
Pre-Germinative Treatments and Morphophysiological Traits in Enterolobium cyclocarpum and Piscidia piscipula (Fabaceae) from the Yucatan Peninsula, Mexico

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Seed Viability
2.2. Imbibed Seeds and Imbibition Rate
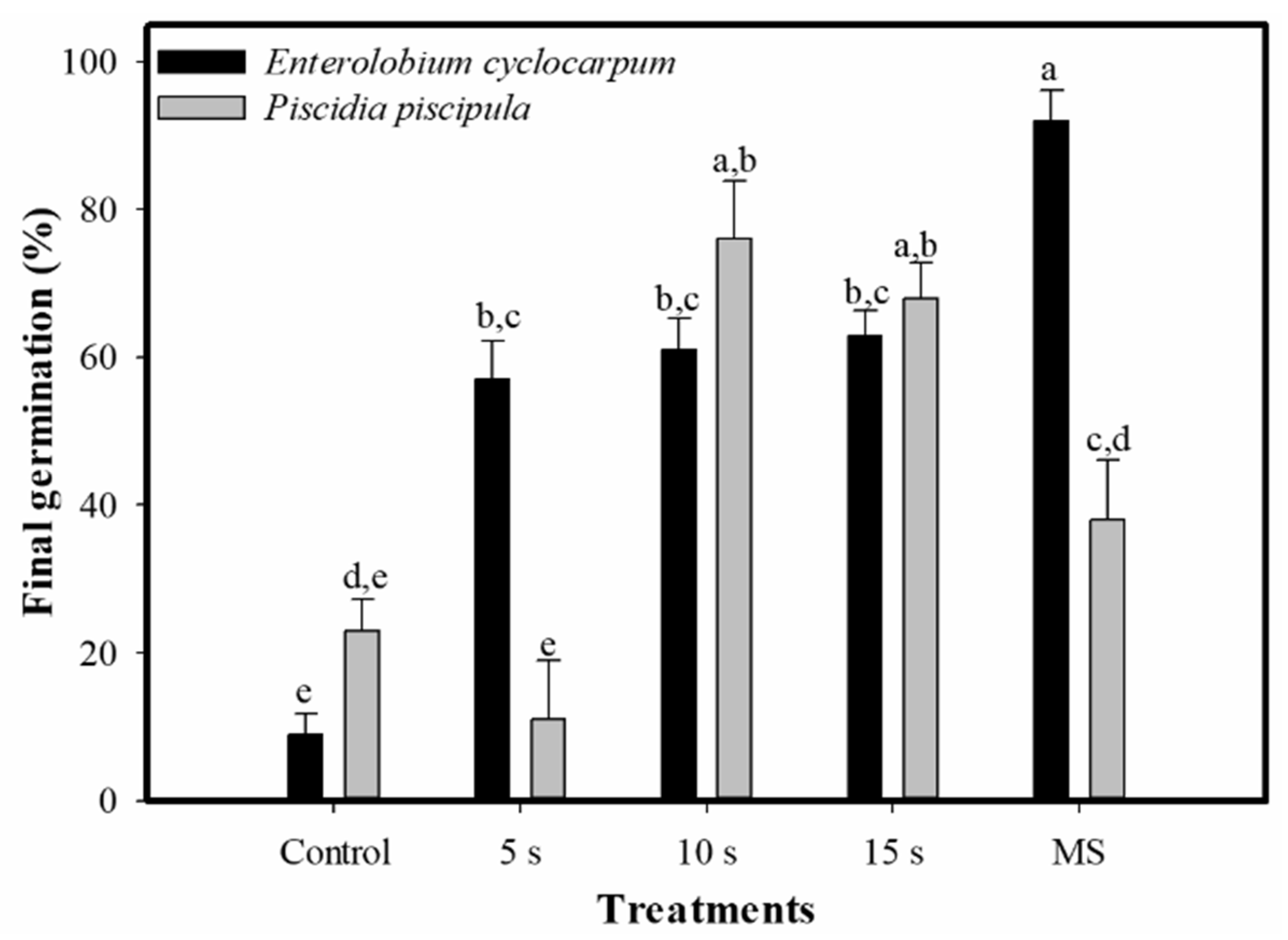
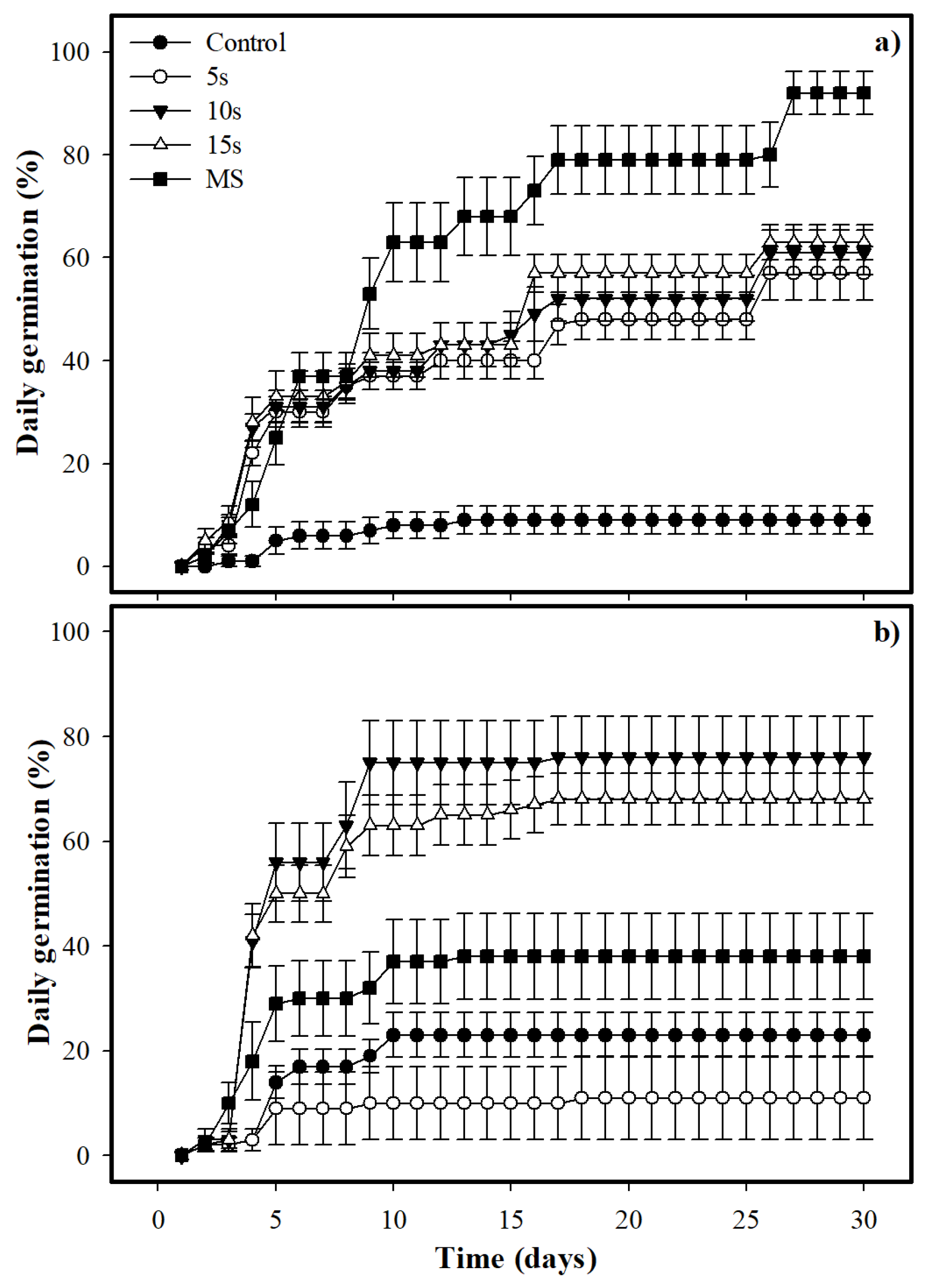
2.3. Seed Germination
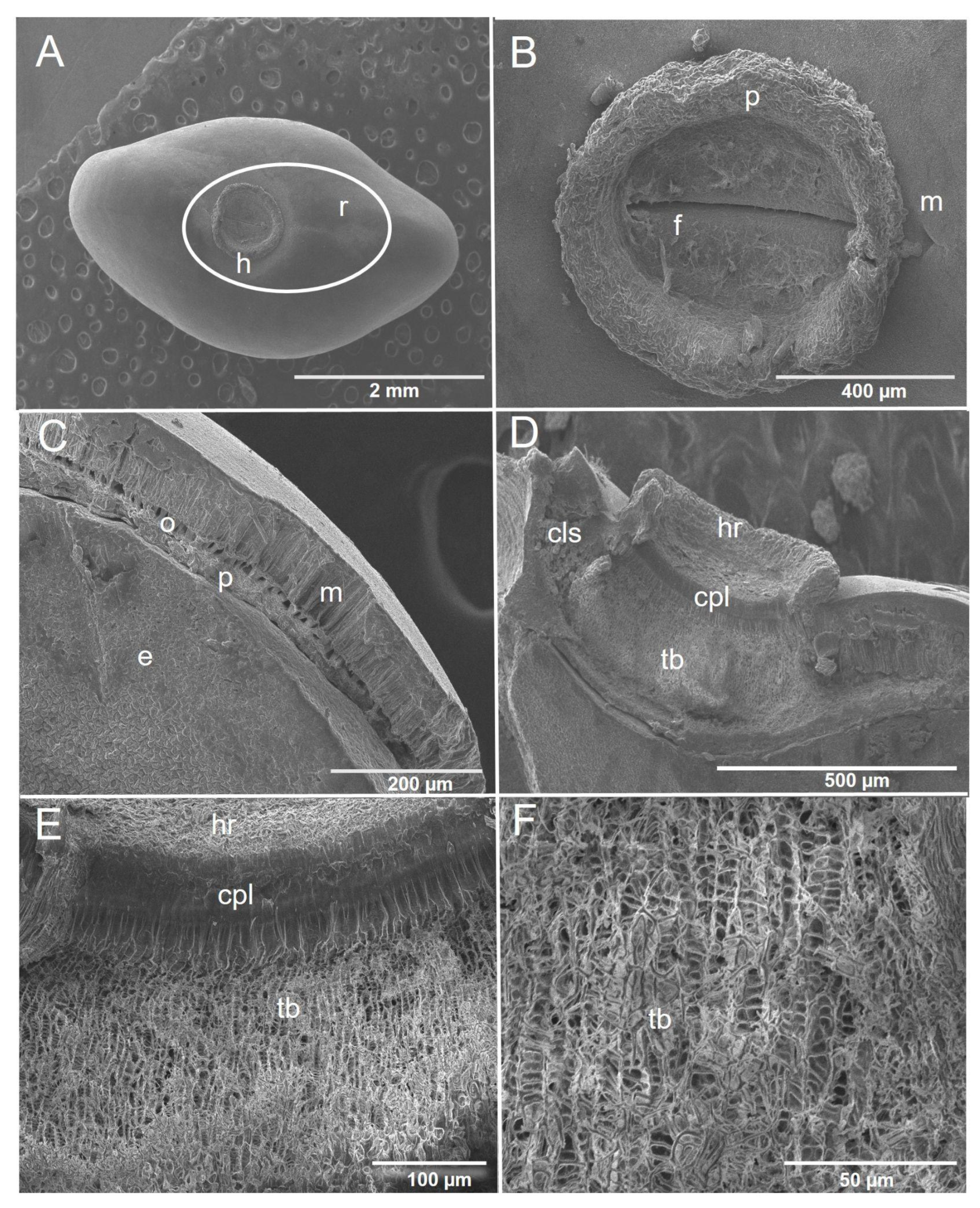
2.4. Morphological Characterization of the Seed Coat
2.4.1. Piscidia piscipula
2.4.2. Enterolobium cyclocarpum
3. Discussion
4. Materials and Methods
4.1. Study Species
4.2. Seed Collection
4.3. Seed Disinfection
4.4. Viability Test
4.5. Imbibition Rate
4.6. Effect of Pre-Germinative Treatments in Fresh Seeds
4.7. Morphological Characterization of the Seed Coat
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic Press: Cambridge, CA, USA, 2014; ISBN 9780124166776. [Google Scholar]
- Robles, E.; Jurado, E.; Ruiz, M.; Yáñez, L.; Flores, J. Heat shock effect in breaking physical dormancy in seeds of Lupinus elegans and L. rotundiflorus from Jalisco, México. Bot. Sci. 2014, 92, 123–129. [Google Scholar]
- Gama, N.; Baskin, J.; Geneve, R.; Baskin, C. Identification and characterization of ten new water gaps in seeds and fruits with physical dormancy and classification of water-gap complexes. Ann. Bot. 2013, 112, 69–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baskin, J.M.; Baskin, C.C.; Li, X. Taxonomy, ecology, and evolution of physical dormancy in seeds. Plant Species Biol. 2000, 15, 139–152. [Google Scholar] [CrossRef]
- Perissé, P.; Planchuelo, A. Seed coat morphology of Lupinus albus L. and Lupinus angustifolius L: In relation to water uptake. Seed Sci. Technol. 2004, 32, 69–77. [Google Scholar] [CrossRef]
- Robles, E.; Flores, J.; Yáñez, L. Paths of water entry and structures involved in the breaking of seed dormancy of Lupinus. J. Plant Physiol. 2016, 192, 75–80. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. A geographical perspective on germination ecology: Tropical and subtropical zones. VII. Hot semi deserts and deserts. In Seeds: Ecology, Biogeography and Evolution of Dormancy and Germination; Baskin, C., Baskin, J.M., Eds.; Academic Press: Cambridge, CA, USA, 2001; pp. 293–329. [Google Scholar]
- Koornneef, M.; Bentsink, L.; Hilhorst, H. Seed dormancy and germination. Current Opinion. Plant Biol. 2002, 5, 33–36. [Google Scholar]
- Mejías, M.; Molist, P.; Pombal, M. Órganos Vegetales: Semillas. Atlas de Histología Vegetal y Animal. Departamento de Biología Funcional y Ciencias de la Salud, Facultad de Biología, Universidad de Vigo: Vigo, Spain, 2018; p. 9. [Google Scholar]
- Moreno, F.G.; Plaza, P.; Magnitskiy, S. Efecto de la testa sobre la germinación de semillas de caucho (Hevea brasiliensis Muell.). Agron. Colomb. 2006, 24, 290–295. [Google Scholar]
- Omar, R.; Albornoz, P.L. Morpho-anatomy, imbibition, viability, and germination of the seed of Anadenanthera colubrina var. cebil (Fabaceae). Rev. Biol. Trop. 2013, 61, 1109–1118. [Google Scholar]
- Sánchez, J.; Pernús, M.; Torres-Arias, Y.; Barrios, D.; Dupuig, Y. Dormancia y germinación en semillas de árboles y arbustos de Cuba: Implicaciones para la restauración ecológica. Act. Bot. Cub. 2019, 218, 77–108. [Google Scholar]
- Langer, R.; Hill, G. Fabaceae. In Agricultural Plants; Cambridge University Press: Cambridge, UK; Lincoln University: Canterbury, New Zealand, 1991; pp. 217–282. ISBN 9781139170284. [Google Scholar]
- Judd, W.S.; Campbell, C.S.; Kellogg, E.A.; Stevens, P.F.; Donoghue, M.J. Plant Systematics: A Phylogenetic Approach, 4th ed.; Sinauer Associates: Sunderland, MA, USA, 2016; ISBN 978-1-60535-389-0. [Google Scholar]
- Ahmad, F.; Anwar, F.; Hira, S. Review on medicinal importance of Fabaceae family. Pahrmacology OnLine 2016, 3, 151–156. [Google Scholar]
- Villaseñor, J.L. Checklist of the native vascular plants of Mexico. Rev. Mex. Biodivers. 2016, 87, 559–902. [Google Scholar] [CrossRef] [Green Version]
- Fernández, G.C.; Tapia, J.L.; Duno de Stefano, R.; Ramírez, I.M. Flora Ilustrada de la Península de Yucatán: Listado Florístico; Centro de Investigación Científica de Yucatán: Mérida, México, 2010; ISBN 9686077823070. [Google Scholar]
- Fernández, G.C.; Tapia, J.L.; Duno de Stefano, R.; Ramírez, I.M.; Can, L.; Hernández, S.; Castillo, A. La Flora de la Península de Yucatán Mexicana: 250 años de conocimiento florístico. CONABIO. Biodiversitas 2012, 101, 6–10. [Google Scholar]
- Viveros, H.; Hernández, J.D.; Velasco, M.V.; Robles, R.; Ruiz, C.; Aparicio, A.; Martínez, M.J.; Hernández, J.; Hernández, M.L. Análisis de semilla, tratamientos pregerminativos de Enterolobium cyclocarpum (Jacq.) Griseb. y su crecimiento inicial. Rev. Mex. Cienc. For. 2015, 6, 52–65. [Google Scholar]
- González-Valdivia, N.A.; Dzib, B.B.; Carballo, J.I. Emergencia y crecimiento de plántulas de Piscidia piscipula (L.) Sarg. en condiciones de vivero. Acta Univ. 2020, 30, 1–9. [Google Scholar]
- Burrows, G.E.; Alden, R.; Robinson, W.A. Markedly different patterns of imbibition in seeds of 48 Acacia species. Seed Sci. Res. 2019, 29, 270–282. [Google Scholar] [CrossRef] [Green Version]
- Galíndez, G.; Malagrina, G.; Ceccato, D.; Ledesma, T.; Lindow, L.; Ortega, P. Dormición física y conservación ex situ de semillas de Amburana cearensis y Myroxylon peruiferum (Fabaceae). Bol. Soc. Argent. Bot. 2015, 50, 153–161. [Google Scholar] [CrossRef]
- Baskin, C.C.; Wurts, J.; Liu, Z.; Baskin, J.M. A method for breaking physical dormancy in seeds of the endemic Tibetan Plateau shrub Sophora moorcroftiana var. moorcroftiana (Fabaceae) and implications for restoration. Nat. Areas J. 2007, 27, 118–123. [Google Scholar] [CrossRef]
- Rodríguez, A.G.; Faria, J.M.; Vaz, T.A.; Nakamura, A.T.; José, A.C. Physical dormancy in Senna multijuga (Fabaceae: Caesalpinioideae) seeds: The role of seed structures in water uptake. Seed Sci. Res. 2014, 24, 147–157. [Google Scholar] [CrossRef]
- Duarte, M.M.; Paula, S.R.P.D.; Ferreira, F.R.D.L.; Nogueira, A.C. Morphological characterization of fruit, seed and seedling and germination of Hymenaea courbaril L. (Fabaceae) (‘Jatobá’). J. Seed Sci. 2016, 38, 204–211. [Google Scholar] [CrossRef]
- Jaganathan, G.K.; Wu, G.R.; Han, Y.Y.; Liu, B.L. Role of the lens in controlling physical dormancy break and germination of Delonix regia (Fabaceae: Caesalpinioideae). Plant Biol. 2017, 19, 53–60. [Google Scholar] [CrossRef]
- Ferreras, A.E.; Marcora, P.I.; Venier, M.P.; Funes, G. Different strategies for breaking physical seed dormancy in field conditions in two fruit morphs of Vachellia caven (Fabaceae). Seed Sci. Res. 2018, 28, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Carruggio, F.; Onofri, A.; Impelluso, C.; Giusso del Galdo, G.; Scopece, G.; Cristaudo, A. Seed dormancy breaking and germination in Bituminaria basaltica and B. bituminosa (Fabaceae). Plants 2020, 9, 1110. [Google Scholar] [CrossRef] [PubMed]
- Soltani, E.; Baskin, J.M.; Baskin, C.C.; Benakashani, F. A meta-analysis of the effects of treatments used to break dormancy in seeds of the megagenus Astragalus (Fabaceae). Seed Sci. Res. 2020, 30, 224–233. [Google Scholar] [CrossRef]
- Tang, L.; Baskin, C.; Baskin, J.; Luo, K.; Yu, X.; Huang, W.; Chen, Y. Methods of breaking physical dormancy in seeds of the invasive weed Mimosa pudica (Fabaceae) and a comparison with 36 other species in the genus. PeerJ 2022, 10, e13567. [Google Scholar] [CrossRef]
- Ezenwa, I. Preliminary evaluation of the suitability of Enterolobium cyclocarpum for use in intensive feed gardens in southwestern Nigeria. Agrofor. Syst. 1988, 44, 13–19. [Google Scholar] [CrossRef]
- Hernández, G.; Sánchez, L.R.; Aragón, F. Tratamientos pregerminativos en cuatro especies arbóreas de uso forrajero de la selva baja caducifolia de la sierra de Manantlán. For. Veracruzana 2001, 3, 9–15. [Google Scholar]
- Patiño, V.F.; Villagómez, A.Y. Los Análisis de Semillas y su Utilización en la Propagación de Especies Forestales. Boletín 40; Instituto Nacional de Investigaciones Forestales: Distrito Federal, México, 1976; p. 26. [Google Scholar]
- Vázquez, C.; Yvanosky, J. Leguminosas (Fabaceae): Características, Hábitat, Cultivo, Usos. Available online: https://www.lifeder.com/leguminosas-fabaceae/ (accessed on 30 May 2020). Tallinn Estonia.
- Ávila, L.E.; Herrera, M.A. Effect of extractives on three physical properties in Enterolobium cyclocarpum from Michoacan, Mexico. Bosque 2012, 33, 227–232. [Google Scholar]
- Hillis, W.E. Chemical features. In Heartwood and Tree Exudates; Springer: Berlin, Germany, 1987; pp. 76–119. ISBN 978-3-642-72536-4. [Google Scholar]
- Fengel, D.; Wegener, G. Wood: Chemistry, Ultrastructure, Reactions; Walter de Gruyter: Berlin, Germany, 2011; pp. 81–82. [Google Scholar]
- Ankli, A.; Heinrich, M.; Bork, P.; Wolfram, L.; Bauerfeind, P.; Brun, R.; Sticher, O. Yucatec Mayan medicinal plants: Evaluation based on indigenous uses. J. Ethnopharmacol. 2002, 79, 43–52. [Google Scholar] [CrossRef]
- Acevedo, P. The occurrence of piscicides and stupefactants in the plant kingdom. New Directions in the Study of Plants and People: Research Contributions from the Institute of Economic Botany. Adv. Econ. Bot. 1990, 8, 1–23. [Google Scholar]
- Cáceres, A.; López, B.R.; Giron, M.A.; Logemann, H. Plants used in Guatemala for the treatment of dermatophytic infections. Screening for antimycotic activity of 44 plant extracts. J. Ethnopharmacol. 1991, 31, 263–276. [Google Scholar] [CrossRef]
- Hernández-Epigmenio, S.; Rodríguez, D.A.; Granados, D.; Cadena, J.A. Latencia física, morfoanatomía y análisis proximal de la semilla de Enterolobium cyclocarpum (Jacq.) Griseb. Entreciencias 2021, 9, 1–15. [Google Scholar] [CrossRef]
- Lersten, N.R. Tracheid bar and vestured pits in legume seeds (Leguminosae: Papilionoideae). Am. J. Bot. 1982, 69, 98–107. [Google Scholar] [CrossRef]
- Souza, F.H.; Marcos, J. The seed coat as a modulator of seed-environment relationships in Fabaceae. Braz. J. Biol. 2001, 24, 365–375. [Google Scholar] [CrossRef] [Green Version]
- Maiti, R.K.; Hernández, J.L.; Valdez, M. Seed ultrastructure and germination of some species of Cactaceae. Phyton 1994, 55, 97–105. [Google Scholar]
- Morpeth, D.R.; Hall, A.M. Microbial enhancement of seed germination in Rosa corymbifera ‘Laxa’. Seed Sci. Res. 2000, 10, 489–494. [Google Scholar] [CrossRef]
- Janzen, D.H. Enterolobium cyclocarpum seed passage rate and survival in horses, Costa Rican Pleistocene seed dispersal agents. Ecology 1981, 62, 593–601. [Google Scholar] [CrossRef]
- Álvarez, C.; Barradas, L.; Ponce, O.; Williams, G. Soil seed bank, seed removal, and germination in a seasonally dry tropical forest in Veracruz, Mexico. Bot. Sci. 2014, 92, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Pennington, T.D.; Sarukhán, J. Árboles Tropicales de México. Manual Para la Identificación de las Principales Especies; UNAM-Fondo de Cultura Económica: Ciudad de México, México, 2005; p. 523. ISSN 9789703216437. [Google Scholar]
- Flores, J.S.; Espejel, I. Tipos de Vegetación de la Península de Yucatán; Etnoflora Yucatanense, Fascículo 3; Universidad Autónoma de Yucatán: Mérida, México, 1994; p. 135. [Google Scholar]
- Aguilera, R.M. Enterolobium cyclocarpum (Jacq.) Griseb. CONAFOR: 2011. Available online: http://www.conafor.gob.mx:8080/documentos/docs/13/920Enterolobium%20cyclocarpum.pdf (accessed on 19 February 2022). Ciudad de México, México.
- SDS. Jabín. Secretaría de Desarrollo Sustentable; Gobierno de Yucatán: Mérida, México, 2019; Available online: https://www.yucatan.gob.mx/?p=jabin (accessed 22 June 2020).
- Centro de Investigación Científica de Yucatán CICY. Flora de la Península de Yucatán. Piscidia; Centro de Investigación Científica de Yucatán CICY: Mérida, México, 2010; Available online: https://www.cicy.mx/sitios/flora%20digital/ficha_virtual.php?especie=1653 (accessed on 22 June 2020).
- Kanan-Kab. El Jabín: Piscidia piscipula; Kanan Kab AC: Mérida, México, 2017; Available online: https://kanankab.wordpress.com/2017/04/09/el-jabin-piscidia-piscipula/ (accessed on 27 June 2020).
- INEGI. Cuentame…información por identidad. Campeche. Flora y Fauna. INEGI n.d. Available online: https://cuentame.inegi.org.mx/monografias/informacion/camp/default.aspx?tema=me&e=04 (accessed on 27 June 2022).
- Zamora, P.; Barrientos, R.C.; Plasencia, A.H.; Villegas, P.; Domínguez, M.R.; Gutiérrez, C.; Aguirre, F.J.; Vargas, J.A.; Uc, S.; Poot, I.G. Estructura y composición florística de la selva mediana subcaducifolia en Hobomó, Campeche, Campeche, México. Madera y Bosques 2020, 26, e2611899. [Google Scholar]
- EcuRed. San Francisco de Campeche, México. Ecured n.d. Available online: https://www.ecured.cu/San_Francisco_de_Campeche_(México) (accessed on 29 August 2022).
- Comisión Nacional del Agua (CONAGUA). Base de Datos Mensuales Climatológicos Correspondientes a la Estación de Campeche; Dirección Local Campeche, Residencia Técnica: Campeche, México, 2010. [Google Scholar]
- Climate-Data. Clima de San Francisco de Campeche. Climate-Data n.d. Available online: https://es.climate-data.org/america-del-norte/mexico/campeche/san-francisco-de-campeche-3373/ (accessed on 29 August 2022).
- Aragón, J.L.; Flores, J.; Jurado, E.; Ramírez, H.M.; Robles, E.; Rodas, J.P.; Yáñez, L. Potential impact of global warming on seed bank, dormancy, and germination of three succulent species from the Chihuahuan Desert. Seed Sci. Res. 2018, 28, 312–318. [Google Scholar] [CrossRef]
- Euan, J.L.; González, E.A.; Ramírez, J.E.; Zamora, P.; Vargas, J.A.; Rodríguez, U.; Enríquez, J.K.; Caamal, J.H.; Aragón, J.L. Storage time effect on the germination of Haematoxylum campechianum in Campeche, Mexico. Madera Bosques 2001, 27, e2732263. [Google Scholar] [CrossRef]
- ISTA. International Rules for Seed Testing; ISTA: Shenzhen, China, 2014. [Google Scholar]
- Copeland, L.O.; McDonald, M.B. Seed Viability and Viability Testing. Principles of Seed Science and Technology; Springer: London, UK, 2001; pp. 124–139. [Google Scholar]
- López, L.; Navarro, D.; Martínez, V.; Toorop, P.E.; Iannetta, P.P. A spectrophotometric assay for robust viability testing of seed batches using 2, 3, 5-triphenyl tetrazolium chloride: Using Hordeum vulgare L. as a model. Front. Plant Sci. 2017, 8, 747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- TeKrony, D.M.; Egli, D.B.; Phillips, A.D. Effect of field weathering on the viability and vigor of soybean seed. Agronomy J. 1980, 72, 749–753. [Google Scholar] [CrossRef]
- Baskin, C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic Press: New York, NY, USA, 1998. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arceo-Gómez, T.M.; Robles-Díaz, E.; Manrique-Ortega, M.D.; Martínez-Campos, Á.R.; Aragón-Gastélum, J.L.; Aguirre-Crespo, F.J.; Ramírez-Albores, J.E.; Pérez-Suárez, M.; Robles, R.; Reyes-Trujeque, J.; et al. Pre-Germinative Treatments and Morphophysiological Traits in Enterolobium cyclocarpum and Piscidia piscipula (Fabaceae) from the Yucatan Peninsula, Mexico. Plants 2022, 11, 2844. https://doi.org/10.3390/plants11212844
Arceo-Gómez TM, Robles-Díaz E, Manrique-Ortega MD, Martínez-Campos ÁR, Aragón-Gastélum JL, Aguirre-Crespo FJ, Ramírez-Albores JE, Pérez-Suárez M, Robles R, Reyes-Trujeque J, et al. Pre-Germinative Treatments and Morphophysiological Traits in Enterolobium cyclocarpum and Piscidia piscipula (Fabaceae) from the Yucatan Peninsula, Mexico. Plants. 2022; 11(21):2844. https://doi.org/10.3390/plants11212844
Chicago/Turabian StyleArceo-Gómez, Thomas Martín, Erika Robles-Díaz, Mayra D. Manrique-Ortega, Ángel Roberto Martínez-Campos, José Luis Aragón-Gastélum, Francisco Javier Aguirre-Crespo, Jorge E. Ramírez-Albores, Marlín Pérez-Suárez, Rafael Robles, Javier Reyes-Trujeque, and et al. 2022. "Pre-Germinative Treatments and Morphophysiological Traits in Enterolobium cyclocarpum and Piscidia piscipula (Fabaceae) from the Yucatan Peninsula, Mexico" Plants 11, no. 21: 2844. https://doi.org/10.3390/plants11212844