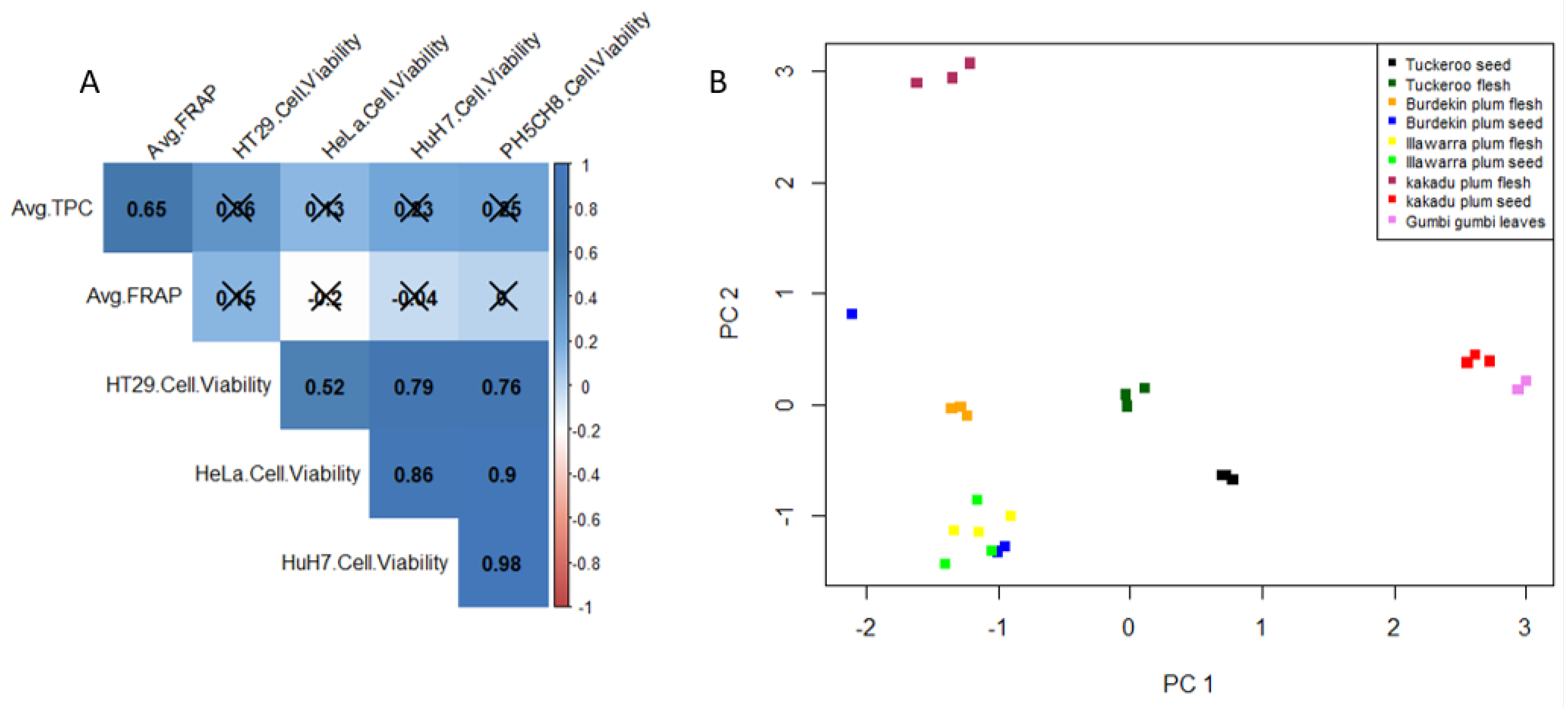
2.1. Total Phenolic Content (TPC) and Antioxidant Capacity
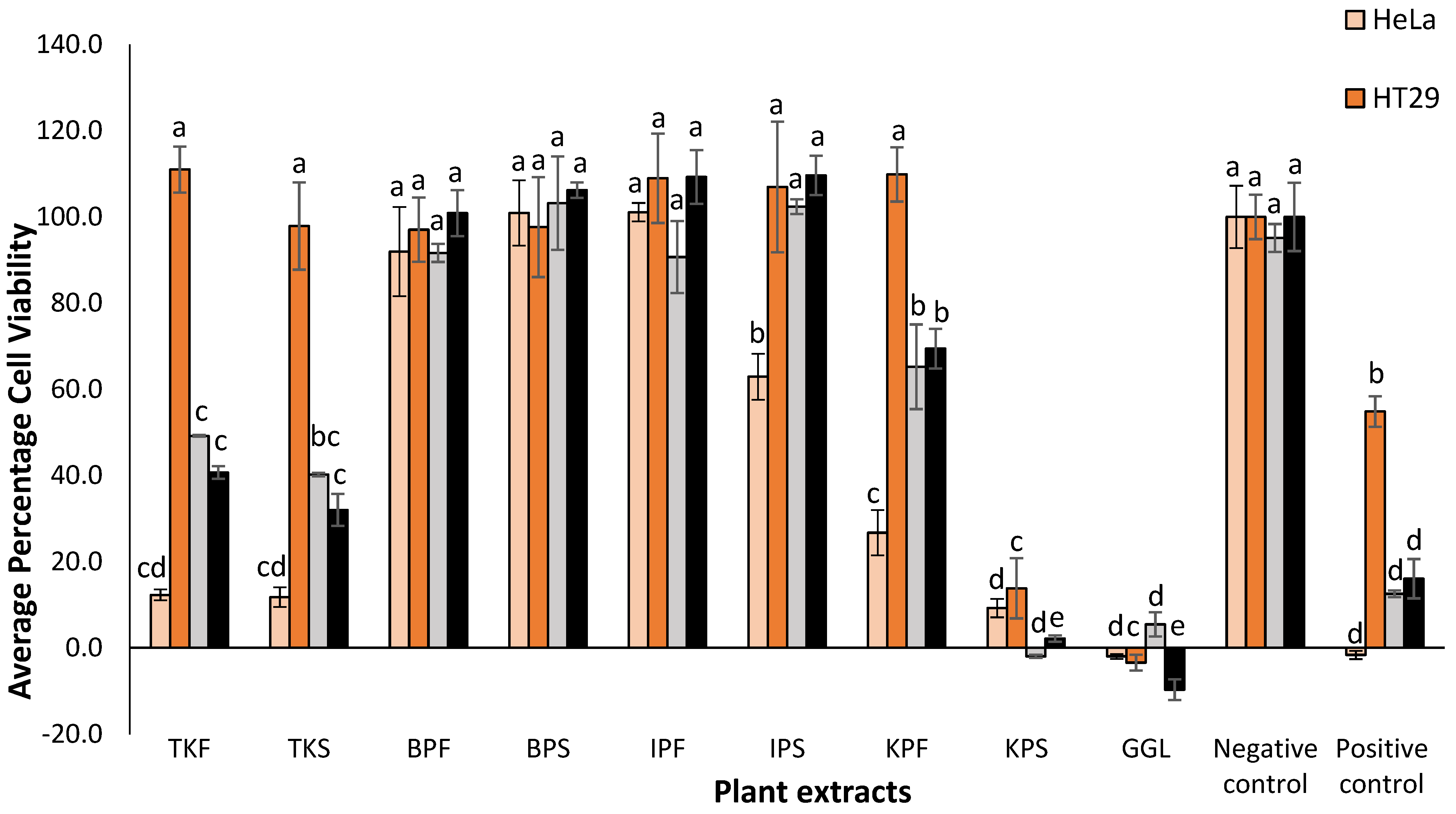
Among the five different native Australian plants studied, KPF exhibited the highest antioxidant capacity and TPC value (
Table 1). Notable values were also seen in BPF and TKF and GGL. High antioxidative capacity and total phenolic content often dictates the bioactive potential of a plant matrix [
2,
4,
5]. Hence, it was postulated that KPF would show high bioactivity in the anticancer and antimicrobial bioassays. Moreover, Kakadu plum and Illawarra plum have previously been shown to exhibit significant in vitro antioxidant, anti-inflammatory, and proapoptotic anticancer activity [
6,
7,
8,
9,
10]. Furthermore, Kakadu plum fruit have the highest recorded concentrations of ascorbic acid of any fruit in the world [
11].
However, the flesh and seeds of these fruits have not been separately tested for their phenolic content, antioxidant capacity, and bioactivity. Data obtained herein shows that the TPC and FRAP content are significantly different (p-value < 0.05) between the flesh and seeds of the plum samples. Although, the seeds have lower TPC and antioxidant capacity compared to the flesh and are often discarded, they still have valuable amounts of phenolic compounds and antioxidants which may be harnessed for therapeutic or nutritional uses.
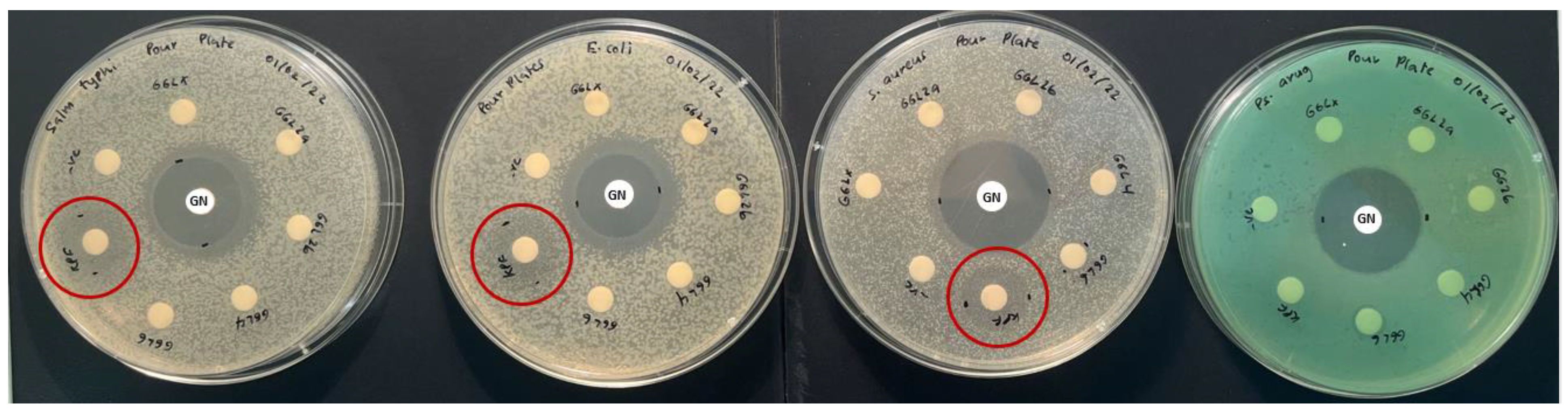
2.3. Antibacterial Activity
The antibacterial activity of the selected plant species was investigated against food poisoning bacteria using the disc diffusion method, with the results presented in
Table 3. The results revealed that only the KPF extracts were potentially effective in suppressing microbial growth of all tested bacterial strains apart from
P. aeruginosa, although it did show weaker activity than the positive control (
Figure 5). Additionally, TKS and TKF were only slightly effective against
S. aureus. Other plants extracts showed no antibacterial activity against the pathogenic bacterial strains at the tested concentration.
Previous studies have also demonstrated KPF extracts ability to inhibit the growth of both Gram-positive and Gram-negative bacteria [
10,
11,
19]. Particularly, inhibitory effects against
E. coli and
S. aureus was noted at concentrations between 3.1–15.6 mg/mL [
11]. Although Kakadu plum has shown exceptionally high ascorbic acid content, this compound alone may not be responsible for the broad antibacterial activity. Studies have shown ascorbic acid to have weak antibacterial activity towards
E. coli and
S. aureus [
11,
17,
20]. Hence, it is more likely that the interaction between ascorbic acid and polyphenols is prompting the antibacterial activity in this fruit. Akter et al. (2021) proposed that the Kakadu plum extracts possibly interfere with the bacterial cell walls and induce lysis or the presence of phenolic compounds and other lower molecular weight compounds such as K
+ and PO
43− may also be enhancing the loss of other intracellular molecules such as proteins, DNA, RNA, and other higher molecular weight materials.
Although some phytochemical studies [
21,
22] and cytotoxicity studies [
16] of tuckeroo fruit exists, no substantial studies report the antibacterial activity of this fruit. This study may be the first to report some antibacterial activity in the flesh and seed of this fruit (
Figure 6). The zone of inhibitions of the extracts at 20 mg/mL are quite small hence future dose response studies to calculate IC
50 values is suggested.
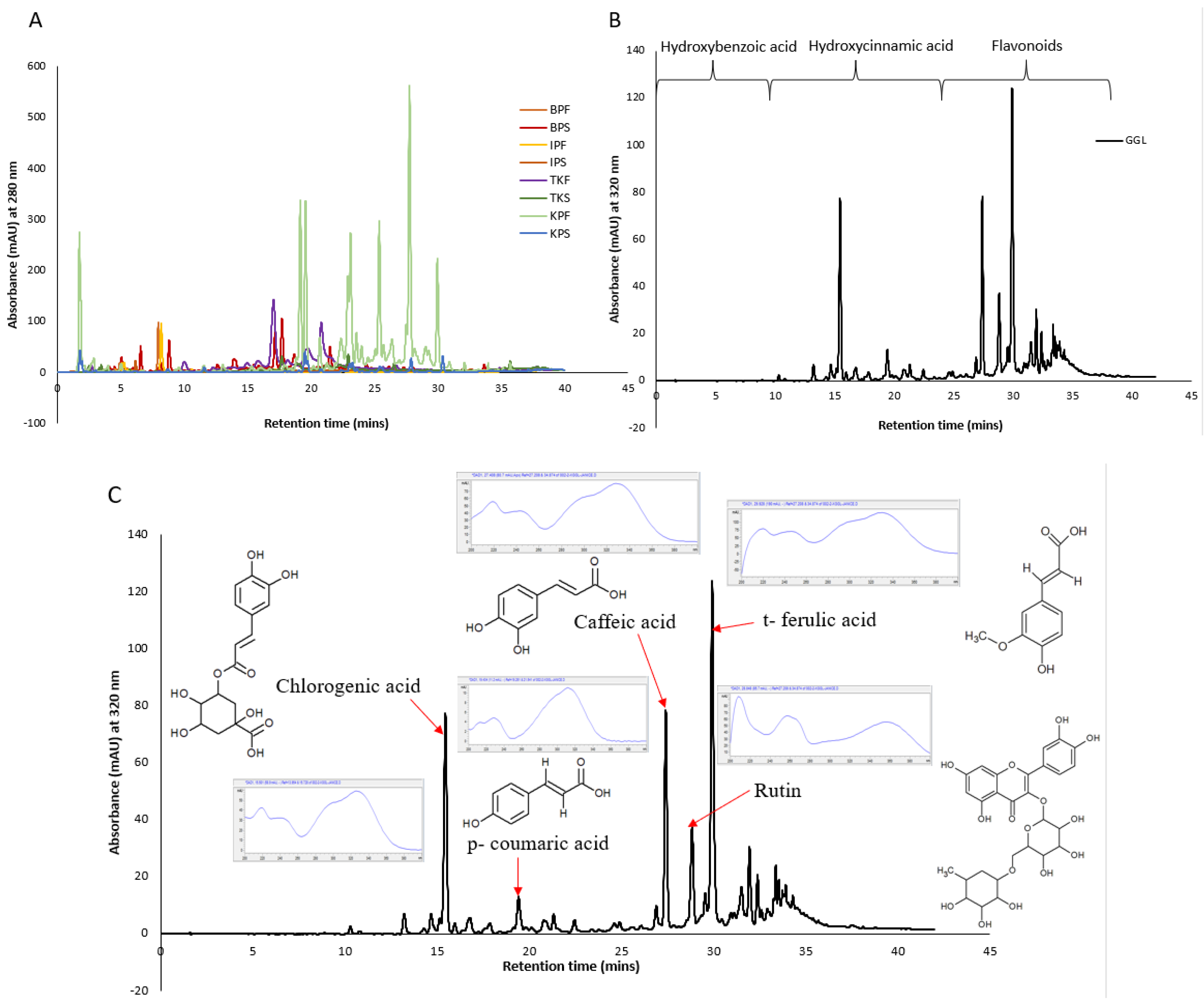
2.4. HPLC Profiling and Characterization of the Crude Extracts
HPLC phenolic profiling of the crude extracts were performed, and the chromatograms obtained are given in
Figure 7 (Panel A and B). With reference to retention time, the elution of polyphenols can be generalized as hydroxybenzoic acid < hydroxycinnamic acids < flavonoid. Hydroxybenzoic derivatives in plants include p-hydroxybenzoic, gallic, syringic, protocatechuic, and vanillic acids which are present as bound components in complex structures like lignins and hydrolysable tannins or attached to cell walls and proteins [
23]. Whereas hydroxycinnamic acid derivatives include p-coumaric, ferulic, caffeic, and sinapic acids which occur in bound forms connected to cell wall structures such as cellulose, lignin, sugars, and proteins through ester bonds. Flavonoids are ubiquitous in many plants and most commonly found ones include quercetin and rutin [
24]. High content of flavonoids (e.g., anthocyanins, flavones, flavonols and chalcones) may be found in a single structure of plants such as its seeds. Numerous peak overlaps were evident in the fruit extracts suggesting occurrence of similar compounds albeit in various concentrations as indicated by the peak intensities (
Figure 7 Panel A).
The KPF extracts demonstrated the strongest peaks which was coherent with the fact that it had the highest TPC and antioxidant capacity values. The higher concentrations of the phenolic compounds in KPF may also explain its strong antibacterial activity determined earlier (
Section 3.3). However, it is interesting to note that although KPS show lower concentrations of the phenolic compounds, it was found to be one of the most potent extracts against the cancer cell lines. These discrepancies in the data could mean that the cytotoxic effect of KPS extracts may not be due to the phenolic compounds but rather glycosidic forms of the aglycone phenols that have not been captured in the chromatogram. A previous study has successfully demonstrated that two isolated flavonoid glycosides (2′ -hydroxyl neophellamuretin and 200-O-rhamnosylswertisin) isolated from
Desmodium caudatum (Thunb.) DC had a certain growth inhibitory effect on HeLa cells in a dose dependent manner [
25]. The chromatographic separation, isolation and characterization of glycosylated phenolic compound is often challenging due to their complex structural conformity arising through varying degree of plausible glycosylation patterns and high polarity [
26].
Moreover, since GGL and Kakadu plum extracts (KPS and KPF) extracts had shown interesting bioactivity, further peak characterization was warranted. In our previous work we had tentatively identified the peaks of the GGL by comparing their UV spectrums and retention times to that of authentic standards as shown in
Figure 7 (Panel C) [
3]. A high occurrence of t-ferulic acid in the GGL extracts (
Figure 7C) was found which may explain the cytotoxic activity of GGL extracts. Ferulic acid isolated from
Ferula foetida, which is a perennial herb, has shown anticancer activity against various types of cancers such as colon and lung cancer, and central nervous system tumors previously [
27]. Additionally, studies have also demonstrated cytotoxicity of flavonoids against cancer cells and have found them to also have high free radical scavenging activity [
28]. Purified flavonoids have shown anticancer activities against a number of human carcinomas which include hepatoma (Hep-G2), cervical carcinoma (Hela), and breast cancer (MCF-7) [
28].
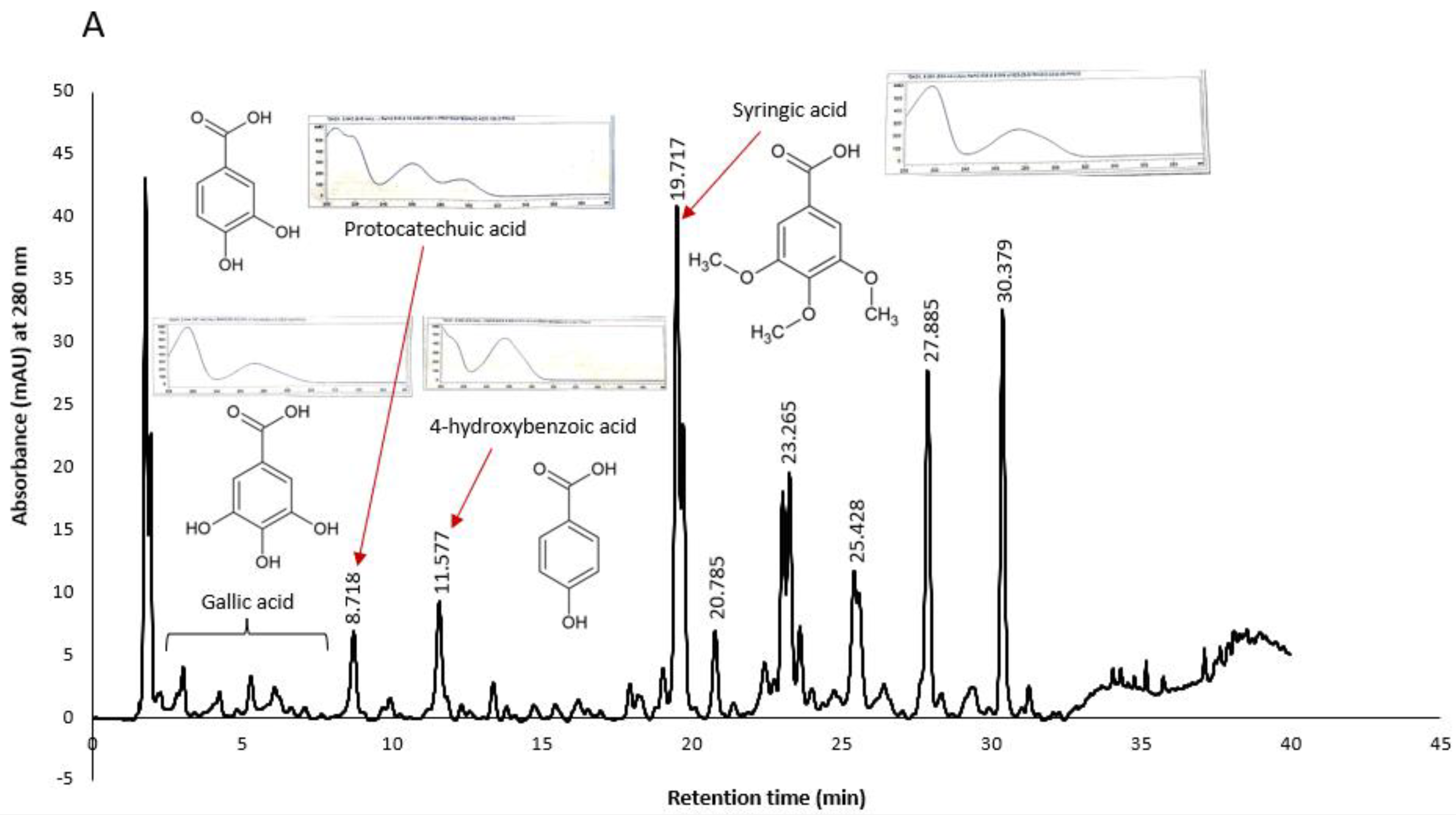
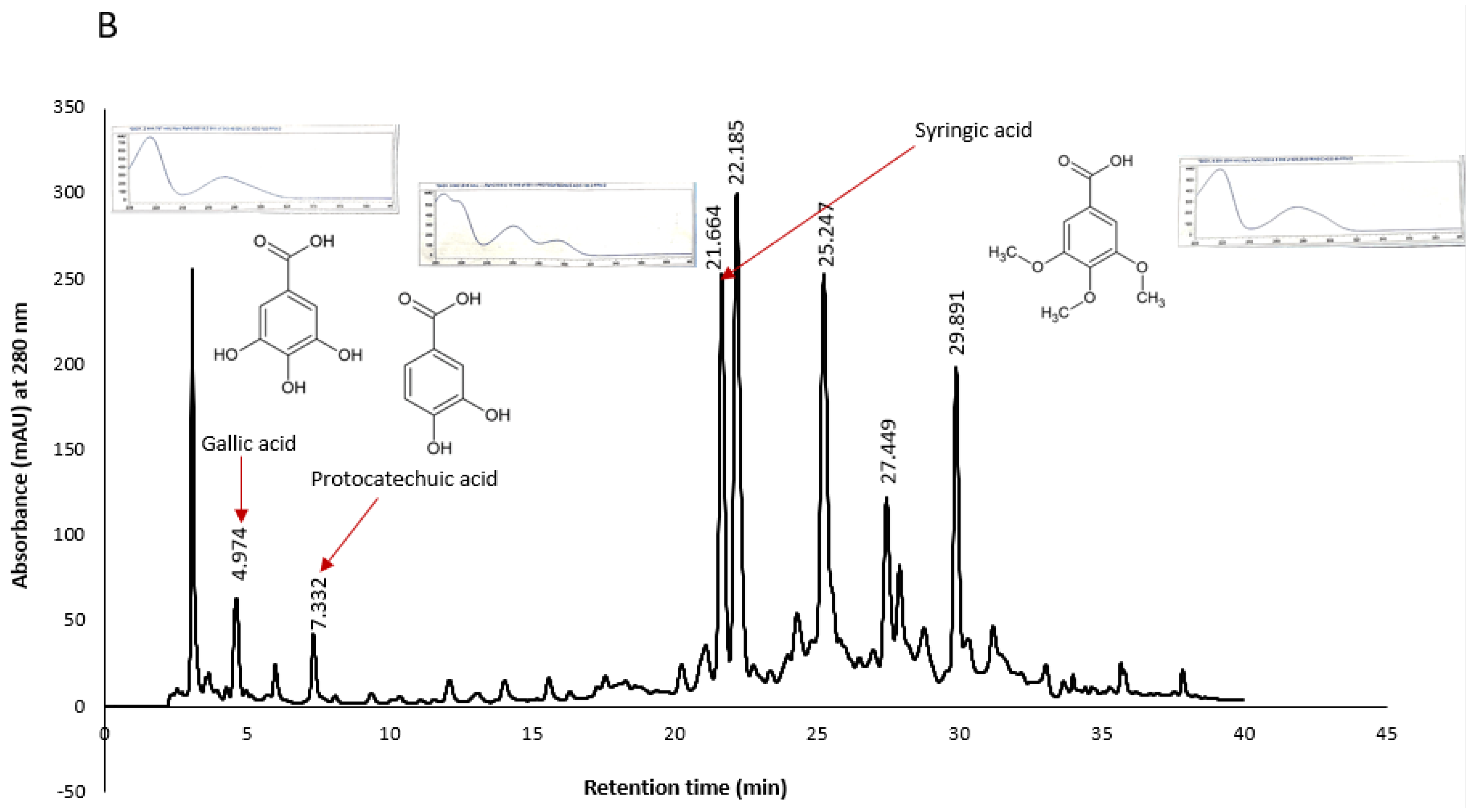
Similarly, some peaks in KPS and KPF extracts were tentatively identified as shown in
Figure 8. It was worth noting that chromatograms of both KPS and KPF extracts measured at a wavelength of 280 nm showed peaks of compounds belonging to the hydroxybenzoic acid class. More specifically both the UV spectral characteristics and retention times of the extract’s peaks closely matched that of gallic acid, protocatechuic acid, 4-hydroxybenzoic acid and syringic acid standards analyzed under similar HPLC conditions.
Konczak et al. (2014) have previously identified hydrolysable tannins and ellagic acids as the major phenolic compounds in Kakadu plum. Kakadu plum belong in the family Combretacea, order Myrtale, which has a significant chemotaxonomic feature which is the presence of ellagic acid and ellagitannins [
29]. In this study however, we were unable to identify ellagic acid peaks due to the unavailability of the standard. None the less, the spectral characteristics of the later peaks (RT > 20 min.) match literature UV spectrum of ellagic acid [
29,
30]. Therefore, it may be presumed that these peaks may be that of ellagic acid. Kakadu plum is unique in that it is the only fruit with the highest ascorbic acid content and that unlike other common fruits, hydroxycinnamic acid and flavonoid compounds are not its major phytochemicals. Further fractionation of Kakadu plum extracts and structural elucidation of compounds for the development of novel antioxidant and/or anticancer drugs is merited.
2.5. Cytotoxic Activity of GGL Fractions
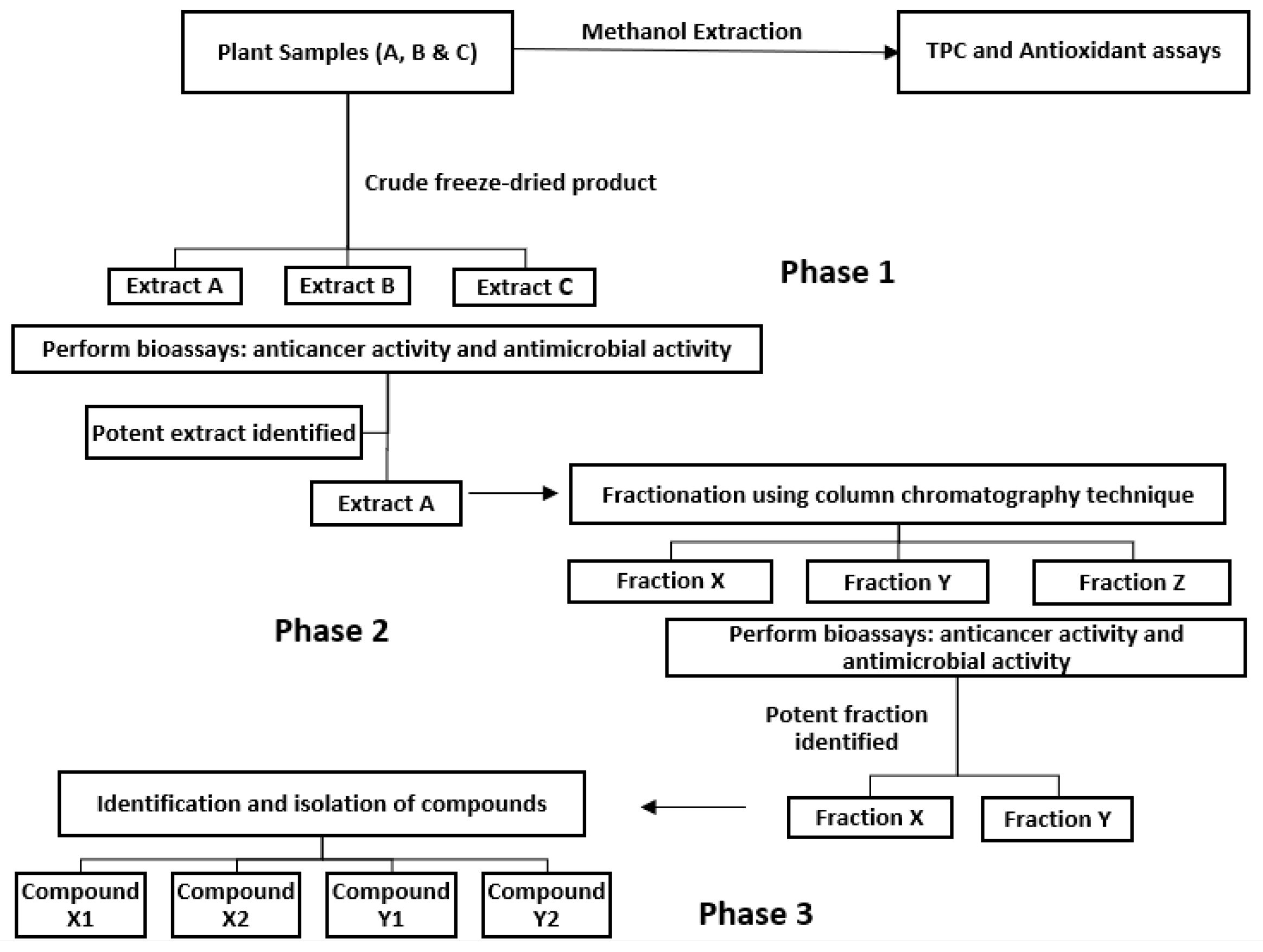
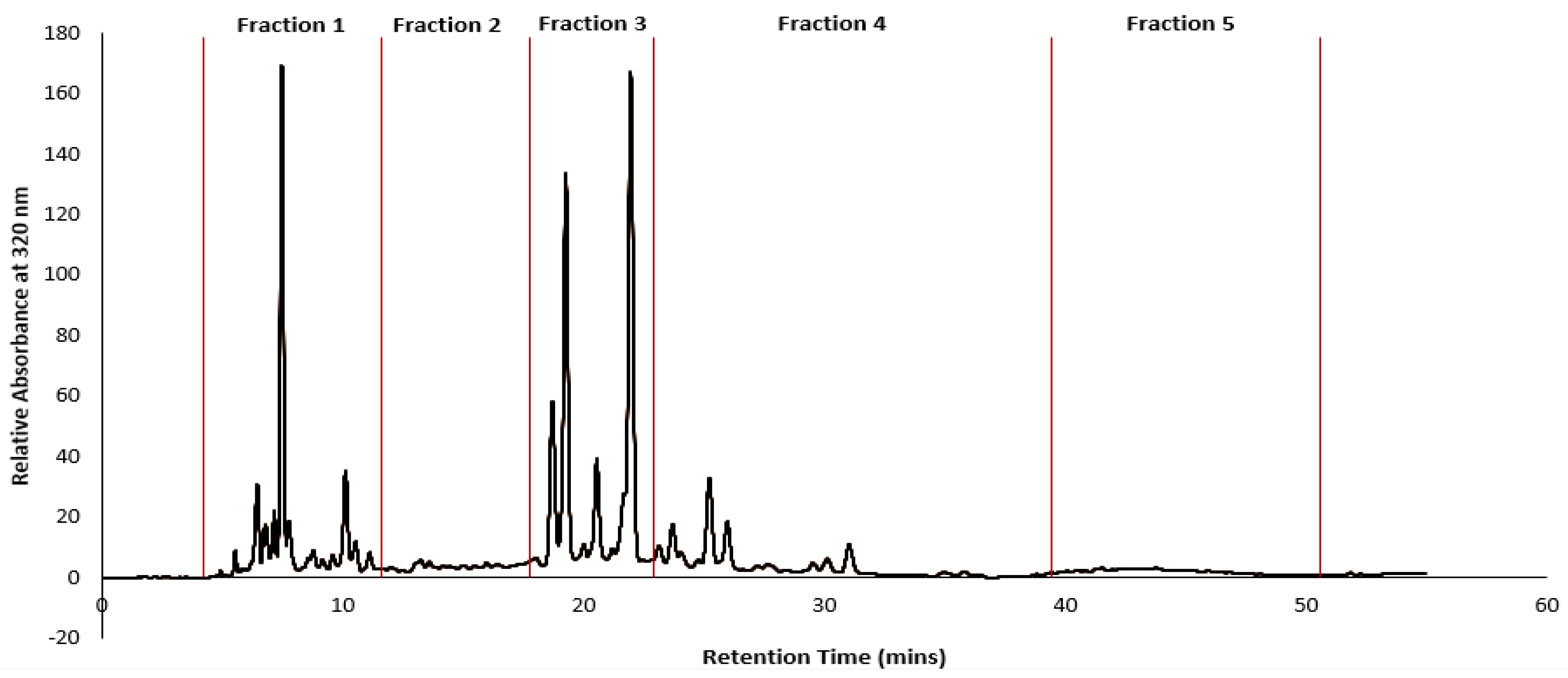
Given the complete inhibition of cancer cell lines subjected to GGL crude extracts, HPLC time-based fractionation of the extracts were performed as shown in
Figure 9 (Phase 2- Bioassay guided fractionation protocol,
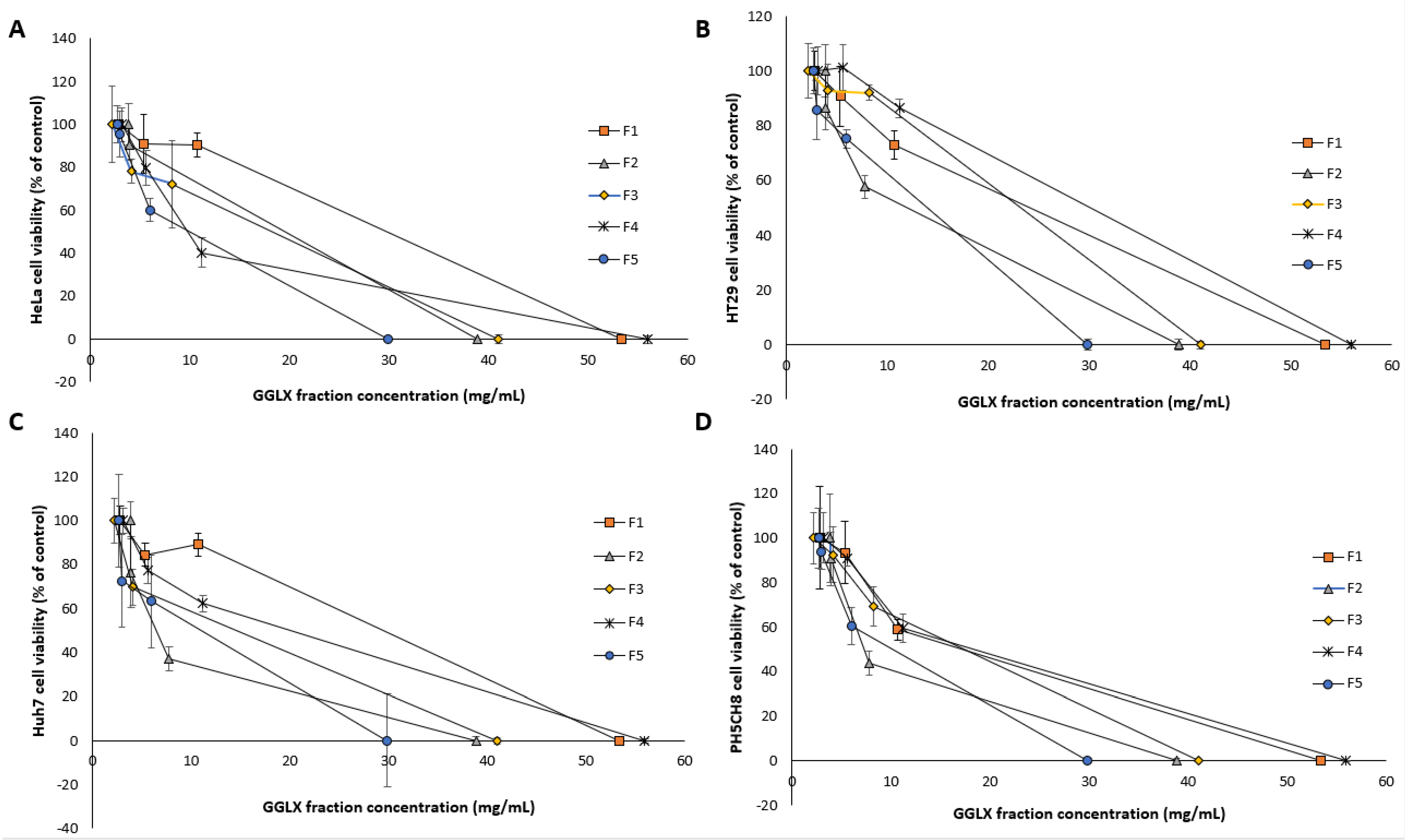
Figure 1). Dose dependent inhibitory effects of the fractions were determined, and the normalized percentage cell viability data are given in
Figure 10. The IC
50 and SI values of the GGL fractions (F1–F5) against the tested cell lines were calculated and is presented in
Table 4.
Highest cytotoxic activities were postulated for Fractions 1 (F1) and Fraction 3 (F3) due to high peak intensities. However, the IC
50 values of Fraction 2 (F2) was the lowest (0.6–0.92 mg/mL) against the cancer cell lines, deeming it to be most potent. In the F2 time frame of the chromatogram (
Figure 9), absence of phenolic peaks suggest that the cytotoxic activity of this fraction may likely be due to other compounds in the matrix rather than polyphenols. Although F2 presented ideal IC
50 values in comparison to the other extracts, its SI values was quite low (
Table 5).
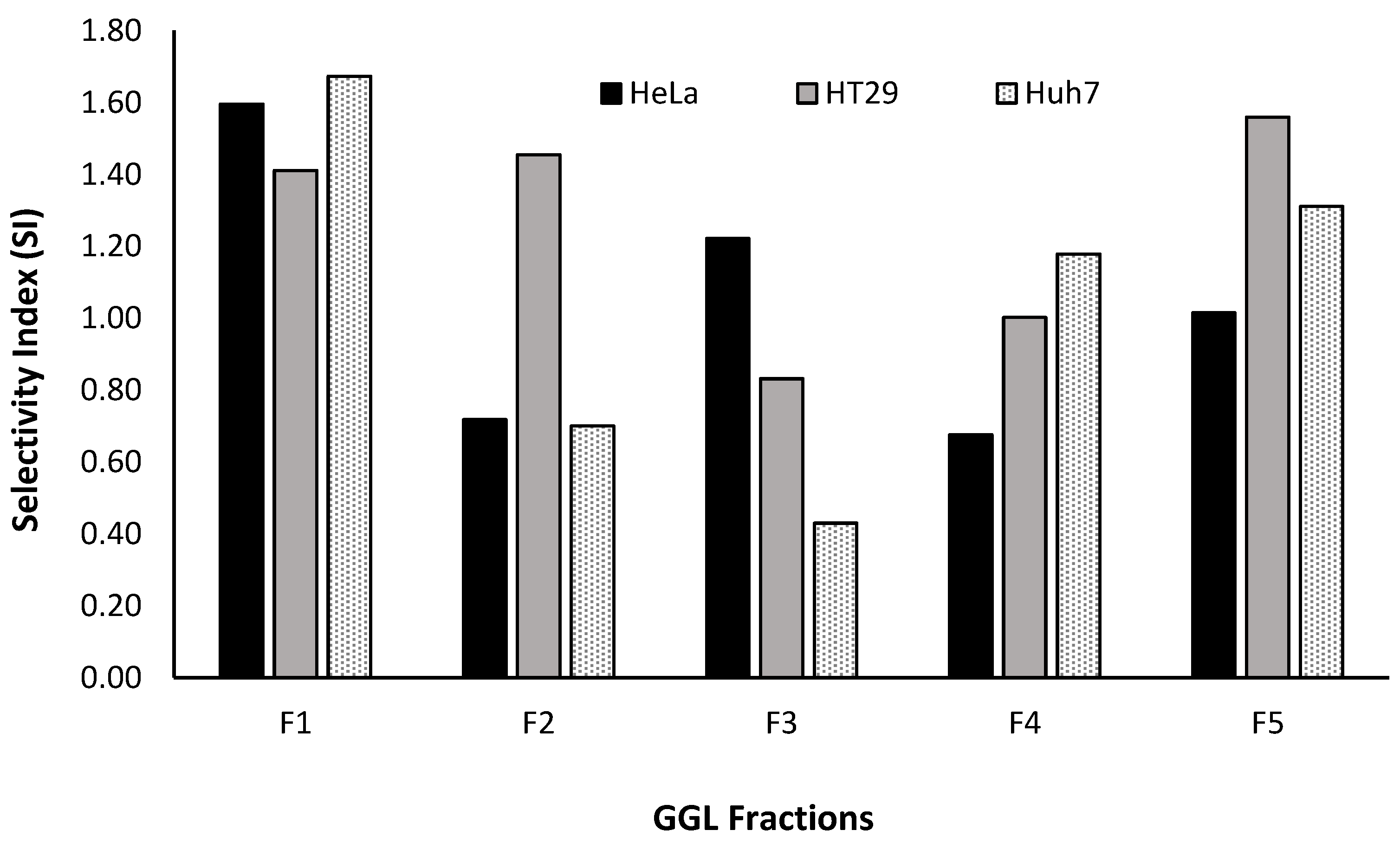
The SI values of the fractions were graphed as shown in
Figure 11 and it was construed that F1, having the highest SI value for HeLa, HT29 and HuH7 (1.60, 1.41 and 1.67, respectively) would be an ideal choice for further fractionation and isolation process (Phase 3- Bioassay guided fractionation protocol,
Figure 1). Thus far, the proposed bioassay guided fractionation protocol has successfully demonstrated an effective and robust screening process of the native Australian fruits and GGL samples analyzed. In last decade this technique has been successfully utilized to isolate various effective compounds from plants such as quinine, morphine, paclitaxel, camptothecin, etopoide, mevastin and astemisinin [
31].
Previously taraxastane-type triterpene saponins and triterpene glycosides, which have also demonstrated antiproliferative activity have been isolated from GGL [
32,
33,
34,
35]. However, due to the occurrence of several varietals of this plant which yield thousands of complex compounds, there still remains research opportunities. Hence, it is anticipated that further fractionation of GGL F1 may possibly lead to the isolation and purification of lead or novel bioactive compound(s).



 ,
, 
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}