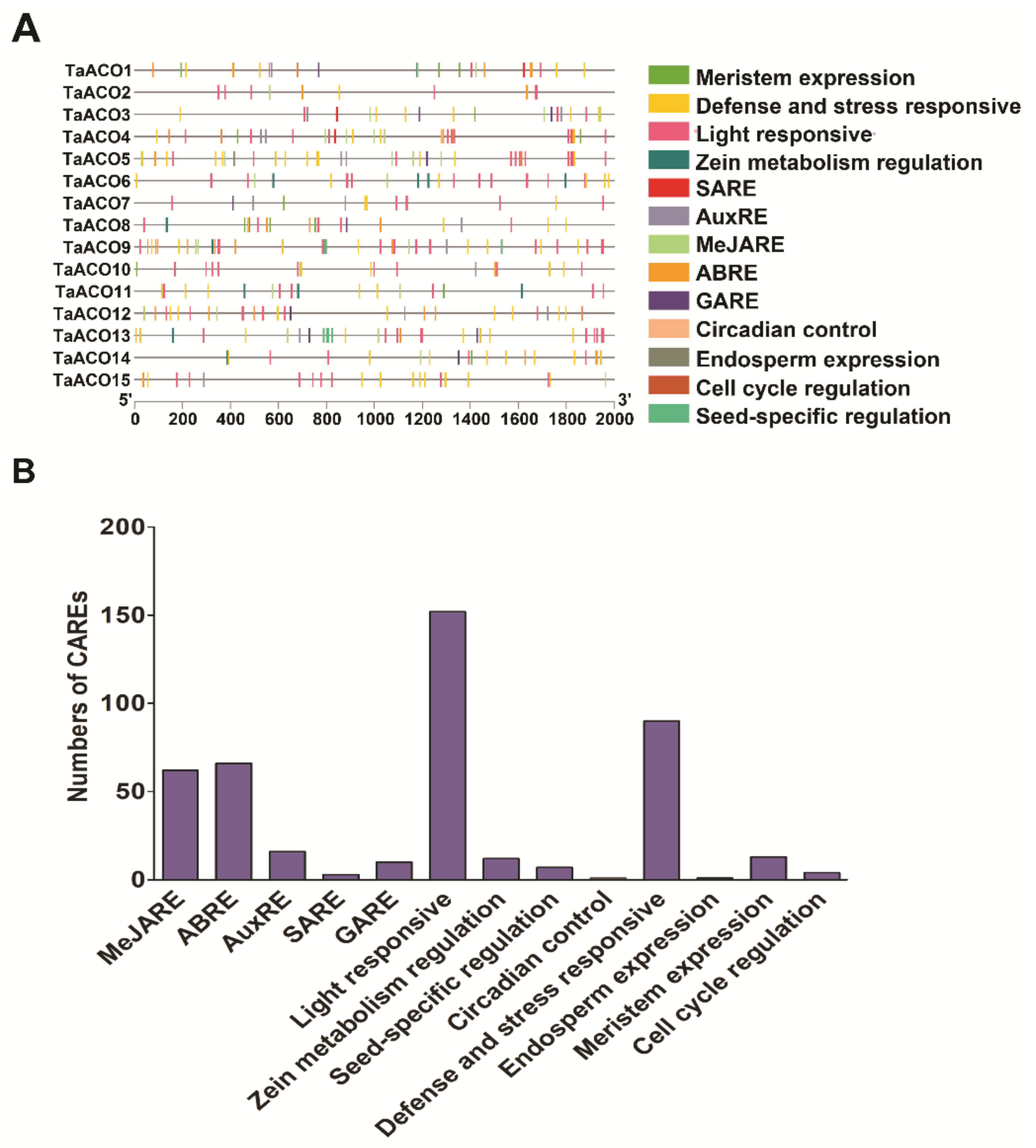
2.4. Prediction of Cis-Acting Regulatory Elements (CAREs) in TaACO Genes
2.5. Gene Ontology Analysis of TaACO Genes
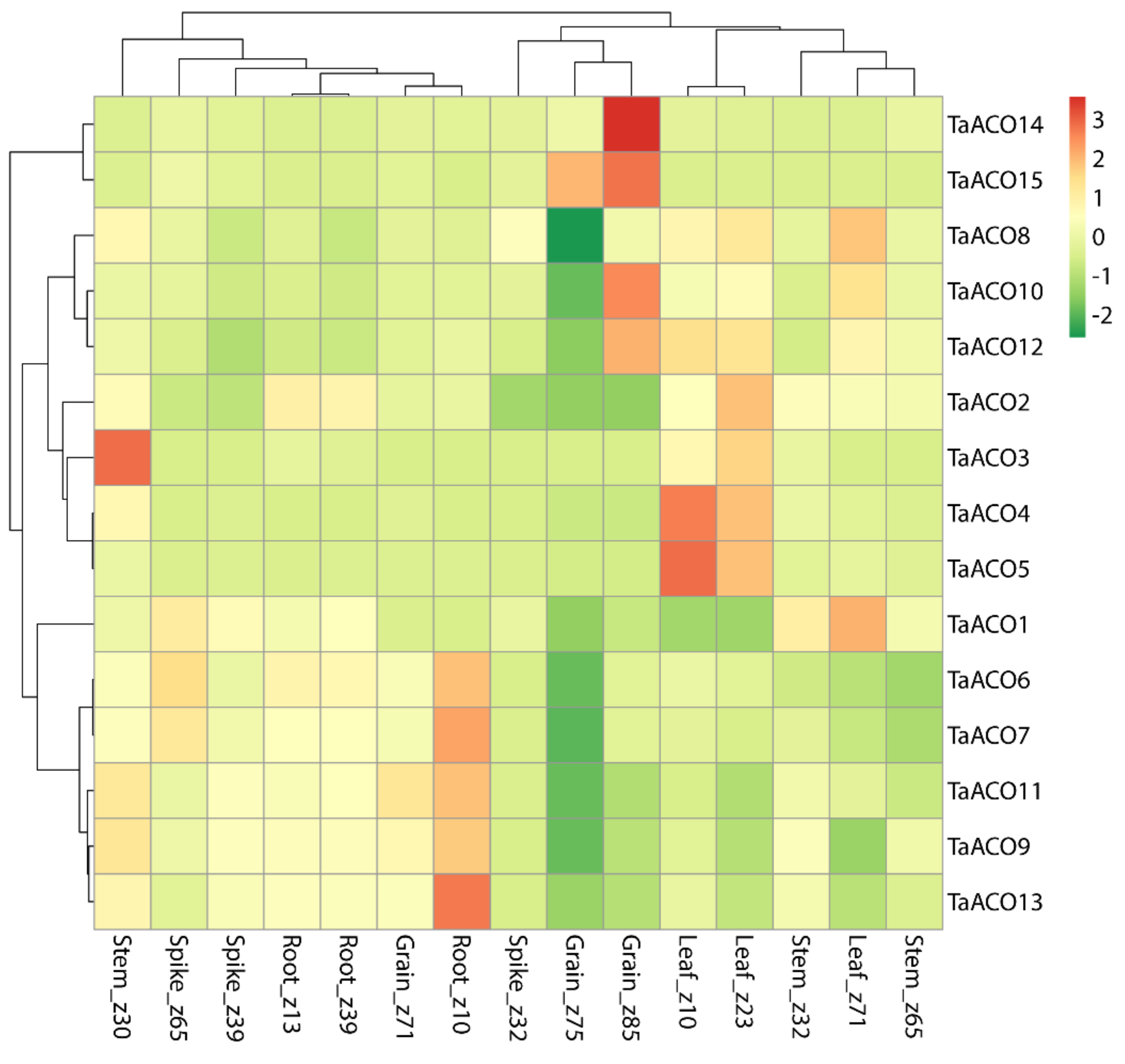
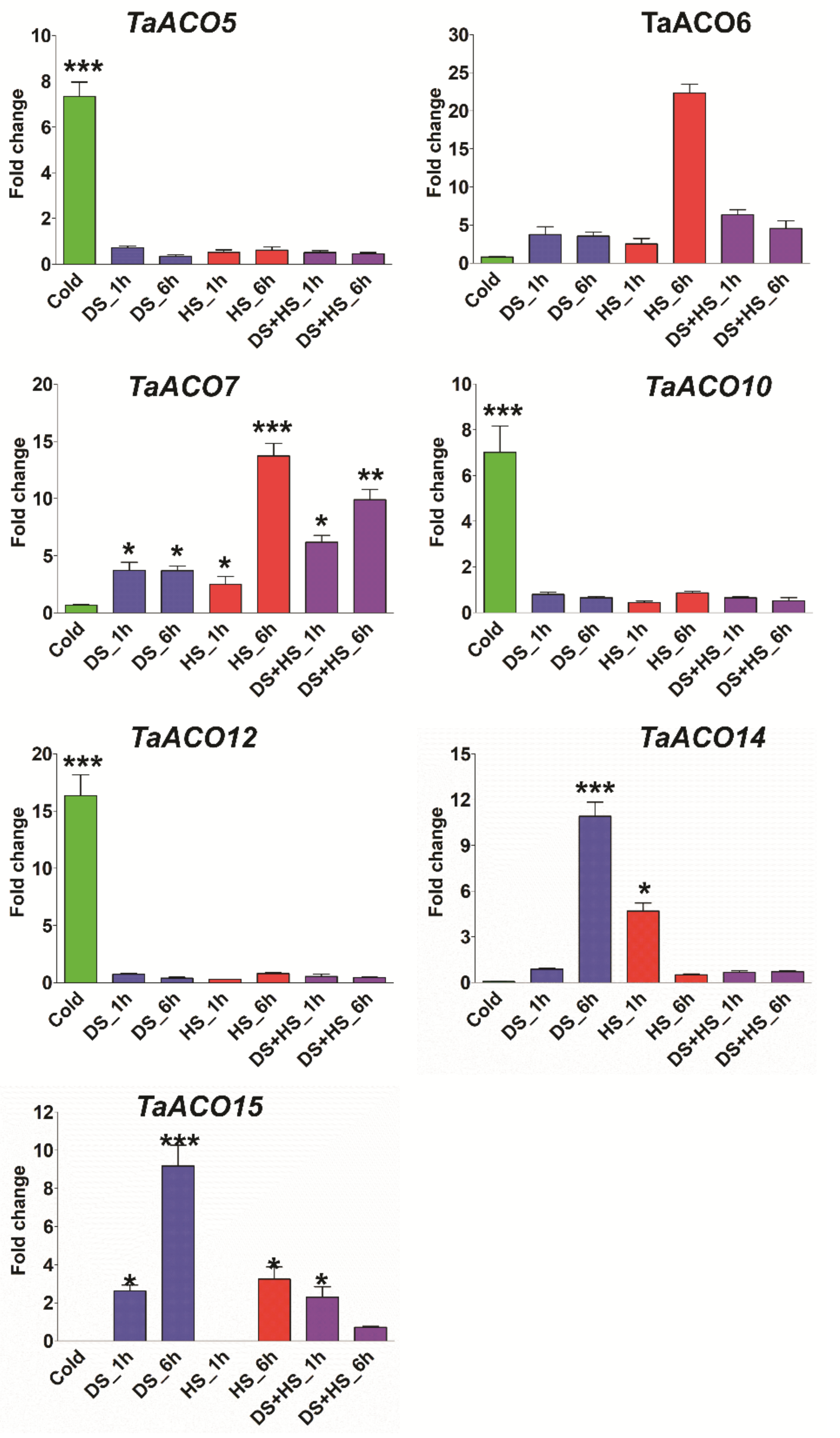
2.6. Expression Profiling of TaACO Genes in Different Tissues, Developmental Stages, and Stress Conditions
2.7. Protein–Protein Interaction Network Analysis of TaACO Genes
3. Discussion
4. Materials and Methods
4.1. Identification of ACO Genes in Wheat
4.2. Biophysical Characteristics, Subcellular Localization Genomic Localization, Gene Duplication and Synteny Analysis
4.3. Gene Structure, Protein Motif, and 3D Structure
4.4. Gene Ontology, Promoter Cis-Acting Regulatory Elements (CAREs), and Protein Interaction Network Analysis
4.5. Expression Analysis of TaACO Genes
4.6. Plant Growth, Stress Conditions, and Quantitative Reverse Transcription PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.-M.; Yang, Q.; Liu, Y.-J.; Yang, H.-L. Molecular evolution and expression divergence of the aconitase (ACO) gene family in land plants. Front. Plant Sci. 2016, 7, 1879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnarrenberger, C.; Martin, W. Evolution of the enzymes of the citric acid cycle and the glyoxylate cycle of higher plants: A case study of endosymbiotic gene transfer. Eur. J. Biochem. 2002, 269, 868–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunes-Nesi, A.; Araújo, W.L.; Obata, T.; Fernie, A.R. Regulation of the mitochondrial tricarboxylic acid cycle. Curr. Opin. Plant Biol. 2013, 16, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Peyret, P.; Perez, P.; Alric, M. Structure, Genomic Organization, and Expression of the Arabidopsis thaliana Aconitase Gene: Plant aconitase show significant homology with mammalian iron-responsive element-binding protein (∗). J. Biol. Chem. 1995, 270, 8131–8137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beinert, H.; Kennedy, M.C.; Stout, C.D. Aconitase as iron− sulfur protein, enzyme, and iron-regulatory protein. Chem. Rev. 1996, 96, 2335–2374. [Google Scholar] [CrossRef]
- Kesawat, M.S.; Das, B.K.; Kumar, M.; Bhaganagare, G.R. An overview on Fe–S protein biogenesis from prokaryotes to eukaryotes. Biol. Nitrogen Fixat. 2015, 57–74. [Google Scholar] [CrossRef]
- Kesawat, M.S.; Das, B.K.; Bhaganagare, G.R. Genome-wide identification, evolutionary and expression analyses of putative Fe–S biogenesis genes in rice (Oryza sativa). Genome 2012, 55, 571–583. [Google Scholar] [CrossRef]
- Matasova, L.; Popova, T. Aconitate hydratase of mammals under oxidative stress. Biochemistry 2008, 73, 957–964. [Google Scholar] [CrossRef]
- Lushchak, O.V.; Piroddi, M.; Galli, F.; Lushchak, V.I. Aconitase post-translational modification as a key in linkage between Krebs cycle, iron homeostasis, redox signaling, and metabolism of reactive oxygen species. Redox Rep. 2014, 19, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Hentze, M.W.; Kühn, L.C. Molecular control of vertebrate iron metabolism: mRNA-based regulatory circuits operated by iron, nitric oxide, and oxidative stress. Proc. Natl. Acad. Sci. USA 1996, 93, 8175–8182. [Google Scholar] [CrossRef]
- Navarre, D.A.; Wendehenne, D.; Durner, J.; Noad, R.; Klessig, D.F. Nitric oxide modulates the activity of tobacco aconitase. Plant Physiol. 2000, 122, 573–582. [Google Scholar] [CrossRef] [Green Version]
- Arnaud, N.; Ravet, K.; Borlotti, A.; Touraine, B.; Boucherez, J.; Fizames, C.; Briat, J.-F.; Cellier, F.; Gaymard, F. The iron-responsive element (IRE)/iron-regulatory protein 1 (IRP1)–cytosolic aconitase iron-regulatory switch does not operate in plants. Biochem. J. 2007, 405, 523–531. [Google Scholar] [CrossRef] [Green Version]
- Moeder, W.; Del Pozo, O.; Navarre, D.A.; Martin, G.B.; Klessig, D.F. Aconitase plays a role in regulating resistance to oxidative stress and cell death in Arabidopsis and Nicotiana benthamiana. Plant Mol. Biol. 2007, 63, 273–287. [Google Scholar] [CrossRef]
- Kruft, V.; Eubel, H.; Jänsch, L.; Werhahn, W.; Braun, H.-P. Proteomic approach to identify novel mitochondrial proteins in Arabidopsis. Plant Physiol. 2001, 127, 1694–1710. [Google Scholar] [CrossRef]
- Heazlewood, J.L.; Tonti-Filippini, J.S.; Gout, A.M.; Day, D.A.; Whelan, J.; Millar, A.H. Experimental analysis of the Arabidopsis mitochondrial proteome highlights signaling and regulatory components, provides assessment of targeting prediction programs, and indicates plant-specific mitochondrial proteins. Plant Cell 2004, 16, 241–256. [Google Scholar] [CrossRef] [Green Version]
- Carrari, F.; Nunes-Nesi, A.; Gibon, Y.; Lytovchenko, A.; Loureiro, M.E.; Fernie, A.R. Reduced expression of aconitase results in an enhanced rate of photosynthesis and marked shifts in carbon partitioning in illuminated leaves of wild species tomato. Plant Physiol. 2003, 133, 1322–1335. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, M.; De Bellis, L.; Alpi, A.; Nishimura, M. Cytosolic aconitase participates in the glyoxylate cycle in etiolated pumpkin cotyledons. Plant Cell Physiol. 1995, 36, 669–680. [Google Scholar]
- Eastmond, P.J.; Graham, I.A. Re-examining the role of the glyoxylate cycle in oilseeds. Trends Plant Sci. 2001, 6, 72–78. [Google Scholar] [CrossRef]
- Borek, S.; Nuc, K. Sucrose controls storage lipid breakdown on gene expression level in germinating yellow lupine (Lupinus luteus L.) seeds. J. Plant Physiol. 2011, 168, 1795–1803. [Google Scholar] [CrossRef]
- Verniquet, F.; Gaillard, J.; Neuburger, M.; Douce, R. Rapid inactivation of plant aconitase by hydrogen peroxide. Biochem. J. 1991, 276, 643–648. [Google Scholar] [CrossRef] [Green Version]
- Terol, J.; Soler, G.; Talon, M.; Cercos, M. The aconitate hydratase family from Citrus. BMC Plant Biol. 2010, 10, 222. [Google Scholar] [CrossRef]
- Pascual, J.; Rahikainen, M.; Angeleri, M.; Alegre, S.; Gossens, R.; Shapiguzov, A.; Heinonen, A.; Trotta, A.; Durian, G.; Winter, Z. ACONITASE 3 is part of theANAC017 transcription factor-dependent mitochondrial dysfunction response. Plant Physiol. 2021, 186, 1859–1877. [Google Scholar] [CrossRef] [PubMed]
- Eprintsev, A.T.; Fedorin, D.N.; Nikitina, M.V.; Igamberdiev, A.U. Expression and properties of the mitochondrial and cytosolic forms of aconitase in maize scutellum. J. Plant Physiol. 2015, 181, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Eprintsev, A.T.; Fedorin, D.N.; Cherkasskikh, M.V.; Igamberdiev, A.U. Regulation of expression of the mitochondrial and cytosolic forms of aconitase in maize leaves via phytochrome. Plant Physiol. Biochem. 2020, 146, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Sweetlove, L.; Heazlewood, J.; Herald, V.; Holtzapffel, R.; Day, D.; Leaver, C.; Millar, A. The impact of oxidative stress on Arabidopsis mitochondria. Plant J. 2002, 32, 891–904. [Google Scholar] [CrossRef]
- Kesawat, M.S.; Kherawat, B.S.; Singh, A.; Dey, P.; Kabi, M.; Debnath, D.; Saha, D.; Khandual, A.; Rout, S.; Ali, A. Genome-wide identification and characterization of the brassinazole-resistant (BZR) gene family and its expression in the various developmental stage and stress conditions in wheat (Triticum aestivum L.). Int. J. Mol. Sci. 2021, 22, 8743. [Google Scholar] [CrossRef]
- Kumar, M.; Kherawat, B.S.; Dey, P.; Saha, D.; Singh, A.; Bhatia, S.K.; Ghodake, G.S.; Kadam, A.A.; Kim, H.-U.; Chung, S.-M. Genome-wide identification and characterization of PIN-FORMED (PIN) gene family reveals role in developmental and various stress conditions in Triticum aestivum L. Int. J. Mol. Sci. 2021, 22, 7396. [Google Scholar] [CrossRef]
- Kesawat, M.S.; Kherawat, B.S.; Singh, A.; Dey, P.; Routray, S.; Mohapatra, C.; Saha, D.; Ram, C.; Siddique, K.H.; Kumar, A. Genome-Wide Analysis and Characterization of the Proline-Rich Extensin-like Receptor Kinases (PERKs) Gene Family Reveals Their Role in Different Developmental Stages and Stress Conditions in Wheat (Triticum aestivum L.). Plants 2022, 11, 496. [Google Scholar] [CrossRef]
- Consortium, I.W.G.S.; Appels, R.; Eversole, K.; Stein, N.; Feuillet, C.; Keller, B.; Rogers, J.; Pozniak, C.J.; Choulet, F.; Distelfeld, A. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar]
- Shiferaw, B.; Smale, M.; Braun, H.-J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global Food Security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef] [Green Version]
- Macauley, H.; Ramadjita, T. Cereal Crops: Rice, Maize, Millet, Sorghum, Wheat; Abdou Diouf International Conference Centre: Sebi Ponty, Senegal, 2015. [Google Scholar]
- Joshi, A.; Mishra, B.; Chatrath, R.; Ortiz Ferrara, G.; Singh, R.P. Wheat improvement in India: Present status, emerging challenges and future prospects. Euphytica 2007, 157, 431–446. [Google Scholar] [CrossRef]
- Kumar, P.; Yadava, R.; Gollen, B.; Kumar, S.; Verma, R.K.; Yadav, S. Nutritional contents and medicinal properties of wheat: A review. Life Sci. Med. Res. 2011, 22, 1–10. [Google Scholar]
- Sharma, I.; Tyagi, B.; Singh, G.; Venkatesh, K.; Gupta, O. Enhancing wheat production-A global perspective. Indian J. Agricul. Sci. 2015, 85, 3–13. [Google Scholar]
- Laghari, K.A.; Sial, M.A.; Arain, M.A. Effect of high temperature stress on grain yield and yield components of wheat (Triticum aestivum L.). J. Sci. Technol. Devel. 2012, 31, 83–90. [Google Scholar]
- Afzal, F.; Chaudhari, S.K.; Gul, A.; Farooq, A.; Ali, H.; Nisar, S.; Sarfraz, B.; Shehzadi, K.J.; Mujeeb-Kazi, A. Bread wheat (Triticum aestivum L.) under biotic and abiotic stresses: An overview. Crop Prod. Glob. Environ. Issues 2015, 293–317. [Google Scholar] [CrossRef]
- Limbalkar, O.M.; Meena, K.; Singh, M.; Sunilkumar, V. Genetic improvement of wheat for biotic and abiotic stress tolerance. Int J Curr Microbiol App Sci 2018, 7, 1962–1971. [Google Scholar] [CrossRef]
- He, Y.; Mawhinney, T.P.; Preuss, M.L.; Schroeder, A.C.; Chen, B.; Abraham, L.; Jez, J.M.; Chen, S. A redox-active isopropylmalate dehydrogenase functions in the biosynthesis of glucosinolates and leucine in Arabidopsis. Plant J. 2009, 60, 679–690. [Google Scholar] [CrossRef]
- He, Y.; Chen, L.; Zhou, Y.; Mawhinney, T.P.; Chen, B.; Kang, B.H.; Hauser, B.A.; Chen, S. Functional characterization of Arabidopsis thaliana isopropylmalate dehydrogenases reveals their important roles in gametophyte development. New Phytol. 2011, 189, 160–175. [Google Scholar] [CrossRef]
- Wiegand, G.; Remington, S.J. Citrate synthase: Structure, control, and mechanism. Annu. Rev. Biophys. Biophys. Chem. 1986, 15, 97–117. [Google Scholar] [CrossRef]
- La Cognata, U.; Landschütze, V.; Willmitzer, L.; Müller-Röber, B. Structure and expression of mitochondrial citrate synthases from higher plants. Plant Cell Physiol. 1996, 37, 1022–1029. [Google Scholar] [CrossRef] [Green Version]
- Lespinet, O.; Wolf, Y.I.; Koonin, E.V.; Aravind, L. The role of lineage-specific gene family expansion in the evolution of eukaryotes. Genome Res. 2002, 12, 1048–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawton-Rauh, A. Evolutionary dynamics of duplicated genes in plants. Mol. Phylogenet. Evol. 2003, 29, 396–409. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faraji, S.; Heidari, P.; Amouei, H.; Filiz, E. Abdullah, and Poczai, P. Investigation and computational analysis of the sulfotransferase (Sot) gene family in potato (solanum tuberosum): Insights into sulfur adjustment for proper development and stimuli responses. Plants 2021, 10, 2597. [Google Scholar] [CrossRef] [PubMed]
- Heidari, P.; Faraji, S.; Poczai, P. Magnesium transporter gene family: Genome-wide identification and characterization in Theobroma cacao, Corchorus capsularis, and Gossypium hirsutum of family malvaceae. Agronomy 2021, 11, 1651. [Google Scholar] [CrossRef]
- William Roy, S.; Gilbert, W. The evolution of spliceosomal introns: Patterns, puzzles and progress. Nat. Rev. Genet. 2006, 7, 211–221. [Google Scholar] [CrossRef]
- Roy, S.W.; Penny, D. Patterns of intron loss and gain in plants: Intron loss–dominated evolution and genome-wide comparison of O. sativa and A. thaliana. Mol. Biol. Evol. 2007, 24, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Lyu, H.M.; Zhu, K.; Van de Peer, Y.; Cheng, Z.M. The emergence and evolution of intron-poor and intronless genes in intron-rich plant gene families. Plant J. 2021, 105, 1072–1082. [Google Scholar] [CrossRef]
- Lecharny, A.; Boudet, N.; Gy, I.; Aubourg, S.; Kreis, M. Introns in, introns out in plant gene families: A genomic approach of the dynamics of gene structure. J. Struct. Funct. Genom. 2003, 3, 111–116. [Google Scholar] [CrossRef]
- Yang, Z.; Gong, Q.; Wang, L.; Jin, Y.; Xi, J.; Li, Z.; Qin, W.; Yang, Z.; Lu, L.; Chen, Q. Genome-wide study of YABBY genes in upland cotton and their expression patterns under different stresses. Front. Genet. 2018, 9, 33. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Garcia, C.M.; Finer, J.J. Identification and validation of promoters and cis-acting regulatory elements. Plant Sci. 2014, 217, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.L.; Sen, R.; Roeder, R.G. Enhancer–promoter communication and transcriptional regulation of Igh. Trends Immunol. 2011, 32, 532–539. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.L.; Singer, D.S. Core promoters in transcription: Old problem, new insights. Trends Biochem. Sci. 2015, 40, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wong, C.-H.; Birnbaum, R.Y.; Li, G.; Favaro, R.; Ngan, C.Y.; Lim, J.; Tai, E.; Poh, H.M.; Wong, E. Chromatin connectivity maps reveal dynamic promoter–enhancer long-range associations. Nature 2013, 504, 306–310. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Tang, Q.; Li, Y.; Zhang, Y.; Zhao, C.; Yan, J.; You, H. Folding/unfolding kinetics of G-quadruplexes upstream of the P1 promoter of the human BCL-2 oncogene. J. Biol. Chem. 2019, 294, 5890–5895. [Google Scholar] [CrossRef]
- Fankhauser, C.; Chory, J. Light control of plant development. Annu. Rev. Cell Dev. Biol. 1997, 13, 203–229. [Google Scholar] [CrossRef] [Green Version]
- Rasmusson, A.G.; Escobar, M.A. Light and diurnal regulation of plant respiratory gene expression. Physiol. Plant. 2007, 129, 57–67. [Google Scholar] [CrossRef]
- Tepperman, J.M.; Hudson, M.E.; Khanna, R.; Zhu, T.; Chang, S.H.; Wang, X.; Quail, P.H. Expression profiling of phyB mutant demonstrates substantial contribution of other phytochromes to red-light-regulated gene expression during seedling de-etiolation. Plant J. 2004, 38, 725–739. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Marekov, L.; Bubber, P.; Browne, S.E.; Stavrovskaya, I.; Lee, J.; Steinert, P.M.; Blass, J.P.; Beal, M.F.; Gibson, G.E. Mitochondrial aconitase is a transglutaminase 2 substrate: Transglutamination is a probable mechanism contributing to high-molecular-weight aggregates of aconitase and loss of aconitase activity in Huntington disease brain. Neurochem. Res. 2005, 30, 1245–1255. [Google Scholar] [CrossRef]
- Regev-Rudzki, N.; Karniely, S.; Ben-Haim, N.N.; Pines, O. Yeast aconitase in two locations and two metabolic pathways: Seeing small amounts is believing. Mol. Biol. Cell 2005, 16, 4163–4171. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.J.; Wang, X.; Butow, R.A. Yeast aconitase binds and provides metabolically coupled protection to mitochondrial DNA. Proc. Natl. Acad. Sci. USA 2007, 104, 13738–13743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Z.; Tsuda, M.; Kishita, Y.; Sato, Y.; Aigaki, T. Impaired energy metabolism in a Drosophila model of mitochondrial aconitase deficiency. Biochem. Biophys. Res. Commun. 2013, 433, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Fernie, A.R.; Carrari, F.; Sweetlove, L.J. Respiratory metabolism: Glycolysis, the TCA cycle and mitochondrial electron transport. Curr. Opin. Plant Biol. 2004, 7, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Bota, D.A.; Van Remmen, H.; Davies, K.J. Modulation of Lon protease activity and aconitase turnover during aging and oxidative stress. FEBS Lett. 2002, 532, 103–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanlerberghe, G.C.; Mclntosh, L. Signals regulating the expression of the nuclear gene encoding alternative oxidase of plant mitochondria. Plant Physiol. 1996, 111, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Hunter, S.; Apweiler, R.; Attwood, T.K.; Bairoch, A.; Bateman, A.; Binns, D.; Bork, P.; Das, U.; Daugherty, L.; Duquenne, L. InterPro: The integrative protein signature database. Nucleic Acids Res. 2009, 37 (Suppl. S1), D211–D215. [Google Scholar] [CrossRef] [Green Version]
- Bilmes, J.A. What HMMs can do. IEICE Trans. Inf. Syst. 2006, 89, 869–891. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Kozlowski, L.P. IPC–isoelectric point calculator. Biol. Direct 2016, 11, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.K.; Kesawat, M.S.; Hong, C.B. One gene member of the ADP-ribosylation factor family is heat-inducible and enhances seed germination in Nicotiana tabacum. Genes Genom. 2017, 39, 1353–1365. [Google Scholar] [CrossRef]
- Kesawat, M.S.; Kim, D.K.; Zeba, N.; Suh, M.C.; Xia, X.; Hong, C.B. Ectopic RING zinc finger gene from hot pepper induces totally different genes in lettuce and tobacco. Mol. Breed. 2018, 38, 1–24. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Narancio, R.; John, U.; Mason, J.; Spangenberg, G. Selection of optimal reference genes for quantitative RT-PCR transcript abundance analysis in white clover (Trifolium repens L.). Funct. Plant Biol. 2018, 45, 737–744. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proposed Gene Name | Gene ID | Genomic Location | Orientation | CDS Length (bp) | Protein Length (aa) | Protein Name | Molecular Weight (KDa) | Isoelectric Point (pI) | GRAVY | Predicted Subcellular Localization |
|---|---|---|---|---|---|---|---|---|---|---|
| TaACO1 | TraesCS2B02G181900 | 2B:156914707-156922970 | Reverse | 1506 | 501 | Hypothetical protein CFC21_022353 | 54.79 | 6.09 | −0.202 | mitochondrion |
| TaACO2 | TraesCS3A02G184000 | 3A:214122773-214123221 | Reverse | 3024 | 1007 | Putative aconitate hydratase, cytoplasmic isoform | 109.32 | 6.50 | −0.132 | chloroplast thylakoid membrane |
| TaACO3 | TraesCS3B02G213800 | 3B:253633139-253633475 | Reverse | 1971 | 656 | Putative aconitate hydratase, cytoplasmic | 71.29 | 5.92 | −0.155 | cytoplasm |
| TaACO4 | TraesCS3B02G213900 | 3B:253666186-253666845 | Reverse | 3015 | 1004 | Putative aconitate hydratase, cytoplasmic isoform | 108.95 | 6.59 | −0.153 | chloroplast thylakoid membrane |
| TaACO5 | TraesCS3D02G188200 | 3D:174907841-174908490 | Reverse | 3015 | 1004 | Putative aconitate hydratase, cytoplasmic isoform | 108.89 | 6.38 | −0.123 | mitochondrion |
| TaACO6 | TraesCS4B02G335100 | 4B:626011015-626011628 | Reverse | 2400 | 799 | Putative aconitate hydratase, cytoplasmic | 86.7 | 5.98 | −0.073 | extracellular space |
| TaACO7 | TraesCS5A02G505000 | 5A:670410810-670411419 | Reverse | 2988 | 995 | Putative aconitate hydratase, cytoplasmic | 106.93 | 6.60 | −0.128 | chloroplast thylakoid membrane |
| TaACO8 | TraesCS6A02G080600 | 6A:49189960-49195152 | Forward | 1533 | 510 | 3-isopropylmalate dehydratase large subunit, chloroplastic-like | 55.16 | 6.43 | −0.218 | chloroplast thylakoid membrane |
| TaACO9 | TraesCS6A02G400000 | 6A:609449354-609450370 | Forward | 2931 | 976 | Putative aconitate hydratase, cytoplasmic | 106.2 | 6.41 | −0.184 | chloroplast thylakoid lumen |
| TaACO10 | TraesCS6B02G108100 | 6B:88711659-88715726 | Forward | 1434 | 477 | Hypothetical protein CFC21_101948 | 51.92 | 5.97 | −0.221 | chloroplast thylakoid membrane |
| TaACO11 | TraesCS6B02G440200 | 6B:705380681-705381704 | Forward | 2934 | 977 | Putative aconitate hydratase, cytoplasmic | 106.31 | 6.37 | −0.209 | chloroplast |
| TaACO12 | TraesCS6D02G074300 | 6D:37921196-37926065 | Forward | 1533 | 510 | 3-isopropylmalate dehydratase large subunit, chloroplastic-like | 55.13 | 6.43 | −0.227 | chloroplast thylakoid membrane |
| TaACO13 | TraesCS6D02G384200 | 6D:462627391-462628393 | Forward | 2928 | 975 | Putative aconitate hydratase, cytoplasmic | 106.15 | 6.35 | −0.174 | chloroplast thylakoid lumen |
| TaACO14 | TraesCS7A02G219100 | 7A:186527123-186528324 | Reverse | 2715 | 904 | Aconitate hydratase, cytoplasmic-like | 99.8 | 5.67 | −0.145 | cytoplasm |
| TaACO15 | TraesCS7D02G223000 | 7D:184190377-184191564 | Forward | 2067 | 688 | Aconitate hydratase, cytoplasmic isoform | 75.71 | 6.19 | −0.156 | cytoplasm |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kesawat, M.S.; Kherawat, B.S.; Ram, C.; Singh, A.; Dey, P.; Gora, J.S.; Misra, N.; Chung, S.-M.; Kumar, M. Genome-Wide Identification and Expression Profiling of Aconitase Gene Family Members Reveals Their Roles in Plant Development and Adaptation to Diverse Stress in Triticum aestivum L. Plants 2022, 11, 3475. https://doi.org/10.3390/plants11243475
Kesawat MS, Kherawat BS, Ram C, Singh A, Dey P, Gora JS, Misra N, Chung S-M, Kumar M. Genome-Wide Identification and Expression Profiling of Aconitase Gene Family Members Reveals Their Roles in Plant Development and Adaptation to Diverse Stress in Triticum aestivum L. Plants. 2022; 11(24):3475. https://doi.org/10.3390/plants11243475
Chicago/Turabian StyleKesawat, Mahipal Singh, Bhagwat Singh Kherawat, Chet Ram, Anupama Singh, Prajjal Dey, Jagan Singh Gora, Namrata Misra, Sang-Min Chung, and Manu Kumar. 2022. "Genome-Wide Identification and Expression Profiling of Aconitase Gene Family Members Reveals Their Roles in Plant Development and Adaptation to Diverse Stress in Triticum aestivum L." Plants 11, no. 24: 3475. https://doi.org/10.3390/plants11243475