
Plant Diversity of Mts. Oligirtos and Farmakas (NE Peloponnisos, Greece) with Emphasis on Their Endemic Flora


Abstract
:1. Introduction
2. Results
2.1. Floristic Analysis of Mts. Oligirtos and Farmakas
2.1.1. Total Flora
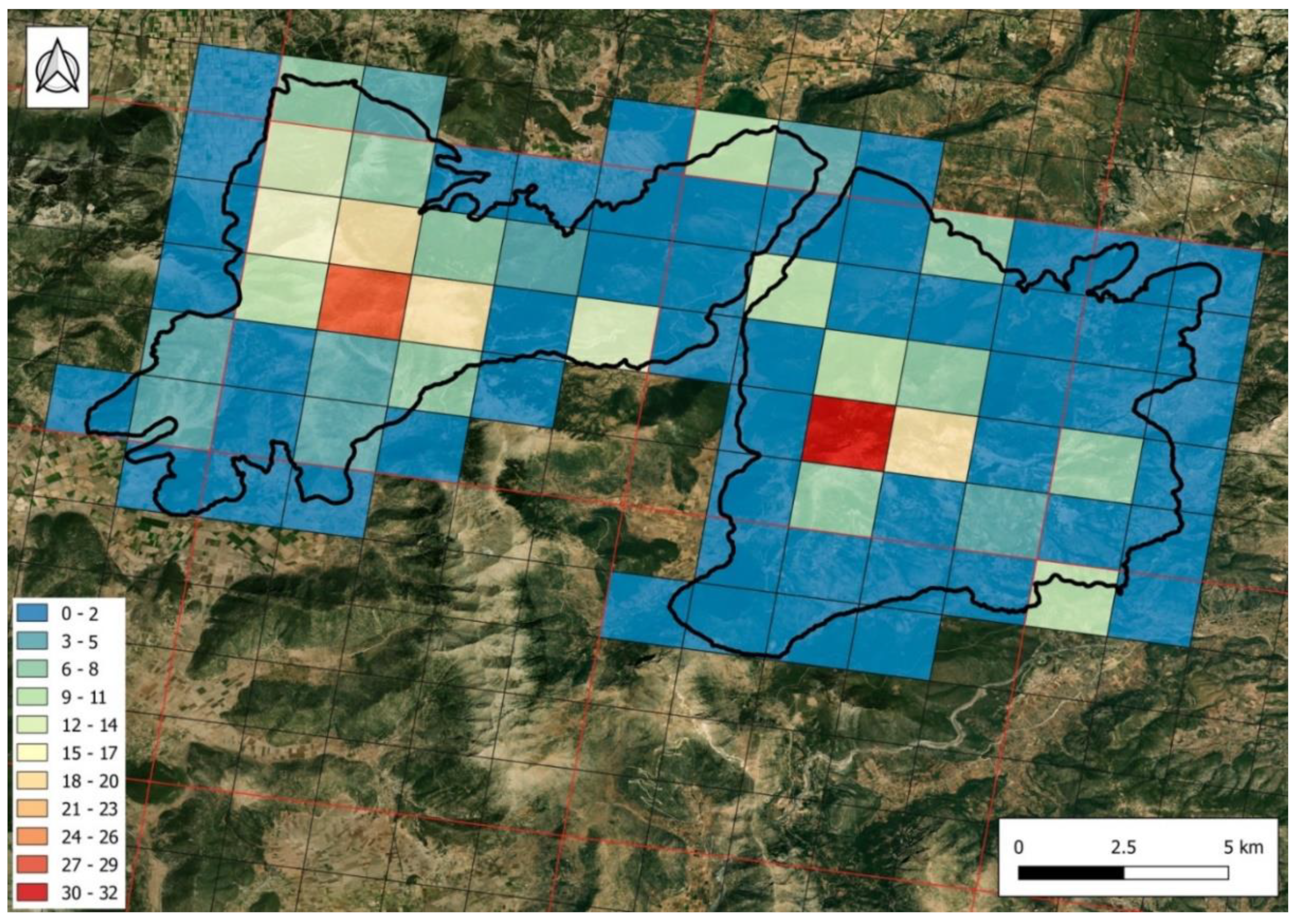
2.1.2. Endemic Flora
2.1.3. Threatened Endemic Taxa and Prioritization
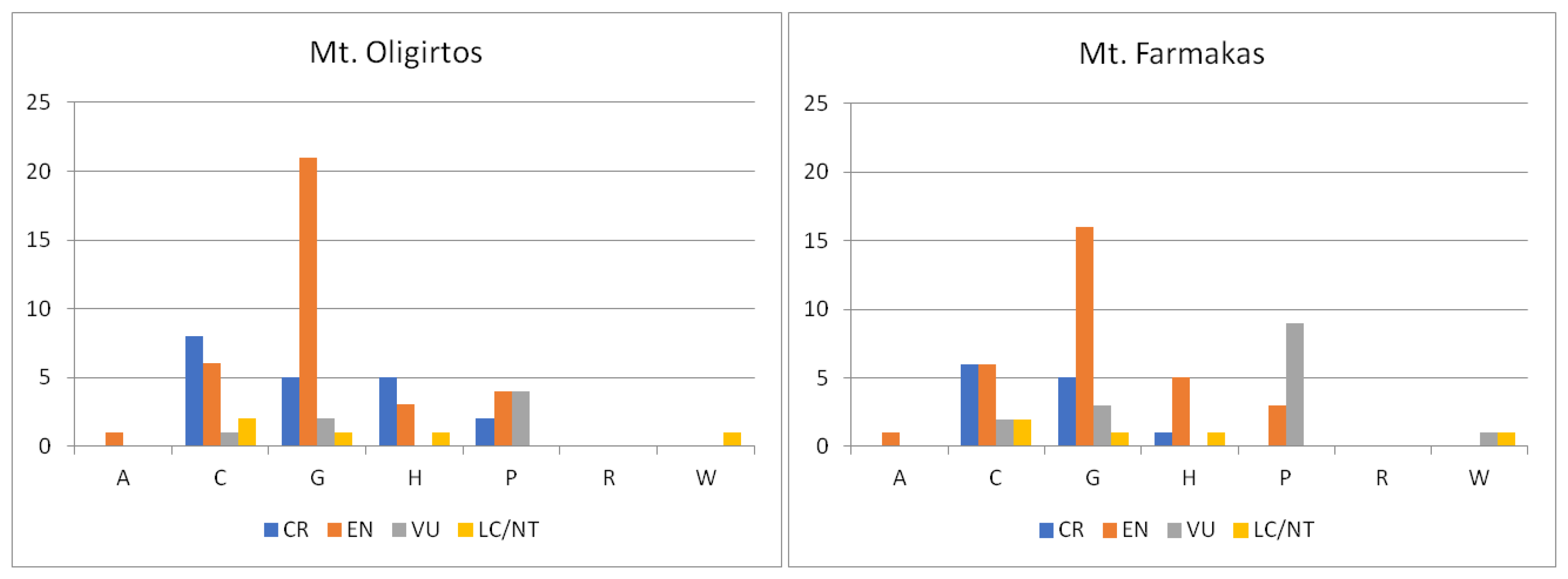
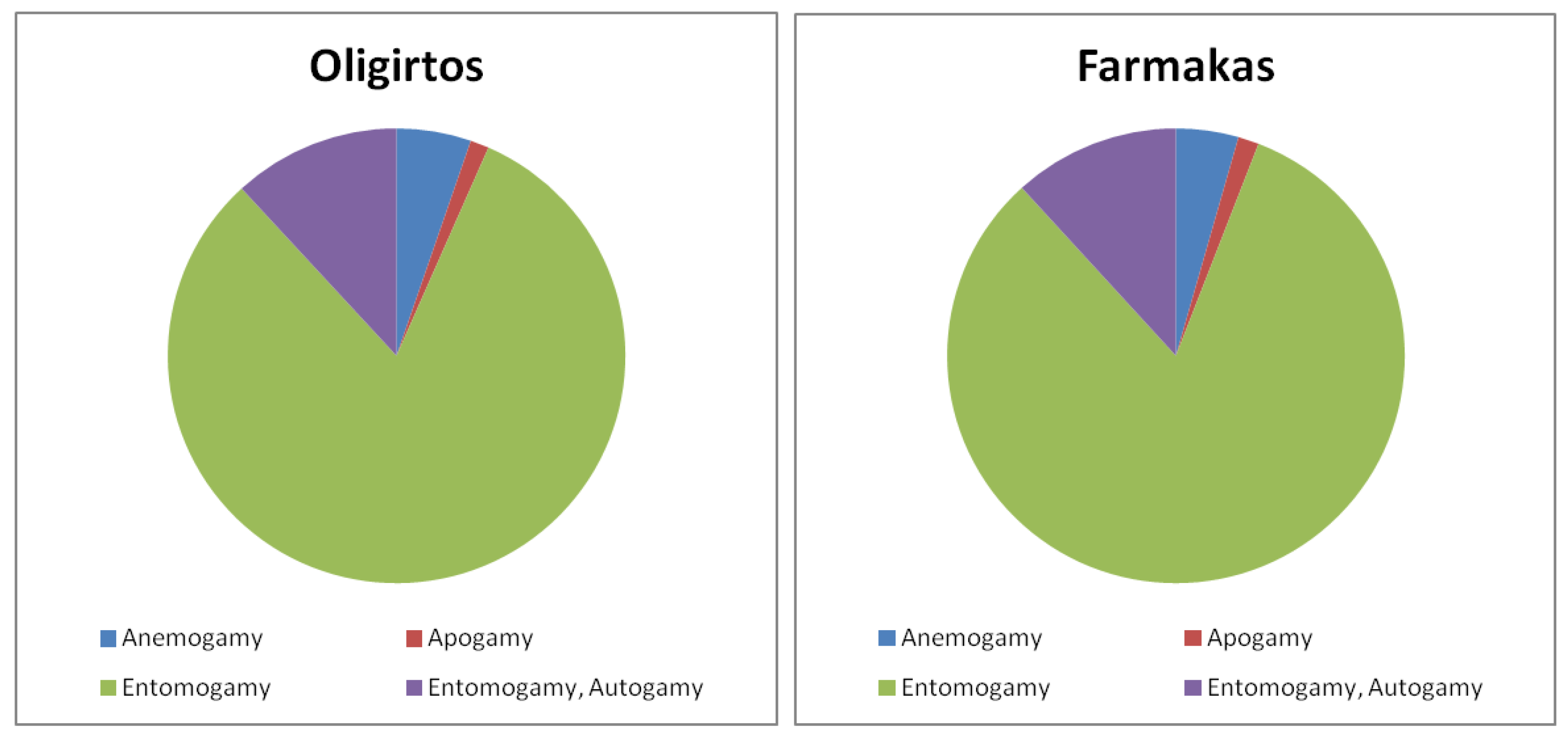
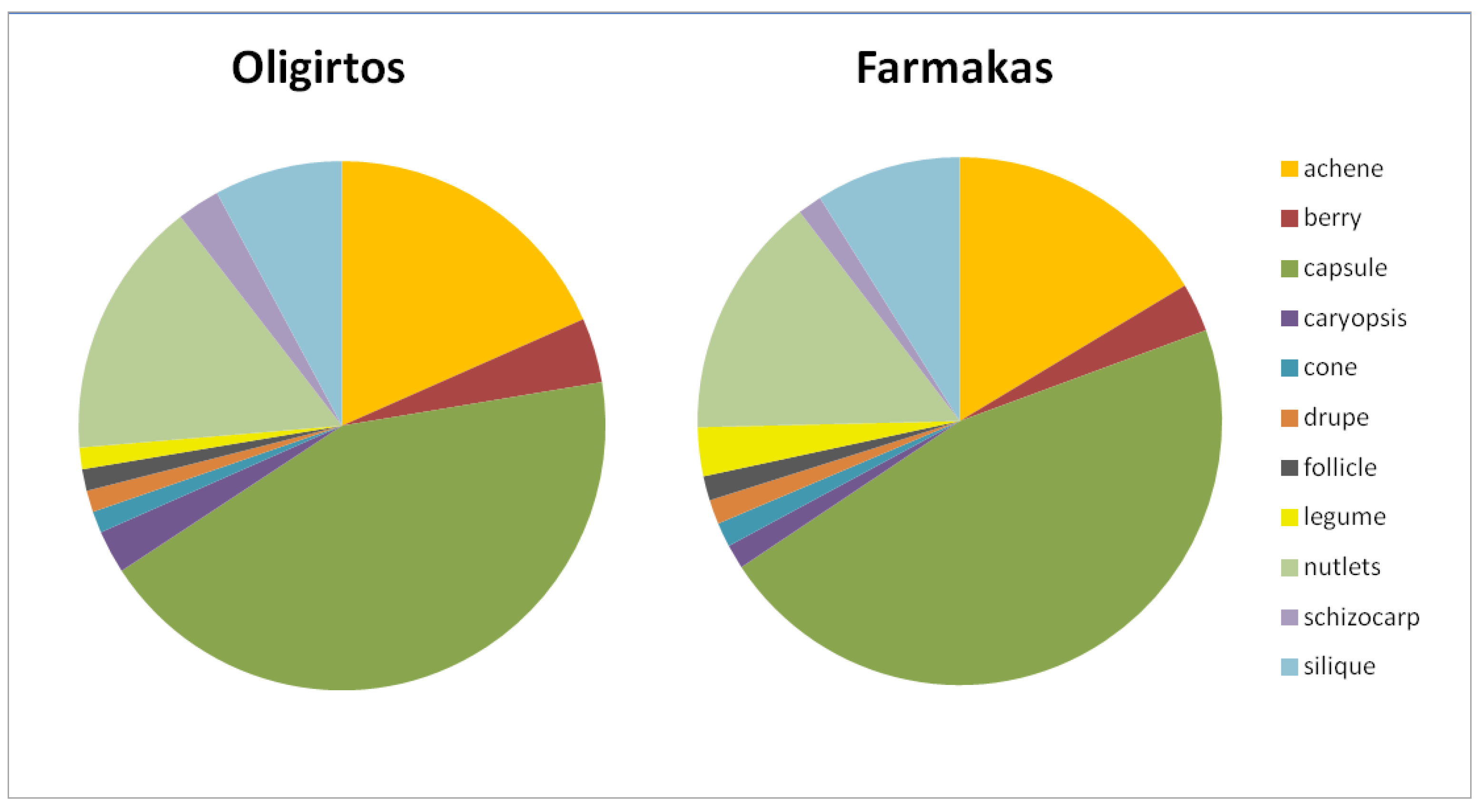
2.1.4. Biological Traits of Endemics
2.2. Comparative Analysis of Endemic Flora among Mountains of N Peloponnisos
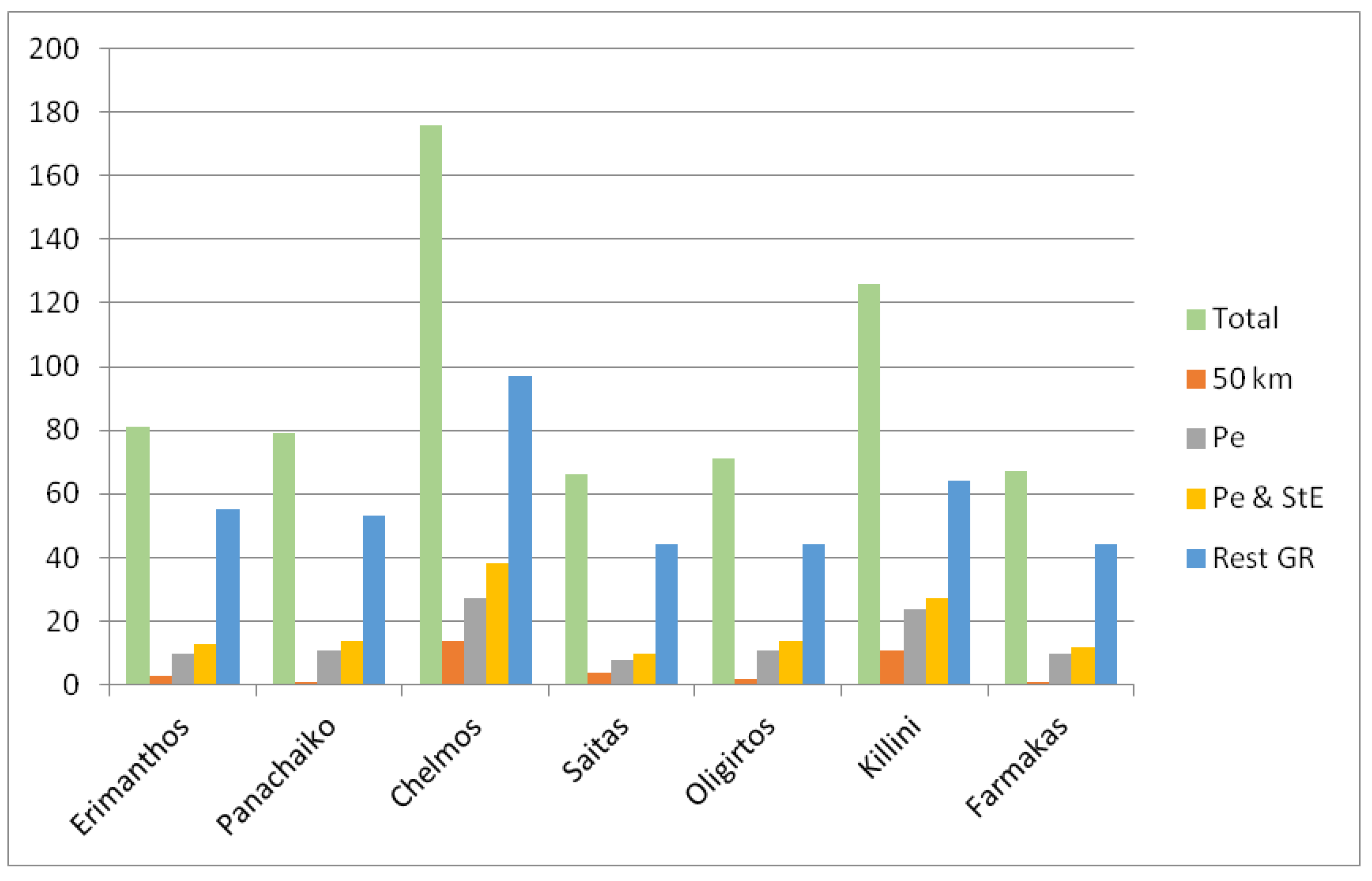
2.2.1. Diversity of Endemic Taxa
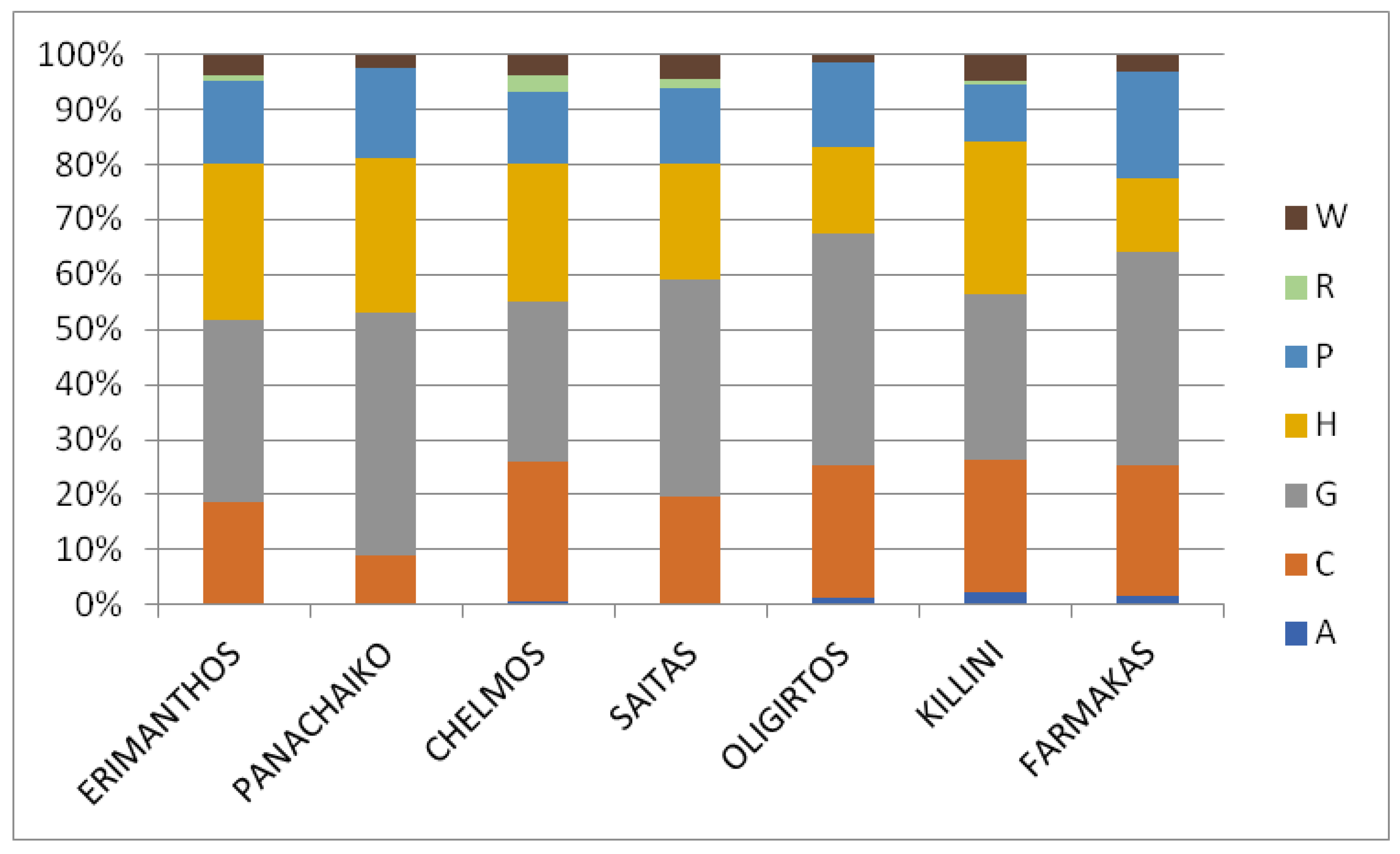
2.2.2. Life Forms
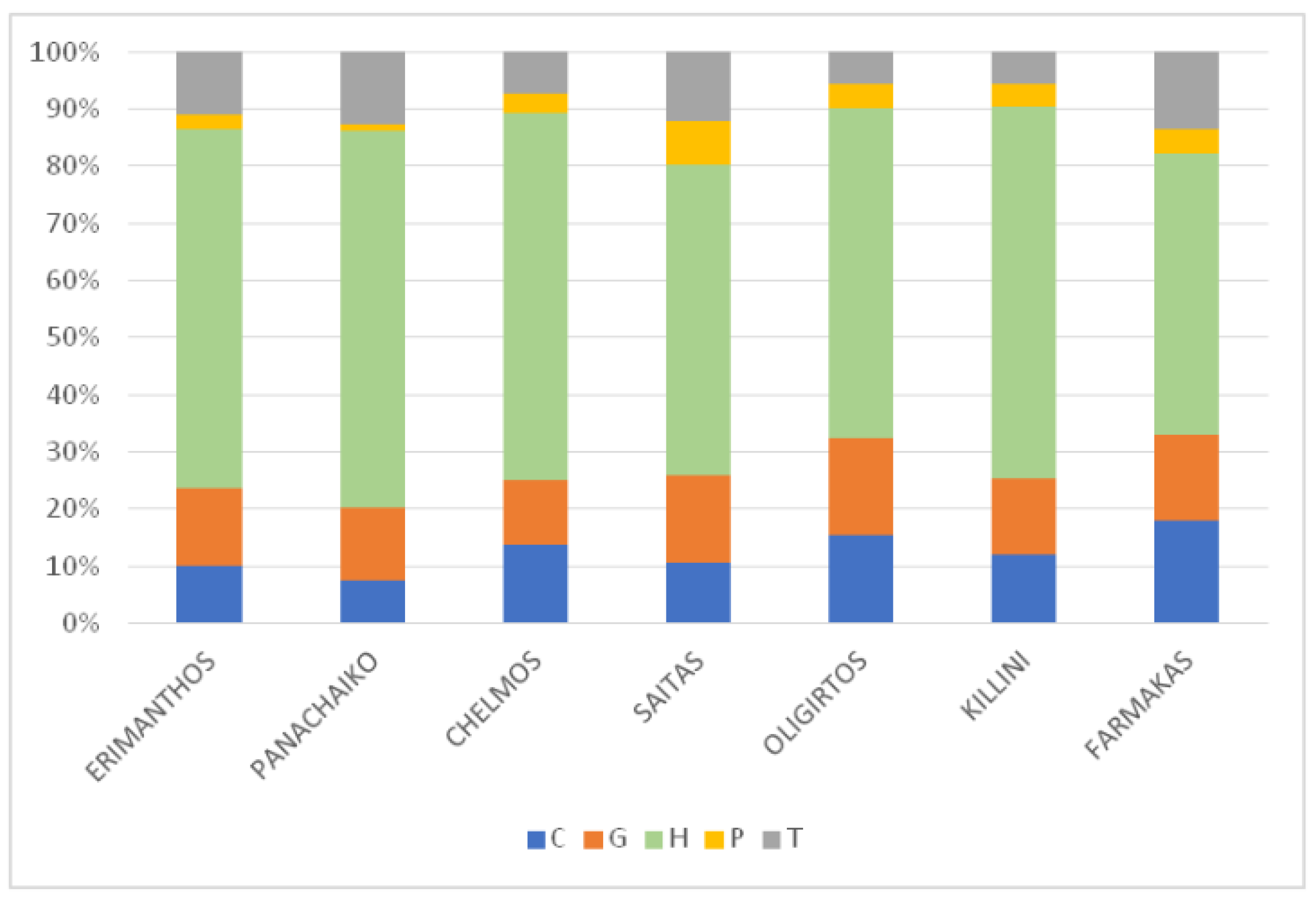
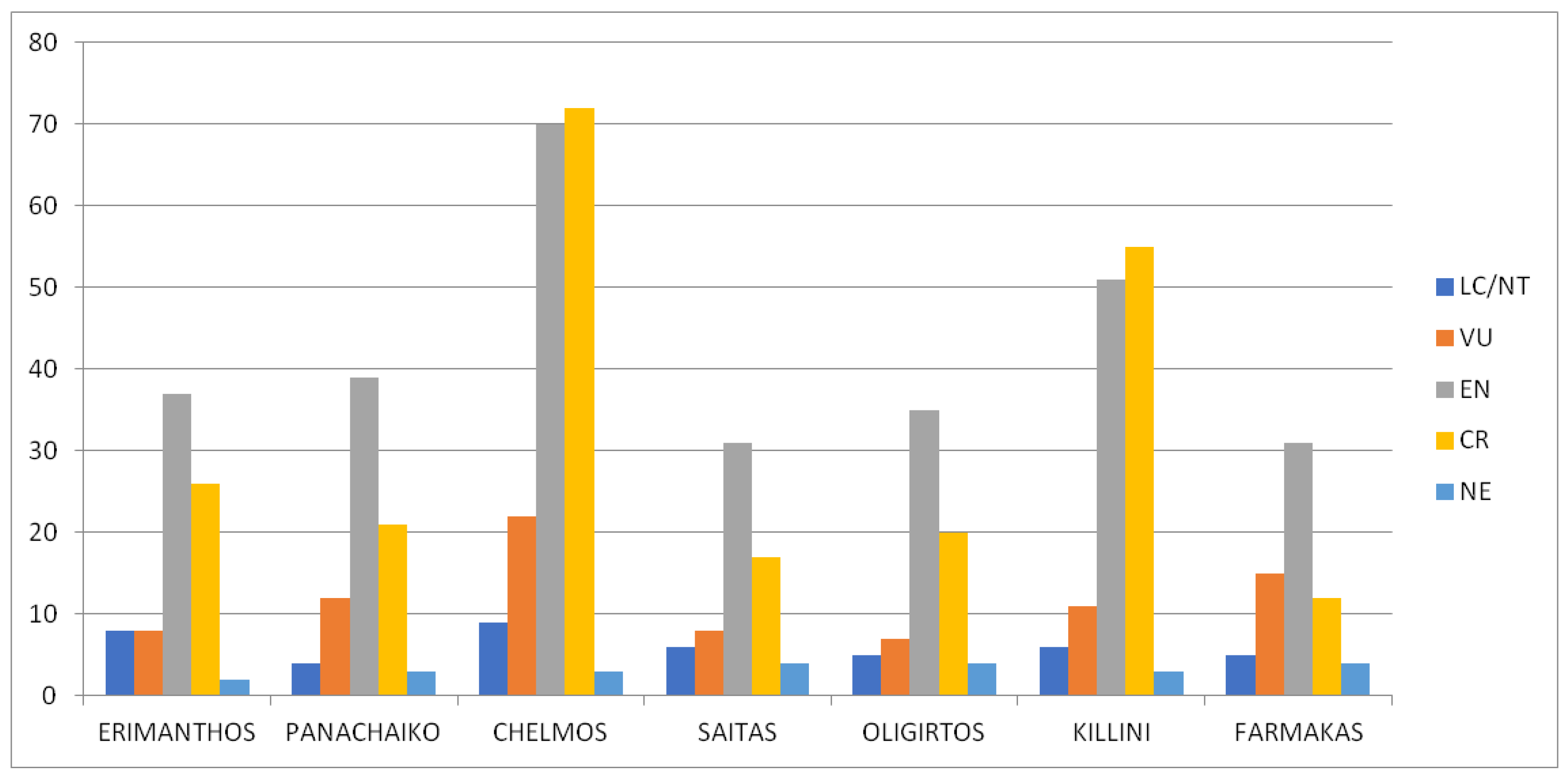
2.2.3. Habitats and IUCN Categories of Threatened Species
3. Discussion
3.1. Floristic Analysis of Mts. Oligirtos and Farmakas
3.1.1. Patterns of Endemism
3.1.2. Biological Traits of Endemics
3.1.3. Conservation Priorities
3.2. Comparative Analysis of Endemic Flora among Mountains of N Peloponnisos
3.2.1. Diversity of Endemic Taxa
3.2.2. Life Forms
3.2.3. Habitats and IUCN Categories of Threatened Species
4. Materials and Methods
4.1. Analyses of Original Data
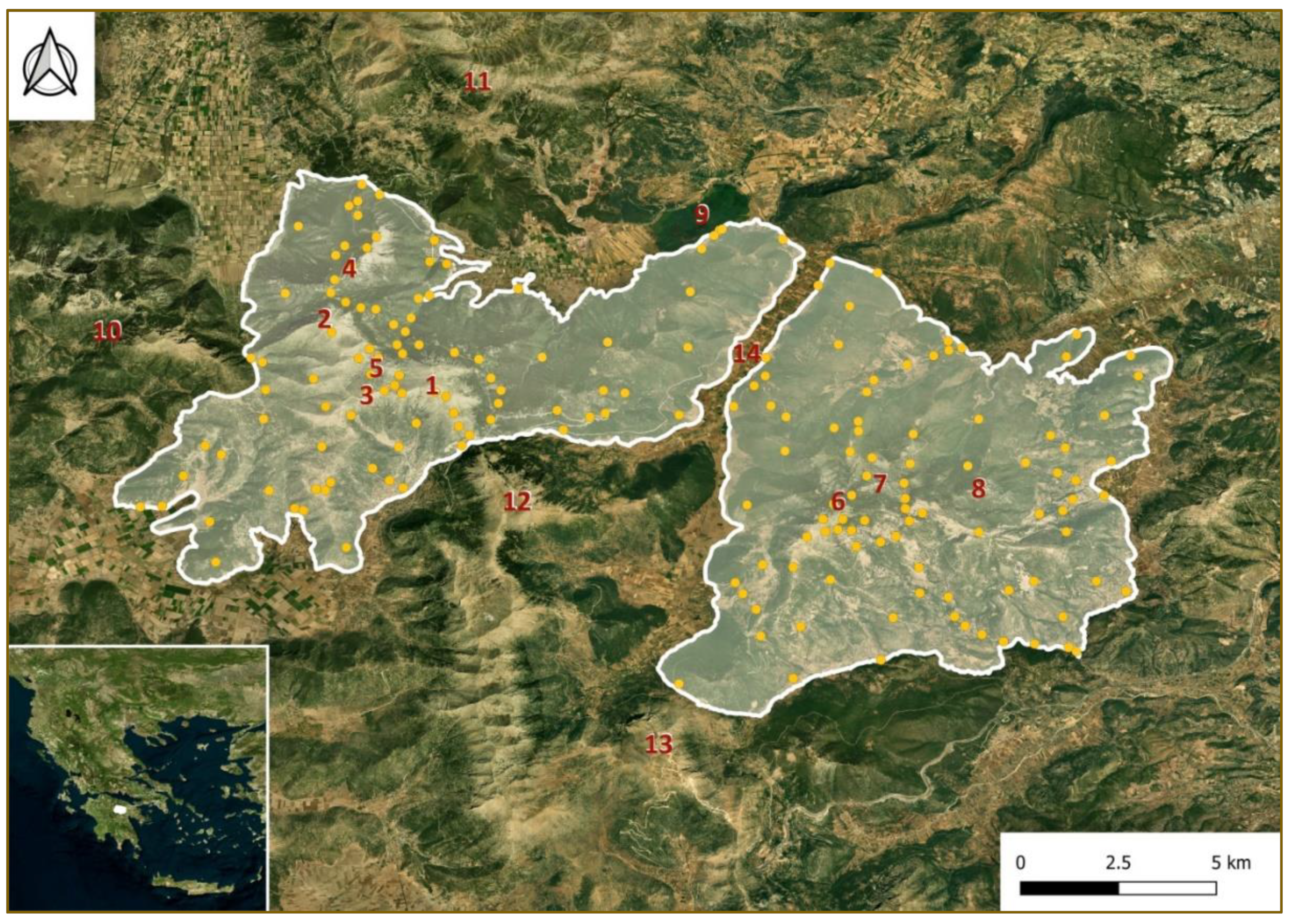
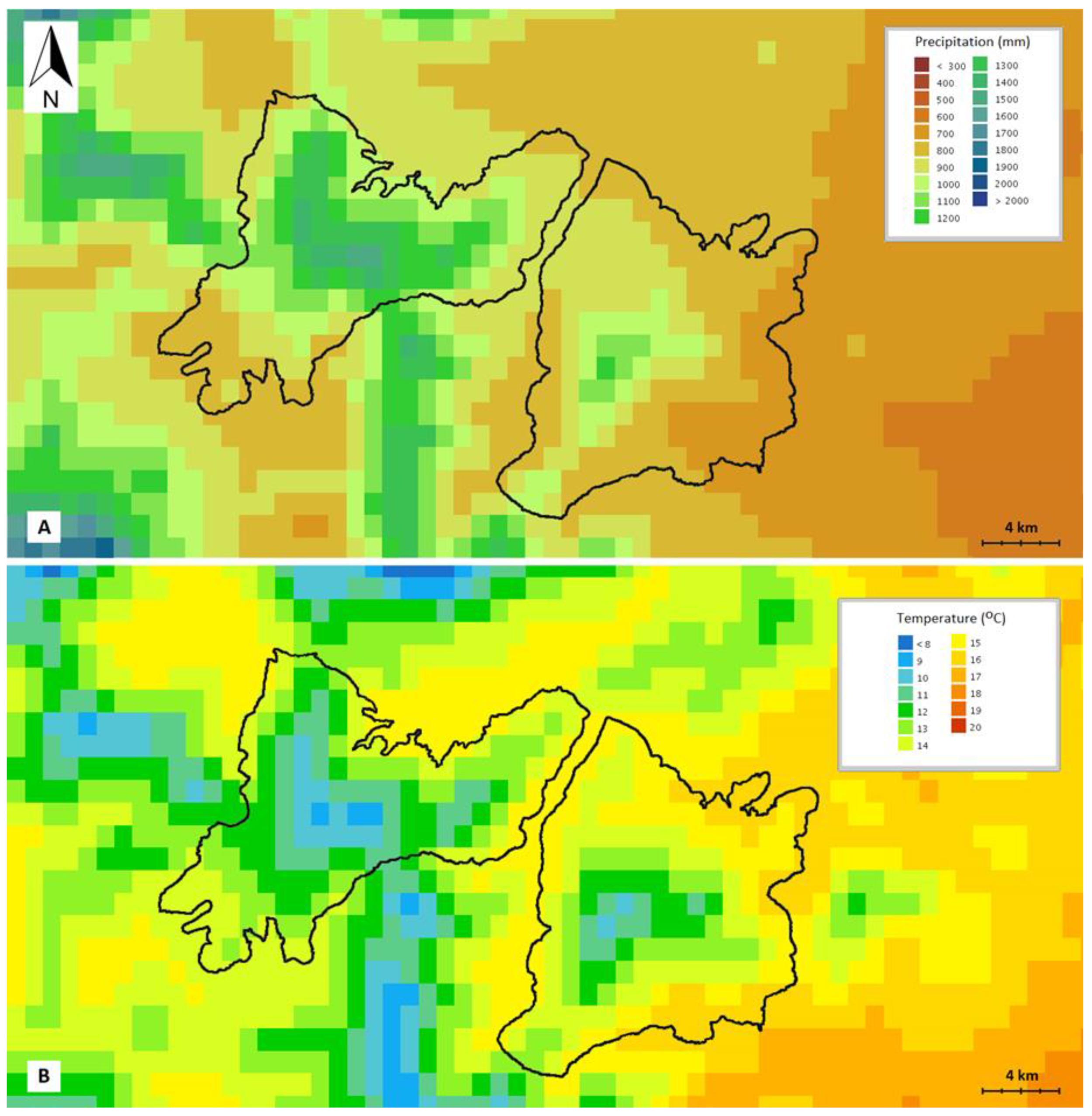
4.1.1. Study Area
4.1.2. Literature Review
4.1.3. Data Collection in the Field
4.1.4. Nomenclature
4.1.5. Chorology, Life Forms, Habitats, and IUCN categories
4.1.6. Mapping Hotspots of Endemic Taxa
4.1.7. Biological Traits of Endemics
4.1.8. Prioritization of Plant Taxa
- i.
- ii.
- iii.
- iv.
- v.
- vi.
4.1.9. Statistical Analyses Software
4.2. Comparative Analysis of Endemic Flora among Mountains of N Peloponnisos
Supplementary Materials
Author Contributions
Funding

Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cuttelod, A.; García, N.; Abdul Malak, D.; Temple, H.; Katariya, V. The Mediterranean: A biodiversity hotspot under threat. In Wildlife in a Changing World—The 2008 Review of The IUCN Red List of Threatened Species; Vié, J.C., Hilton-Taylor, C.C., Stuart, S.N., Eds.; IUCN: Gland, Switzerland, 2008; pp. 89–101. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Médail, F.; Quézel, P. Hot-Spots Analysis for conservation of Plant Biodiversity in the Mediterranean Basin. Ann. Missouri Bot. Gard. 1997, 84, 112–127. [Google Scholar] [CrossRef]
- Dimopoulos, P.; Raus, T.; Bergmeier, E.; Constantinidis, T.; Iatrou, G.; Kokkini, S.; Strid, A.; Tzanoudakis, D. Vascular plants of Greece: An annotated checklist. Englera 2013, 31, 1–372. Available online: http://www.jstor.org/stable/24365847 (accessed on 28 August 2022). [CrossRef] [Green Version]
- Dimopoulos, P.; Raus, T.; Bergmeier, E.; Constantinidis, T.; Iatrou, G.; Kokkini, S.; Strid, A.; Tzanoudakis, D. Vascular plants of Greece: An annotated checklist. Supplement. Willdenowia 2016, 46, 301–347. [Google Scholar] [CrossRef] [Green Version]
- Strid, A.; Tan, K. Flora Hellenica; Koeltz: Königstein, Germany, 1997; Volume 1. [Google Scholar]
- Mermygkas, D.; Zikos, A.; Constantinidis, T. Biological traits, habitat preferences and endemism in the flora of Peloponnisos, Greece. Fl. Medit. 2021, 31, 37–52. [Google Scholar] [CrossRef]
- Cheminal, A.; Kokkoris, I.P.; Zotos, A.; Strid, A.; Dimopoulos, P. Assessing the Ecosystem Services Potential of Endemic Floras: A Systematic Review on the Greek Endemics of Peloponnese. Sustainability 2022, 14, 5926. [Google Scholar] [CrossRef]
- Tan, K.; Iatrou, G. Endemic Plants of Greece—The Peloponnese; Gad Publishers Ltd.: Copenhagen, Denmark, 2001. [Google Scholar]
- Strid, A.; Tan, K. Recent progress in plant taxonomy and floristic studies in Greece. Bot. Serbica 2017, 41, 123–152. [Google Scholar] [CrossRef]
- Trigas, P.; Tsiftsis, S.; Tsiripidis, I.; Iatrou, G. Distribution Patterns and Conservation Perspectives of the Endemic Flora of Peloponnese (Greece). Folia Geobot. 2012, 47. [Google Scholar] [CrossRef]
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Strid, A.; Dimopoulos, P. Extinction Risk Assessment of the Greek Endemic Flora. Biology 2021, 10, 195. [Google Scholar] [CrossRef] [PubMed]
- European Commission Council. Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Communities 1992, 206, 7–50. [Google Scholar]
- Commission of the European Communities. Interpretation Manual of European Union Habitats—EUR 27; European Commission DG Environment—Nature and Biodiversity: Brussels, Belgium, 2007. [Google Scholar]
- Mermygkas, D.; Yannitsaros, A. A floristic report from Mt Saitas, North Peloponnese, Greece. Phytol. Balcan. 2015, 21, 315–349. [Google Scholar]
- Tsakiri, M.; Koumoutsou, E.; Kokkoris, I.P.; Trigas, P.; Iliadou, E.; Tzanoudakis, D.; Dimopoulos, P.; Iatrou, G. National Park and UNESCO Global Geopark of Chelmos-Vouraikos (Greece): Floristic Diversity, Ecosystem Services and Management Implications. Land 2022, 11, 33. [Google Scholar] [CrossRef]
- Dimopoulos, P. Floristic and Phytosociological Research of Mountain Killini—Ecological Approach. Ph.D. Thesis, University of Patras, Patras, Greece, 1993. (In Greek, with English summary). [Google Scholar]
- Georghiou, K.; Delipetrou, P. Patterns and traits of the endemic plants of Greece. Bot. J. Linn. Soc. 2010, 162, 130–422. [Google Scholar] [CrossRef] [Green Version]
- Pérez-García, F.J.; Medina-Cazorla, J.M.; Martínez-Hernández, F.; Garrido-Becerra, J.A.; Mendoza-Fernández, A.J.; Salmerón-Sánchez, E.; Mota, J.F. Iberian Baetic endemic flora and the implications for a conservation policy. Ann. Bot. Fennici 2012, 49, 43–54. [Google Scholar] [CrossRef]
- Panitsa, M.; Kokkoris, I.P.; Kougioumoutzis, K.; Kontopanou, A.; Bazos, I.; Strid, A.; Dimopoulos, P. Linking Taxonomic, Phylogenetic and Functional Plant Diversity with Ecosystem Services of Cliffs and Screes in Greece. Plants 2021, 10, 992. [Google Scholar] [CrossRef]
- Sciandrello, S.; Guarino, R.; Minissale, P.; Spampinato, G. The endemic vascular flora of Peloritani Mountains (NE Sicily): Plant functional traits and phytogeographical relationships in the most isolated and fragmentary micro-plate of the Alpine orogeny. Plant Biosyst. 2015, 149, 838–854. [Google Scholar] [CrossRef] [Green Version]
- Di Musciano, M.; Carranza, M.L.; Frate, L.; Di Cecco, V.; Di Martino, L.; Frattaroli, A.R.; Stanisci, A. Distribution of Plant Species and Dispersal Traits along Environmental Gradients in Central Mediterranean Summits. Diversity 2018, 10, 58. [Google Scholar] [CrossRef] [Green Version]
- Faegri, K.; Van Der Pijl, L. Principles of Pollination Ecology; Pergamon Press: Oxford, UK, 1979. [Google Scholar]
- Médail, F.; Quézel, P. Biodiversity Hotspots in the Mediterranean Basin: Setting Global Conservation Priorities. Conserv. Biol. 1999, 13, 1510–1513. [Google Scholar] [CrossRef]
- Grau, O.; Ninot, J.M.; Ferré, A.; Font, X.; Grytnes, J. Altitudinal species richness patterns of vascular plants in the south-eastern Pyrenees and nearby mountains of Catalonia. Plant Ecol. Divers. 2012, 5, 115–126. [Google Scholar] [CrossRef]
- Nezis, N. The Greek Mountain; Anavasis, Hellenic Federation of Mountaineering and Climbing: Athens, Greece, 2010; Volume 2. (In Greek) [Google Scholar]
- Lekkas, E.; Skourtsos, E.; Karalemas, N.; Papaspyropoulos, K.; Filis, C. Investigation of the Possibility of Increasing the Water Abstraction Quantities by the Municipal Enterprise for Water Supply and Sewerage of Argos–Mycenae with Water from the Springs of Agios Georgios Kiveri (Anavalos); University of Athens—Department of Geology and Geoenvironment: Athens, Greece, 2017. (In Greek) [Google Scholar]
- Bagnouls, F.; Gaussen, H. Les climats biologiques et leur classification. Ann. Géogr. 1957, 66, 193–220. [Google Scholar] [CrossRef]
- HNMS (Hellenic National Meteorological Service), Climatic Data for Selected Stations in Greece. Available online: http://www.emy.gr/emy/en/climatology/climatology_city?perifereia=Peloponnese&poli=Tripoli (accessed on 20 July 2022).
- Emberger, L. Une classification biogéographique des climats. Recueil Trav. Lab. Bot. Fac. Sci. Univ. Montpellier Sér. Bot. 1955, 7, 3–43. [Google Scholar]
- Emberger, L. Orientation Actuelle au Service de la C.G.V. de la Cartographie Physiologique Appliqué; Centre National de la Recherche Scientifique: Paris, France, 1959; Volume 4. [Google Scholar]
- Sauvage, C. Le quotient pluviothermique d’ Emberger, son utilization et la représentation géographique de ses variations au Maroc. Ann. Ser. Phys. Globe Météo. Inst. Sc. Chér. 1963, 20, 11–23. [Google Scholar]
- HNMS (Hellenic National Meteorological Service), Climatic Atlas of Greece 1971–2000. Available online: http://climatlas.hnms.gr/sdi/ (accessed on 17 June 2022).
- Strid, A. Mountain Flora of Greece; Cambridge University Press: Cambridge, UK, 1986; Volume 1. [Google Scholar]
- Strid, A.; Tan, K. Mountain Flora of Greece; Edinburgh University Press: Edinburgh, UK, 1991; Volume 2. [Google Scholar]
- Vladimirov, V.; Dane, F.; Tan, K. Comp. New floristic records in the Balkans: 11. Phytol. Balcan. 2009, 15, 273–289. [Google Scholar]
- Vladimirov, V.; Dane, F.; Tan, K. Comp. New floristic records in the Balkans: 17. Phytol. Balcan. 2011, 17, 361–384. [Google Scholar]
- Vladimirov, V.; Dane, F.; Stevanović, V.; Tan, K. Comp. New floristic records in the Balkans: 22. Phytol. Balcan. 2013, 19, 267–303. [Google Scholar]
- Vladimirov, V.; Dane, F.; Matevski, V.; Tan, K. Comp. New floristic records in the Balkans: 25. Phytol. Balcan. 2014, 20, 267–310. [Google Scholar]
- Vladimirov, V.; Dane, F.; Matevski, V.; Tan, K. Comp. New floristic records in the Balkans: 27. Phytol. Balcan. 2015, 21, 189–219. [Google Scholar]
- Vladimirov, V.; Dane, F.; Matevski, V.; Tan, K. Comp. New floristic records in the Balkans: 29. Phytol. Balcan. 2016, 22, 93–123. [Google Scholar]
- Vladimirov, V.; Dane, F.; Tan, K. Comp. New floristic records in the Balkans: 30. Phytol. Balcan. 2016, 22, 259–292. [Google Scholar]
- Vladimirov, V.; Aybeke, M.; Tan, K. Comp. New floristic records in the Balkans: 36. Phytol. Balcan. 2018, 24, 263–292. [Google Scholar]
- Vladimirov, V.; Aybeke, M.; Tan, K. Comp. New floristic records in the Balkans: 39. Phytol. Balcan. 2019, 25, 203–238. [Google Scholar]
- Vladimirov, V.; Aybeke, M.; Tan, K. Comp. New floristic records in the Balkans: 40. Phytol. Balcan. 2019, 25, 295–335. [Google Scholar]
- Strid, A. Adonis cyllenea (Ranunculaceae) and Helichrysum taenari (Asteraceae) rediscovered in Peloponnisos [with an appendix by G. Sfikas]. Ann. Mus. Goulandris 1986, 7, 221–231. [Google Scholar]
- Phitos, D.; Constantinidis, T.H.; Kamari, G. The Red Data Book of Rare and Threatened Plants of Greece; Hellenic Botanical Society: Patra, Greece, 2009; Volume 1. (In Greek) [Google Scholar]
- POWO (Plants of the World Online). Facilitated by the Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org (accessed on 2 July 2022).
- Teppner, H. Onosma kaheirei spec. nova and O. erectum (Boraginaceae) from Greece. Phyton 1988, 28, 115–131. [Google Scholar]
- Dimopoulos, P.; Raus, T.; Strid, A.; Flora of Greece Web. Vascular Plants of Greece. An Annotated Checklist. Version IV (July 2022). Available online: http://portal.cybertaxonomy.org/floragreece/ (accessed on 30 July 2022).
- Tutin, T.G.; Burges, N.A.; Chater, A.O.; Edmondson, J.R.; Heywood, V.H.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. (Eds.) Flora Europaea, 2nd ed.; Cambridge University Press: Cambridge, UK, 1993; Volume 1. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. (Eds.) Flora Europaea; Cambridge University Press: Cambridge, UK, 1968–1980; Volumes 2–5. [Google Scholar]
- Strid, A.; Tan, K. Flora Hellenica; Koeltz: Königstein, Germany, 2002; Volume 2. [Google Scholar]
- Strid, A. Atlas of the Aegean Flora, Part 1: Text & Plates; Botanischer Garten und Botanisches Museum Berlin-Dahlem: Berlin, Germany, 2016. [Google Scholar]
- Strid, A. Atlas of the Aegean Flora, Part 2: Maps; Botanischer Garten und Botanisches Museum Berlin-Dahlem: Berlin, Germany, 2016. [Google Scholar]
- Aghababyan, M.V. A revision of Papaver sect. Argemonidium Spach (Papaveraceae). Takhtajania 2011, 1, 38–43. [Google Scholar]
- Antonopoulos, Z.; Tsiftsis, S. Atlas of the Greek Orchids; Mediterraneo Editions: Rethymno, Greece, 2017; Volume 2. [Google Scholar]
- Barberá, P.; Romero-Zarco, C.; Aedo, C. Taxonomic Revision of Trisetum Sect. Trisetum (Poaceae: Pooideae: Aveninae) from Eurasia and North Africa. Ann. Missouri Bot. Gard. 2018, 103, 350–392. [Google Scholar] [CrossRef]
- Bardy, K.E.; Albach, D.C.; Schneeweiss, G.M.; Fischer, M.A.; Schönswetter, P. Disentangling phylogeography, polyploid evolution and taxonomy of a woodland herb (Veronica chamaedrys group, Plantaginaceae s.l.) in southeastern Europe. Mol. Phylogenet. Evol. 2010, 57, 771–786. [Google Scholar] [CrossRef]
- Belyaeva, I. 2009. Nomenclature of Salix fragilis L. and a new species, S. euxina (Salicaceae). Taxon 2009, 58, 1344–1348. [Google Scholar] [CrossRef]
- Boratyński, A.; Browicz, K.; Zieliński, J. Chorology of Trees and Shrubs in Greece; Poznanska Drukarnia Naukowa: Poznań, Poland, 1992. [Google Scholar]
- Browicz, K. The genus Colutea L. A monograph. Monogr. Bot. 1963, 14, 3–136+28 plates. [Google Scholar]
- Hardion, L.; Verlaque, R.; Baumel, A.; Juin, M.; Vila, B. Revised systematics of Mediterranean Arundo (Poaceae) based on AFLP fingerprints and morphology. Taxon 2012, 61, 1217–1226. [Google Scholar] [CrossRef]
- Ietswaart, J.H. A Taxonomic Revision of the Genus Origanum (Labiatae); Leiden University Press: The Hague, The Netherlands, 1980. [Google Scholar]
- Kadereit, J.W. A revision of Papaver section Argemonidium. Notes Roy. Bot. Gard. Edinburgh 1986, 44, 25–43. [Google Scholar]
- Kadereit, J.W. A revision of Papaver L. section Rhoeadium Spach. Notes Roy. Bot. Gard. Edinburgh 1989, 45, 225–286. [Google Scholar]
- Kokkini, S.; Karousou, R.; Vokou, D. Pattern of geographic variations of Origanum vulgare trichomes and essential oil content in Greece. Biochem. Syst. Ecol. 1994, 22, 517–528. [Google Scholar] [CrossRef]
- Mathew, B.F. The Crocus. A Revision of the Genus crocus (Iridacae); B.T. Batsford Ltd.: London, UK, 1982. [Google Scholar]
- Mermygkas, D.; Tan, K.; Yannitsaros, A. A new species of Iris (Iridaceae) from the northern Peloponnese (Greece). Phytol. Balcan. 2010, 16, 263–266. [Google Scholar]
- Persson, K. New species of Colchicum (Colchicaceae) from the Greek mountains. Willdenowia 1988, 18, 29–46. [Google Scholar]
- Rechinger, K.H. Der Formenkreis der Inula candida und seine Verbreitung. Oesterr. Bot. Z. 1938, 87, 81–100. [Google Scholar] [CrossRef]
- Robson, N.K. Studies in the genus Hypericum L. (Hypericaceae) 9. Addenda, corrigenda, keys, lists and general discussion. Phytotaxa 2012, 72, 1–111. [Google Scholar] [CrossRef] [Green Version]
- Širjaev, G. Generis Ononis L. revisio critica. Beih. Bot. Centralbl. 1932, 49, 381–665. [Google Scholar]
- Snogerup, S.; Snogerup, B. Bupleurum L. (Umbelliferae) in Europe - 1. The annuals, B. sect. Bupleurum and sect. Aristata. Willdenowia 2001, 31, 205–308. [Google Scholar] [CrossRef] [Green Version]
- Thomas, S.M. A taxonomic clarification of Petrorhagia section Kohlrauschia (Caryophyllaceae). Bot. J. Linn. Soc. 1983, 87, 55–75. [Google Scholar] [CrossRef]
- Wood, J.R.I.; Williams, B.R.M.; Mitchell, T.C.; Carine, M.A.; Harris, D.J.; Scotland, R.W. A foundation monograph of Convolvulus L. (Convolvulaceae). PhytoKeys 2015, 51, 1–278. [Google Scholar] [CrossRef] [Green Version]
- Zieliński, J. The genus Rosa in Greece. Arbor. Kórnickie 1991, 35, 3–45. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Version 2022-1. Available online: https://www.iucnredlist.org (accessed on 22 July 2022).
- Julve, P. Baseflor. Index botanique, écologique et chronologique de la Flore de France. Programme Catminat, 2016. [Google Scholar] [CrossRef]
- Filipe, A.F.; Lawrence, J.E.; Bonada, N. Vulnerability of stream biota to climate change in mediterranean climate regions: A synthesis of ecological responses and conservation challenges. Hydrobiologia 2013, 719, 331–351. [Google Scholar] [CrossRef]
- Cramer, W.; Guiot, J.; Fader, M.; Garrabou, J.; Gattuso, J.P.; Iglesias, A.; Lange, M.A.; Lionello, P.; Llasat, M.C.; Paz, S. Climate change and interconnected risks to sustainable development in the Mediterranean. Nat. Clim. Chang. 2018, 8, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Calleja, J.A.; Escolà, M.; Carvalho, J.; Forcadell, J.M.; Serrano, E.; Bartolomé, J. Cattle grazing fails to control shrub encroachment in Mediterranean landscapes. Rangel. Ecol. Manag. 2019, 72, 803–811. [Google Scholar] [CrossRef]
- Nadal-Romero, E.; Otal-Laín, I.; Lasanta, T.; Sánchez-Navarrete, P.; Errea, P.; Cammeraat, E. Woody encroachment and soil carbon stocks in subalpine areas in the Central Spanish Pyrenees. Sci. Total Environ. 2018, 636, 727–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bugalho, M.N.; Abreu, J.M.F. The multifunctional role of grasslands. In Sustainable Mediterranean Grasslands and Their Multi-Functions; Porqueddu, C., Tavares de Sousa, M.M., Eds.; International Centre for Advanced Mediterranean Agronomic Studies (CIHEAM): Paris, France; Food and Agriculture Organization (FAO): Rome, Italy; East Norfolk Medical Practice (ENMP): Great Yarmouth, UK; Seychelles People’s Progressive Front (SPPF): Paris, France, 2008; pp. 25–30. [Google Scholar]
- Moriondo, M.; Good, P.; Durao, R.; Bindi, M.; Giannakopoulos, C.; Corte-Real, J. Potential impact of climate change on fire risk in the Mediterranean area. Clim. Res. 2006, 31, 85–95. [Google Scholar] [CrossRef]
- Sarris, D.; Christopoulou, A.; Angelonidi, E.; Koutsias, N.; Fulé, P.Z.; Arianoutsou, M. Increasing extremes of heat and drought associated with recent severe wildfires in southern Greece. Reg. Environ. Chang. 2014, 14, 1257–1268. [Google Scholar] [CrossRef]
- Christopoulou, A.; Kazanis, D.; Fyllas, N.M.; Arianoutsou, M. Post-fire recovery of Abies cephalonica forest communities: The case of Mt Parnitha National Park, Attica, Greece. iForest 2018, 11, 757–764. [Google Scholar] [CrossRef] [Green Version]
- Urban, M.C. Escalator to extinction. Proc. Natl. Acad. Sci. USA 2018, 115, 11871–11873. [Google Scholar] [CrossRef] [Green Version]
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Trigas, P.; Strid, A.; Dimopoulos, P. Plant Diversity Patterns and Conservation Implications under Climate-Change Scenarios in the Mediterranean: The Case of Crete (Aegean, Greece). Diversity 2020, 12, 270. [Google Scholar] [CrossRef]
- Arianoutsou-Faraggitaki, M.; Margaris, N.S. Producers and the fire cycle in a phryganic ecosystem. In Components of Productivity of Mediterranean Climate Regions: Basic and Applied Aspects; Margaris, N.S., Mooney, N.A., Eds.; Dr. W Junk Publishers: The Hague, The Netherlands, 1981; Volume 4, pp. 181–190. [Google Scholar]
- Arianoutsou-Faraggitaki, M.; Margaris, N.S. Phryganic (East Mediterranean) ecosystems and fire. Ecol. Medit. 1982, 8, 473–480. [Google Scholar] [CrossRef]
- Calvo, L.; Baeza, J.; Marcos, E.; Santana, V.; Papanastasis, V.P. Post-Fire management of shrublands. In Post-Fire Management and Restoration of Southern European Forests, Managing Forest Ecosystems 24; Moreira, F., Arianoutsou, M., Corona, P., De las Heras, J., Eds.; Springer Science+Business Media B.V.: Dordrecht, The Netherlands, 2012; pp. 293–319. [Google Scholar]
- Dimopoulos, P.; Bergmeier, E.; Papastergiadou, E.; Sykora, K.; Dafis, S. Evaluation system of the relative importance of vegetation syntaxa for nature conservation in Greece. In Proceedings of the first Balkan Botanical Congress: Progress in Botanical Research, Thessaloniki, Greece, 19–22 September 1997. [Google Scholar]
- Boteva, D.; Griffiths, G.; Dimopoulos, P. Evaluation and mapping of the conservation significance of habitats using GIS: An example from Crete, Greece. J. Nat. Conserv. 2004, 12, 237–250. [Google Scholar] [CrossRef]
- Kontopanou, A.; Panitsa, M. Habitat Islands on the Aegean Islands (Greece): Elevational Gradient of Chasmophytic Diversity, Endemism, Phytogeographical Patterns and need for Monitoring and Conservation. Diversity 2020, 12, 33. [Google Scholar] [CrossRef] [Green Version]
- Liu, U.; Kenney, S.; Breman, E.; Cossu, T.A. A multicriteria decision making approach to prioritise vascular plants for species-based conservation. Biol. Conserv. 2019, 234, 221–240. [Google Scholar] [CrossRef]
- Maroulis, G. Flora and Vegetation of Mount Erimanthos Ecosystems (NW Peloponnisos). Ph.D. Thesis, University of Patras, Patras, Greece, 2003. (In Greek, with English summary). [Google Scholar]
- Kokkoris, I. Study on the Flora and Vegetation of Mt. Panachaiko. Ecological Evaluation, Management Proposals and Establishment of a Bio-Monitoring Program Using Remote Sensing Methods and Geographical Information Systems. Ph.D. Thesis, University of Patras, Patras, Greece, 2014. (In Greek, with English summary). [Google Scholar]
- Vladimirov, V.; Tan, K. Comp. New floristic records in the Balkans: 16. Phytol. Balcan. 2011, 17, 246–264. [Google Scholar]
- Vladimirov, V.; Dane, F.; Tan, K. Comp. New floristic records in the Balkans: 19. Phytol. Balcan. 2012, 18, 205–230. [Google Scholar]
- Vladimirov, V.; Tan, K. Comp. New floristic records in the Balkans: 24. Phytol. Balcan. 2014, 20, 99–137. [Google Scholar]
- Vladimirov, V.; Dane, F.; Tan, K. Comp. New floristic records in the Balkans: 26. Phytol. Balcan. 2015, 21, 53–91. [Google Scholar]
- Vladimirov, V.; Dane, F.; Tan, K. Comp. New floristic records in the Balkans: 28. Phytol. Balcan. 2015, 21, 367–399. [Google Scholar]
- Vladimirov, V.; Aybeke, M.; Matevski, V.; Tan, K. Comp. New floristic records in the Balkans: 33. Phytol. Balcan. 2017, 23, 281–329. [Google Scholar]
- Vladimirov, V.; Aybeke, M.; Tan, K. Comp. New floristic records in the Balkans: 37. Phytol. Balcan. 2018, 24, 397–461. [Google Scholar]
- Vladimirov, V.; Aybeke, M.; Tan, K. Comp. New floristic records in the Balkans: 46. Phytol. Balcan. 2021, 27, 373–411. [Google Scholar]
- Constantinidis, T.; Kalpoutzakis, E. Plant Guide to Μount Parnon and Moustos Wetland Protected Area; Management Body Μount Parnon & Moustos Wetland: Astros, Greece, 2015. [Google Scholar]
- Sørensen, Τ. A method of establishing groups of equal amplitude in plant sociology based on similarity of species content. K. Dansk. Vidensk. Selsk. Biol. Skr. 1948, 5, 1–34. [Google Scholar]
- Diserud, O.H.; Ødegaard, F. A multiple-site similarity measure. Biol. Lett. 2007, 3, 20–22. [Google Scholar] [CrossRef]
- Georgiou, O. Petrorhagia (Ser.) Link. In Flora Hellenica; Strid, A., Tan, K., Eds.; Koeltz: Königstein, Germany, 1997; Volume 1, pp. 333–343. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Total Score | IUCN Category |
|---|---|---|
| Adonis cyllenea Boiss., Heldr. and Orph. | 20 | CR |
| Viola oligyrtia Tiniakou | 20 | EN |
| Crataegus pycnoloba Boiss. and Heldr. | 15 | LC |
| Erysimum pectinatum Bory and Chaub. | 15 | EN |
| Cirsium hypopsilum Boiss. and Heldr. | 15 | CR |
| Noccaea graeca (Jord.) F.K. Mey. | 15 | CR |
| Onosma erecta Sm. subsp. malickyi Teppner | 15 | CR |
| Convolvulus mairei Halácsy | 12 | EN |
| Endemism Category | rs | p |
|---|---|---|
| Total | 0.8929 | 0.01 |
| 50 km | 0.7208 | 0.07 |
| Pe | 0.7456 | 0.05 |
| Pe and StE | 0.8108 | 0.03 |
| Rest GR | 0.8524 | 0.01 |
| Panachaiko | Chelmos | Saitas | Oligirtos | Killini | Farmakas | |
|---|---|---|---|---|---|---|
| Erimanthos | 0.75 | 0.54 | 0.63 | 0.57 | 0.57 | 0.58 |
| Panachaiko | 0.49 | 0.57 | 0.55 | 0.49 | 0.55 | |
| Chelmos | 0.47 | 0.46 | 0.70 | 0.43 | ||
| Saitas | 0.73 | 0.59 | 0.66 | |||
| Oligirtos | 0.52 | 0.75 | ||||
| Killini | 0.49 |
| Habitat Abbreviation | Habitat Category Descriptor |
|---|---|
| A | Freshwater habitats (aquatic habitats, springs and fens, reedbeds and damp tall herb vegetation, seasonally flooded depressions, damp and seepage meadows, streambanks, river and lake shores) |
| C | Cliffs, rocks, walls, ravines, boulders |
| G | Temperate and submediterranean grasslands (lowland to montane dry and mesic meadows and pastures, rock outcrops and stony ground, grassy non-ruderal verges and forest edges) |
| H | High mountain vegetation (subalpine and alpine grasslands, screes and rocks, scrub above the tree line) |
| M | Coastal habitats (marine waters and mudflats, salt marshes, sand dunes, littoral rocks, halo-nitrophilous scrub) |
| P | Xeric Mediterranean phrygana and grasslands (Mediterranean dwarf shrub formations, annual-rich pastures, and lowland screes) |
| R | Agricultural and ruderal habitats (fields, gardens and plantations, roadsides and trampled sites, frequently disturbed and pioneer habitats) |
| W | Woodlands and scrub (broadleaved and coniferous forest, riparian and mountain forest and scrub, hedges, shady woodland margins) |
| Habitat Category | Score |
| A | 6 |
| G | 5 |
| W | 4 |
| H | 3 |
| P | 2 |
| C | 1 |
| Endemism Category | Score |
| 50 km | 4 |
| Pe | 3 |
| Pe and StE | 2 |
| Rest GR | 1 |
| Mountain | Highest Peak [m a.s.l.] | Lowest Elevation Approx. [m a.s.l.] | Elevation Range Approx. [m] | Area Approx. [km2] | Main Geological Substrates * | Latitudinal Range | Longitudinal Range | Taxa (nr. of sp. and subsp.) |
|---|---|---|---|---|---|---|---|---|
| Erimanthos | 2223 | 400 | 1800 | 510 | L, F, C | 37°51’ to 38°04’ N | 21°43’ to 22°01’ E | 979 |
| Panachaiko | 1924 | 200 | 1700 | 177 | L, F, Sc, C | 38°07’ to 38°17’ N | 21°47’ to 21°57’ E | 829 |
| Chelmos | 2355 | 25 | 2300 | 655 | L, D, Sa, C, P | 37°47’ to 38°11’ N | 21°57’ to 22°19’ E | 1478 |
| Saitas | 1812 | 450 | 1400 | 127 | L, D, F, Sa | 37°43’ to 37°52’ N | 22°08’ to 22°20’ E | 800 |
| Oligirtos | 1935 | 540 | 1400 | 105 | L, D, F | 37°44’ to 37°52’ N | 22°17’ to 22°29’ Ε | 740 |
| Killini | 2375 | 200 | 2200 | 233 | L, D, Sa, C | 37°53’ to 38°03’ N | 22°21’ to 22°31’ E | 1021 |
| Farmakas | 1615 | 210 | 1400 | 115 | L, D, C | 37°42’ to 37°50’ N | 22°27’ to 22°36’ Ε | 762 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zikos, A.; Constantinidis, T. Plant Diversity of Mts. Oligirtos and Farmakas (NE Peloponnisos, Greece) with Emphasis on Their Endemic Flora. Plants 2022, 11, 2649. https://doi.org/10.3390/plants11192649
Zikos A, Constantinidis T. Plant Diversity of Mts. Oligirtos and Farmakas (NE Peloponnisos, Greece) with Emphasis on Their Endemic Flora. Plants. 2022; 11(19):2649. https://doi.org/10.3390/plants11192649
Chicago/Turabian StyleZikos, Andreas, and Theophanis Constantinidis. 2022. "Plant Diversity of Mts. Oligirtos and Farmakas (NE Peloponnisos, Greece) with Emphasis on Their Endemic Flora" Plants 11, no. 19: 2649. https://doi.org/10.3390/plants11192649