
Impacts of the Invasive Alien Carpobrotus spp. on Coastal Habitats on a Mediterranean Island (Giglio Island, Central Italy)
 , , ,
, , ,
Abstract
:1. Introduction
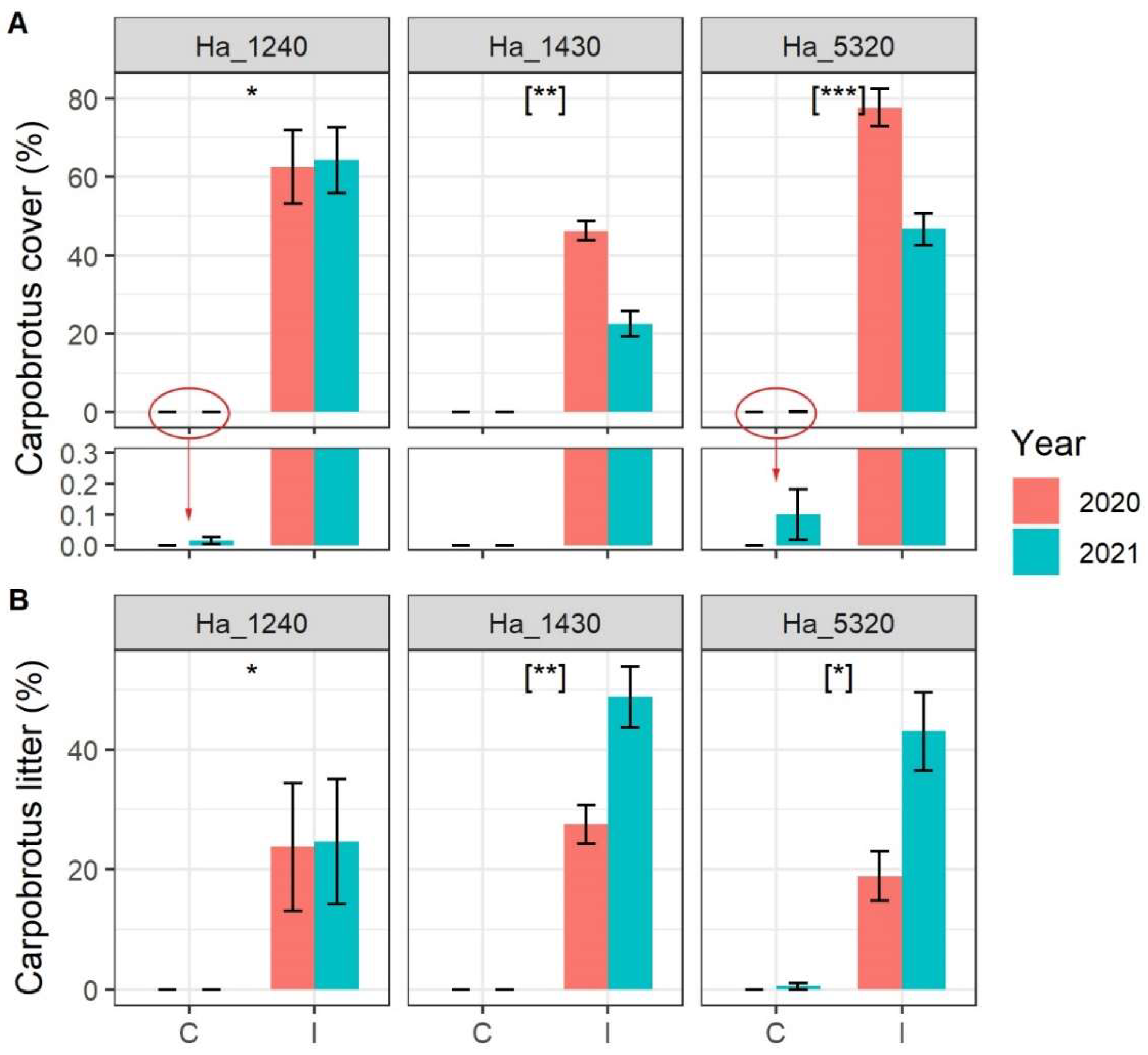
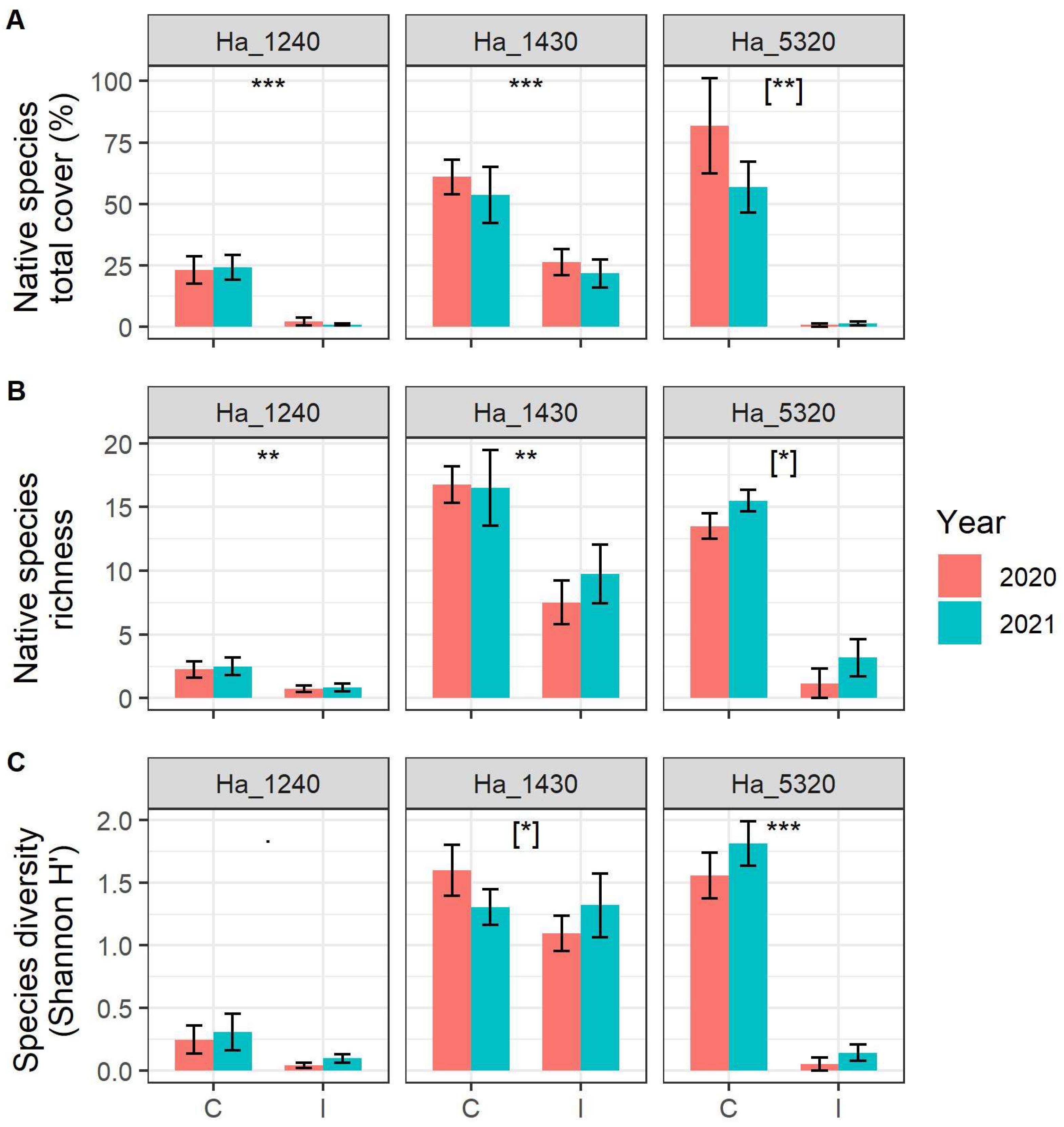
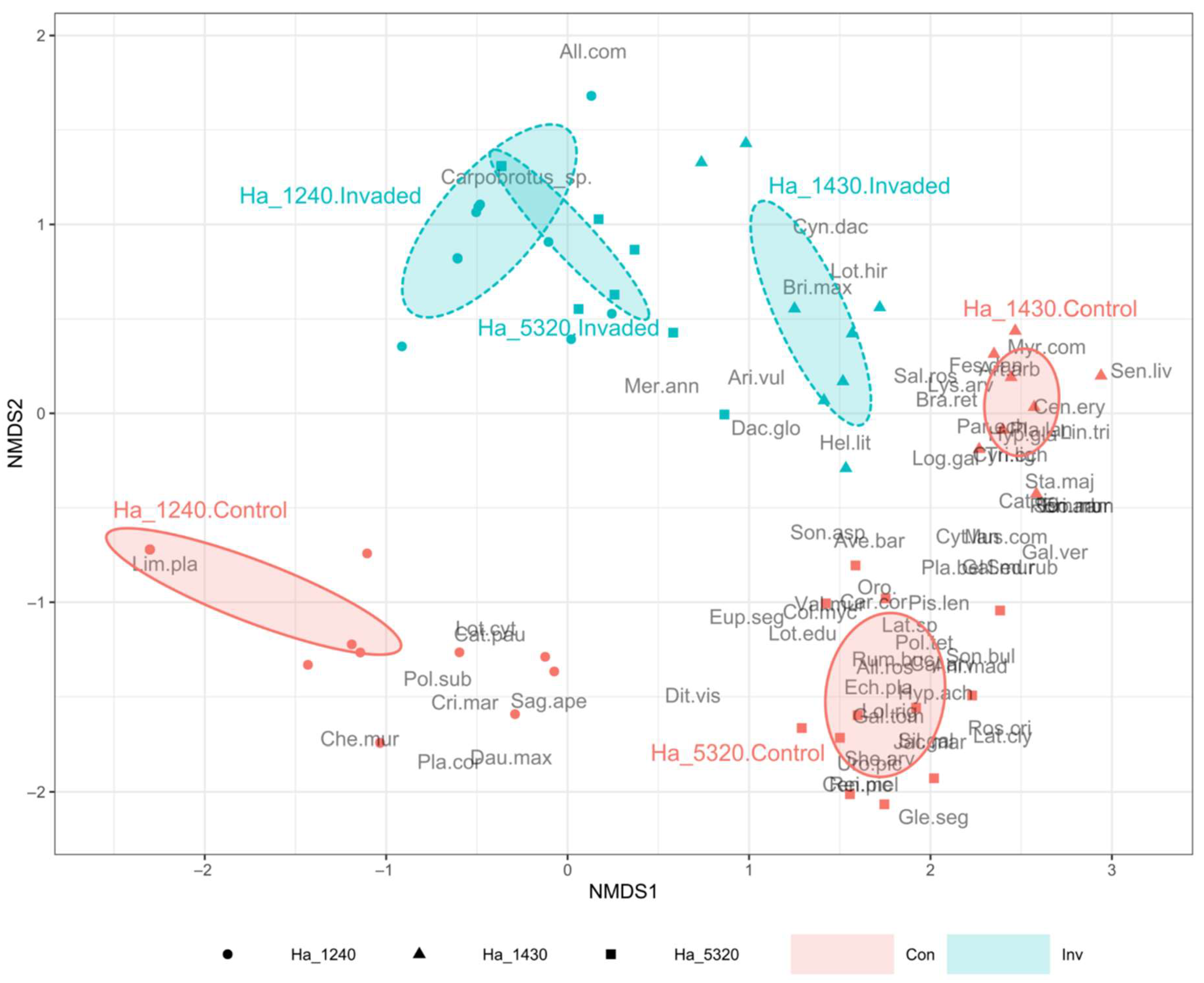
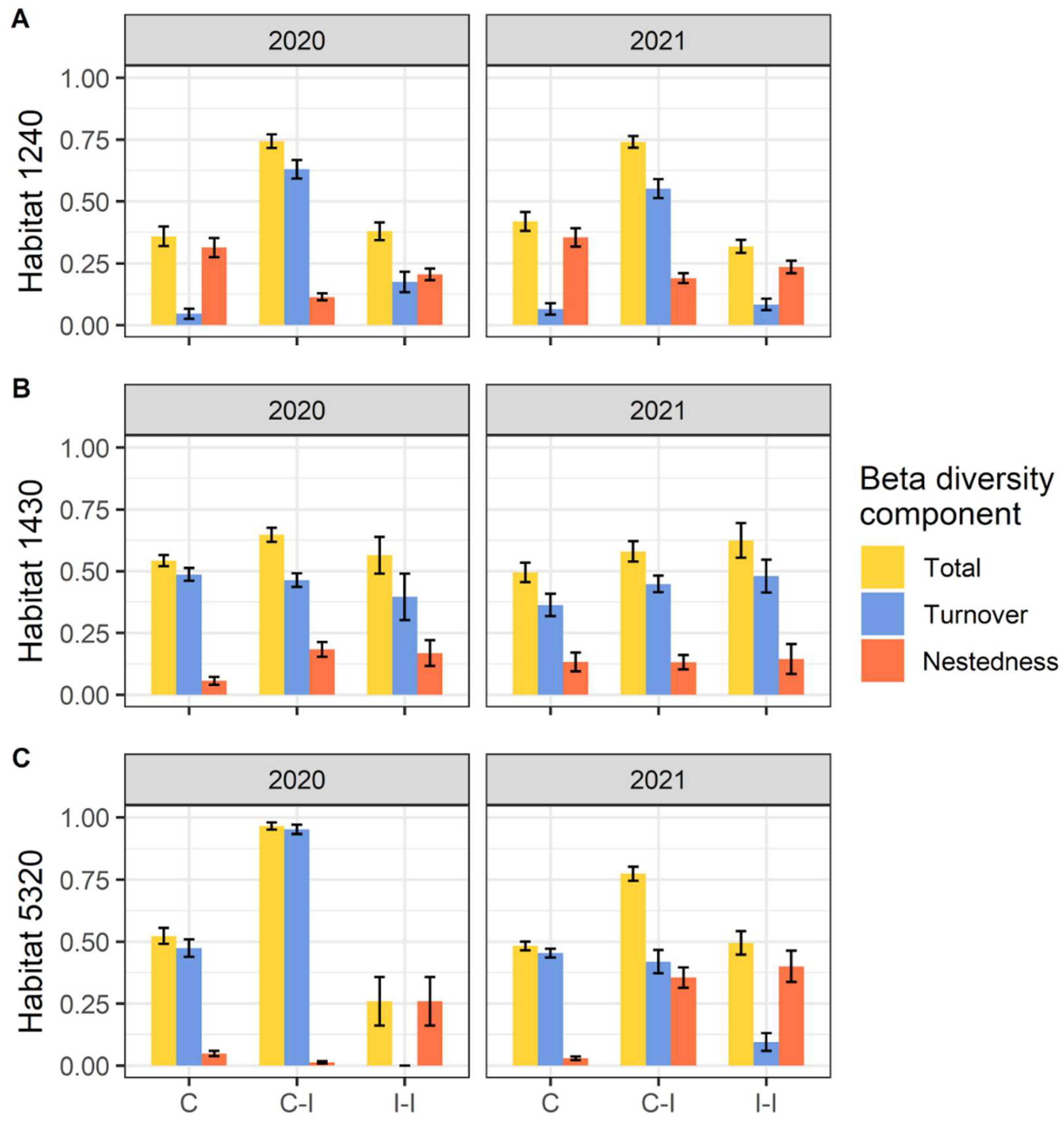
2. Results
3. Discussion
4. Materials and Methods
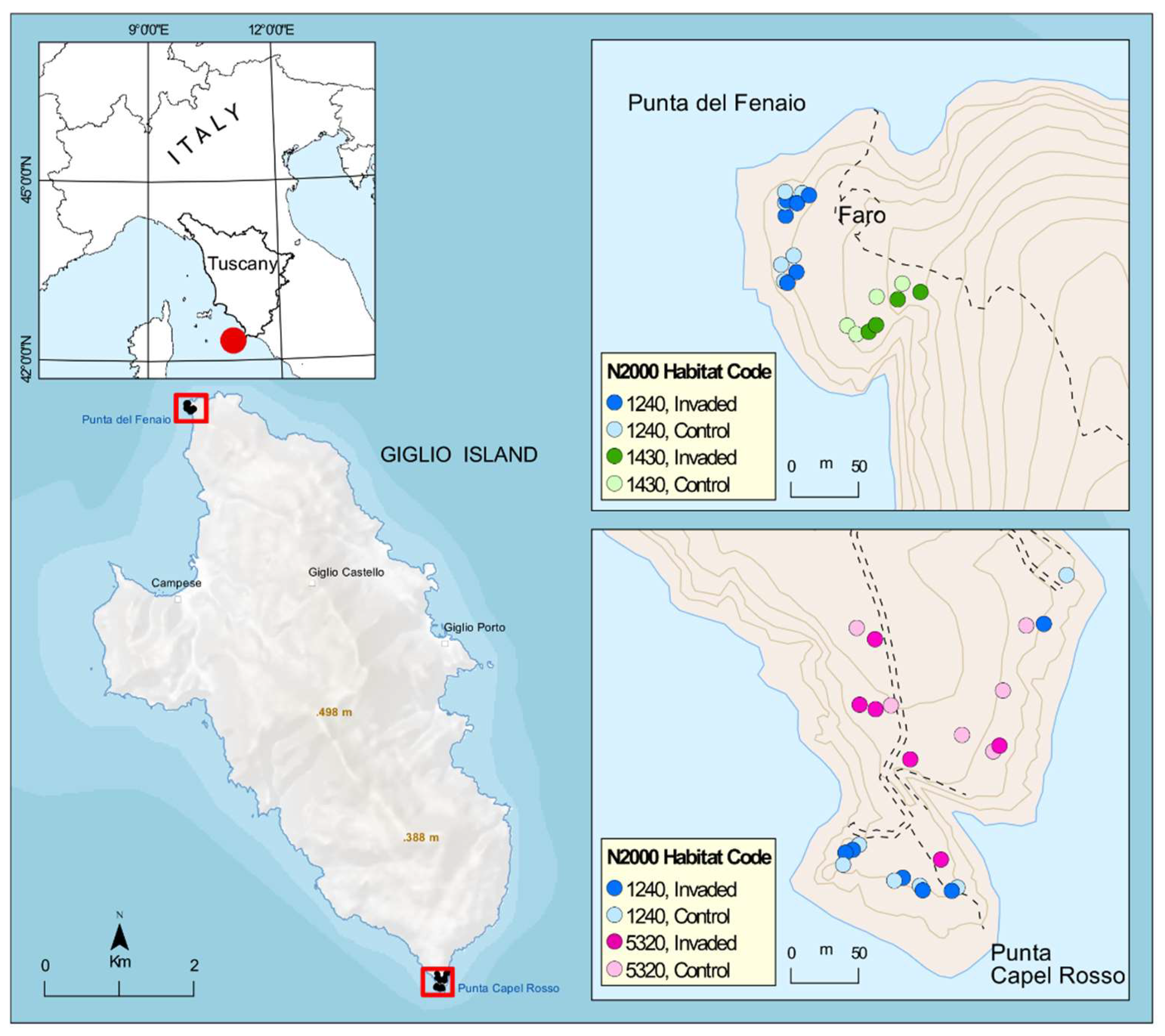
4.1. Study Area
4.2. Data Collection
4.3. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Díaz, S.M.; Settele, J.; Brondízio, E.; Ngo, H.; Guèze, M.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.; Butchart, S.; et al. The Global Assessment Report on Biodiversity and Ecosystem Services: Summary for Policy Makers; Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services secretariat: Bonn, Germany, 2019; ISBN 978-3-947851-13-3. [Google Scholar]
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ Warning on Invasive Alien Species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef] [PubMed]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M.; et al. No Saturation in the Accumulation of Alien Species Worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef] [PubMed]
- Hejda, M.; Pyšek, P.; Jarošík, V. Impact of Invasive Plants on the Species Richness, Diversity and Composition of Invaded Communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological Impacts of Invasive Alien Plants: A Meta-Analysis of Their Effects on Species, Communities and Ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef]
- Vilà, M.; Rohr, R.P.; Espinar, J.L.; Hulme, P.E.; Pergl, J.; Le Roux, J.J.; Schaffner, U.; Pyšek, P. Explaining the Variation in Impacts of Non-Native Plants on Local-Scale Species Richness: The Role of Phylogenetic Relatedness. Glob. Ecol. Biogeogr. 2015, 24, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Vilà, M.; Hulme, P.E. Impact of Biological Invasions on Ecosystem Services; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Kumar Rai, P.; Singh, J.S. Invasive Alien Plant Species: Their Impact on Environment, Ecosystem Services and Human Health. Ecol. Indic. 2020, 111, 106020. [Google Scholar] [CrossRef]
- Olden, J.D.; Rooney, T.P. On Defining and Quantifying Biotic Homogenization. Glob. Ecol. Biogeogr. 2006, 15, 113–120. [Google Scholar] [CrossRef]
- Dawson, W.; Moser, D.; van Kleunen, M.; Kreft, H.; Pergl, J.; Pyšek, P.; Weigelt, P.; Winter, M.; Lenzner, B.; Blackburn, T.M.; et al. Global Hotspots and Correlates of Alien Species Richness across Taxonomic Groups. Nat. Ecol. Evol. 2017, 1, 0186. [Google Scholar] [CrossRef] [Green Version]
- Gaertner, M.; Den Breeyen, A.; Hui, C.; Richardson, D.M. Impacts of Alien Plant Invasions on Species Richness in Mediterranean-Type Ecosystems: A Meta-Analysis. Prog. Phys. Geogr. Earth Environ. 2009, 33, 319–338. [Google Scholar] [CrossRef]
- Brundu, G. Invasive Alien Plants in Protected Areas in Mediterranean Islands: Knowledge Gaps and Main Threats. In Plant Invasions in Protected Areas: Patterns, Problems and Challenges; Foxcroft, L.C., Pyšek, P., Richardson, D.M., Genovesi, P., Eds.; Invading Nature—Springer Series in Invasion Ecology; Springer: Dordrecht, The Netherlands, 2013; pp. 395–422. ISBN 978-94-007-7750-7. [Google Scholar]
- Celesti-Grapow, L.; Bassi, L.; Brundu, G.; Camarda, I.; Carli, E.; D’Auria, G.; Del Guacchio, E.; Domina, G.; Ferretti, G.; Foggi, B.; et al. Plant Invasions on Small Mediterranean Islands: An Overview. Plant Biosyst.-Int. J. Deal. Asp. Plant Biol. 2016, 150, 1119–1133. [Google Scholar] [CrossRef]
- Lazzaro, L.; Foggi, B.; Ferretti, G.; Brundu, G. Priority Invasive Alien Plants in the Tuscan Archipelago (Italy): Comparing the EPPO Prioritization Scheme with the Australian WRA. Biol. Invasions 2016, 18, 1317–1333. [Google Scholar] [CrossRef]
- Lazzaro, L.; Bolpagni, R.; Buffa, G.; Gentili, R.; Lonati, M.; Stinca, A.; Acosta, A.T.R.; Adorni, M.; Aleffi, M.; Allegrezza, M.; et al. Impact of Invasive Alien Plants on Native Plant Communities and Natura 2000 Habitats: State of the Art, Gap Analysis and Perspectives in Italy. J. Environ. Manag. 2020, 274, 111140. [Google Scholar] [CrossRef] [PubMed]
- Maes, J. A Model for the Assessment of Habitat Conservation Status in the EU; Publication Office of the European Commission JRC scientific and policy report; European Commission Joint Research Centre Institute for Environment and Sustainability: Luxembourg, 2013. [Google Scholar]
- Pyšek, P.; Jarošík, V.; Hulme, P.E.; Pergl, J.; Hejda, M.; Schaffner, U.; Vilà, M. A Global Assessment of Invasive Plant Impacts on Resident Species, Communities and Ecosystems: The Interaction of Impact Measures, Invading Species’ Traits and Environment. Glob. Chang. Biol. 2012, 18, 1725–1737. [Google Scholar] [CrossRef]
- Guerra, C.; Baquero, R.A.; Gutiérrez-Arellano, D.; Nicola, G.G. Is the Natura 2000 Network Effective to Prevent the Biological Invasions? Glob. Ecol. Conserv. 2018, 16, e00497. [Google Scholar] [CrossRef]
- Mazaris, A.D.; Katsanevakis, S. The Threat of Biological Invasions Is Under-Represented in the Marine Protected Areas of the European Natura 2000 Network. Biol. Conserv. 2018, 225, 208–212. [Google Scholar] [CrossRef]
- Campoy, J.G.; Acosta, A.T.R.; Affre, L.; Barreiro, R.; Brundu, G.; Buisson, E.; González, L.; Lema, M.; Novoa, A.; Retuerto, R.; et al. Monographs of Invasive Plants in Europe: Carpobrotus. Bot. Lett. 2018, 165, 440–475. [Google Scholar] [CrossRef]
- Carranza, M.L.; Carboni, M.; Feola, S.; Acosta, A.T.R. Landscape-Scale Patterns of Alien Plant Species on Coastal Dunes: The Case of Iceplant in Central Italy. Appl. Veg. Sci. 2010, 13, 135–145. [Google Scholar] [CrossRef]
- Molinari, N.; D’Antonio, C.; Thomson, G.; Cuddington, K.; Byers, J.; Hastings, A.; Wilson, W. Carpobrotus as a Case Study of the Complexities of Species Impacts. In Ecosystem Engineers: Plants to Protists; Therotical ecology series; Elsevier: London, UK, 2007; pp. 139–162. [Google Scholar]
- Santoro, R.; Jucker, T.; Carboni, M.; Acosta, A.T.R. Patterns of Plant Community Assembly in Invaded and Non-Invaded Communities along a Natural Environmental Gradient. J. Veg. Sci. 2012, 23, 483–494. [Google Scholar] [CrossRef]
- Vilà, M.; Tessier, M.; Suehs, C.M.; Brundu, G.; Carta, L.; Galanidis, A.; Lambdon, P.; Manca, M.; Médail, F.; Moragues, E.; et al. Local and Regional Assessments of the Impacts of Plant Invaders on Vegetation Structure and Soil Properties of Mediterranean Islands. J. Biogeogr. 2006, 33, 853–861. [Google Scholar] [CrossRef]
- Acosta, A.; Izzi, C.F.; Stanisci, A. Comparison of Native and Alien Plant Traits in Mediterranean Coastal Dunes. Community Ecol. 2006, 7, 35–41. [Google Scholar] [CrossRef]
- Santoro, R.; Carboni, M.; Carranza, M.L.; Acosta, A.T.R. Focal Species Diversity Patterns Can Provide Diagnostic Information on Plant Invasions. J. Nat. Conserv. 2012, 20, 85–91. [Google Scholar] [CrossRef]
- Jucker, T.; Carboni, M.; Acosta, A.T.R. Going beyond Taxonomic Diversity: Deconstructing Biodiversity Patterns Reveals the True Cost of Iceplant Invasion. Divers. Distrib. 2013, 19, 1566–1577. [Google Scholar] [CrossRef]
- Badalamenti, E.; Gristina, L.; Laudicina, V.A.; Novara, A.; Pasta, S.; La Mantia, T. The Impact of Carpobrotus Cfr. Acinaciformis (L.) L. Bolus on Soil Nutrients, Microbial Communities Structure and Native Plant Communities in Mediterranean Ecosystems. Plant Soil 2016, 409, 19–34. [Google Scholar] [CrossRef]
- Santoro, R.; Jucker, T.; Carranza, M.; Acosta, A. Assessing the Effects of Carpobrotus Invasion on Coastal Dune Soils. Does the Nature of the Invaded Habitat Matter? Community Ecol. 2011, 12, 234–240. [Google Scholar] [CrossRef]
- Novoa, A.; González, L.; Moravcová, L.; Pyšek, P. Constraints to Native Plant Species Establishment in Coastal Dune Communities Invaded by Carpobrotus Edulis: Implications for Restoration. Biol. Conserv. 2013, 164, 1–9. [Google Scholar] [CrossRef]
- Vieites-Blanco, C.; González-Prieto, S.J. Effects of Carpobrotus Edulis Invasion on Soil Gross N Fluxes in Rocky Coastal Habitats. Sci. Total Environ. 2018, 619–620, 966–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novoa, A.; González, L. Impacts of Carpobrotus Edulis (L.) N.E.Br. on the Germination, Establishment and Survival of Native Plants: A Clue for Assessing Its Competitive Strength. PLoS ONE 2014, 9, e107557. [Google Scholar] [CrossRef] [Green Version]
- Novoa, A.; González, L.; Moravcová, L.; Pyšek, P. Effects of Soil Characteristics, Allelopathy and Frugivory on Establishment of the Invasive Plant Carpobrotus Edulis and a Co-Occuring Native, Malcolmia Littorea. PLoS ONE 2012, 7, e53166. [Google Scholar] [CrossRef] [Green Version]
- Brunel, S.; Brundu, G.; Fried, G. Eradication and control of invasive alien plants in the Mediterranean Basin: Towards better coordination to enhance existing initiatives. EPPO Bull. 2013, 43, 290–308. [Google Scholar] [CrossRef]
- Lazzaro, L.; Ferretti, G.; Giuliani, C.; Foggi, B. A Checklist of the Alien Flora of the Tuscan Archipelago (Italy). Webbia 2014, 69, 157–176. [Google Scholar] [CrossRef]
- Hulme, P.E.; Pyšek, P.; Jarošík, V.; Pergl, J.; Schaffner, U.; Vilà, M. Bias and Error in Understanding Plant Invasion Impacts. Trends Ecol. Evol. 2013, 28, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Baselga, A. Partitioning the Turnover and Nestedness Components of Beta Diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Chase, J.M.; Myers, J.A. Disentangling the Importance of Ecological Niches from Stochastic Processes across Scales. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2351–2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leprieur, F.; Olden, J.D.; Lek, S.; Brosse, S. Contrasting Patterns and Mechanisms of Spatial Turnover for Native and Exotic Freshwater Fish in Europe. J. Biogeogr. 2009, 36, 1899–1912. [Google Scholar] [CrossRef]
- Marini, L.; Bertolli, A.; Bona, E.; Federici, G.; Martini, F.; Prosser, F.; Bommarco, R. Beta-Diversity Patterns Elucidate Mechanisms of Alien Plant Invasion in Mountains. Glob. Ecol. Biogeogr. 2013, 22, 450–460. [Google Scholar] [CrossRef]
- Fried, G.; Laitung, B.; Pierre, C.; Chagué, N.; Panetta, F.D. Impact of Invasive Plants in Mediterranean Habitats: Disentangling the Effects of Characteristics of Invaders and Recipient Communities. Biol. Invasions 2014, 16, 1639–1658. [Google Scholar] [CrossRef]
- Lazzaro, L.; Tondini, E.; Lombardi, L.; Giunti, M. The Eradication of Carpobrotus Spp. in the Sand-Dune Ecosystem at Sterpaia (Italy, Tuscany): Indications from a Successful Experience. Biologia 2020, 75, 199–208. [Google Scholar] [CrossRef]
- Buisson, E.; Braschi, J.; Chenot-Lescure, J.; Hess, M.C.M.; Vidaller, C.; Pavon, D.; Ramone, H.; Amy-Krebs, E.; Cottaz, C.; Passetti, A. Native Plant Community Recovery after Carpobrotus (Ice Plant) Removal on an Island—Results of a 10-Year Project. Appl. Veg. Sci. 2021, 24, e12524. [Google Scholar] [CrossRef]
- Foggi, B.; Pancioli, V. Contribution to the Knowledge of the Vegetation of the Giglio Island (Tuscan Archipelago, Southern Tuscany). Webbia 2008, 63, 25–48. [Google Scholar] [CrossRef]
- Conser, C.; Connor, E.F. Assessing the Residual Effects of Carpobrotus Edulis Invasion, Implications for Restoration. Biol. Invasions 2009, 11, 349–358. [Google Scholar] [CrossRef]
- Malavasi, M.; Santoro, R.; Cutini, M.; Acosta, A.T.R.; Carranza, M.L. The Impact of Human Pressure on Landscape Patterns and Plant Species Richness in Mediterranean Coastal Dunes. Plant Biosyst.-Int. J. Deal. Asp. Plant Biol. 2016, 150, 73–82. [Google Scholar] [CrossRef]
- Spyreas, G.; Wilm, B.W.; Plocher, A.E.; Ketzner, D.M.; Matthews, J.W.; Ellis, J.L.; Heske, E.J. Biological consequences of invasion by reed canary grass (Phalaris arundinacea). Biol. Invasions 2010, 12, 1253–1267. [Google Scholar] [CrossRef]
- Benesperi, R.; Giuliani, C.; Zanetti, S.; Gennai, M.; Mariotti Lippi, M.; Guidi, T.; Nascimbene, J.; Foggi, B. Forest plant diversity is threatened by Robinia pseudoacacia (black-locust) invasion. Biodivers. Conserv. 2012, 21, 3555–3568. [Google Scholar] [CrossRef]
- Chen, G.Q.; Zhang, C.B.; Ling, M.A.; Qiang, S.; Silander, J.A.; Qi, L.L. Biotic homogenization caused by the invasion of Solidago canadensis in China. J. Integr. Agric. 2013, 12, 835–845. [Google Scholar] [CrossRef]
- Chen, H.; Qian, H.; Spyreas, G.; Crossland, M. Native-exotic species richness relationships across spatial scales and biotic homogenization in wetland plant communities of Illinois, USA. Divers. Distrib. 2010, 16, 737–743. [Google Scholar] [CrossRef]
- Nascimbene, J.; Lazzaro, L.; Benesperi, R. Patterns of β-diversity and similarity reveal biotic homogenization of epiphytic lichen communities associated with the spread of black locust forests. Fungal Ecol. 2015, 14, 1–7. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Jagodziński, A.M. Impacts of Invasive Trees on Alpha and Beta Diversity of Temperate Forest Understories. Biol. Invasions 2021, 23, 235–252. [Google Scholar] [CrossRef]
- Marzialetti, F.; Cascone, S.; Frate, L.; Di Febbraro, M.; Acosta, A.T.R.; Carranza, M.L. Measuring Alpha and Beta Diversity by Field and Remote-Sensing Data: A Challenge for Coastal Dunes Biodiversity Monitoring. Remote Sens. 2021, 13, 1928. [Google Scholar] [CrossRef]
- Baldini, R.M. The Vascular Flora of the Giglio Island. (Tuscan Archipelago): Taxonomic Revision and Updating. Webbia 1998, 52, 307–404. [Google Scholar] [CrossRef]
- Foggi, B.; Viciani, D.; Baldini, R.M.; Carta, A.; Guidi, T. Conservation Assessment of the Endemic Plants of the Tuscan Archipelago, Italy. Oryx 2015, 49, 118–126. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species Assemblages and Indicator Species:The Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D. Mixed-Effects Models in S and S-PLUS; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2006; p. 528. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package, 2020.
- Baselga, A.; Orme, D.; Villeger, S.; De Bortoli, J.; Leprieur, F.; Baselga, M.A. Package ‘betapart’. Partitioning beta diversity into turnover and nestedness components, version, 1.5.6. 2022.
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response Variable | Habitat | Variable | numDF | denDF | F Value | p Value | |
|---|---|---|---|---|---|---|---|
| Native species total cover | 1240 £ | Invasion Status | 1 | 44 | 155.35 | <0.001 | *** |
| Year | 1 | 44 | 0.25 | 0.616 | |||
| Invasion Status:Year | 1 | 44 | 0.54 | 0.466 | |||
| 1430 £ | Invasion Status | 1 | 12 | 19.69 | <0.001 | *** | |
| Year | 1 | 12 | 4.46 | 0.056 | ˙ | ||
| Invasion Status:Year | 1 | 12 | 0.15 | 0.705 | |||
| 5320 £ | Invasion Status | 1 | 20 | 101.44 | <0.001 | *** | |
| Year | 1 | 20 | 0.13 | 0.720 | |||
| Invasion Status:Year | 1 | 20 | 9.21 | 0.007 | ** | ||
| Native species richness | 1240 £ | Invasion Status | 1 | 44 | 8.32 | 0.006 | ** |
| Year | 1 | 44 | 0.45 | 0.504 | |||
| Invasion Status:Year | 1 | 44 | 0.11 | 0.743 | |||
| 1430 | Invasion Status | 1 | 12 | 18.24 | 0.001 | ** | |
| Year | 1 | 12 | 0.51 | 0.487 | |||
| Invasion Status:Year | 1 | 12 | 0.80 | 0.388 | |||
| 5320 £ | Invasion Status | 1 | 20 | 35.09 | <0.001 | *** | |
| Year | 1 | 20 | 15.75 | <0.001 | *** | ||
| Invasion Status:Year | 1 | 20 | 8.12 | 0.010 | * | ||
| Species diversity (H’) | 1240 £ | Invasion Status | 1 | 44 | 3.93 | 0.054 | ˙ |
| Year | 1 | 44 | 4.80 | 0.034 | * | ||
| Invasion Status:Year | 1 | 44 | 0.29 | 0.595 | |||
| 1430 | Invasion Status | 1 | 12 | 3.46 | 0.088 | ˙ | |
| Year | 1 | 12 | 0.15 | 0.703 | |||
| Invasion Status:Year | 1 | 12 | 9.29 | 0.010 | * | ||
| 5320 | Invasion Status | 1 | 20 | 93.61 | <0.001 | *** | |
| Year | 1 | 20 | 3.23 | 0.087 | ˙ | ||
| Invasion Status:Year | 1 | 20 | 0.74 | 0.400 |
| Species | Invasion Status | Habitat | Indicator Value | p Value | |
|---|---|---|---|---|---|
| Limonium planesiae Pignatti | Control plots | 1240 | 0.95 | <0.001 | *** |
| Brachypodium retusum (Pers.) P.Beauv. | 1430 | 0.70 | <0.001 | *** | |
| Artemisia arborescens (Vaill.) L. | 0.69 | <0.001 | *** | ||
| Lysimachia arvensis (L.) U.Manns & Anderb. | 0.64 | <0.001 | *** | ||
| Helichrysum litoreum Guss. | 0.52 | <0.001 | *** | ||
| Paronychia echinulata Chater | 0.50 | <0.001 | *** | ||
| Linum trigynum L. | 0.38 | 0.001 | ** | ||
| Festuca danthonii Asch. & Graebn. subsp. danthonii | 0.34 | 0.001 | ** | ||
| Lotus hirsutus L. | 0.34 | 0.003 | ** | ||
| Hypochaeris glabra L. | 0.30 | 0.007 | ** | ||
| Arisarum vulgare O.Targ.Tozz. subsp. vulgare | 0.29 | 0.015 | * | ||
| Myrtus communis L. | 0.25 | 0.014 | * | ||
| Plantago lanceolata L. | 0.25 | 0.015 | * | ||
| Stachys major (L.) Bartolucci & Peruzzi | 0.25 | 0.015 | * | ||
| Sedum rubens L. | 0.23 | 0.029 | * | ||
| Muscari comosum (L.) Mill. | 0.22 | 0.038 | * | ||
| Cytisus laniger DC. | 0.20 | 0.026 | * | ||
| Allium roseum L. subsp. roseum | 5320 | 0.89 | <0.001 | *** | |
| Euphorbia segetalis L. | 0.80 | <0.001 | *** | ||
| Lotus edulis L. | 0.75 | <0.001 | *** | ||
| Polycarpon tetraphyllum (L.) L. | 0.69 | <0.001 | *** | ||
| Hypochaeris achyrophorus L. | 0.57 | <0.001 | *** | ||
| Carlina corymbosa L. | 0.57 | <0.001 | *** | ||
| Glebionis segetum (L.) Fourr. | 0.50 | <0.001 | *** | ||
| Urospermum picroides (L.) Scop. ex F.W.Schmidt | 0.50 | <0.001 | *** | ||
| Rumex bucephalophorus L. | 0.49 | <0.001 | *** | ||
| Coleostephus myconis (L.) Cass. ex Rchb.f. | 0.48 | <0.001 | *** | ||
| Valantia muralis L. | 0.42 | <0.001 | *** | ||
| Lolium rigidum Gaudin | 0.40 | <0.001 | *** | ||
| Jacobaea maritima (L.) Pelser & Meijden subsp. maritima | 0.39 | <0.001 | *** | ||
| Silene gallica L. | 0.33 | 0.002 | ** | ||
| Echium plantagineum L. | 0.32 | 0.006 | ** | ||
| Avena barbata Pott ex Link | 0.31 | 0.022 | * | ||
| Pistacia lentiscus L. | 0.28 | 0.019 | * | ||
| Anisantha madritensis (L.) Nevski subsp. madritensis | 0.27 | 0.018 | * | ||
| Rostraria cristata (L.) Tzvelev | 0.25 | 0.024 | * | ||
| Plantago bellardii All. | 0.23 | 0.028 | * | ||
| Calendula arvensis (Vaill.) L. | 0.17 | 0.049 | * | ||
| Dactylis glomerata L. | Invaded plots | 1430 | 0.59 | <0.001 | *** |
| Briza maxima L. | 0.34 | 0.005 | ** | ||
| Cynodon dactylon (L.) Pers. | 0.26 | 0.008 | ** | ||
| Carpobrotus spp. | 5320 | 0.39 | <0.001 | *** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mugnai, M.; Benesperi, R.; Viciani, D.; Ferretti, G.; Giunti, M.; Giannini, F.; Lazzaro, L. Impacts of the Invasive Alien Carpobrotus spp. on Coastal Habitats on a Mediterranean Island (Giglio Island, Central Italy). Plants 2022, 11, 2802. https://doi.org/10.3390/plants11202802
Mugnai M, Benesperi R, Viciani D, Ferretti G, Giunti M, Giannini F, Lazzaro L. Impacts of the Invasive Alien Carpobrotus spp. on Coastal Habitats on a Mediterranean Island (Giglio Island, Central Italy). Plants. 2022; 11(20):2802. https://doi.org/10.3390/plants11202802
Chicago/Turabian StyleMugnai, Michele, Renato Benesperi, Daniele Viciani, Giulio Ferretti, Michele Giunti, Francesca Giannini, and Lorenzo Lazzaro. 2022. "Impacts of the Invasive Alien Carpobrotus spp. on Coastal Habitats on a Mediterranean Island (Giglio Island, Central Italy)" Plants 11, no. 20: 2802. https://doi.org/10.3390/plants11202802