
Bioprospecting Soil Bacteria from Arid Zones to Increase Plant Tolerance to Drought: Growth and Biochemical Status of Maize Inoculated with Plant Growth-Promoting Bacteria Isolated from Sal Island, Cape Verde


Abstract
:1. Introduction
2. Results
2.1. Bacterial Plant Growth-Promoting (PGP) Traits
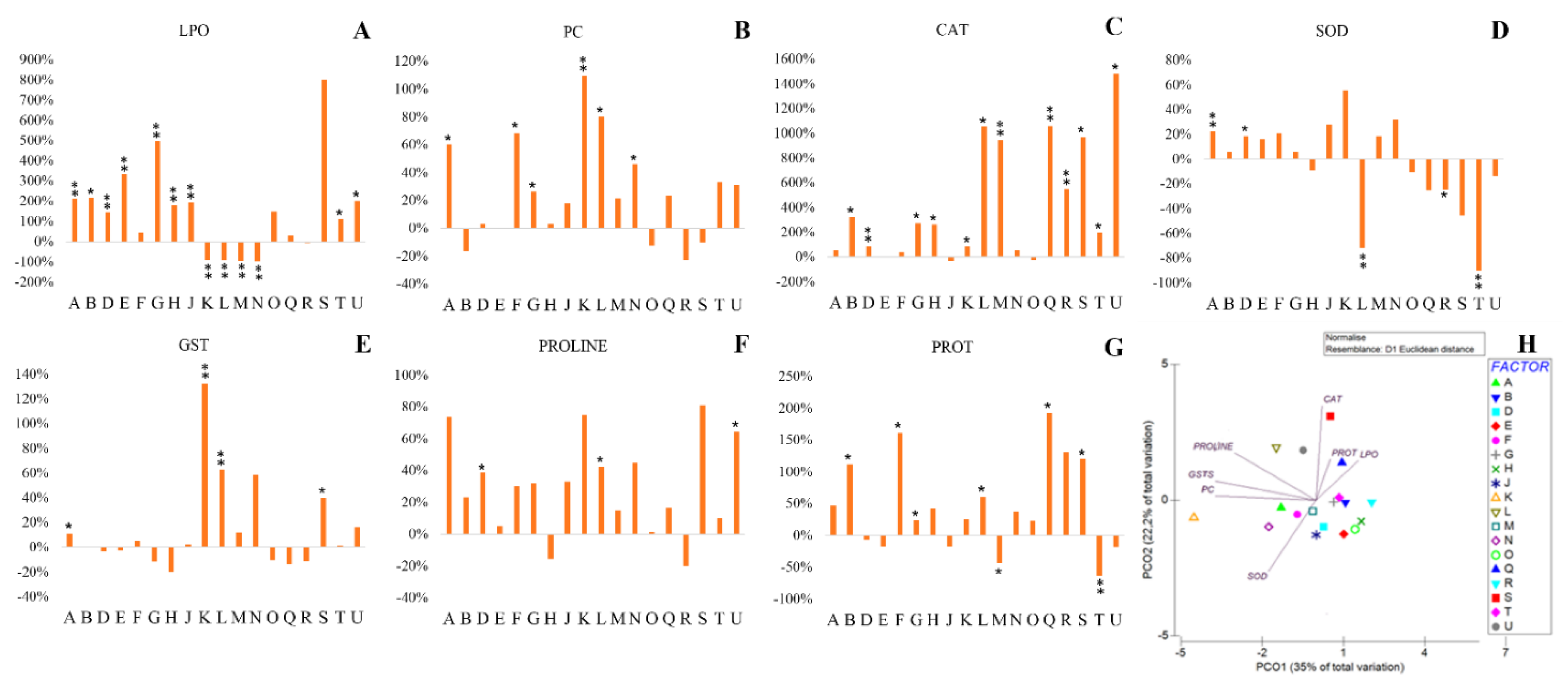
2.2. Exposure of Bacteria to PEG
2.2.1. Cellular Damage
2.2.2. Antioxidant Response
2.2.3. Osmolyte Production
2.2.4. Protein Content
2.2.5. Multivariate Analysis
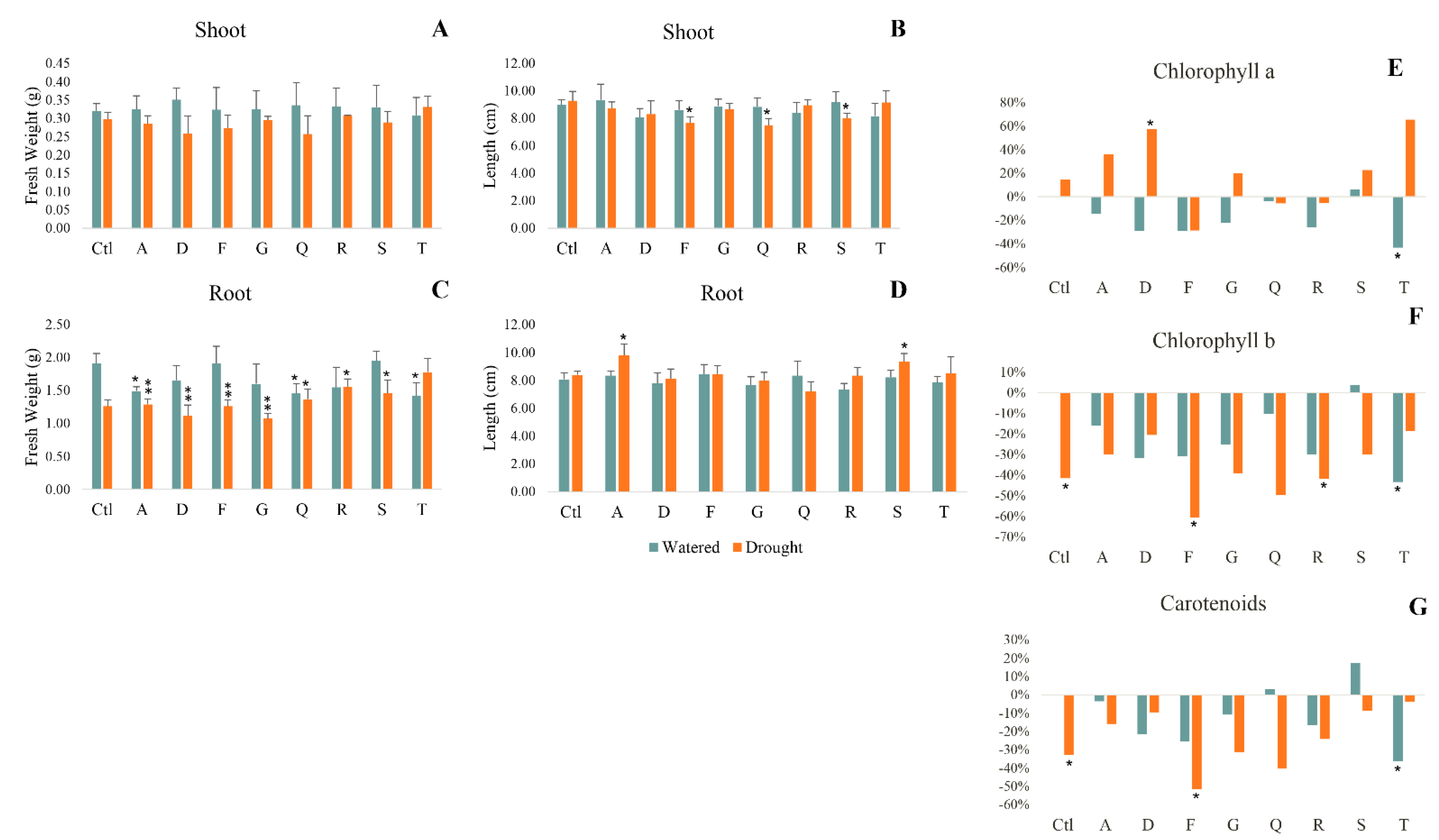
2.3. Biochemical Response of Plants to Drought
2.3.1. Biometric Parameters
2.3.2. Photosynthetic Pigments
2.3.3. Cellular Damage
2.3.4. Antioxidant Response
2.3.5. Metabolism
2.3.6. Osmolyte Production
2.3.7. Multivariate Analysis
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Tolerance to Polyethylene Glycol (PEG)
4.3. Bacterial Ability to Promote Plant Growth
4.3.1. Siderophore Production
4.3.2. Alginate Production
4.3.3. Indole Acetic acid Production
4.3.4. Phosphate Solubilization
4.4. Greenhouse Experiment
Photosynthetic Pigments
4.5. Biochemical Analysis
4.5.1. Extraction
4.5.2. Lipid Peroxidation
4.5.3. Superoxide Dismutase
4.5.4. Catalase
4.5.5. Glutathione S-Transferases
4.5.6. Protein
4.5.7. Protein Carbonylation
4.5.8. ETS
4.5.9. Soluble Sugars
4.5.10. Proline
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Nations, U. Population | United Nations. Available online: https://www.un.org/en/global-issues/population (accessed on 19 July 2022).
- United Nations Food and Agriculture Organisation. How to Feed the World in 2050. Insights from an Expert Meet; FAO: Rome, Italy, 2009; Volume 2050, pp. 1–35. [Google Scholar]
- Lutz, W.; Kc, S. Dimensions of Global Population Projections: What Do We Know about Future Population Trends and Structures? Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2779–2791. [Google Scholar] [CrossRef] [PubMed]
- Prosekov, A.Y.; Ivanova, S.A. Food Security: The Challenge of the Present. Geoforum 2018, 91, 73–77. [Google Scholar] [CrossRef]
- Ahmad, I.; Ahmad, B.; Boote, K.; Hoogenboom, G. Adaptation Strategies for Maize Production under Climate Change for Semi-Arid Environments. Eur. J. Agron. 2020, 115, 126040. [Google Scholar] [CrossRef]
- Ayub, M.; Ashraf, M.Y.; Kausar, A.; Saleem, S.; Anwar, S.; Altay, V.; Ozturk, M. Growth and Physio-Biochemical Responses of Maize (Zea mays L.) to Drought and Heat Stresses. Plant Biosyst. 2021, 155, 535–542. [Google Scholar] [CrossRef]
- Cairns, J.E.; Hellin, J.; Sonder, K.; Araus, J.L.; MacRobert, J.F.; Thierfelder, C.; Prasanna, B.M. Adapting Maize Production to Climate Change in Sub-Saharan Africa. Food Secur. 2013, 5, 345–360. [Google Scholar] [CrossRef] [Green Version]
- Kogo, B.K.; Kumar, L.; Koech, R.; Langat, P. Modelling Impacts of Climate Change on Maize (Zea mays L.) Growth and Productivity: A Review of Models, Outputs and Limitations. J. Geosci. Environ. Prot. 2019, 7, 76–95. [Google Scholar] [CrossRef] [Green Version]
- FAO. The Impact of Natural Hazards and Disasters on Agriculture; Food and Agriculture Organization of the United Nations: Rome, Italy, 2015. [Google Scholar]
- FAO. The Impact of Disasters and Crises on Agriculture and Food Security: 2021. In Impact of Disasters and Crises on Agriculture and Food Security; FAO: Rome, Italy, 2021; Volume 2021. [Google Scholar] [CrossRef]
- WHO. Drought Overview. Available online: https://www.who.int/health-topics/drought#tab=tab_1 (accessed on 18 October 2022).
- Daniel, T.; Miriam, M.; Patrick, A.; Huang, Z. Drought in Numbers. Restor. Readiness Resil. 2022, 51. Available online: https://www.unccd.int/sites/default/files/2022-05/Drought%20in%20Numbers.pdf (accessed on 18 October 2022).
- Rojas, O. Agricultural Extreme Drought Assessment at Global Level Using the FAO-Agricultural Stress Index System (ASIS). Weather Clim. Extrem. 2020, 27, 100184. [Google Scholar] [CrossRef]
- Brás, T.A.; Seixas, J.; Carvalhais, N.; Jagermeyr, J. Severity of Drought and Heatwave Crop Losses Tripled over the Last Five Decades in Europe. Environ. Res. Lett. 2021, 16, 065012. [Google Scholar] [CrossRef]
- Aroca, R. Plant Responses to Drought Stress: From Morphological to Molecular Features; Springer: Cham, Switzerland, 2013; pp. 1–466. [Google Scholar] [CrossRef]
- Abdelaal, K.; Alkahtani, M.; Attia, K.; Hafez, Y.; Király, L.; Künstler, A. The Role of Plant Growth-Promoting Bacteria in Alleviating the Adverse Effects of Drought on Plants. Biology 2021, 10, 520. [Google Scholar] [CrossRef]
- Keyvan, S. The Effects of Drought Stress on Yield, Relative Water Content, Proline, Soluble Carbohydrates and Chlorophyll of Bread Wheat Cultivars. J. Anim. Plant Sci. 2010, 8, 1051–1060. [Google Scholar]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of Salinity Stress on Plants and Its Tolerance Strategies: A Review. Environ. Sci. Pollut. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef] [PubMed]
- Masih, I.; Maskey, S.; Mussá, F.E.F.; Trambauer, P. A Review of Droughts on the African Continent: A Geospatial and Long-Term Perspective. Hydrol. Earth Syst. Sci. 2014, 18, 3635–3649. [Google Scholar] [CrossRef] [Green Version]
- FAO Infographic: Drought & Agriculture. Available online: https://www.fao.org/3/i7378e/i7378e.pdf (accessed on 18 October 2022).
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. Sustain. Agric. 2009, 29, 153–188. [Google Scholar] [CrossRef]
- Wery, J.; Silim, S.N.; Knights, E.J.; Malhotra, R.S.; Cousin, R. Screening Techniques and Sources of Tolerance to Extremes of Moisture and Air Temperature in Cool Season Food Legumes; Springer: Dordrecht, The Netherlands, 1994; pp. 439–456. [Google Scholar] [CrossRef]
- Badri, D.V.; Weir, T.L.; van der Lelie, D.; Vivanco, J.M. Rhizosphere Chemical Dialogues: Plant–Microbe Interactions. Curr. Opin. Biotechnol. 2009, 20, 642–650. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The Rhizosphere: A Playground and Battlefield for Soilborne Pathogens and Beneficial Microorganisms. Plant Soil 2008, 321, 341–361. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The Rhizosphere Microbiome and Plant Health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Yadav, V.K.; Bhagat, N.; Sharma, S.K. Modulation in Plant Growth and Drought Tolerance of Wheat Crop upon Inoculation of Drought-Tolerant-Bacillus Species Isolated from Hot Arid Soil of India. J. Pure Appl. Microbiol. 2022, 16, 246–263. [Google Scholar] [CrossRef]
- Benabdellah, K.; Abbas, Y.; Abourouh, M.; Aroca, R.; Azcón, R. Influence of Two Bacterial Isolates from Degraded and Non-Degraded Soils and Arbuscular Mycorrhizae Fungi Isolated from Semi-Arid Zone on the Growth of Trifolium Repens under Drought Conditions: Mechanisms Related to Bacterial Effectiveness. Eur. J. Soil Biol. 2011, 47, 303–309. [Google Scholar] [CrossRef]
- Glick, B.R. Plant Growth-Promoting Bacteria: Mechanisms and Applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [Green Version]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of Action of Plant Growth Promoting Bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karmakar, J.; Goswami, S.; Pramanik, K.; Maiti, T.K.; Kar, R.K.; Dey, N. Growth Promoting Properties of Mycobacterium and Bacillus on Rice Plants under Induced Drought. Plant Sci. Today 2021, 8, 49–57. [Google Scholar] [CrossRef]
- Camaille, M.; Fabre, N.; Clément, C.; Barka, E.A. Advances in Wheat Physiology in Response to Drought and the Role of Plant Growth Promoting Rhizobacteria to Trigger Drought Tolerance. Microorganisms 2021, 9, 687. [Google Scholar] [CrossRef]
- Danish, S.; Zafar-Ul-Hye, M.; Mohsin, F.; Hussain, M. ACC-Deaminase Producing Plant Growth Promoting Rhizobacteria and Biochar Mitigate Adverse Effects of Drought Stress on Maize Growth. PLoS ONE 2020, 15, e0230615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashry, N.M.; Alaidaroos, B.A.; Mohamed, S.A.; Badr, O.A.M.; El-Saadony, M.T.; Esmael, A. Utilization of Drought-Tolerant Bacterial Strains Isolated from Harsh Soils as a Plant Growth-Promoting Rhizobacteria (PGPR): Utilization of Drought-Tolerant Bacterial Strains. Saudi J. Biol. Sci. 2022, 29, 1760–1769. [Google Scholar] [CrossRef]
- Chakraborty, U.; Chakraborty, B.N.; Chakraborty, A.P.; Dey, P.L. Water Stress Amelioration and Plant Growth Promotion in Wheat Plants by Osmotic Stress Tolerant Bacteria. World J. Microbiol. Biotechnol. 2013, 29, 789–803. [Google Scholar] [CrossRef]
- García, J.E.; Maroniche, G.; Creus, C.; Suárez-Rodríguez, R.; Ramirez-Trujillo, J.A.; Groppa, M.D. In Vitro PGPR Properties and Osmotic Tolerance of Different Azospirillum Native Strains and Their Effects on Growth of Maize under Drought Stress. Microbiol. Res. 2017, 202, 21–29. [Google Scholar] [CrossRef]
- Sá, C.; Cardoso, P.; Figueira, E. Alginate as a Feature of Osmotolerance Differentiation among Soil Bacteria Isolated from Wild Legumes Growing in Portugal. Sci. Total Environ. 2019, 681, 312–319. [Google Scholar] [CrossRef]
- Kajić, S.; Puljko, A.; Rajnović, I.; Sikora, S. Resistance of Indigenous Bradyrhizobium Japonicum Strains on Moisture Deficiency Stress. J. Cent. Eur. Agric. 2020, 21, 285–291. [Google Scholar] [CrossRef]
- Cardoso, P.; Alves, A.; Silveira, P.; Sá, C.; Fidalgo, C.; Freitas, R.; Figueira, E. Bacteria from Nodules of Wild Legume Species: Phylogenetic Diversity, Plant Growth Promotion Abilities and Osmotolerance. Sci. Total Environ. 2018, 645, 1094–1102. [Google Scholar] [CrossRef]
- Bano, Q.; Ilyas, N.; Bano, A.; Zafar, N.; Akram, A.; Fayaz, A.; Hassan, U.L. Effect of azospirillum inoculation on maize (Zea mays L.) under drought stress. Pak. J. Bot 2013, 45, 13–20. [Google Scholar]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; Enshasy, H. El Plant Growth Promoting Rhizobacteria (PGPR) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Behrooz, A.; Vahdati, K.; Rejali, F.; Lotfi, M.; Sarikhani, S.; Leslie, C. Arbuscular Mycorrhiza and Plant Growth-Promoting Bacteria Alleviate Drought Stress in Walnut. HortScience 2019, 54, 1087–1092. [Google Scholar] [CrossRef] [Green Version]
- Devi, N.S.A.; Kumutha, K.; Anandham, R.; Krishnamoorthy, R. Induction of Moisture Stress Tolerance by Bacillus and Paenibacillus in Pigeon Pea (Cajanus cajan. L.). 3 Biotech 2021, 11, 355. [Google Scholar] [CrossRef] [PubMed]
- Grobelak, A.; Napora, A.; Kacprzak, M. Using Plant Growth-Promoting Rhizobacteria (PGPR) to Improve Plant Growth. Ecol. Eng. 2015, 84, 22–28. [Google Scholar] [CrossRef]
- Vardharajula, S.; Ali, S.Z.; Grover, M.; Reddy, G.; Bandi, V. Drought-Tolerant Plant Growth Promoting Bacillus Spp.: Effect on Growth, Osmol Ytes, and Antioxidant Status of Maize under Drought Stress. J. Plant Interact. 2011, 6, 1–14. [Google Scholar] [CrossRef]
- Rudolph, N.; Labuschagne, N.; Aveling, T.A.S. The Effect of Plant Growth Promoting Rhizobacteria on Seed Germination and Seedling Growth of Maize. Seed Sci. Technol. 2015, 43, 507–518. [Google Scholar] [CrossRef]
- Guo, R.; Hao, W.; Gong, D. Effects of Water Stress on Germination and Growth of Linseed Seedlings (Linum usitatissimum L.), Photosynthetic Efficiency and Accumulation of Metabolites. J. Agric. Sci. 2012, 4, 253. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Gopal, M. Siderophore Production by Plant Growth Promoting Rhizobacteria. Indian J. Agric. Res. 2008, 42, 153–156. [Google Scholar]
- Wieser, C.; Pilet, P.-E. Maize Root Extension: Effects of Indol-3yl-Acetic and Gibberellic Acids. Z. Pflanzenphysiol. 1984, 113, 263–270. [Google Scholar] [CrossRef]
- Arzanesh, M.H.; Alikhani, H.A.; Khavazi, K.; Rahimian, H.A.; Miransari, M. Wheat (Triticum aestivum L.) Growth Enhancement by Azospirillum Sp. under Drought Stress. World J. Microbiol. Biotechnol. 2011, 27, 197–205. [Google Scholar] [CrossRef]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-Acetic Acid in Plant-Microbe Interactions. Antonie van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2014, 106, 85–125. [Google Scholar] [CrossRef]
- Pustovoitova, T.N.; Zhdanova, N.E.; Zholkevich, V.N. Changes in the Levels of IAA and ABA in Cucumber Leaves under Progressive Soil Drought. Russ. J. Plant Physiol. 2004, 51, 513–517. [Google Scholar] [CrossRef]
- Ullah, A.; Nisar, M.; Ali, H.; Hazrat, A.; Hayat, K.; Keerio, A.A.; Ihsan, M.; Laiq, M.; Ullah, S.; Fahad, S.; et al. Drought Tolerance Improvement in Plants: An Endophytic Bacterial Approach. Appl. Microbiol. Biotechnol. 2019, 103, 7385–7397. [Google Scholar] [CrossRef] [PubMed]
- Uzma, M.; Iqbal, A.; Hasnain, S. Drought Tolerance Induction and Growth Promotion by Indole Acetic Acid Producing Pseudomonas Aeruginosa in Vigna Radiata. PLoS ONE 2022, 17, e0262932. [Google Scholar] [CrossRef] [PubMed]
- Ahemad, M.; Zaidi, A.; Khan, M.S.; Oves, M. Biological Importance of Phosphorus and Phosphate Solubilizing Microbes—An Overview. In Phosphate Solubilizing Microbes Crop Improv; Nova Science Publishers: Hauppauge, NY, USA, 2009; pp. 1–14. [Google Scholar]
- Kour, D.; Rana, K.L.; Yadav, A.N.; Sheikh, I.; Kumar, V.; Dhaliwal, H.S.; Saxena, A.K. Amelioration of Drought Stress in Foxtail Millet (Setaria italica L.) by P-Solubilizing Drought-Tolerant Microbes with Multifarious Plant Growth Promoting Attributes. Environ. Sustain. 2020, 3, 23–34. [Google Scholar] [CrossRef]
- Kour, D.; Lata Rana, K.; Sheikh, I.; Kumar, V.; Nath Yadav, A.; Singh Dhaliwal, H.; Kumar Saxena, A. Alleviation of Drought Stress and Plant Growth Promotion by Pseudomonas Libanensis EU-LWNA-33, a Drought-Adaptive Phosphorus-Solubilizing Bacterium. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2020, 90, 785–795. [Google Scholar] [CrossRef]
- Kour, D.; Rana, K.L.; Yadav, A.N.; Yadav, N.; Kumar, V.; Kumar, A.; Sayyed, R.Z.; Hesham, A.E.-L.; Dhaliwal, H.S.; Saxena, A.K. Drought-Tolerant Phosphorus-Solubilizing Microbes: Biodiversity and Biotechnological Applications for Alleviation of Drought Stress in Plants. In Plant Growth Promoting Rhizobacteria for Sustainable Stress Management; Springer: Singapore, 2019; pp. 255–308. [Google Scholar] [CrossRef]
- Pang, Z.; Zhao, Y.; Xu, P.; Yu, D. Microbial Diversity of Upland Rice Roots and Their Influence on Rice Growth and Drought Tolerance. Microorganisms 2020, 8, 1329. [Google Scholar] [CrossRef]
- Anjum, S.A.; Xie, X.Y.; Wang, L.C.; Saleem, M.F.; Man, C.; Lei, W. Morphological, Physiological and Biochemical Responses of Plants to Drought Stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar] [CrossRef]
- Yamada, M.; Morishita, H.; Urano, K.; Shiozaki, N.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Yoshiba, Y. Effects of Free Proline Accumulation in Petunias under Drought Stress. J. Exp. Bot. 2005, 56, 1975–1981. [Google Scholar] [CrossRef]
- Rajendrakumar, C.S.V.; Reddy, B.V.B.; Reddy, A.R. Proline-Protein Interactions: Protection of Structural and Functional Integrity of M4 Lactate Dehydrogenase. Biochem. Biophys. Res. Commun. 1994, 201, 957–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.; Bano, A.; Zandi, P. Effects of Exogenously Applied Plant Growth Regulators in Combination with Pgpr on the Physiology and Root Growth of Chickpea (Cicer Arietinum) and Their Role in Drought Tolerance. J. Plant Interact. 2018, 13, 239–247. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Yadav, V.; Tuteja, N.; Johri, A.K. Antioxidant Enzyme Activities in Maize Plants Colonized with Piriformospora Indica. Microbiology 2009, 155, 780–790. [Google Scholar] [CrossRef] [Green Version]
- Marasco, R.; Rolli, E.; Vigani, G.; Borin, S.; Sorlini, C.; Ouzari, H.; Zocchi, G.; Daffonchio, D. Are Drought-Resistance Promoting Bacteria Cross-Compatible with Different Plant Models? Plant Signal. Behav. 2013, 8, e26741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saikia, J.; Sarma, R.K.; Dhandia, R.; Yadav, A.; Bharali, R.; Gupta, V.K.; Saikia, R. Alleviation of Drought Stress in Pulse Crops with ACC Deaminase Producing Rhizobacteria Isolated from Acidic Soil of Northeast India. Sci. Rep. 2018, 8, 3560. [Google Scholar] [CrossRef] [PubMed]
- Kleinwächter, M.; Selmar, D. New Insights Explain That Drought Stress Enhances the Quality of Spice and Medicinal Plants: Potential Applications. Agron. Sustain. Dev. 2015, 35, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.; Bano, A.M.D.; Babar, A. Impacts of Plant Growth Promoters and Plant Growth Regulators on Rainfed Agriculture. PLoS ONE 2020, 15, e0231426. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Bahadur, I.; Maurya, B.R.; Raghuwanshi, R.; Meena, V.S.; Singh, D.K.; Dixit, J. Does a Plant Growth Promoting Rhizobacteria Enhance Agricultural Sustainability? J. Pure Appl. Microbiol. 2015, 9, 715–724. [Google Scholar]
- Vishnupradeep, R.; Bruno, L.B.; Taj, Z.; Karthik, C.; Challabathula, D.; Tripti; Kumar, A.; Freitas, H.; Rajkumar, M. Plant Growth Promoting Bacteria Improve Growth and Phytostabilization Potential of Zea Mays under Chromium and Drought Stress by Altering Photosynthetic and Antioxidant Responses. Environ. Technol. Innov. 2022, 25, 102154. [Google Scholar] [CrossRef]
- Dalle-Donne, I.; Rossi, R.; Giustarini, D.; Milzani, A.; Colombo, R. Protein Carbonyl Groups as Biomarkers of Oxidative Stress. Clin. Chim. Acta 2003, 329, 23–38. [Google Scholar] [CrossRef]
- Gietler, M.; Nykiel, M.; Orzechowski, S.; Fettke, J.; Zagdańska, B. Protein Carbonylation Linked to Wheat Seedling Tolerance to Water Deficiency. Environ. Exp. Bot. 2017, 137, 84–95. [Google Scholar] [CrossRef]
- Tiepo, A.N.; Constantino, L.V.; Madeira, T.B.; Gonçalves, L.S.A.; Pimenta, J.A.; Bianchini, E.; de Oliveira, A.L.M.; Oliveira, H.C.; Stolf-Moreira, R. Plant Growth-Promoting Bacteria Improve Leaf Antioxidant Metabolism of Drought-Stressed Neotropical Trees. Planta 2020, 251, 83. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Galván, A.; Romero-Perdomo, F.A.; Estrada-Bonilla, G.; Meneses, C.H.S.G.; Bonilla, R.R. Dry-Caribbean Bacillus Spp. Strains Ameliorate Drought Stress in Maize by a Strain-Specific Antioxidant Response Modulation. Microorganisms 2020, 8, 823. [Google Scholar] [CrossRef] [PubMed]
- Sheteiwy, M.S.; Abd Elgawad, H.; Xiong, Y.C.; Macovei, A.; Brestic, M.; Skalicky, M.; Shaghaleh, H.; Alhaj Hamoud, Y.; El-Sawah, A.M. Inoculation with Bacillus Amyloliquefaciens and Mycorrhiza Confers Tolerance to Drought Stress and Improve Seed Yield and Quality of Soybean Plant. Physiol. Plant. 2021, 172, 2153–2169. [Google Scholar] [CrossRef] [PubMed]
- Naseem, H.; Bano, A. Role of Plant Growth-Promoting Rhizobacteria and Their Exopolysaccharide in Drought Tolerance of Maize. J. Plant Interact. 2014, 9, 689–701. [Google Scholar] [CrossRef] [Green Version]
- Sandhya, V.; Ali, S.Z.; Grover, M.; Reddy, G.; Venkateswarlu, B. Effect of Plant Growth Promoting Pseudomonas Spp. on Compatible Solutes, Antioxidant Status and Plant Growth of Maize under Drought Stress. Plant Growth Regul. 2010, 62, 21–30. [Google Scholar] [CrossRef]
- Sebastiana, M.; da Silva, A.B.; Matos, A.R.; Alcântara, A.; Silvestre, S.; Malhó, R. Ectomycorrhizal Inoculation with Pisolithus Tinctorius Reduces Stress Induced by Drought in Cork Oak. Mycorrhiza 2018, 28, 247–258. [Google Scholar] [CrossRef]
- Maleki, A.; Naderi, A.; Naseri, R.; Fathi, A.; Bahamin, S.; Maleki, R. Physiological Performance of Soybean Cultivars under Drought Stress. The Effects of Supplementary Irrigation and Different Plant Densities on Morphological Traits, Yield and Its Components and Protein Content of Chickpea (Cicer arietinum L.). Sirvan Regi. Bull. Environ. Pharmacol. Life Sci. 2013, 2, 38–44. [Google Scholar]
- Ashraf, M.; Mehmood, S. Response of Four Brassica Species to Drought Stress. Environ. Exp. Bot. 1990, 30, 93–100. [Google Scholar] [CrossRef]
- Lehninger, A. Biochemistry; Worth Publishers: London, UK, 1970. [Google Scholar]
- Sá, C.; Matos, D.; Pires, A.; Cardoso, P.; Figueira, E. Effects of Volatile Sulfur Compounds on Growth and Oxidative Stress of Rhizobium Leguminosarum E20-8 Exposed to Cadmium. Sci. Total Environ. 2021, 800, 149478. [Google Scholar] [CrossRef]
- Pastore, D.; Trono, D.; Laus, M.N.; Di Fonzo, N.; Flagella, Z. Possible Plant Mitochondria Involvement in Cell Adaptation to Drought Stress A Case Study: Durum Wheat Mitochondria. J. Exp. Bot. 2007, 58, 195–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germ, M.; Kreft, I.; Stibilj, V.; Urbanc-Berčič, O. Combined Effects of Selenium and Drought on Photosynthesis and Mitochondrial Respiration in Potato. Plant Physiol. Biochem. 2007, 45, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Heidari, M.; Golpayegani, A. Effects of Water Stress and Inoculation with Plant Growth Promoting Rhizobacteria (PGPR) on Antioxidant Status and Photosynthetic Pigments in Basil (Ocimum basilicum L.). J. Saudi Soc. Agric. Sci. 2012, 11, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Wang, H.; Zhao, X.; Lu, Z.; Sun, X.; Ding, G. Role of Suillus Placidus in Improving the Drought Tolerance of Masson Pine (Pinus Massoniana Lamb.) Seedlings. Forests 2021, 12, 332. [Google Scholar] [CrossRef]
- Arora, N.K.; Verma, M. Modified Microplate Method for Rapid and Efficient Estimation of Siderophore Produced by Bacteria. 3 Biotech 2017, 7, 381. [Google Scholar] [CrossRef] [Green Version]
- Johnson, A.S.; O’Sullivan, E.; D’Aoust, L.N.; Omer, A.; Bonner-Weir, S.; Fisher, R.J.; Weir, G.C.; Colton, C.K. Quantitative Assessment of Islets of Langerhans Encapsulated in Alginate. Tissue Eng. Part C Methods 2011, 17, 435–449. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indoleacetic acid. Plant Physiol. 1951, 26, 192. [Google Scholar] [CrossRef] [Green Version]
- Lopes, T.; Cardoso, P.; Matos, D.; Rocha, R.; Pires, A.; Marques, P.; Figueira, E. Graphene Oxide Influence in Soil Bacteria Is Dose Dependent and Changes at Osmotic Stress: Growth Variation, Oxidative Damage, Antioxidant Response, and Plant Growth Promotion Traits of a Rhizobium Strain. Nanotoxicology 2022, 1–17. [Google Scholar] [CrossRef]
- Nautiyal, C.S. An Efficient Microbiological Growth Medium for Screening Phosphate Solubilizing Microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef]
- Wellburn, A.R.; Lichtenthaler, H. Formulae and Program to Determine Total Carotenoids and Chlorophylls A and B of Leaf Extracts in Different Solvents. In Advances in Photosynthesis Research; Sybesma, C., Ed.; Springer: Dordrecht, The Netherlands, 1984; Volume 2, pp. 9–12. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal Lipid Peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide Dismutase: Improved Assays and an Assay Applicable to Acrylamide Gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Johansson, L.H.; Håkan Borg, L.A. A Spectrophotometric Method for Determination of Catalase Activity in Small Tissue Samples. Anal. Biochem. 1988, 174, 331–336. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-Transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Robinson, H.W.; Hogden, C.G.; Co-investigator, N. The Biuret Reaction in the Determination of Serum Proteins. I. A Study of the Conditions Necessary for the Production of a Stable Colour Which Bears a Quantitative Relationship to the Protein Concentration. J. Biol. Chem. 1940, 135, 707–725. [Google Scholar] [CrossRef]
- Mesquita, C.S.; Oliveira, R.; Bento, F.; Geraldo, D.; Rodrigues, J.V.; Marcos, J.C. Simplified 2,4-Dinitrophenylhydrazine Spectrophotometric Assay for Quantification of Carbonyls in Oxidized Proteins. Anal. Biochem. 2014, 458, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Udenigwe, C.C.; Udechukwu, M.C.; Yiridoe, C.; Gibson, A.; Gong, M. Antioxidant Mechanism of Potato Protein Hydrolysates against in Vitro Oxidation of Reduced Glutathione. J. Funct. Foods 2016, 20, 195–203. [Google Scholar] [CrossRef]
- King, F.D.; Packard, T.T. Respiration and the Activity of the Respiratory Electron Transport System in Marine Zooplankton1. Limnol. Oceanogr. 1975, 20, 849–854. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Accession Number | Code | Isolated from | IC50 PEG | Siderophores (PSU/OD) | Alginate (μg/mL/OD) | Phosphate Solubilization (Ratio) | IAA (μg/mL/OD) | Seedling Emergence |
|---|---|---|---|---|---|---|---|---|---|
| Pantoea sp. | OM985627 | A | Acacia albida | 7.5 | 22.04 ± 2.73 | 19.25 ± 1.01 | 1.18 ± 0.09 | 4.23 ± 0.48 | ↑ |
| Klebsiella sp. | OM985630 | B | Acacia albida | 7.5 | 13.96 ± 4.39 | 19.75 ± 0.56 | 1.25 ± 0.07 | 5.60 ± 0.33 | ↔ |
| Pseudomonas sp. | OM985635 | D | Acacia albida | 7.5 | 23.43 ± 4.25 | 24.88 ± 0.94 | 1.25 ± 0.14 | 4.44 ± 0.39 | ↑ |
| Pseudomonas sp. | OM985638 | E | Acacia albida | 7.5 | 18.94 ± 3.14 | 23.42 ± 0.89 | − | 4.76 ± 0.43 | ↔ |
| Acinetobacter sp. | OM985640 | F | Acacia albida | 7.5 | 32.23 ± 5.45 | 22.66 ± 0.59 | 1.71 ± 0.21 | 3.72 ± 0.52 | ↑ |
| Stenotrophomonas sp. | OM985622 | G | Acacia albida | 7.5 | 16.12 ± 2.29 | 13.56 ± 0.80 | 1.03 ± 0.09 | 4.36 ± 0.45 | ↑ |
| Enterobacter sp. | OM985625 | H | Acacia albida | 7.5 | 13.90 ± 8.64 | 15.79 ± 0.50 | 1.97 ± 0.19 | 4.75 ± 0.46 | ↔ |
| Enterobacter sp. | OM985623 | J | Amaranthus viridis | 7.5 | 25.83 ± 1.80 | 18.27 ± 2.72 | 1.88 ± 0.27 | 9.62 ± 0.83 | ↔ |
| Pantoea sp. | OM985626 | K | Amaranthus viridis | 7.5 | 22.05 ± 7.83 | 25.55 ± 0.94 | 1.32 ± 0.12 | 9.87 ± 1.23 | ↔ |
| Pseudomonas sp. | OM985629 | L | Amaranthus viridis | 7.5 | 57.54 ± 4.89 | 15.07 ± 1.82 | 1.40 ± 0.07 | 4.68 ± 1.33 | ↔ |
| Rhizobium sp. | OM985632 | M | Acacia albida | 7.5 | 22.54 ± 5.76 | 14.12 ± 1.71 | 1.08 ± 0.14 | 4.17 ± 0.52 | ↔ |
| Paenarthrobacter sp. | OM985634 | N | Acacia albida | 12.5 | 49.94 ± 4.64 | 7.43 ± 0.76 | − | 5.71 ± 0.80 | ↔ |
| Ochrobactrum sp. | OM985637 | O | Acacia albida | 12.5 | 17.77 ± 0.96 | 12.84 ± 0.82 | 1.94 ± 0.14 | 6.15 ± 0.80 | ↓ |
| Pseudomonas sp. | OM985621 | Q | Acacia albida | 10 | 15.77 ± 5.69 | 15.94 ± 3.31 | 1.04 ± 0.03 | 6.39 ± 0.88 | ↑ |
| Rhizobium sp. | OM985624 | R | Acacia albida | 10 | 23.29 ± 8.10 | 14.71 ± 1.66 | 1.15 ± 0.03 | 4.67 ± 0.34 | ↑ |
| Stenotrophomonas sp. | OM985631 | S | Acacia albida | 7.5 | 51.83 ± 5.77 | 25.16 ± 4.08 | − | 7.23 ± 0.41 | ↑ |
| Pseudomonas sp. | OM985633 | T | Acacia albida | 10 | 12.04 ± 3.77 | 13.24 ± 1.51 | 1.38 ± 0.24 | 4.34 ± 0.61 | ↑ |
| Enterobacter sp. | OM985636 | U | Acacia albida | 10 | 13.91 ± 1.32 | 19.67 ± 2.94 | 1.88 ± 0.17 | 3.80 ± 0.57 | ↔ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz, C.; Cardoso, P.; Santos, J.; Matos, D.; Figueira, E. Bioprospecting Soil Bacteria from Arid Zones to Increase Plant Tolerance to Drought: Growth and Biochemical Status of Maize Inoculated with Plant Growth-Promoting Bacteria Isolated from Sal Island, Cape Verde. Plants 2022, 11, 2912. https://doi.org/10.3390/plants11212912
Cruz C, Cardoso P, Santos J, Matos D, Figueira E. Bioprospecting Soil Bacteria from Arid Zones to Increase Plant Tolerance to Drought: Growth and Biochemical Status of Maize Inoculated with Plant Growth-Promoting Bacteria Isolated from Sal Island, Cape Verde. Plants. 2022; 11(21):2912. https://doi.org/10.3390/plants11212912
Chicago/Turabian StyleCruz, Catarina, Paulo Cardoso, Jacinta Santos, Diana Matos, and Etelvina Figueira. 2022. "Bioprospecting Soil Bacteria from Arid Zones to Increase Plant Tolerance to Drought: Growth and Biochemical Status of Maize Inoculated with Plant Growth-Promoting Bacteria Isolated from Sal Island, Cape Verde" Plants 11, no. 21: 2912. https://doi.org/10.3390/plants11212912