
Cyclodextrins Increase Triterpene Production in Solanum lycopersicum Cell Cultures by Activating Biosynthetic Genes


Abstract
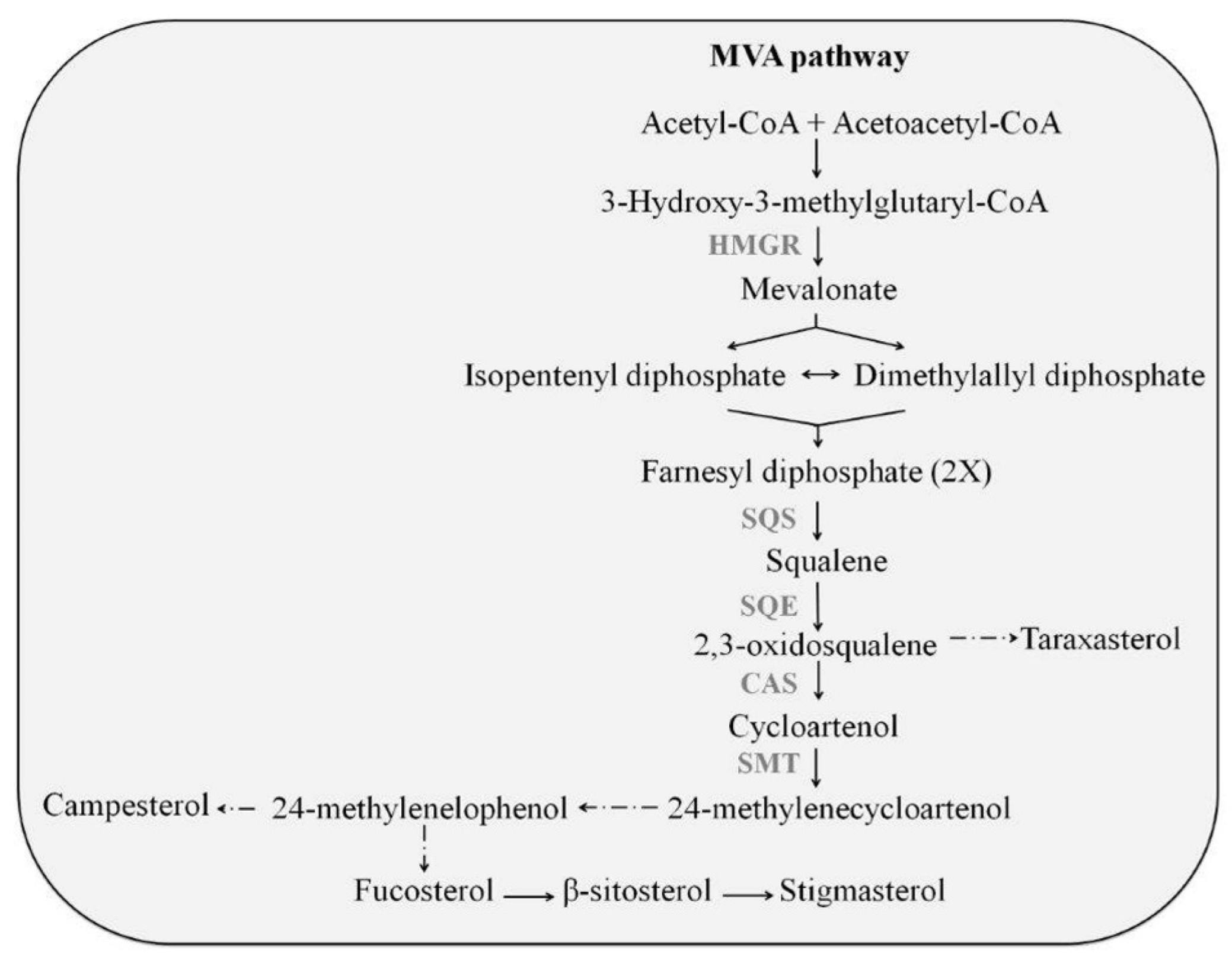
:1. Introduction
2. Results and Discussion
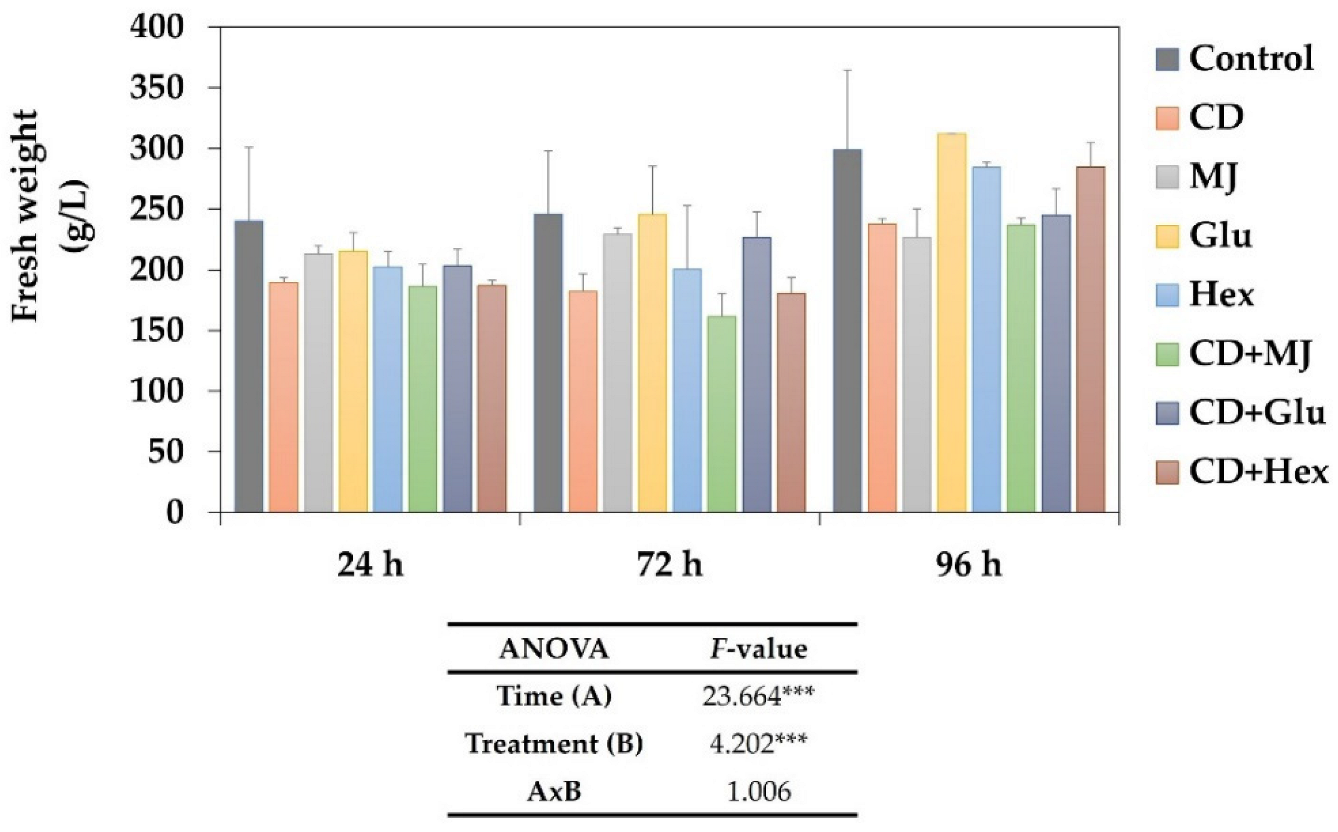
2.1. Effect of Elicitors on the Growth of MT Tomato SCC
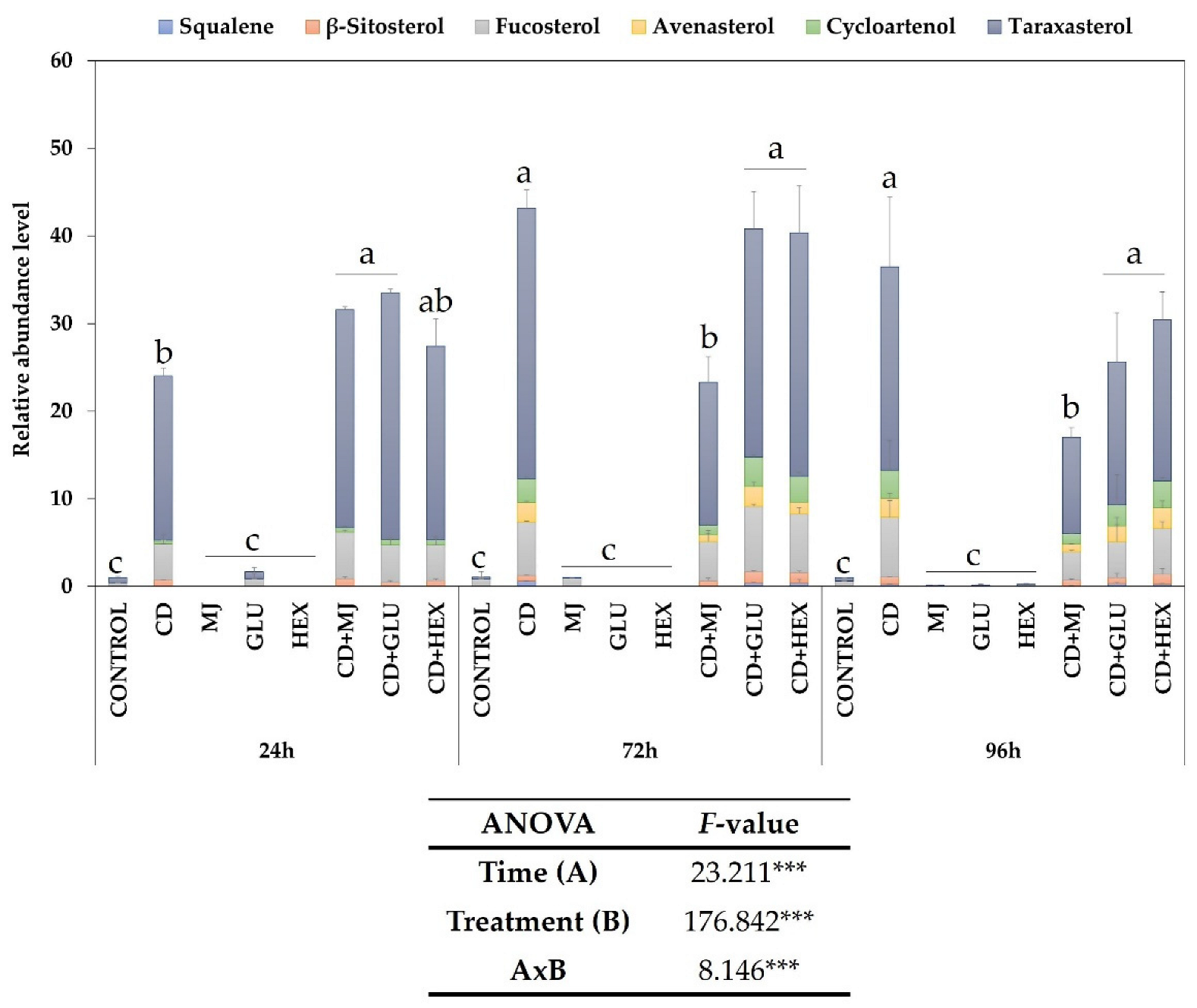
2.2. Effect of Elicitors on Total Triterpene Content in MT Tomato SCC
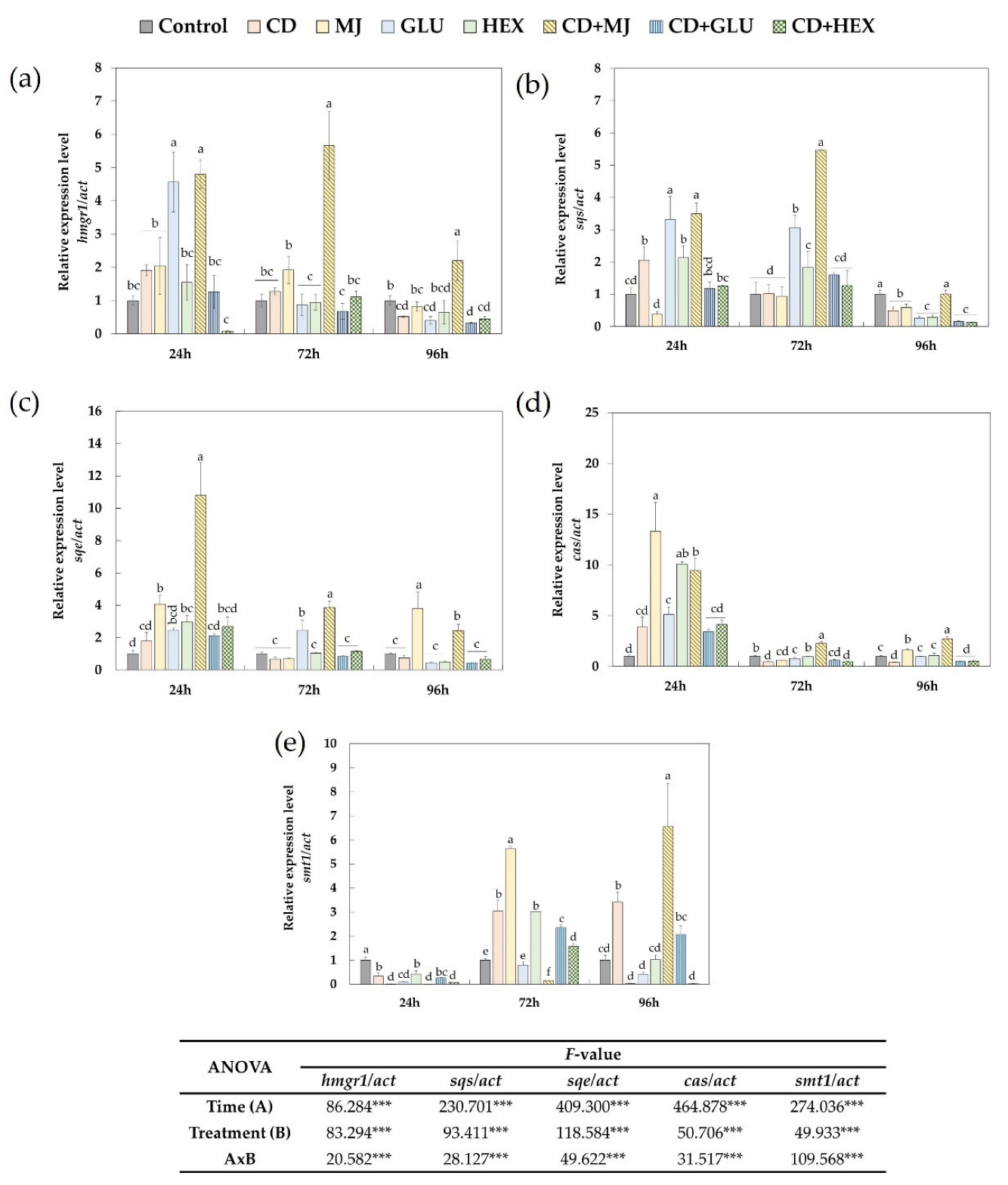
2.3. Effect of Elicitors on the Expression of Triterpene Biosynthetic Genes
3. Materials and Methods
3.1. Plant Materials
3.2. Elicitation of MT Tomato SCC
3.3. Extraction and Identification of Triterpene Compounds
3.4. Quantitative Real-Time RT-PCR (qRT-PCR)
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aoki, K.; Yano, K.; Suzuki, A.; Kawamura, S.; Sakurai, N.; Suda, K.; Kurabayashi, A.; Suzuki, T.; Tsugane, T.; Watanabe, M.; et al. Large-scale analysis of full-length cDNAs from the tomato (Solanum lycopersicum) cultivar Micro-Tom.; a reference system for the Solanaceae genomics. BMC Genom. 2010, 11, 210. [Google Scholar] [CrossRef] [Green Version]
- Mannucci, A.; Mariotti, L.; Castagna, A.; Santin, M.; Trivellini, A.; Reyes, T.H.; Mensuali- Sodi, A.; Ranieri, A.M.; Quartacci, M.F. Hormone profile changes occur in roots and leaves of Micro-Tom tomato plants when exposing the aerial part to low doses of UV-B radiation. Plant Physiol. Biochem. 2020, 148, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Barreto, R.F.; de Mello Prado, R.; Lúcio, J.C.B.; López-Díaz, I.; Carrera, E.; Carvalho, R.F. Ammonium toxicity alleviation by silicon is dependent on cytokinins in tomato cv. Micro-tom. J. Plant Growth Regul. 2022, 41, 417–428. [Google Scholar] [CrossRef]
- Kosma, D.K.; Parsons, E.P.; Isaacson, T.; Lü, S.; Rose, J.K.; Jenks, M.A. Fruit cuticle lipid composition during development in tomato ripening mutants. Physiol. Plant. 2010, 139, 107–117. [Google Scholar] [CrossRef]
- Wang, Z.; Guhling, O.; Yao, R.; Li, F.; Yeats, T.H.; Rose, J.K.; Jetter, R. Two oxidosqualene cyclases responsible for biosynthesis of tomato fruit cuticular triterpenoids. Plant Physiol. 2011, 155, 540–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szakiel, A.; Pączkowski, C.; Pensec, F.; Bertsch, C. Fruit cuticular waxes as a source of biologically active triterpenoids. Phytochem. Rev. 2012, 11, 263–284. [Google Scholar] [CrossRef] [Green Version]
- Haliński, Ł.P.; Kalkowska, M.; Kalkowski, M.; Piorunowska, J.; Topolewska, A.; Stepnowski, P. Cuticular wax variation in the tomato (Solanum lycopersicum L.), related wild species and their interspecific hybrids. Biochem. Syst. Ecol. 2015, 60, 215–224. [Google Scholar] [CrossRef]
- Romero, P.; Rose, J.K. A relationship between tomato fruit softening, cuticle properties and water availability. Food Chem. 2019, 295, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Sabater-Jara, A.B.; Almagro, L.; Belchí-Navarro, S.; Ferrer, M.Á.; Barceló, A.R.; Pedreño, M.Á. Induction of sesquiterpenes.; phytoesterols and extracellular pathogenesis-related proteins in elicited cell cultures of Capsicum annuum. J. Plant Physiol. 2010, 167, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Briceño, Z.; Almagro, L.; Sabater-Jara, A.B.; Calderón, A.A.; Pedreño, M.A.; Ferrer, M.A. Enhancement of phytosterols taraxasterol and induction of extracellular pathogenesis-related proteins in cell cultures of Solanum lycopersicum cv Micro-Tom elicited with cyclodextrins and methyl jasmonate. J. Plant Physiol. 2012, 169, 1050–1058. [Google Scholar] [CrossRef] [PubMed]
- Devarenne, T.P.; Ghosh, A.; Chappell, J. Regulation of squalene synthase a key enzyme of sterol biosynthesis in tobacco. Plant Physiol. 2002, 129, 1095–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posé, D.; Castanedo, I.; Borsani, O.; Nieto, B.; Rosado, A.; Taconnat, L.; Ferrer, A.; Dolan, L.; Valpuesta, V.; Botella, M.A. Identification of the Arabidopsis dry2/sqe1-5 mutant reveals a central role for sterols in drought tolerance and regulation of reactive oxygen species. Plant J. 2009, 59, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Griebel, T.; Zeier, J. A role for β-sitosterol to stigmasterol conversion in plant-pathogen interactions. Plant J. 2010, 63, 254–268. [Google Scholar] [CrossRef]
- Vranová, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef]
- Fu, J.; Liu, G.; Yang, M.; Wang, X.; Chen, X.; Chen, F.; Yang, Y. Isolation and functional analysis of squalene synthase gene in tea plant Camellia sinensis. Plant Physiol. Biochem. 2019, 142, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Thimmappa, R.; Geisler, K.; Louveau, T.; O’Maille, P.; Osbourn, A. Triterpene biosynthesis in plants. Annu. Rev. Plant Biol. 2014, 65, 225–257. [Google Scholar] [CrossRef]
- Han, J.Y.; Jo, H.J.; Kwon, E.K.; Choi, Y.E. Cloning and characterization of oxidosqualene cyclases involved in taraxasterol, taraxerol and bauerenol triterpene biosynthesis in Taraxacum coreanum. Plant Cell Physiol. 2019, 60, 1595–1603. [Google Scholar] [CrossRef] [PubMed]
- Gas-Pascual, E.; Berna, A.; Bach, T.J.; Schaller, H. Plant oxidosqualene metabolism: Cycloartenol synthase–dependent sterol biosynthesis in Nicotiana benthamiana. PLoS ONE 2014, 9, e109156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vriese, K.; Pollier, J.; Goossens, A.; Beeckman, T.; Vanneste, S. Dissecting cholesterol and phytosterol biosynthesis via mutants and inhibitors. J. Exp. Bot. 2021, 72, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Campestrini, L.H.; Melo, P.S.; Peres, L.E.; Calhelha, R.C.; Ferreira, I.C.; Alencar, S.M. A new variety of purple tomato as a rich source of bioactive carotenoids and its potential health benefits. Heliyon 2019, 5, e02831. [Google Scholar] [CrossRef] [PubMed]
- Nabi, N.; Singh, S.; Saffeullah, P. Responses of in vitro cell cultures to elicitation: Regulatory role of jasmonic acid and methyl jasmonate: A review. In Vitro Cell Dev. Biol. Plant. 2021, 57, 341–355. [Google Scholar] [CrossRef]
- Bru, R.; Sellés, S.; Casado-Vela, J.; Belchí-Navarro, S.; Pedreño, M.A. Modified cyclodextrins are chemically defined glucan inducers of defense responses in grapevine cell cultures. J. Agric. Food Chem. 2006, 54, 65–71. [Google Scholar] [CrossRef]
- Almagro, L.; García-Pérez, P.; Belchí-Navarro, S.; Sánchez-Pujante, P.J.; Pedreño, M.A. New strategies for the use of Linum usitatissimum cell factories for the production of bioactive compounds. Plant Physiol. Biochem. 2016, 99, 73–78. [Google Scholar] [CrossRef]
- Giri, C.C.; Zaheer, M. Chemical elicitors versus secondary metabolite production in vitro using plant cell tissue and organ cultures: Recent trends and a sky eye view appraisal. Plant Cell Tissue Organ Cult. 2016, 126, 1–18. [Google Scholar] [CrossRef]
- Demole, E.; Lederer, E.; Mercier, D. Isolation and determination of the structure of methyl jasmonate a constituent odor characteristic of jasmine oil. Helv. Chim. Acta 1962, 45, 675–685. [Google Scholar] [CrossRef]
- Bertini, L.; Palazzi, L.; Proietti, S.; Pollastri, S.; Arrigoni, G.; Polverino de Laureto, P.; Caruso, C. Proteomic analysis of MeJa-induced defense responses in rice against wounding. Int. J. Mol. Sci. 2019, 20, 2525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelberth, J.; Contreras, C.F.; Dalvi, C.; Li, T.; Engelberth, M. Early transcriptome analyses of Z-3-hexenol-treated Zea mays revealed distinct transcriptional networks and anti-herbivore defense potential of green leaf volatiles. PLoS ONE 2013, 8, e77465. [Google Scholar]
- Vidal-Limon, H.R.; Almagro, L.; Moyano, E.; Palazon, J.; Pedreño, M.A.; Cusido, R.M. Perfluorodecalins and hexenol as inducers of secondary metabolism in Taxus media and Vitis vinifera cell cultures. Front Plant Sci. 2018, 9, 335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okinaka, Y.; Mimori, K.; Takeo, K.; Kitamura, S.; Takeuchi, Y.; Yamaoka, N.; Yoshikawa, M. A structural model for the mechanisms of elicitor release from fungal cell walls by plant β-1.; 3-endoglucanase. Plant Physiol. 1995, 109, 839–845. [Google Scholar] [CrossRef] [Green Version]
- Cheong, J.J.; Hahn, M.G. A specific high-affinity binding site for the hepta-beta- glucoside elicitor exists in soybean membranes. Plant Cell. 1991, 3, 137–147. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Yamada, A.; Hong, N.; Ogawa, T.; Ishii, T.; Shibuya, N. Differences in the recognition of glucan elicitor signals between rice and soybean: β-glucan fragments from the rice blast disease fungus Pyricularia oryzae that elicit phytoalexin biosynthesis in suspension-cultured rice cells. Plant Cell. 2000, 12, 817–826. [Google Scholar] [PubMed]
- Almagro, L.; Carbonell-Bejerano, P.; Belchí-Navarro, S.; Bru, R.; Martínez-Zapater, J.M.; Lijavetzky, D.; Pedreño, M.A. Dissecting the transcriptional response to elicitors in Vitis vinifera cells. PLoS ONE 2014, 9, e109777. [Google Scholar]
- Patil, R.A.; Lenka, S.K.; Normanly, J.; Walker, E.L.; Roberts, S.C. Methyl jasmonate represses growth and affects cell cycle progression in cultured Taxus cells. Plant Cell Rep. 2014, 33, 1479–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiorucci, A.S. To grow or defend? More on the plant Cornelian dilemma. Plant Physiol. 2020, 183, 437–438. [Google Scholar] [CrossRef] [PubMed]
- Pauwels, L.; Morreel, K.; De Witte, E.; Lammertyn, F.; Van Montagu, M.; Boerjan, W.; Inze, D.; Goossens, A. Mapping methyl jasmonate-mediated transcriptional reprogramming of metabolism and cell cycle progression in cultured Arabidopsis cells. Proc. Natl. Acad. Sci. USA 2008, 105, 1380–1385. [Google Scholar] [CrossRef] [Green Version]
- Noctor, G.; Reichheld, J.P.; Foyer, C.H. ROS-related redox regulation and signaling in plants. Semin. Cell Dev. Biol. 2018, 80, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Belchí-Navarro, S.; Almagro, L.; Lijavetzky, D.; Bru, R.; Pedreño, M.A. Enhanced extracellular production of trans-resveratrol in Vitis vinifera suspension cultured cells by using cyclodextrins and methyljasmonate. Plant Cell Rep. 2012, 31, 81–89. [Google Scholar] [CrossRef]
- Almagro, L.; Pedreño, M.A. Use of cyclodextrins to improve the production of plant bioactive compounds. Phytochem. Rev. 2020, 19, 1061–1080. [Google Scholar] [CrossRef]
- Sabater-Jara, A.B.; Pedreño, M.A. Use of β-cyclodextrins to enhance phytosterol production in cell suspension cultures of carrot (Daucus carota L.). Plant Cell Tissue Organ Cult. 2013, 114, 249–258. [Google Scholar] [CrossRef]
- Sharma, K.; Zafar, R. Optimization of methyl jasmonate and β-cyclodextrin for enhanced production of taraxerol and taraxasterol in (Taraxacum officinale Weber) cultures. Plant Physiol. Biochem. 2016, 103, 24–30. [Google Scholar] [CrossRef]
- Burg, J.S.; Espenshade, P.J. Regulation of HMG-CoA reductase in mammals and yeast. Prog. Lipid Res. 2011, 50, 403–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miras-Moreno, B.; Almagro, L.; Pedreño, M.A.; Sabater-Jara, A.B. Effect of terbinafine on the biosynthetic pathway of isoprenoid compounds in carrot suspension cultured cells. Plant Cell Rep. 2018, 37, 1011–1019. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Morel, G.; Wetmore, R.H. Fern callus tissue culture. Am. J. Bot. 1951, 38, 141–143. [Google Scholar] [CrossRef]
- Miras-Moreno, B.; Almagro, L.; Pedreño, M.A.; Sabater-Jara, A.B. Enhanced accumulation of phytosterols and phenolic compounds in cyclodextrin-elicited cell suspension culture of Daucus carota. Plant Sci. 2016, 250, 154–164. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Abbreviation | Accession Number | Primer Pair (5′-Forward-3′/5′-Reverse-3′) | Size (pb) |
|---|---|---|---|
| hmgr1 | NM_001309881.1 | CTTCCACTCCCATTGTACCTAAC/GATCTTCTCACGCCACCTTAC | 93 |
| sqs | NM_001247787.2 | ACCCACCGATGTTAAAGTACC/CTGGTCCATGAGAACCTTGT | 108 |
| sqe | XM_026029244.1 | GCACATGCTCCTCTTACAGTAG/GAGGGAACATCAACCTTAGGG | 86 |
| cas | NM_001246855.2 | CGCTTTGTTGGTCCTATCACT/GAGGGTGTGGGTAGTAAAGG | 133 |
| smt1 | XM_004229602.3 | AAGTTCTCTCTGCTGTTGACAAA/ACTCTCCCCATCCGTATTCATAGAAG | 144 |
| act | FJ532351.1 | TCAGGCTGTGCTTTCCTTGT/CGACCAGCAAGATCCAAACG | 141 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabater-Jara, A.B.; Marín-Marín, M.J.; Almagro, L.; Pedreño, M.A. Cyclodextrins Increase Triterpene Production in Solanum lycopersicum Cell Cultures by Activating Biosynthetic Genes. Plants 2022, 11, 2782. https://doi.org/10.3390/plants11202782
Sabater-Jara AB, Marín-Marín MJ, Almagro L, Pedreño MA. Cyclodextrins Increase Triterpene Production in Solanum lycopersicum Cell Cultures by Activating Biosynthetic Genes. Plants. 2022; 11(20):2782. https://doi.org/10.3390/plants11202782
Chicago/Turabian StyleSabater-Jara, Ana Belén, María Jesús Marín-Marín, Lorena Almagro, and María Angeles Pedreño. 2022. "Cyclodextrins Increase Triterpene Production in Solanum lycopersicum Cell Cultures by Activating Biosynthetic Genes" Plants 11, no. 20: 2782. https://doi.org/10.3390/plants11202782