
A Morphometric and Karyological Study of the Anthemis macedonica Group (Asteraceae, Anthemideae) Reveals a New Species from Greece
 , , and
, , and
Abstract
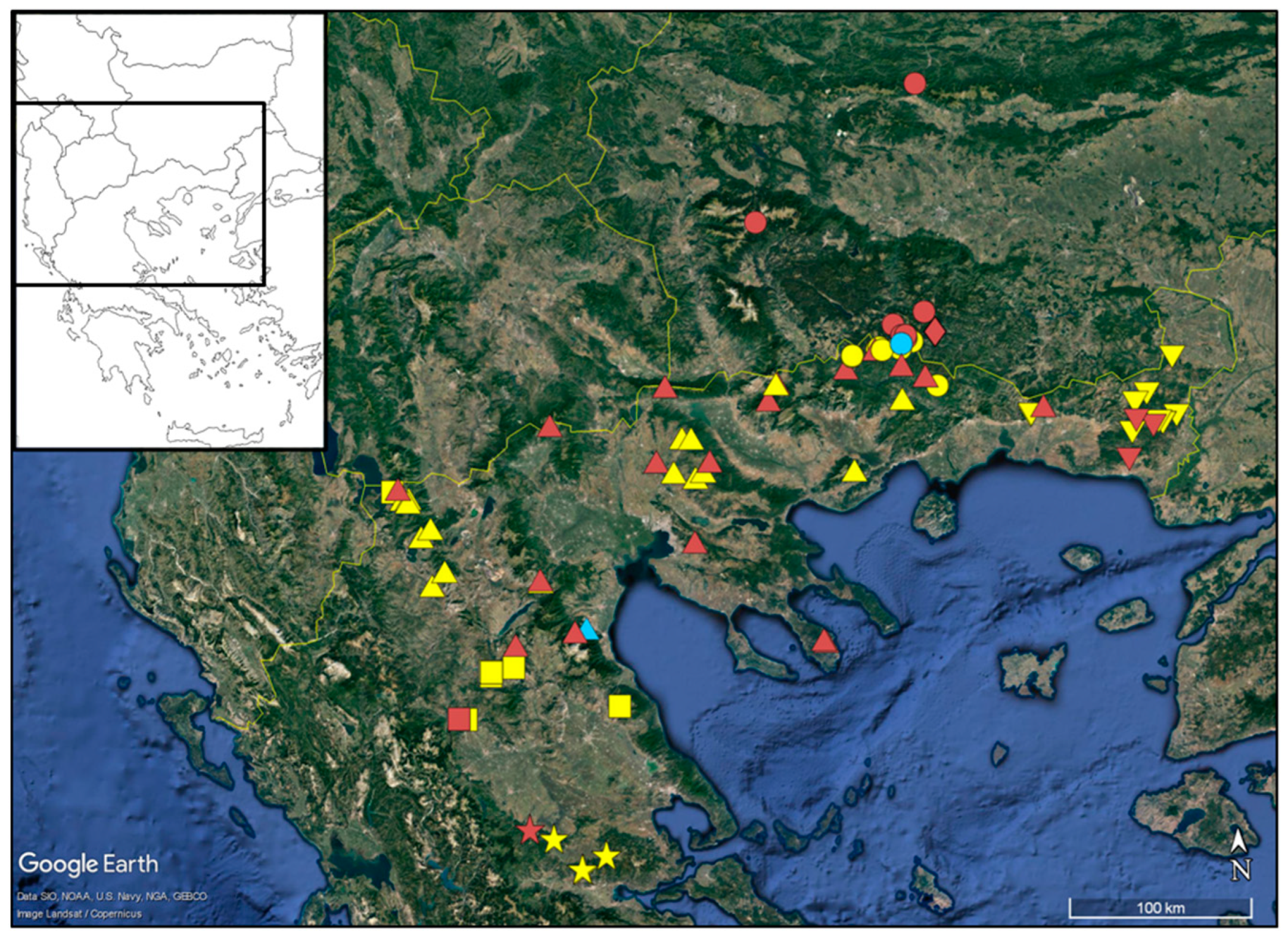
:1. Introduction
2. Results
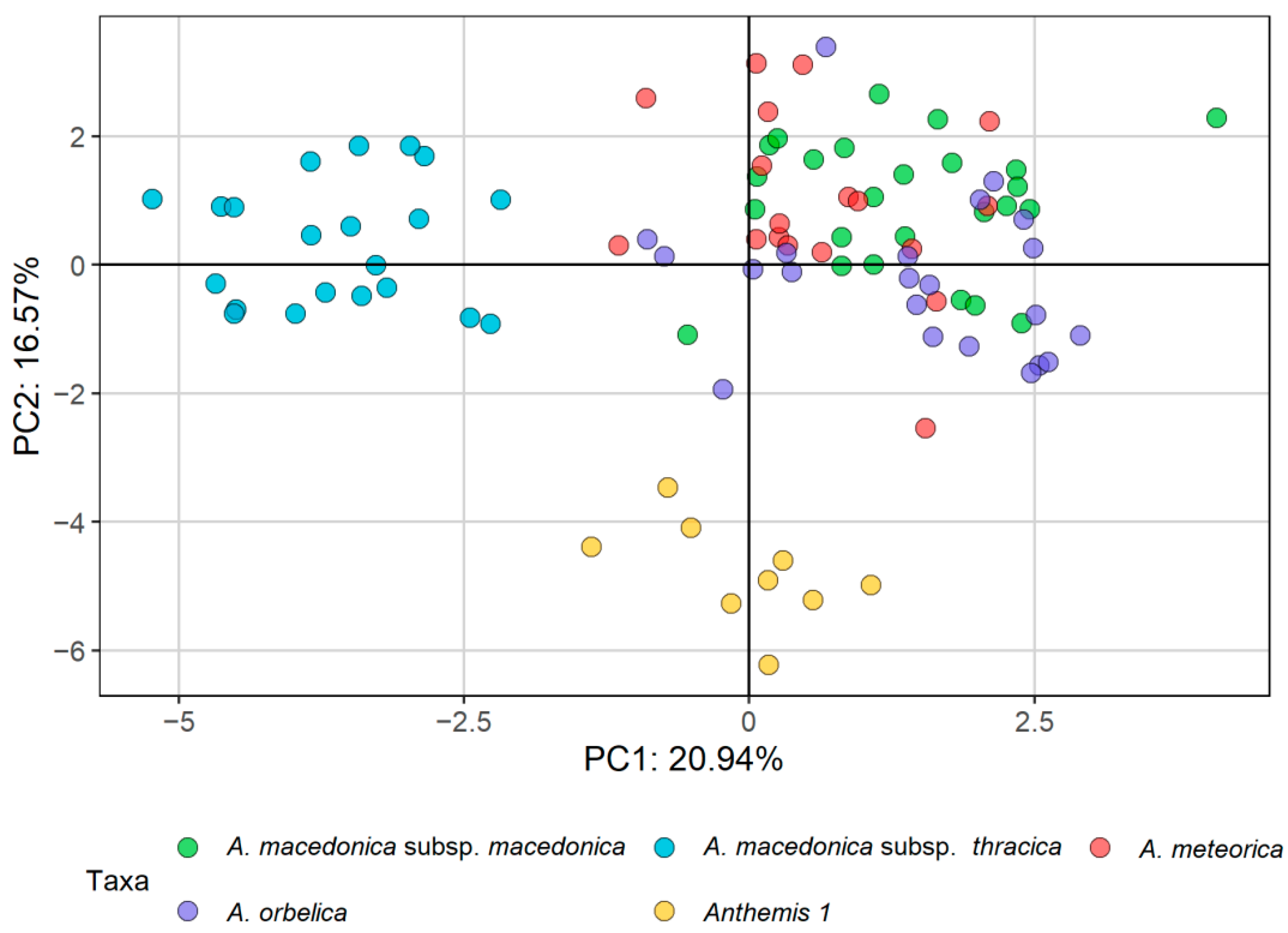
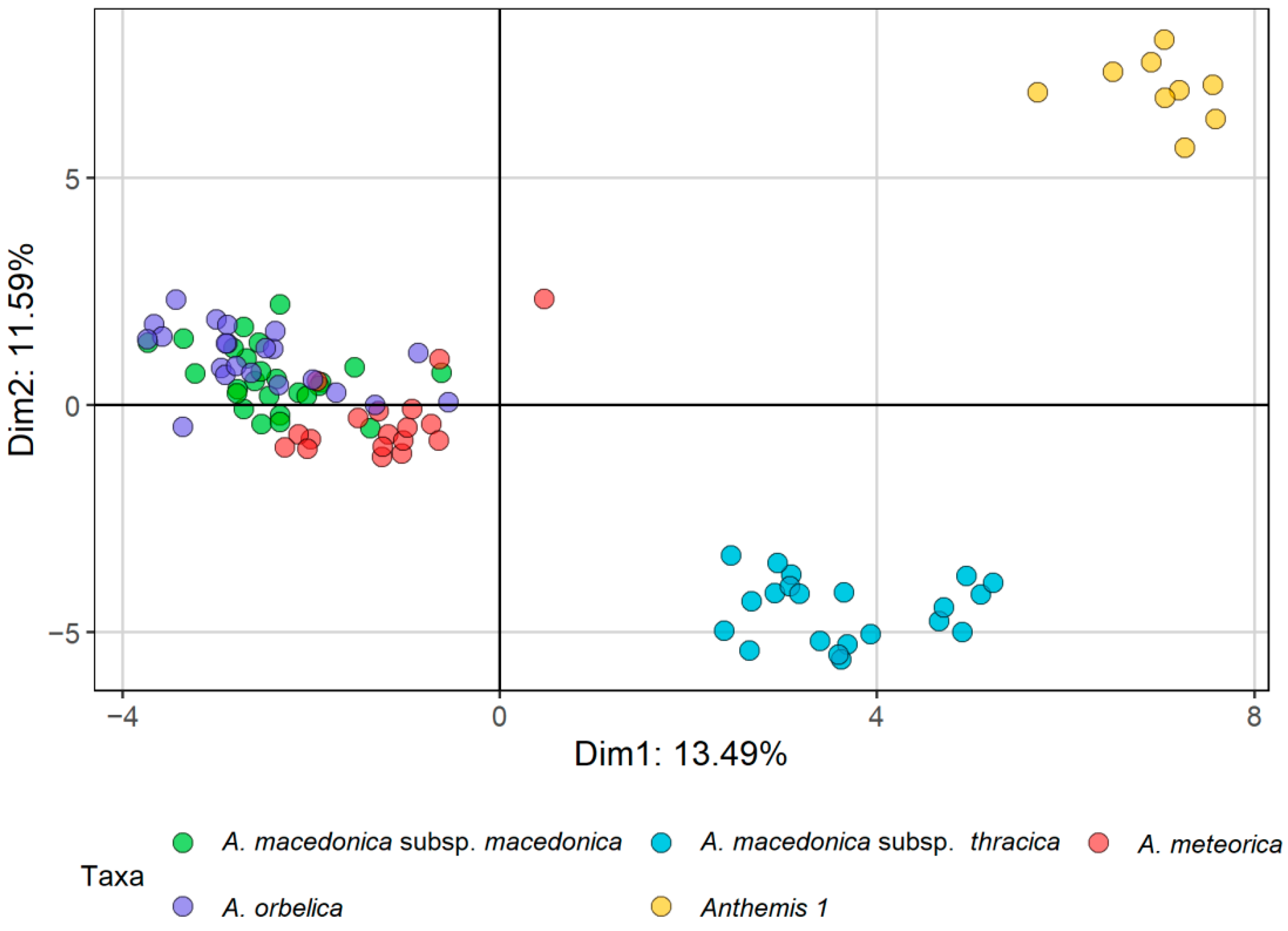
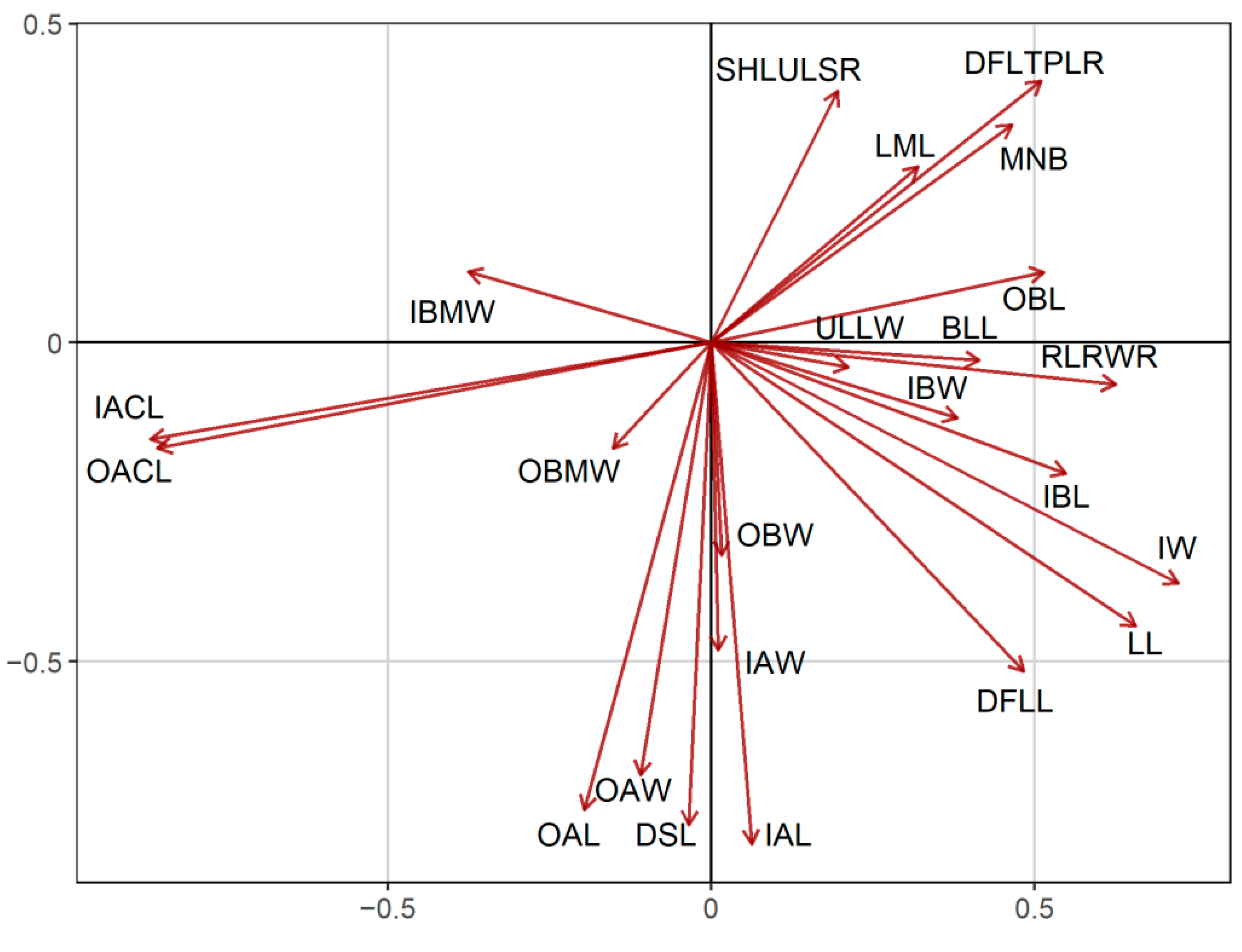
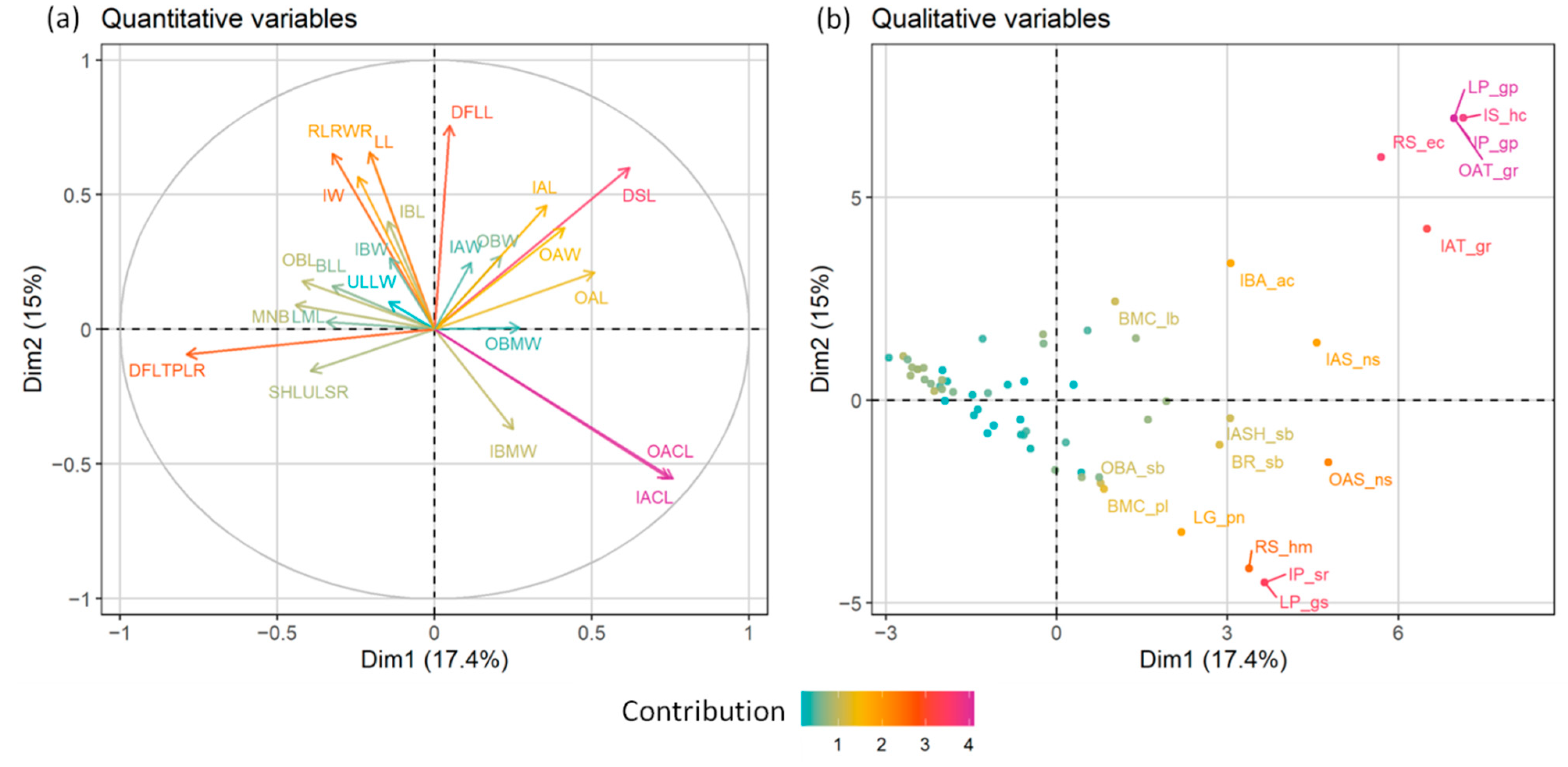
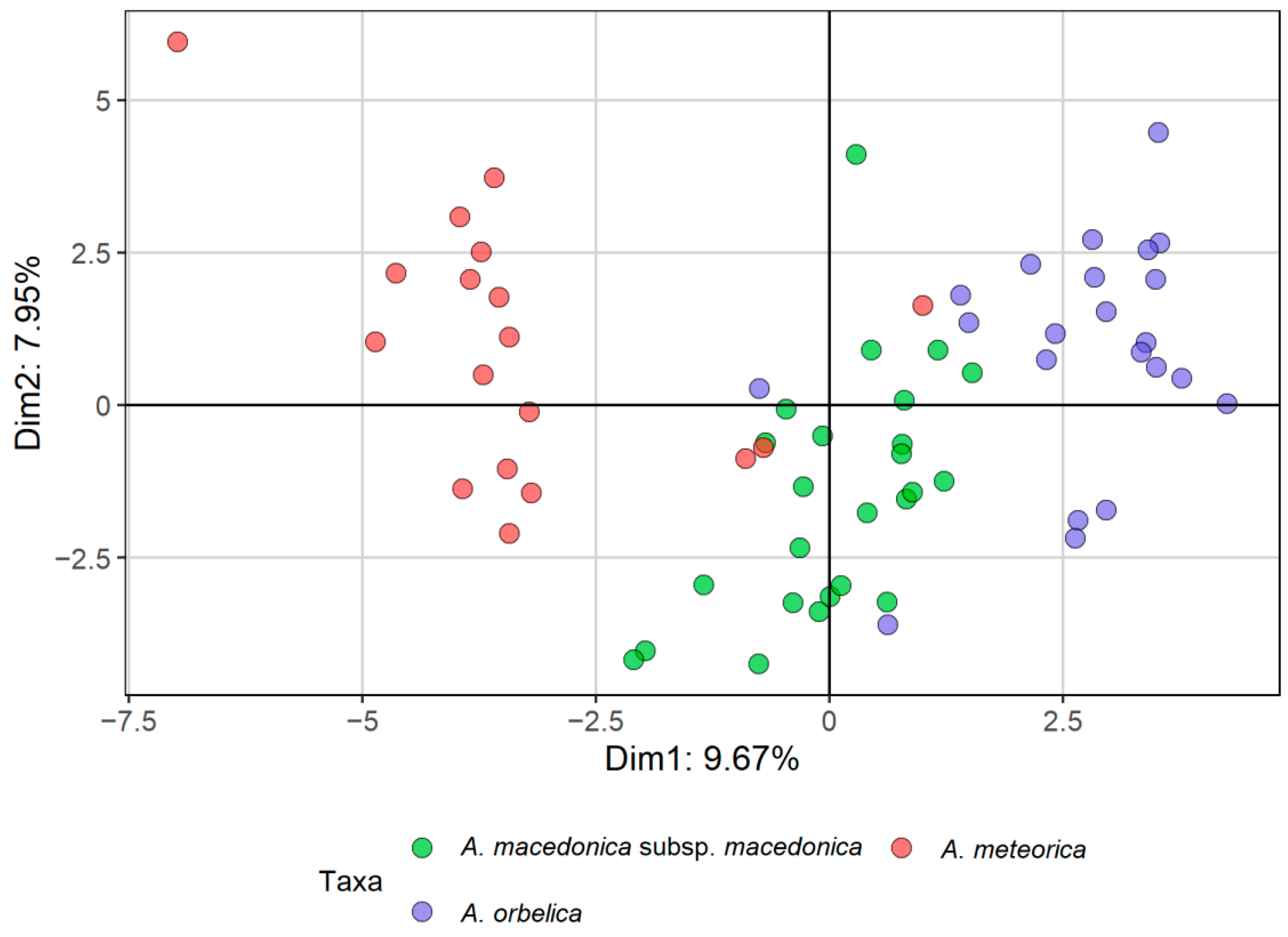
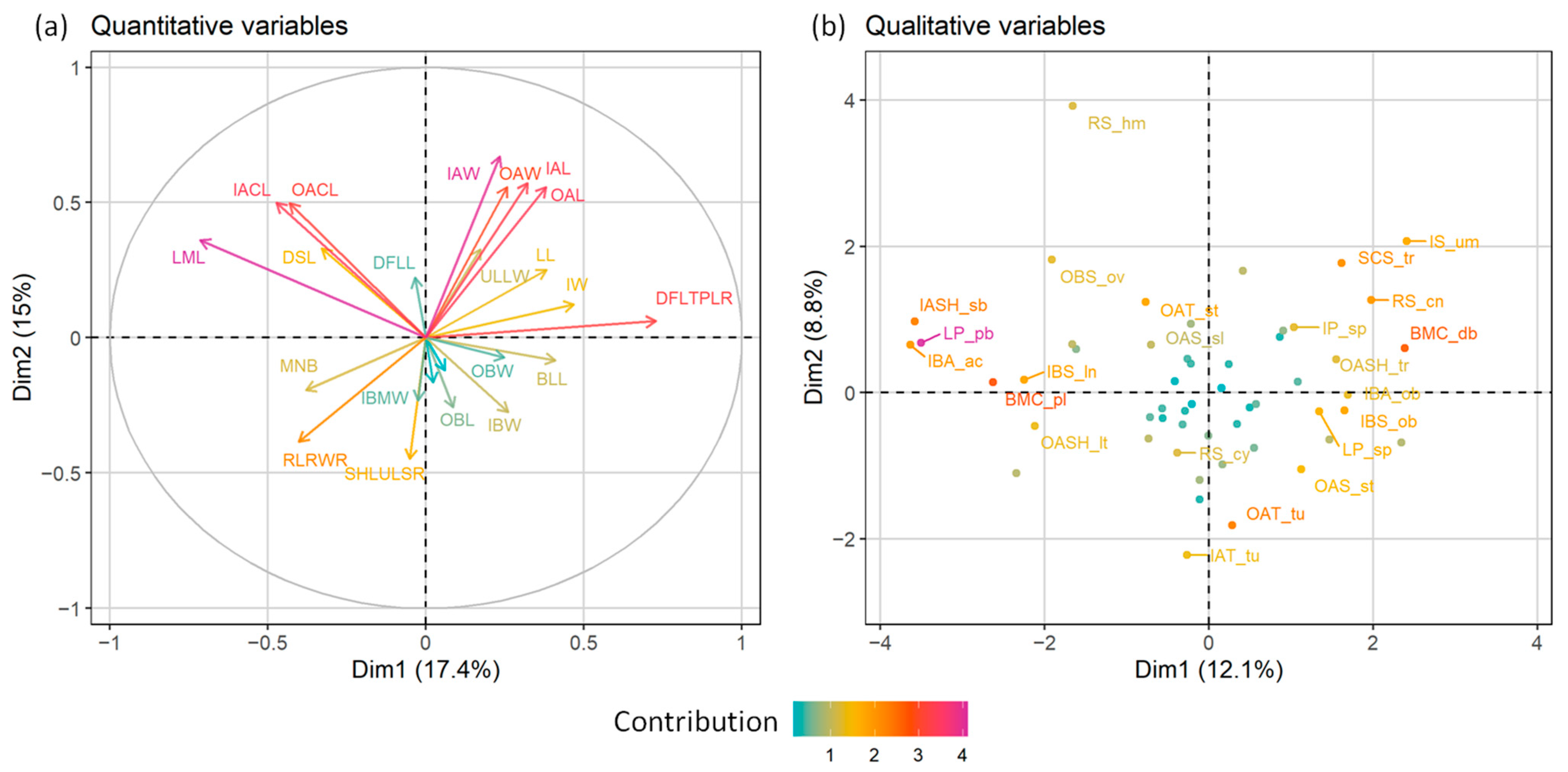
2.1. Morphometric Analyses
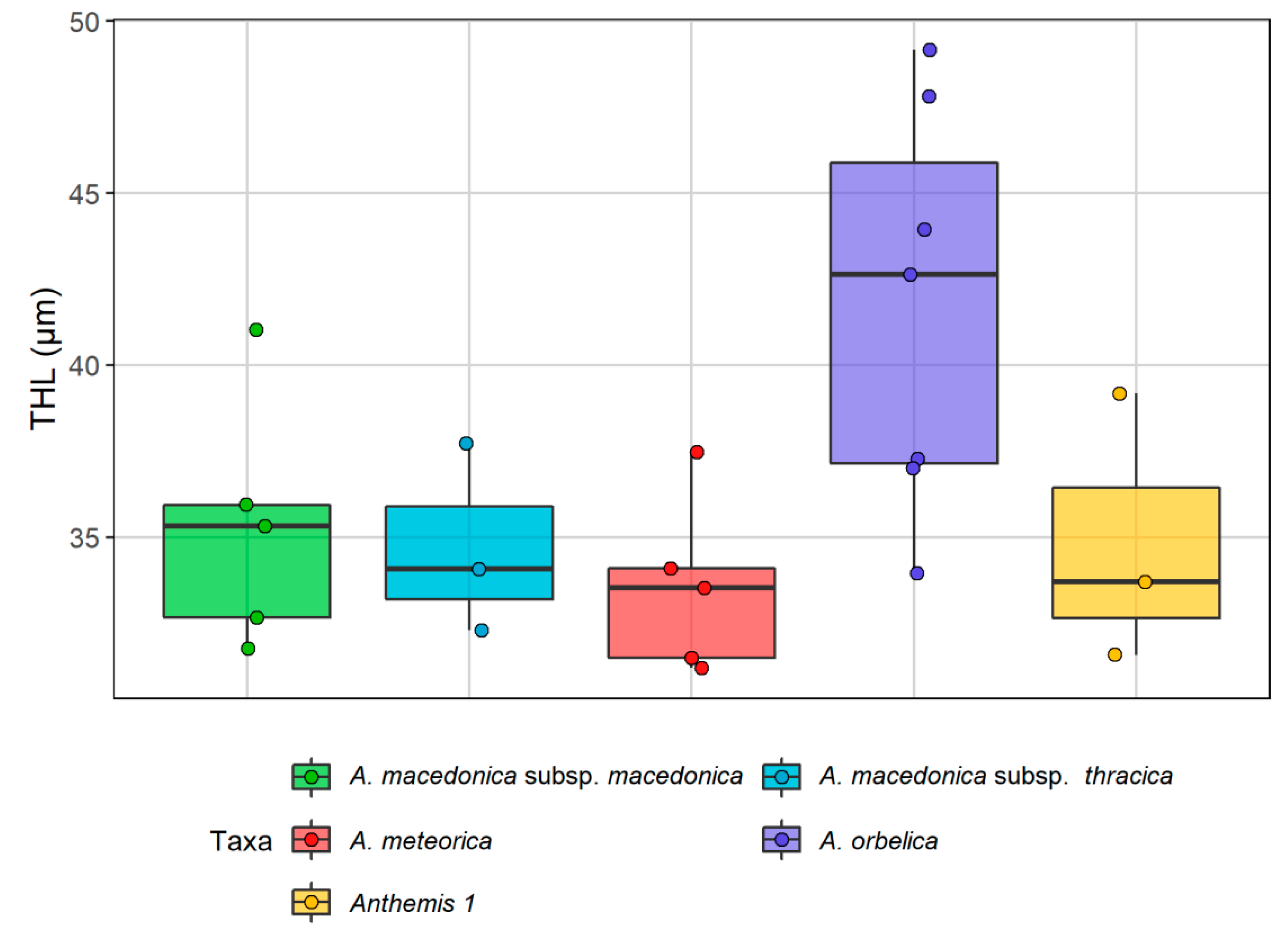
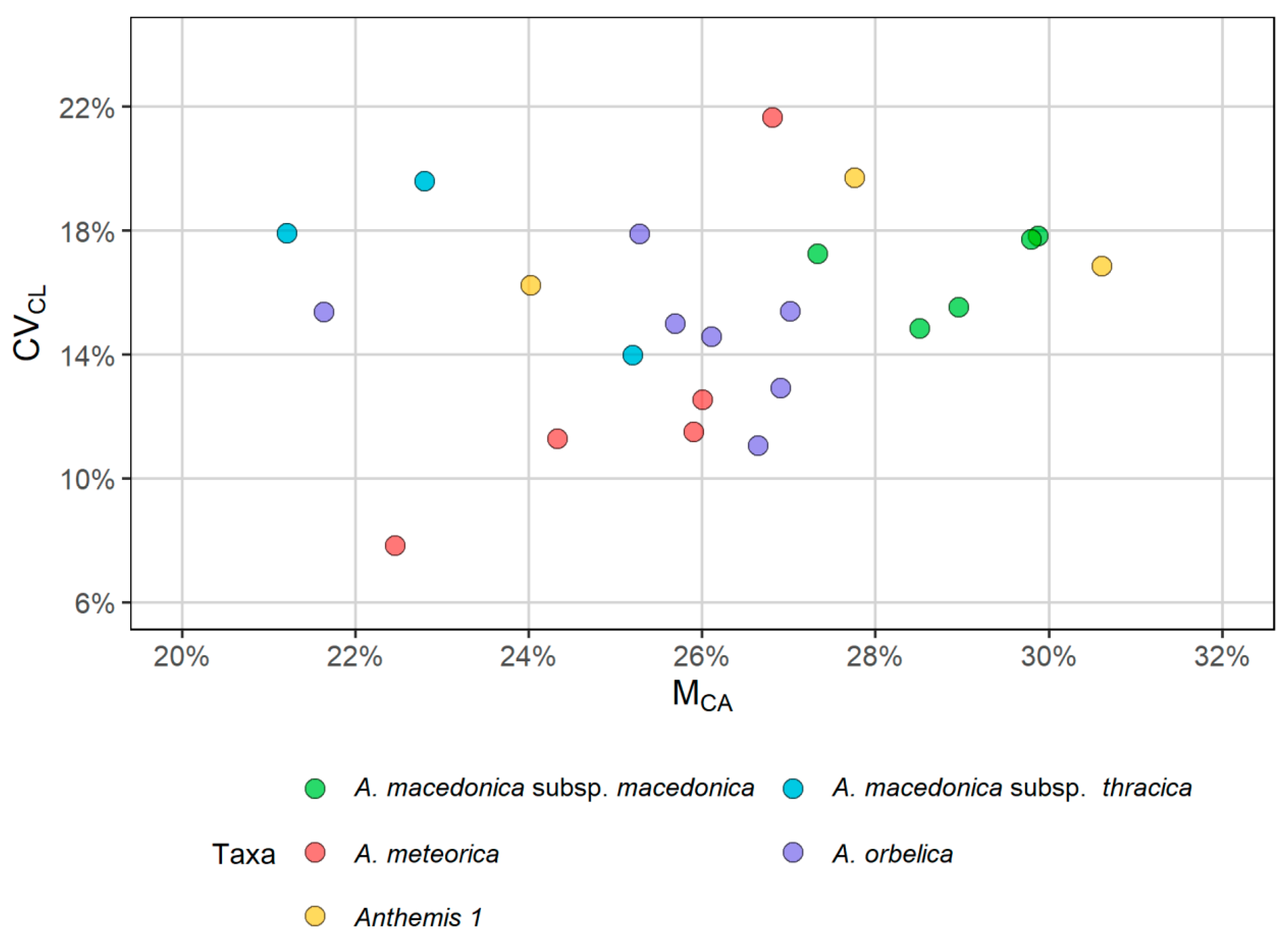
2.2. Karyological Analyses
3. Discussion
3.1. Evaluation of Taxa within the Anthemis macedonica Group
3.2. Key to Taxa
3.3. Taxonomic Treatment
4. Materials and Methods
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Examined Specimens
References
- Oberprieler, C.; Himmelreich, S.; Källersjö, M.; Vallès, J.; Watson, L.E.; Vogt, R. Anthemideae. In Systematics, Evolution, and Biogeography of Compositae; Funk, V.A., Susanna, A., Stuessy, T.F., Bayer, R.J., Eds.; IAPT: Vienna, Austria, 2009; pp. 631–666. [Google Scholar]
- Lo Presti, R.M.; Oppolzer, S.; Oberprieler, C. A molecular phylogeny and a revised classification of the Mediterranean genus Anthemis s.l. (Compositae, Anthemideae) based on three molecular markers and micromorphological characters. Taxon 2010, 59, 1441–1456. [Google Scholar] [CrossRef]
- Oberprieler, C. Phylogenetic relationships in Anthemis L. (Compositae, Anthemideae) based on nrDNA ITS sequence variation. Taxon 2001, 50, 745–762. [Google Scholar] [CrossRef]
- Oberprieler, C. Temporal and spatial diversification of Circum-Mediterranean Compositae-Anthemideae. Taxon 2005, 54, 951–966. [Google Scholar] [CrossRef]
- Oberprieler, C. The Systematics of Anthemis L. (Compositae, Anthemideae) in W and C North Africa. Bocconea 1998, 9, 1–328. [Google Scholar]
- Dimopoulos, P.; Raus, T.; Bergmeier, E.; Constantinidis, T.; Iatrou, G.; Kokkini, S.; Strid, A.; Tzanoudakis, D. Vascular Plants of 545 Greece: An Annotated Checklist. Englera 2013, 31, 1–372. Available online: http://www.jstor.org/stable/24365847 (accessed on 30 June 2022).
- Euro+Med Plantbase—The Information Resource for Euro-Mediterranean Plant Diversity. Available online: http://www.europlusmed.org (accessed on 31 May 2022).
- Grisebach, A. Spicilegium Florae Rumelicae et Bithynicae, Volumen Secundum; F. Vieweg et filium: Brunsvigae, Germany, 1844; pp. 208–209. [Google Scholar]
- Boissier, E. Diagnoses Plantarum Orientalium Novarum; Series secunda, No 6; B. Hermann: Lipsiae, Germany; B. Baillière: Parisiis, France, 1859; pp. 97–98. [Google Scholar]
- Boissier, E. Flora Orientalis, Volumen Tertium; H. Georg.: Lugduni, France, 1875; p. 308. [Google Scholar]
- Pančić, J. Elementa ad Floram Principatus Bulgariae; Kraljevsko-Srpska Drž. Štamparija: Beograd, Serbia, 1883; p. 39. [Google Scholar]
- Pančić, J. Nova Elementa ad Floram Principatus Bulgariae; Kraljevsko-Srpska Drž. Štamparija: Beograd, Serbia, 1886; p. 27. [Google Scholar]
- Formánek, E. Dritter beitrag zur Flora von Serbien und Bulgarien. Verh. Naturf. Vereins Brünn 1898, 36, 6–116. [Google Scholar]
- Velenovský, J. Plantae novae bulgaricae. Österr. Bot. Z. 1902, 52, 154–156. [Google Scholar] [CrossRef] [Green Version]
- Velenovský, J. Nachträge zur Flora von Bulgarien. Sitzungsber. Königl. Böhm. Ges. Wiss., Math.-Naturwiss. Cl. 1903, 28, 1–31. [Google Scholar]
- Vandas, C. Reliquiae Formánekianae; J. Jelínek: Brunae, Czech Republic, 1909; pp. 285–286. [Google Scholar]
- Hayek, A. Prodromus Florae Peninsulae Balcanicae. Repert. Spec. Nov. Regni Veg. Beih. 1931, 30, 1–1152. [Google Scholar]
- Stojanov, N.; Achtarov, B. Material zur Kenntnis der Gattung Anthemis. Notizbl. Königl. Bot. Gart. Berlin 1937, 13, 513–523. [Google Scholar]
- Stojanov, N.; Stefanov, B. Flora na Balgarija, 3rd ed.; Universitetska pecatnica: Sofia, Bulgaria, 1948. [Google Scholar]
- Fernandes, R. Anthemis L. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, England, 1976; Volume 4, pp. 145–159. [Google Scholar]
- Greuter, W.; Oberprieler, C.; Vogt, R. The Euro+Med treatment of Anthemideae (Compositae)—Generic concepts and required new names. Willdenowia 2003, 33, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Haussknecht, C. Symbolae ad floram graecam. Aufzählung der im Sommer 1885 in Griechenland gesammelten Pflanzen. Mitth. Thüring. Bot. Vereins 1893, 5, 41–126. [Google Scholar]
- Franzén, R. Anthemis cretica (Asteraceae) and related species in Greece. Willdenowia 1986, 16, 35–45. [Google Scholar]
- Bornovas, J.; Rondogianni-Tsiambaou, T. Geological map of Greece, 2nd ed.; Institute of Geology and Mineral Exploration: Athens, Greece, 1983. [Google Scholar]
- Constantinidis, T. The floristic diversity of serpentine in Greece 1. An inventory of the Aliki Area (Sterea Ellas, Central Greece). Phyton 2004, 44, 45–67. [Google Scholar]
- Kuzmanov, B.; Thin, N.N.; Georgieva, S. A cytotaxonomic study on Bulgarian Anthemis species. Candollea 1981, 36, 19–76. [Google Scholar]
- Peruzzi, L.; Eroğlu, H.E. Karyotype asymmetry: Again, how to measure and what to measure? Comp. Cytogenet. 2013, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paszco, B. A critical review and a new proposal of karyotype asymmetry indices. Plant Syst. Evol. 2006, 258, 39–48. [Google Scholar] [CrossRef]
- Zarco, C.R. A new method for estimating karyotype asymmetry. Taxon 1986, 35, 526–530. [Google Scholar] [CrossRef]
- Tomović, G.; Niketić, M.; Lakušić, D.; Ranđelović, V.; Stevanović, V. Balkan endemic plants in Central Serbia and Kosovo regions: Distribution patterns, ecological characteristics, and centres of diversity. Bot. J. Linn. 2014, 176, 173–202. [Google Scholar] [CrossRef] [Green Version]
- Assyov, B.; Petrova, A. Conspectus of the Bulgarian Vascular Flora, 4th ed.; Bulgarian Biodiversity Foundation: Sofia, Bulgaria, 2012; pp. 71–72. [Google Scholar]
- Peev, D.; Petrova, A.; Anchev, M.; Temniskova, D.; Denchev, C.M.; Ganeva, A.; Gussev, C.; Vladimirov, V. Red Data Book of the Republic of Bulgaria 1. Plants and Fungi; Academy of Sciences & Ministry of Environment and Water: Sofia, Bulgaria, 2015.
- Pichi-Sermolli, R. Flora e vegetazione delle serpentine e delle alter ofioliti dell’ alta valle del Tevere (Toscana). Webbia 1948, 6, 3–378. [Google Scholar] [CrossRef]
- Brady, K.U.; Kruckeberg, A.R.; Bradshaw, H.D., Jr. Evolutionary ecology of plant adaptation to serpentine soils. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 243–266. [Google Scholar] [CrossRef]
- Stevanović, V.; Tan, K.; Iatrou, G. Distribution of the endemic Balkan flora on serpentine I.—Obligate serpentine endemics. Plant Syst. Evol. 2003, 242, 149–170. [Google Scholar] [CrossRef]
- Proctor, J.; Woodell, S.R.J. The ecology of serpentine soils. Adv. Ecol. Res. 1975, 9, 255–365. [Google Scholar] [CrossRef]
- Kleinsteuber, A.; Ristow, M.; Hassler, M. Flora von Rhodos und Chalki, Band 1; Kleinsteuber Books: Karlsruhe, Germany, 2016; pp. 146–147. [Google Scholar]
- Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 28 February 2022).
- Stearn, W.T. Botanical Latin, 3rd ed.; David & Charles: Newton Abbot, England, 1978; p. 566. [Google Scholar]
- Jolliffe, I.T. Principal Component Analysis; Springer: New York, NY, USA, 2002. [Google Scholar]
- The R Project for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 15 May 2022).
- Pagès, J. Analyse factorielle de données mixtes. Rev. Stat. Appl. 2004, 52, 93–111. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Goula, K.; Constantinidis, T. Taxonomic diversity and karyology of Anthemis rigida (Anthemideae, Asteraceae) in the Aegean, Greece. Phytotaxa 2021, 484, 129–143. [Google Scholar] [CrossRef]
- Peruzzi, L.; Leitch, I.J.; Caparelli, K.F. Chromosome diversity and evolution in Liliaceae. Ann. Bot. 2009, 103, 459–475. [Google Scholar] [CrossRef]
- Altınordu, F.; Peruzzi, L.; Yu, Y.; He, X. A tool for the analysis of chromosomes: KaryoType. Taxon 2016, 65, 586–592. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Quantitative Characters | |||
| MNB | Maximum number of stem branches at the lower part of stem | OBMW | Outer bracts margin width |
| BLL | Basal leaf length | IBL | Inner bracts length |
| DSLs | Number of divisions in stem leaves | IBW | Inner bracts width |
| ULLW | Ultimate leaf lobes width | IBMW | Inner bracts margin width |
| LML | Leaf mucro length | OACL | Outer disc floret achenes corona length |
| LL | Ligule length | OAL | Outer disc floret achenes length |
| IW | Involucre width | OAW | Outer disc floret achenes width |
| DFLL | Disc floret lobes length | IACL | Inner disc floret achenes corona length |
| OBL | Outer bracts length | IAL | Inner disc floret achenes length |
| OBW | Outer bracts width | IAW | Inner disc floret achenes width |
| Ratios | |||
| SHLULSR | Total stem length/upper leafless stem | DFLTPLR | Total disc floret length/thickened part length |
| RLRWR | Receptecle length/receptacle width | ||
| Qualitative Characters | |||
| LG | Longevity | IP | Involucre pubescence |
| BRs | Stem branches | IS | Involucre shape |
| ULLS | Ultimate leaf lobe shape | RS | Receptacle shape |
| LP | Leaf pubescence | OAT | Outer disc floret achenes surface: tuberculation |
| OBS | Outer bracts shape | OAS | Outer disc floret achenes surface: striation |
| OBA | Outer bracts apex | OASH | Outer disc floret achenes shape |
| IBS | Inner bracts shape | IAT | Inner disc floret achenes surface: tuberculation |
| IBA | Inner bracts apex | IAS | Inner disc floret achenes surface: striation |
| BMC | Bracts margin colour | IASH | Inner disc floret achenes shape |
| SCS | Scale shape | ||
| Taxon | Origin | 2n | Formula | THL | CVCL | MCA |
|---|---|---|---|---|---|---|
| A. macedonica subsp. macedonica | (a) Kastoria, (b) Thessaloniki | 18 | 12m + 2sm + 4stsat | 31.76–41.04 | 14.84–17.82 | 27.33–29.88 |
| A. macedonica subsp. thracica | Evros | 18 | 12m + 4sm + 2stsat | 32.29–37.72 | 13.99–19.6 | 21.21–25.2 |
| A. meteorica | (a) Grevena, (b) Trikala | 18 | 12m + 4sm + 2stsat | 31.19–37.47 | 7.84–21.65 | 22.46–26.81 |
| A. orbelica | (a) Drama-Frakto, (b) Drama-Livaditis | 18 | 12m + 2sm + 4stsat | 37–49.16 | 11.05–17.89 | 21.64–27.02 |
| Anthemis 1 | Magnisia | 18 | 12m + 2sm + 4stsat | 31.58–39.18 | 16.23–19.72 | 24.02–30.61 |
| A. macedonica subsp. macedonica | A. macedonica subsp. orbelica | A. meteorica | A. serpentinica (Anthemis 1) | A. thracica | |
|---|---|---|---|---|---|
| Lifespan | Annual to biennial or short lived perennial | Biennial, sometimes flowering from the first year, or short lived perennial | Biennial, sometimes flowering from the first year, or short lived perennial | Predominately biennial | Predominantly perennial, sometimes flowering from the first year |
| Individual leaf rosettes at flowering period | Usually absent | Usually present | Absent | Present | Absent |
| Leaf pubescence | Sparsely pubescent to glabrous | Sparsely pubescent to subglabrous | Usually pubescent | Tomentose | Tomentose to tomentose-sericeous |
| Involucre pubescence | Glabrescent | Slightly pubescent to glabrescent | Slightly pubescent to almost glabrous | Pubescent to tomentose | Tomentose- sericeous |
| Receptacle shape | Shortly conical to hemispherical-cylindrical | Shortly conical to hemispherical-cylindrical | Elongated hemispherical-cylindrical | Elongated conical, apex sharply acute | Hemispherical to shortly conical |
| Inner involucral bracts shape | Oblanceolate to obovate, apex usually obtuse | Oblanceolate to obovate, apex obtuse to subacute | Lanceolate to oblanceolate, apex subacute | Lanceolate, apex acute | Lanceolate to oblanceolate, apex subacute to acute |
| Involucral bracts margin | Light to dark brown | Usually brown to dark brown | Not coloured | Very thin, light brown or pale | Not coloured |
| Involucral bracts midvein | Green, darker than rest of the bract | Green, much darker than rest of the bract | Concolorous or slightly darker than rest of the bract, prominent | Concolorous with the rest of the bract, prominent | Concolorous with the rest of the bract, prominent |
| Disc florets thickened part length | 1–1.2 | 1–1.1 | 1–1.3 (–1.5) | 1.5–2 | 1–1.2 |
| Disc florets length | (2.4–) 2.5–2.8 (–3) | (2.3–) 2.5–2.8 | (2.3–) 2.7–3 | 2.8–3.2 | 2.2–2.3 (–2.5) |
| Achenes corona length | 0.1 | 0.1 (–0.2) | 0.1–0.2 | 0.2–0.3 | 0.4–0.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goula, K.; Touloumis, K.; Dimopoulos, P.; Constantinidis, T. A Morphometric and Karyological Study of the Anthemis macedonica Group (Asteraceae, Anthemideae) Reveals a New Species from Greece. Plants 2022, 11, 3006. https://doi.org/10.3390/plants11213006
Goula K, Touloumis K, Dimopoulos P, Constantinidis T. A Morphometric and Karyological Study of the Anthemis macedonica Group (Asteraceae, Anthemideae) Reveals a New Species from Greece. Plants. 2022; 11(21):3006. https://doi.org/10.3390/plants11213006
Chicago/Turabian StyleGoula, Katerina, Konstantinos Touloumis, Panayotis Dimopoulos, and Theophanis Constantinidis. 2022. "A Morphometric and Karyological Study of the Anthemis macedonica Group (Asteraceae, Anthemideae) Reveals a New Species from Greece" Plants 11, no. 21: 3006. https://doi.org/10.3390/plants11213006