
Hematopoietic Effects of Angelica gigas Nakai Extract on Cyclophosphamide-Induced Myelosuppression

 ,
,
Abstract
:1. Introduction
2. Results
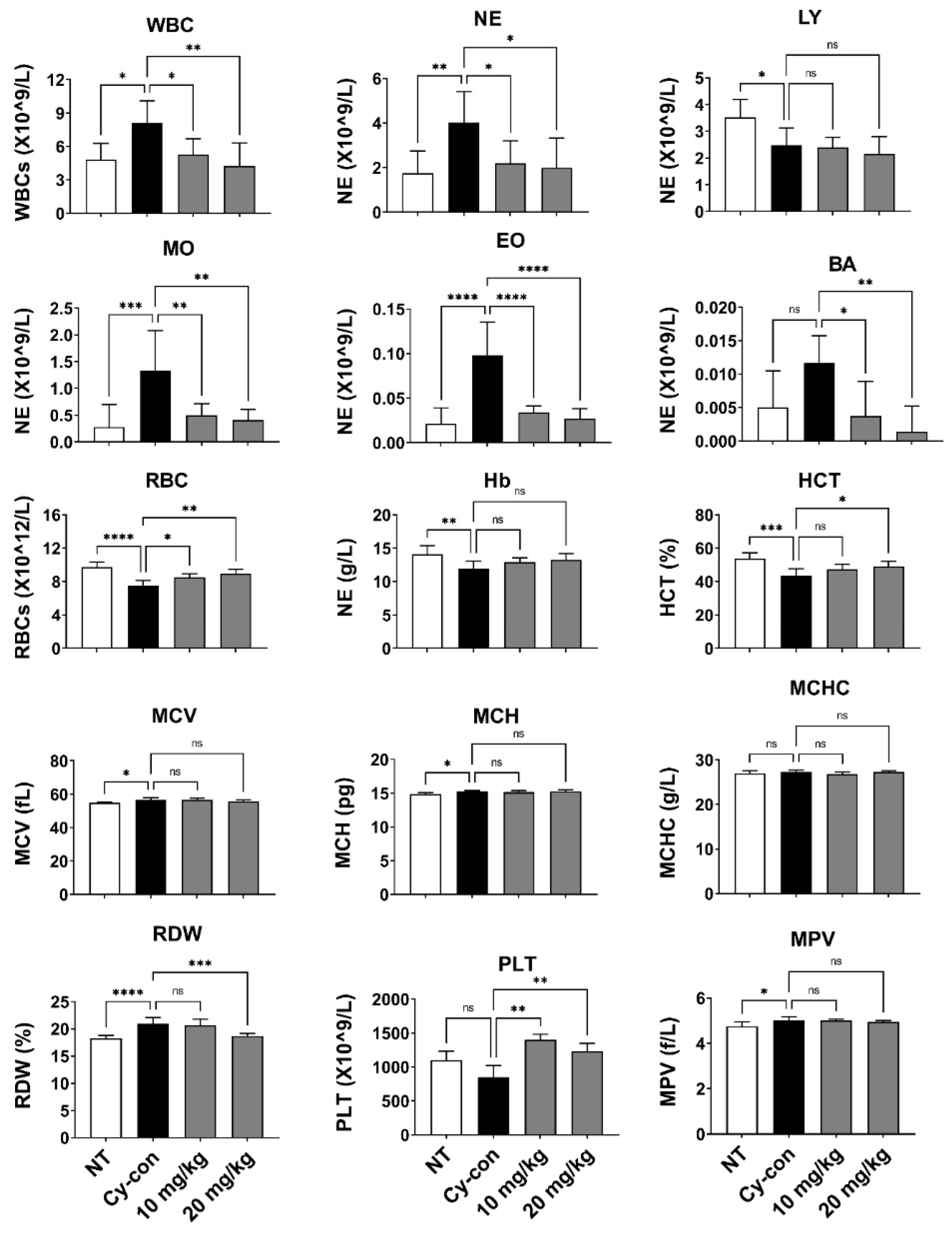
2.1. Characteristics of Myelosuppressive Mice and the Effect of AGNEX in Peripheral Blood
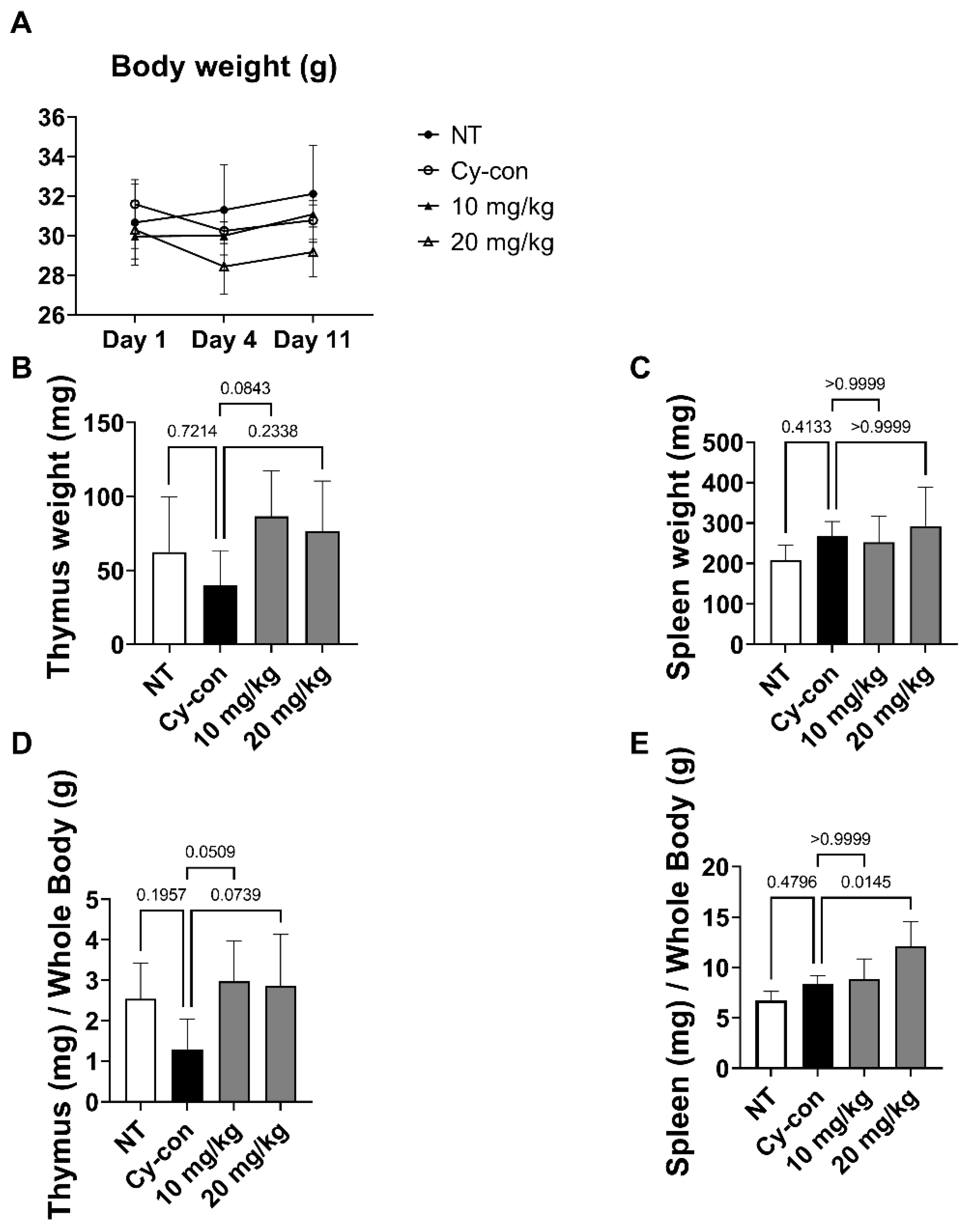
2.2. Effect of AGNEX on the Thymus and Spleen
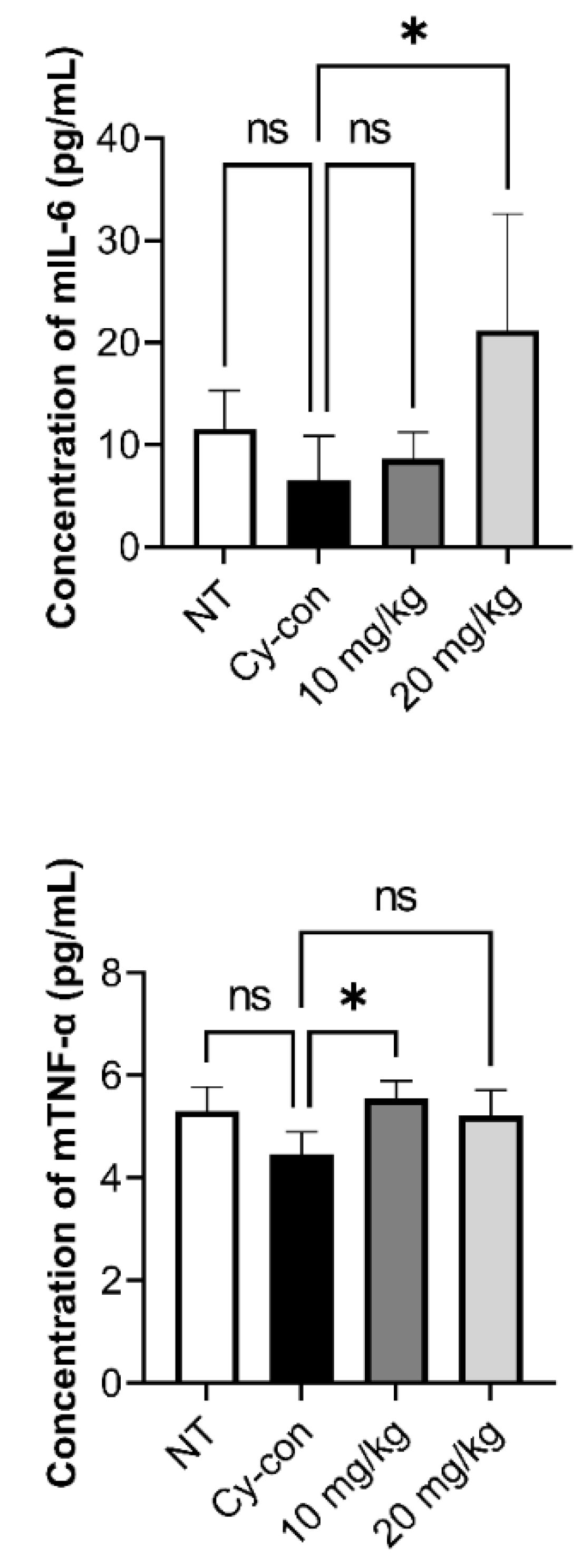
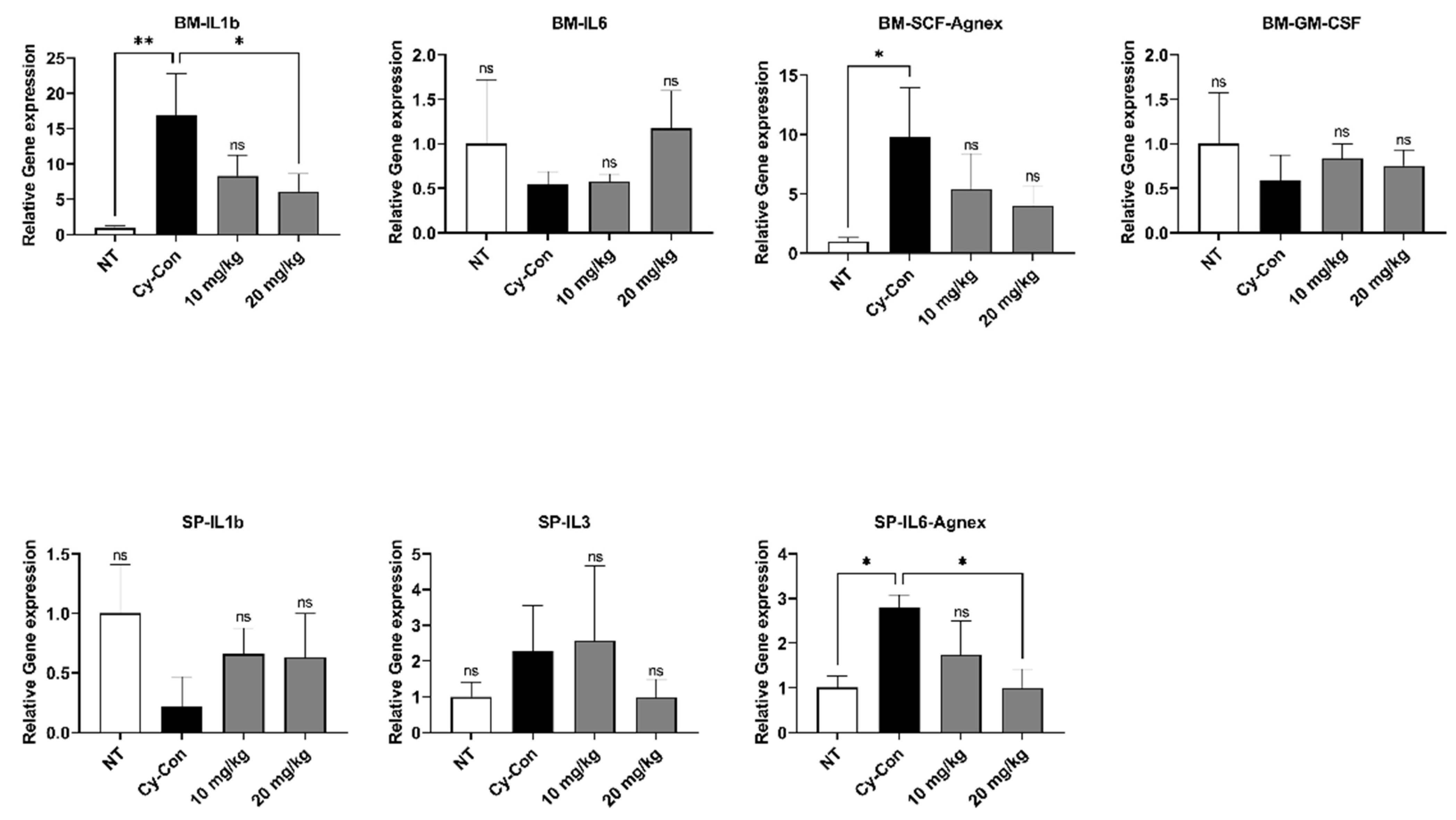
2.3. Correlation between AGNEX and Hematopoietic Factors
2.4. Confirmation of Cell Proliferation and Viability of BM and Splenocytes
2.5. Validation of the AGNEX Efficacy in Erythroleukemia-Type Cells
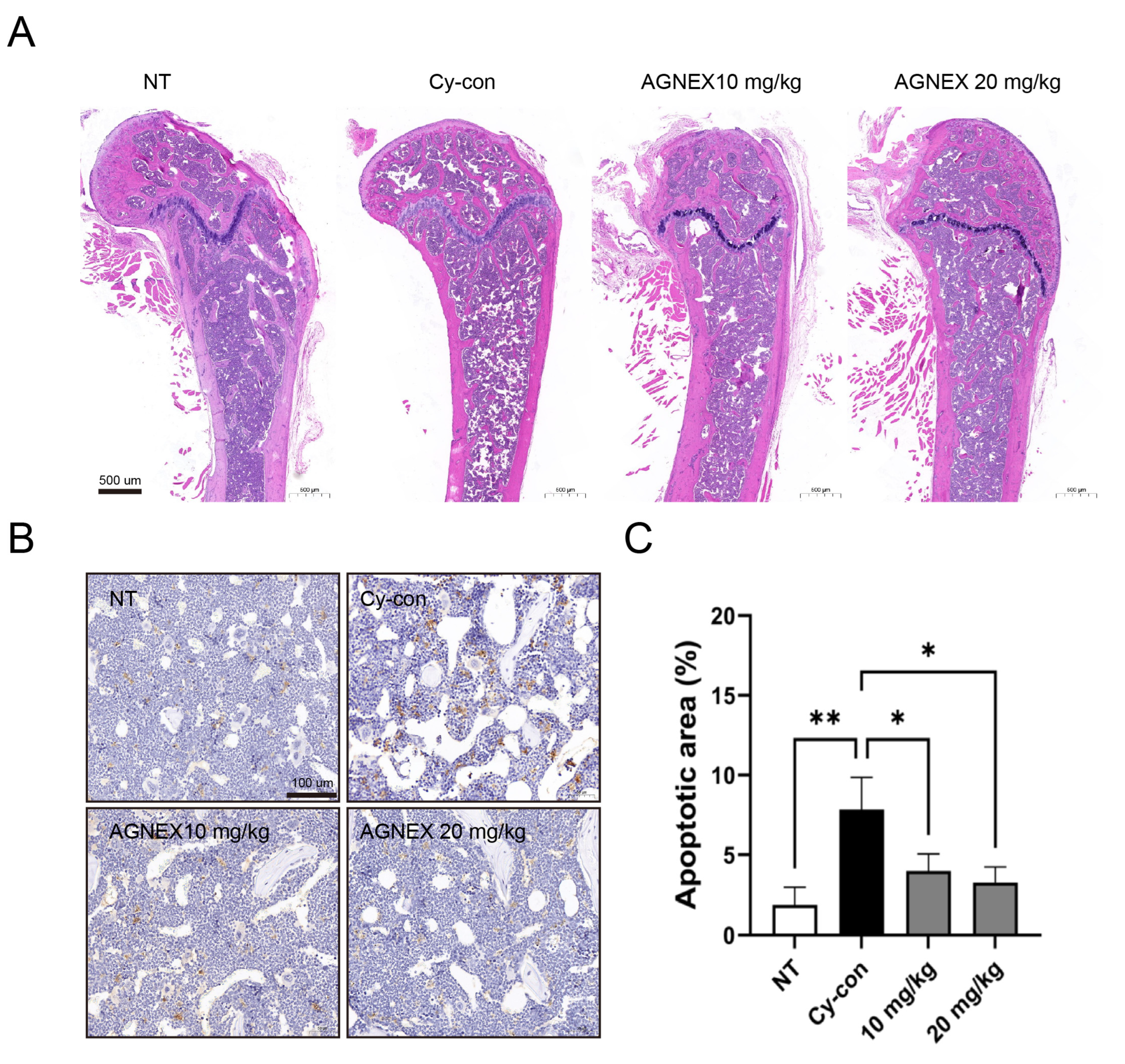
2.6. Validation of the AGNEX Ability to Recover and Differentiate in the Femoral Bone Marrow
3. Discussion
4. Materials and Methods
4.1. Preparation of Angelica gigas Nakai Extract (AGNEX)
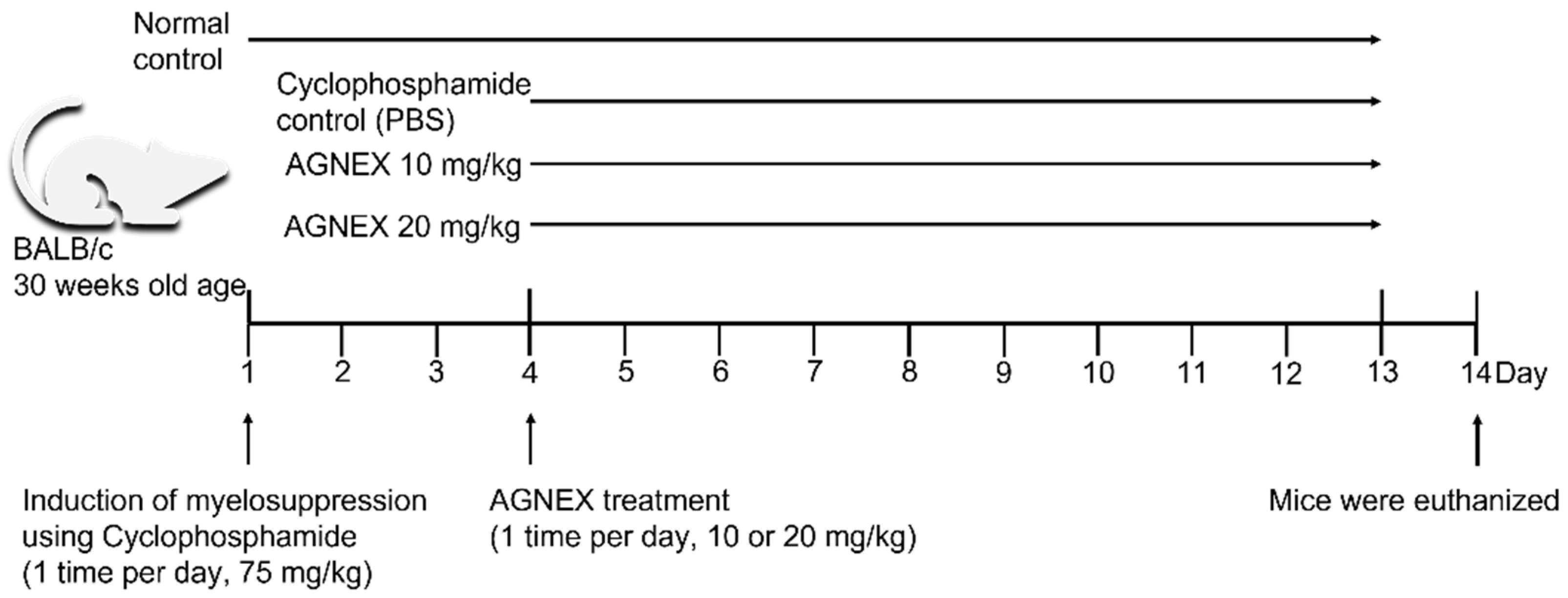
4.2. Animal Experimental Design
4.3. Analysis of Mouse Peripheral Blood
4.4. Histological Analysis
4.5. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.6. Enzyme-Linked Immunosorbent Assay (ELISA)
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hong, S.; Won, Y.J.; Lee, J.J.; Jung, K.W.; Kong, H.J.; Im, J.S.; Seo, H.G.; The Community of Population-Based Regional Cancer Registries. Cancer Statistics in Korea: Incidence, Mortality, Survival, and Prevalence in 2018. Cancer Res. Treat. 2021, 53, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.K.; Park, S.M.; Ahn, J.Y.; Kim, O.H.; Kim, Y.W.; Park, K.-I.; Park, S.-D.; Lee, J.-H. Anticancer activity of herbal formula Jisilhaebaekgyeji-Tang against human breast cancer cells and its mechanism. Mol. Cell. Toxicol. 2022. [Google Scholar] [CrossRef]
- Li, S.; Zhao, S.; Guo, Y.; Yang, Y.; Huang, J.; Wang, J.; Lu, S.; Wang, B.; Chai, C.; Xu, Z.; et al. Clinical Efficacy and Potential Mechanisms of Acupoint Stimulation Combined with Chemotherapy in Combating Cancer: A Review and Prospects. Front. Oncol. 2022, 12, 864046. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Park, P.S.; Jeong, D.H.; Sim, S.; Kim, N.; Park, H.; Jeon, K.S.; Um, Y.; Kim, M.-J. The characteristics of the growth and the active compounds of Angelica gigas Nakai in cultivation sites. Plants 2020, 9, 823. [Google Scholar] [CrossRef]
- Lee, T.K.; Kang, I.J.; Sim, H.; Lee, J.C.; Ahn, J.H.; Kim, D.W.; Park, J.H.; Lee, C.H.; Kim, J.D.; Won, M.H.; et al. Therapeutic Effects of Decursin and Angelica gigas Nakai Root Extract in Gerbil Brain after Transient Ischemia via Protecting BBB Leakage and Astrocyte Endfeet Damage. Molecules 2021, 26, 2161. [Google Scholar] [CrossRef]
- Madgula, V.L.; Avula, B.; Reddy, V.L.N.; Khan, I.A.; Khan, S.I. Transport of decursin and decursinol angelate across Caco-2 and MDR-MDCK cell monolayers: In vitro models for intestinal and blood-brain barrier permeability. Planta Med. 2007, 73, 330–335. [Google Scholar] [CrossRef]
- Lu, J.; Kim, S.H.; Jiang, C.; Lee, H.; Guo, J. Oriental herbs as a source of novel anti-androgen and prostate cancer chemopreventive agents. Acta Pharmacol. Sin. 2007, 28, 1365–1372. [Google Scholar] [CrossRef]
- Kiyonga, A.N.; An, J.H.; Lee, K.Y.; Lim, C.; Suh, Y.G.; Chin, Y.W.; Jung, K. Rapid and Efficient Separation of Decursin and Decursinol Angelate from Angelica gigas Nakai using Ionic Liquid, (BMIm)BF4, Combined with Crystallization. Molecules 2019, 24, 2390. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.H.; Sik Bang, S.; Seok Choi, J.; Han, H.; Kim, I.H. Involvement of PKC and ROS in the cytotoxic mechanism of anti-leukemic decursin and its derivatives and their structure-activity relationship in human K562 erythroleukemia and U937 myeloleukemia cells. Cancer Lett. 2005, 223, 191–201. [Google Scholar] [CrossRef]
- Kim, E.; Nam, J.; Chang, W.; Zulfugarov, I.S.; Okhlopkova, Z.M.; Olennikov, D.; Chirikova, N.K.; Kim, S.W. Angelica gigas Nakai and Decursin Downregulate Myc Expression to Promote Cell Death in B-cell Lymphoma. Sci. Rep. 2018, 8, 10590. [Google Scholar] [CrossRef]
- Lee, H.W.; Kim, H.; Ryuk, J.A.; Kil, K.J.; Ko, B.S. Hemopoietic effect of extracts from constituent herbal medicines of Samul-tang on phenylhydrazine-induced hemolytic anemia in rats. Int. J. Clin. Exp. Pathol. 2014, 7, 6179–6185. [Google Scholar] [PubMed]
- Liu, J.; Wei, J.; Wang, C.; Meng, X.; Chen, H.; Deng, P.; Huandike, M.; Zhang, H.; Li, X.; Chai, L. The combination of Radix Astragali and Radix Angelicae Sinensis attenuates the IFN-gamma-induced immune destruction of hematopoiesis in bone marrow cells. BMC Complement. Altern. Med. 2019, 19, 356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.Q.; Yue, S.J.; Tang, Y.P.; Chen, Y.Y.; Zhou, G.S.; Zhang, J.; Zhu, Z.H.; Liu, P.; Duan, J.A. A network pharmacology approach to investigate the blood enriching mechanism of Danggui buxue Decoction. J. Ethnopharmacol. 2019, 235, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Tang, R.; Chen, L.B.; Zhang, K.S.; Huang, X.P.; Deng, C.Q. Effects of Astragalus Combined with Angelica on Bone Marrow Hematopoiesis Suppression Induced by Cyclophosphamide in Mice. Biol. Pharm. Bull 2017, 40, 598–609. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Bei, H.; Du, R.; Chen, Q.; Wu, F.; Chen, J.; Bo, H. Metabolomic analysis of serum reveals the potential effective ingredients and pathways of Danggui Buxue Tang in promoting erythropoiesis. Complement. Ther. Med. 2020, 48, 102247. [Google Scholar] [CrossRef]
- Randolph-Habecker, J.; Iwata, M.; Torok-Storb, B. Cytomegalovirus mediated myelosuppression. J. Clin. Virol. 2002, 25 (Suppl. 2), S51–S56. [Google Scholar] [CrossRef]
- Barreto, J.N.; McCullough, K.B.; Ice, L.L.; Smith, J.A. Antineoplastic agents and the associated myelosuppressive effects: A review. J. Pharm. Pract. 2014, 27, 440–446. [Google Scholar] [CrossRef]
- Ba, Y.; Shi, Y.; Jiang, W.; Feng, J.; Cheng, Y.; Xiao, L.; Zhang, Q.; Qiu, W.; Xu, B.; Xu, R.; et al. Current management of chemotherapy-induced neutropenia in adults: Key points and new challenges: Committee of Neoplastic Supportive-Care (CONS), China Anti-Cancer Association Committee of Clinical Chemotherapy, China Anti-Cancer Association. Cancer Biol. Med. 2020, 17, 896–909. [Google Scholar] [CrossRef]
- Jagannathan-Bogdan, M.; Zon, L.I. Hematopoiesis. Development 2013, 140, 2463–2467. [Google Scholar] [CrossRef] [Green Version]
- Keohane, E.M.; Smith, L.; Walenga, J. Rodak’s Hematology: Clinical Principles and Applications, 5th ed.; Saunders: Philadelphia, PA, USA, 2015. [Google Scholar]
- Liu, M.; Tan, H.; Zhang, X.; Liu, Z.; Cheng, Y.; Wang, D.; Wang, F. Hematopoietic effects and mechanisms of Fufang ejiao jiang on radiotherapy and chemotherapy-induced myelosuppressed mice. J. Ethnopharmacol. 2014, 152, 575–584. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, L.; Yang, C.; Zhang, Y.; Li, J.; Zhang, X.; Chen, J.; Huang, B.; Zhao, D.; Li, X.; et al. Prevention Effect of Protopanaxadiol-Type Saponins Saponins and Protopanaxatriol-Type Saponins on Myelosuppression Mice Induced by Cyclophosphamide. Front. Pharmacol. 2022, 13, 845034. [Google Scholar] [CrossRef] [PubMed]
- Willard-Mack, C.L.; Elmore, S.A.; Hall, W.C.; Harleman, J.; Kuper, C.F.; Losco, P.; Rehg, J.E.; Ruhl-Fehlert, C.; Ward, J.M.; Weinstock, D.; et al. Nonproliferative and Proliferative Lesions of the Rat and Mouse Hematolymphoid System. Toxicol. Pathol. 2019, 47, 665–783. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, L.; Yang, Z.; Wang, J.; Li, W.; Zhou, J.; Zhang, J. Hematopoietic Effects of Paeoniflorin and Albiflorin on Radiotherapy-Induced Myelosuppression Mice. Evid. Based Complement. Alternat. Med. 2016, 2016, 5789381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.; Wang, Y.; Cai, E.; Zhang, L.; Zhao, Y.; Sun, N.; Zheng, X.; Wang, S. Study of the Effects and Mechanisms of Ginsenoside Compound K on Myelosuppression. J. Agric. Food Chem. 2019, 67, 1402–1408. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Passegue, E. TNF-alpha Coordinates Hematopoietic Stem Cell Survival and Myeloid Regeneration. Cell Stem. Cell 2019, 25, 357–372.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodish, H.F.; Lodish, H.F. Molecular Cell Biology, 6th ed.; W.H. Freeman: New York, NY, USA; Basingstoke, UK, 2008. [Google Scholar]
- Rang, H.P.; Rang, H.P.; Dale, M.M. Rang & Dale’s Pharmacology, 6th ed.; Illustrations; Churchill Livingstone: Edinburgh, UK, 2007; 829p. [Google Scholar]
- Davidoff, A.N.; Mendelow, B.V. Cell-cycle disruptions and apoptosis induced by the cyclophosphamide derivative mafosfamide. Exp. Hematol. 1993, 21, 922–927. [Google Scholar]
- Patel, P.B.; Thakkar, V.R.; Patel, J.S. Cellular Effect of Curcumin and Citral Combination on Breast Cancer Cells: Induction of Apoptosis and Cell Cycle Arrest. J. Breast Cancer 2015, 18, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Mahecha, A.; Hales, B.F.; Robaire, B. Effects of acute and chronic cyclophosphamide treatment on meiotic progression and the induction of DNA double-strand breaks in rat spermatocytes. Biol. Reprod. 2005, 72, 1297–1304. [Google Scholar] [CrossRef] [Green Version]
- Kano, Y.; Akutsu, M.; Tsunoda, S.; Mano, H.; Sato, Y.; Honma, Y.; Furukawa, Y. In vitro cytotoxic effects of a tyrosine kinase inhibitor STI571 in combination with commonly used antileukemic agents. Blood 2001, 97, 1999–2007. [Google Scholar] [CrossRef] [Green Version]
- Peters, R.H.; Jollow, D.J.; Stuart, R.K. Role of glutathione in the in vitro synergism between 4-hydroperoxy-cyclophosphamide and cisplatin in leukemia cell lines. Cancer Res. 1991, 51, 2536–2541. [Google Scholar]
- Ahn, M.-J.; Lee, M.K.; Kim, Y.C.; Sung, S.H. The simultaneous determination of coumarins in Angelica gigas root by high performance liquid chromatography–diode array detector coupled with electrospray ionization/mass spectrometry. J. Pharm. Biomed. Anal. 2008, 46, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, L.; Jiang, C.; Xing, C.; Kim, S.-H.; Lu, J. Anti-cancer and other bioactivities of Korean Angelica gigas Nakai (AGN) and its major pyranocoumarin compounds. Anti-Cancer Agents Med. Chem. (Former. Curr. Med. Chem.-Anti-Cancer Agents) 2012, 12, 1239–1254. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Kang, J.S. Antithrombotic Potential of Red Allium cepa and Angelica gigas Nakai. Indian J. Pharm. Educ. Res. 2021, 55, 837–845. [Google Scholar] [CrossRef]
- Kim, K.-M.; Kim, T.-H.; Park, Y.-J.; Kim, I.-H.; Kang, J.-S. Evaluation of the genotoxicity of decursin and decursinol angelate produced by Angelica gigas Nakai. Mol. Cell. Toxicol. 2009, 5, 83–87. [Google Scholar]
- Carey, P.J. Drug-induced myelosuppression: Diagnosis and management. Drug Saf. 2003, 26, 691–706. [Google Scholar] [CrossRef] [PubMed]
- Zlotoff, D.A.; Bhandoola, A. Hematopoietic progenitor migration to the adult thymus. Ann. N. Y. Acad. Sci. 2011, 1217, 122–138. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, H.C.; Griffiths, K.L.; Periasamy, P.; Hinton, R.A.; Hey, Y.Y.; Petvises, S.; Tan, J.K. Spleen as a site for hematopoiesis of a distinct antigen presenting cell type. Stem. Cells Int. 2011, 2011, 954275. [Google Scholar] [CrossRef]
- Kapila, V.; Wehrle, C.J.; Tuma, F. Physiology, Spleen. In StatPearls; StatPearls: Tampa/St. Petersburg, FL, USA, 2022. [Google Scholar]
- Zhao, J.; He, S.; Xiang, C.; Zhang, S.; Chen, X.; Lu, X.; Yao, Q.; Yang, L.; Ma, L.; Tian, W. KLF9 promotes autophagy and apoptosis in T-cell acute lymphoblastic leukemia cells by inhibiting AKT/mTOR signaling pathway. Mol. Cell. Toxicol. 2022. [Google Scholar] [CrossRef]
- Rehberger, K.; Kropf, C.; Segner, H. In vitro or not in vitro: A short journey through a long history. Environ. Sci. Eur. 2018, 30, 23. [Google Scholar] [CrossRef]
- Hulsart-Billstrom, G.; Dawson, J.I.; Hofmann, S.; Muller, R.; Stoddart, M.J.; Alini, M.; Redl, H.; El Haj, A.; Brown, R.; Salih, V.; et al. A surprisingly poor correlation between in vitro and in vivo testing of biomaterials for bone regeneration: Results of a multicentre analysis. Eur. Cell Mater. 2016, 31, 312–322. [Google Scholar] [CrossRef]
- Xu, Y.; Shrestha, N.; Preat, V.; Beloqui, A. An overview of in vitro, ex vivo and in vivo models for studying the transport of drugs across intestinal barriers. Adv. Drug Deliv. Rev. 2021, 175, 113795. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, K.M.; Kang, J.S. Preventive effect of Angelica gigas Nakai extract oral administration on dry eye syndrome. Asian Pac. J. Trop. Med. 2018, 11, 369. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | |
|---|---|---|
| murine GAPDH | Forward | : 5′-AGGTCGGTGTGAACGGATTTG-3′ |
| Reverse | : 5′-TGTAGACCATGTAGTTGAGGTCA-3′ | |
| murine IL-1β | Forward | : 5′-GCTACCTGTGTCTTTCCCGT-3′ |
| Reverse | : 5′-CGTCACACACCAGCAGGTTA-3′ | |
| murine IL-3 | Forward | : 5′-GGTTCTTGCCAGCTCTACCA-3′ |
| Reverse | : 5′-GGTATCCCGGCCACTGATTG-3′ | |
| murine IL-6 | Forward | : 5′-GTGGCTAAGGACCAAGACCA-3′ |
| Reverse | : 5′-TAACGCACTAGGTTTGCCGA-3′ | |
| murine SCF | Forward | : 5′-ATGTTCCCCGCTCTCTTTGG-3′ |
| Reverse | : 5′-GTGTGGCATAAGGGCTCACT-3′ | |
| murine GM-CSF | Forward | : 5′-GGCCATCAAAGAAGCCCTGA-3′ |
| Reverse | : 5′-TGGTGAAATTGCCCCGTAGA-3′ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, M.; Park, S.; Chung, Y.; Lim, J.-O.; Kang, J.S.; Park, J.H. Hematopoietic Effects of Angelica gigas Nakai Extract on Cyclophosphamide-Induced Myelosuppression. Plants 2022, 11, 3476. https://doi.org/10.3390/plants11243476
Kang M, Park S, Chung Y, Lim J-O, Kang JS, Park JH. Hematopoietic Effects of Angelica gigas Nakai Extract on Cyclophosphamide-Induced Myelosuppression. Plants. 2022; 11(24):3476. https://doi.org/10.3390/plants11243476
Chicago/Turabian StyleKang, Mincheol, Seojin Park, Yuseong Chung, Je-Oh Lim, Jae Seon Kang, and Jun Hong Park. 2022. "Hematopoietic Effects of Angelica gigas Nakai Extract on Cyclophosphamide-Induced Myelosuppression" Plants 11, no. 24: 3476. https://doi.org/10.3390/plants11243476