
Joint Cultivation of Allium ursinum and Armoracia rusticana under Foliar Sodium Selenate Supply

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Morphological Characteristics
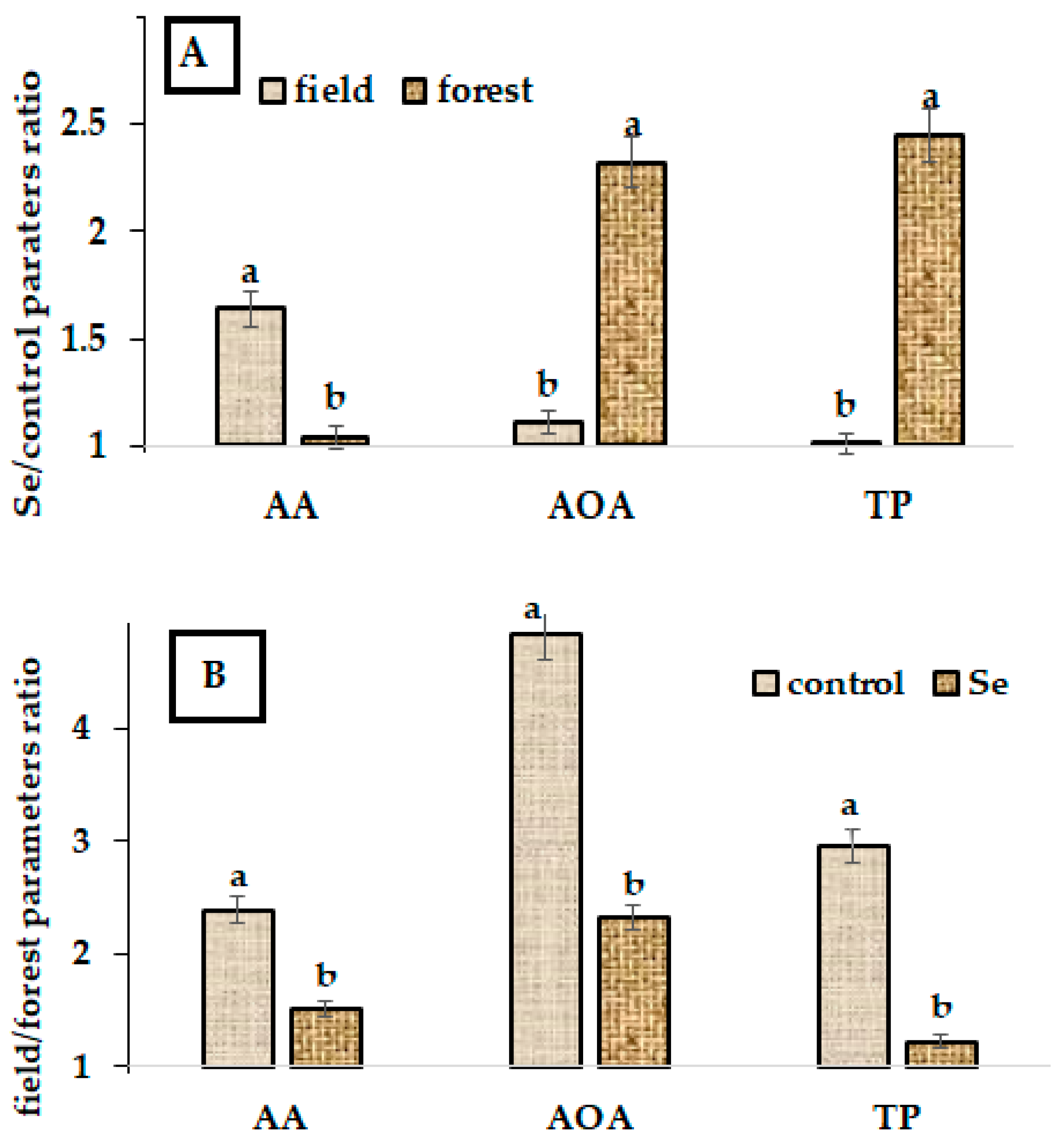
2.2. Biochemical Characteristics
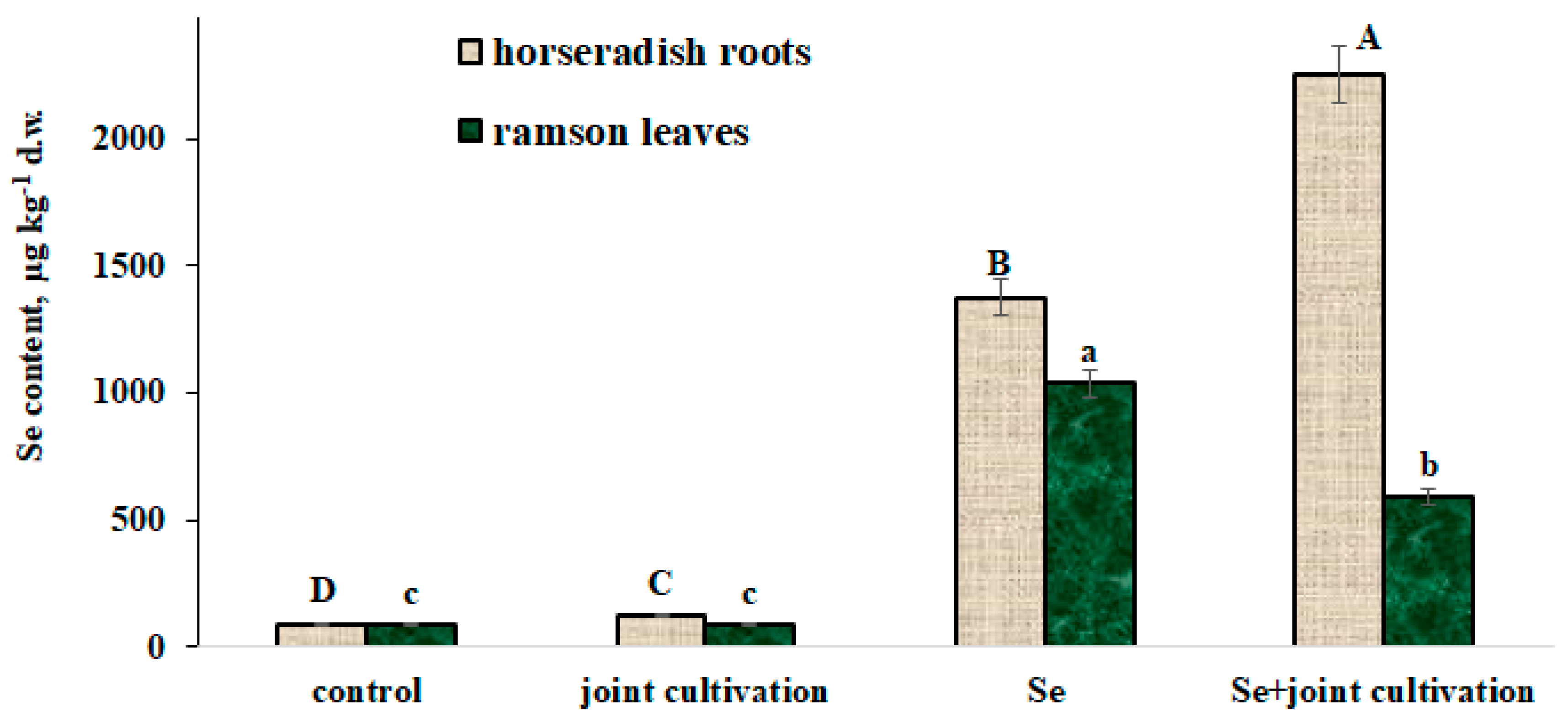
2.3. Se Accumulation
3. Materials and Methods
3.1. Object of Investigation and Cultivation Conditions
3.2. Biochemical Analysis
3.2.1. Dry Matter
3.2.2. Ascorbic Acid
3.2.3. Preparation of Ethanolic Extracts
3.2.4. Polyphenols
3.2.5. Antioxidant Activity (AOA)
3.2.6. Nitrates
3.2.7. Mono- and Di-Saccharides
3.2.8. Selenium
3.3. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Voća, S.; Šic Žlabur, J.; Fabek Uher, S.; Peša, M.; Opačić, N.; Radman, S. Neglected potential of wild garlic (Allium ursinum L.)—Specialized metabolites content and antioxidant capacity of wild populations in relation to location and plant phenophase. Horticulturae 2022, 8, 24. [Google Scholar] [CrossRef]
- Sobolewska, D.; Podolak, I.; Makowska-Was, J. Allium ursinum: Botanical, phytochemical and pharmacological overview. Phytochem. Rev. 2015, 14, 81–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leporatti, M.L.; Ivancheva, S. Preliminary comparative analysis of medicinal plants used in the traditional medicine of Bulgaria and Italy. J. Ethnopharm. 2003, 87, 123–142. [Google Scholar] [CrossRef]
- Pavlović, D.R.; Veljković, M.; Stojanović, N.M.; Gočmanac-Ignjatović, M.; Mihailov-Krstev, T.; Branković, S.; Sokolović, D.; Marčetić, M.; Radulović, N.; Radenković, M. Influence of different wild-garlic (Allium ursinum) extracts on the gastrointestinal system: Spasmolytic, antimicrobial and antioxidant properties. J. Pharm. Pharmacol. 2017, 69, 1208–1218. [Google Scholar] [CrossRef]
- Stanisavljević, N.; Soković Bajić, S.; Jovanović, Ž.; Matić, I.; Tolinaćki, M.; Popović, D.; Popović, N.; Terzic-Vidojević, A.; Golić, N.; Beškoski, V.; et al. Antioxidant’ and antiproliferative activity of Allium ursinum and their associated microbiota during simulated in vitro digestion in the presence of food matrix. Front. Microbiol. 2020, 11, 601616. [Google Scholar] [CrossRef]
- Rietz, B.; Isensee, H.; Strobach, H. Cardioprotective actions of wild garlic Allium ursinum in ischemia and reperfusion. Mol. Cell. Biochem. 1993, 119, 143–150. [Google Scholar] [CrossRef]
- Epure, L.I.; Roman, G.V.; Maracineanu, R. Studies on medicinal and aromatic plants used in the therapeutic recopies in the Bucharest university hospital. Sci. Pap. UASVM 2011, 54, 304–313. [Google Scholar]
- Amagova, Z.; Golubkina, N.; Matsadze, V.; Elmurzaeva, F.; Muligova, R.; Caruso, G. Biochemical characteristics of Allium ursinum L. sprouts as affected by the growing location in Chechen republic. Italus Hortus 2020, 27, 66–81. [Google Scholar] [CrossRef]
- Amagova, Z.A.; Matsadze, V.H.; Golubkina, N.A.; Seredin, T.M.; Caruso, G. Fortification of wild garlic with selenium. Veg. Crop. Russ. 2018, 4, 76–80. [Google Scholar] [CrossRef] [Green Version]
- Kieliszek, M.; Lipinski, B. Selenium supplementation in the prevention of coronavirus infections (COVID-19). Med. Hypotheses 2020, 143, 109878. [Google Scholar] [CrossRef]
- Golubkina, N.; Kharchenko, V.; Caruso, G. Chapter 7: Selenium: Prospects of Functional Food Production with High Antioxidant Activity. In Plant Antioxidants and Health, Reference Series in Phytochemistry; Ekiert, H., Ramawat, K.G., Arora, J., Eds.; Springer: New York, NY, USA, 2021; pp. 149–176. [Google Scholar] [CrossRef]
- Oborny, B.; Botta-Dukát, Z.; Rudolf, K.; Morschhauser, T. Population ecology of Allium ursinum, a space-monopolizing clonal plant. Acta Bot. Hung. 2011, 53, 371–388. [Google Scholar] [CrossRef]
- Djurdjevic, L.; Dinic, A.; Pavlović, P.; Mitrović, M.; Karadzic, B.; Tešević, V. Allelopathic potential of Allium ursinum L. Biochem. Syst. Ecol. 2004, 32, 533–544. [Google Scholar] [CrossRef]
- Ku, K.-M.; Jeffery, E.H.; Juvik, J.A.; Kushad, M.M. Correlation of quinone reductase activity and allyl isothiocyanate formation among different genotypes and grades of horseradish roots. J. Agric. Food Chem. 2015, 63, 2947–2955. [Google Scholar] [CrossRef]
- Agneta, R.; Möllers, C.; Rivelli, A.R. Horseradish (Armoracia rusticana), a neglected medical and condiment species with a relevant glucosinolate profile: A review. Genet. Resour. Crop Evol. 2013, 60, 1923–1943. [Google Scholar] [CrossRef]
- Abdalla, M.A.; Sulieman, S.; Mühling, K.H. Regulation of Selenium/Sulfur Interactions to Enhance Chemopreventive Effects: Lessons to Learn from Brassicaceae. Molecules 2020, 25, 5846. [Google Scholar] [CrossRef]
- Keck, A.-S.; Finley, J.W. Cruciferous Vegetables: Cancer Protective Mechanisms of Glucosinolate Hydrolysis Products and Selenium. Integr. Cancer Ther. 2004, 3, 5–12. [Google Scholar] [CrossRef]
- Revelou, P.-K.; Xagoraris, M.; Kokotou, M.G.; Constantinou-Kokotou, V. Cruciferous vegetables as functional foods: Effects of selenium biofortification. Int. J. Veg. Sci. 2022, 28, 191–210. [Google Scholar] [CrossRef]
- Itani, T.; Nakahata, Y.; Kato-Noguchi, H. Allelopathic activity of some herb plant species. Int. J. Agr. Biol. 2013, 15, 1569–1572. [Google Scholar]
- Nguyen, N.M.; Gonda, S.; Vasas, G. A review on the phytochemical composition and potential medicinal uses of horseradish (Armoracia rusticana) root. Food Rev. Int. 2013, 29, 261–275. [Google Scholar] [CrossRef]
- Malagoli, M.; Schiavon, M.; Dall’Acqua, S.; Pilon-Smits, E.A.H. Effects of selenium biofortification on crop nutritional quality. Front. Plant Sci. 2015, 6, 280. [Google Scholar] [CrossRef] [Green Version]
- Kuršvietienė, L.; Mongirdienė, A.; Bernatonienė, J.; Šulinskienė, J.; Stanevičienė, I. Selenium Anticancer Properties and Impact on Cellular Redox Status. Antioxidants 2020, 9, 80. [Google Scholar] [CrossRef] [PubMed]
- Antoshkina, M.; Golubkina, N.; Sekara, A.; Tallarita, A.; Caruso, G. Effects of selenium application on biochemical characteristics and biofortification level of kohlrabi produce. Front. Biosci.-Landmark 2021, 9, 533–542. [Google Scholar] [CrossRef]
- Gross, E.M. Allelopathy of aquatic autotrophs. Crit. Rev. Plant Sci. 2003, 22, 313–339. [Google Scholar] [CrossRef] [Green Version]
- Sipos, M.; Blidar, C.F.; Bunta, D. Allelopathic effects of aqueous extracts from horseradish (Armoracia rusticana L.) metamorphosed roots on several cereals. Rom. Agric. Res. 2012, 29, 169–173. [Google Scholar]
- Nguyen, T.; Ku, K.-M. Metabolomics and physiological approach to understand allelopathic effect of horseradish extract on onion root and lettuce seed as model organism. Plants 2021, 10, 1992. [Google Scholar] [CrossRef]
- Paciolla, C.; Fortunato, S.; Dipierro, N.; Paradiso, A.; De Leonardis, S.; Mastropasqua, L.; de Pinto, M.C. Vitamin C in plants: From functions to biofortification. Antioxidants 2019, 8, 519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivelli, A.R.; Caruso, M.C.; Galgano, S.; De, M.; Galgano, F. Vitamin C content in leaves and roots of horseradish (Armoracia rusticana): Seasonal variation in fresh tissues and retention as affected by storage conditions. Emirates J. Food Agric 2017, 29, 799–806. [Google Scholar] [CrossRef] [Green Version]
- de Sousa, G.F.; Silva, M.A.; de Morais, E.G.; Van Opbergen, G.A.Z.; Van Opbergen, G.G.A.Z.; de Oliveira, R.R.; Amaral, D.; Brown, P.; Chalfun-Junior, A.; Guilherme, L.R.G. Selenium enhances chilling stress tolerance in coffee species by modulating nutrient, carbohydrates, and amino acids content. Front. Plant Sci. 2022, 13, 1000430. [Google Scholar] [CrossRef]
- Mora, M.L.; Durán, P.; Acuña, J.; Cartes, P.; Demanet, R.; Gianfreda, L. Improving selenium status in plant nutrition and quality. J. Soil Sci. Plant Nutr. 2015, 15, 486–503. [Google Scholar] [CrossRef]
- Walters, S.A. Horseradish: A neglected and underutilized plant species for improving human health. Horticulturae 2021, 7, 167. [Google Scholar] [CrossRef]
- El Mehdawi, A.F.; Quinn, C.F.; Pilon-Smits, E.A.H. Effects of selenium hyperaccumulation on plant–plant interactions: Evidence for elemental allelopathy? New Phytol. 2011, 191, 120–131. [Google Scholar] [CrossRef]
- Golubkina, N.; Logvinenko, L.; Koshevarov, A.; Ushakova, O.; Kosheleva, O.; Shevchuk, O.; Caruso, G. Effects of intercropping on growth, elemental composition and antioxidant activity of Artemisia scoparia and Raphanus sativus under lead supply. Italus Hortus 2021, 28, 44–58. [Google Scholar] [CrossRef]
- Institute of Medicine: Panel on Dietary Antioxidants and Related Compounds. Dietary Reference Intakes for Vitamin C, Vitamin E, Selenium, and Carotenoids; National Academies Press (US): Washington, DC, USA, 2000. Available online: https://www.ncbi.nlm.nih.gov/books/NBK225470/ (accessed on 19 November 2018).
- Ávila, F.W.; Yang, Y.; Faquin, V.; Ramos, S.J.; Guilherme, L.R.G.; Thannhauser, T.W.; Li, L. Impact of selenium supply on Se-methylselenocysteine and glucosinolate accumulation in selenium-biofortified Brassica sprouts. Food Chem. 2014, 165, 578–586. [Google Scholar] [CrossRef]
- Wiesner-Reinhold, M.; Schreiner, M.; Baldermann, S.; Schwarz, D.; Hanschen, F.S.; Kipp, A.P.; Rowan, D.D.; Bentley-Hewitt, K.L.; McKenzie, M.J. Mechanisms of selenium enrichment and measurement in Brassicaceous vegetables, and their application to human health. Front Plant Sci. 2017, 8, 1365. [Google Scholar] [CrossRef] [Green Version]
- Barichman, T.C.; Kopsell, D.A.; Sams, C.E. Selenium influences glucosinolate and isothiocyanates and increases sulfur uptake in Arabidopsis thaliana and rapid-cycling Brassica oleracea. J. Agric. Food Chem. 2013, 61, 202–209. [Google Scholar] [CrossRef]
- AOAC Association Official Analytical Chemists. The Official Methods of Analysis of AOAC International; 22 Vitamin C; AOAC: Rockville, MD, USA, 2012. [Google Scholar]
- Golubkina, N.A.; Kekina, H.G.; Molchanova, A.V.; Antoshkina, M.S.; Nadezhkin, S.M.; Soldatenko, A.V. Plants Antioxidants and Methods of Their Determination; INFRA-M Academic Publishing LLC.: Moscow, Russia, 2020; (In Russian). [Google Scholar] [CrossRef]
- Swamy, P.M. Laboratory Manual on Biotechnology; Rastogi Publications: Meerut, India, 2008; p. 617. [Google Scholar]
- Alfthan, G.V. A micromethod for the determination of selenium in tissues and biological fluids by single-test-tube fluorimetry. Anal. Chim. Acta 1984, 165, 187–194. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameter | Control | Joint Cultivation | Se | Se + Joint Cultivation |
|---|---|---|---|---|
| Plant height (cm) | 35.0 ± 1.3 b | 39.1 ± 1.7 a | 38.7 ± 1.5 a | 40.9 ± 1.8 a |
| Leaf number | 2.3 ± 0.3 a | 2.6 ± 0.3 a | 2.6 ± 0.3 a | 2.7 ± 0.3 a |
| Leaf area (cm2) | 89.3 ± 11.4 b | 104.1 ± 15.5 ab | 116.3 ± 10.2 a | 118.2 ± 11.3 a |
| Plant biomass (g) | 14.8± 1.1 b | 17.4 ± 1.2 ab | 16.8 ± 1.1 ab | 18.9 ± 1.2 a |
| Yield (g m−2) | 980 ±64.2 b | 1069 ± 58.2 ab | 1046 ± 68.1 ab | 1145 ± 66.4 a |
| Parameter | Control | Joint Cultivation | Se | Se + Joint Cultivation |
|---|---|---|---|---|
| Plant height, cm | 89.0 ± 6.6 a | 92.2 ± 7.4 a | 88.7 ± 6.5 a | 94.5 ± 7.2 a |
| Leaf width, cm | 15.0 b | 17.4 a | 15.2 b | 18.0 a |
| Plant total mass, g | 290 ± 31 c | 394 ± 32 b | 320 ± 30 c | 495 ± 38 a |
| Plant root mass, g | 35 ± 2.5 c | 58 ± 3.7 a | 48 ± 4.1 b | 60 ± 5.0 a |
| Plant leaf mass, g | 255 ± 23 c | 336 ± 31 b | 272 ± 25 bc | 435 ± 40 a |
| Aboveground plant mass, kg m−2 | 1.02 ± 0.08c | 1.27 ± 0.10 b | 1.09 ± 0.09 bc | 1.74 ± 0.12 a |
| Parameter | Control | Joint Cultivation | Se | Se + Joint Cultivation |
|---|---|---|---|---|
| Dry matter, % | 11.75 ± 1.1 a | 11.0 ± 1.0 a | 11.70 ± 1.1 a | 11.1 ± 1.1 a |
| Nitrates, mg kg−1 d.w. | 971 ± 78 a | 983 ± 86 a | 1023 ± 91 a | 939 ± 90 a |
| Ascorbic acid, mg 100 g−1 d.w. | 328 ± 2 b | 555 ± 38 a | 532 ± 43 a | 543 ± 42 a |
| AOA *, mg GAE g−1 d.w. | 41.1 ± 2.6 b | 45.8 ± 2.8 ab | 45.7 ± 2.8 ab | 47.5 ± 3.1 a |
| TP **, mg GAE g−1 d.w. | 15.7 ± 1.1 a | 15.1 ± 1.0 a | 15.9 ± 1.2 a | 16.2 ± 1.2 a |
| Roots | ||||
|---|---|---|---|---|
| Parameter | Control | Joint Cultivation | Se | Se + Joint Cultivation |
| Dry matter, % | 23.4 ± 2.1 ab | 26.8 ± 2.2 a | 26.7 ± 2.3 a | 20.6 ± 2.0 b |
| Nitrates, mg kg−1 d.w. | 369 ± 32 b | 457 ± 41 a | 386 ± 33 ab | 324 ± 30 b |
| Ascorbic acid, mg 100 g−1 d.w. | 159 ± 14 b | 224 ± 21 a | 237 ± 22 a | 247 ± 21 a |
| AOA *, mg GAE g−1 d.w. | 17.4 ± 1.1 a | 17.2 ± 1.0 a | 17.6 ± 1.1 a | 19.3 ± 1.2 a |
| TP **, mg GAE g−1 d.w. | 7.9 ± 0.5 b | 9.2 ± 0.6 a | 9.3 ± 0.6 a | 10.2 ± 0.9 a |
| Monosaccharides, % d.w. | 5.5 ± 0.5 b | 5.7 ± 0.5 b | 8.7 ± 0.7 a | 8.0 ± 0.7 a |
| Total sugar, % d.w. | 23.6 ± 2.1 a | 22.3 ± 2.0 a | 19.8 ± 1.7 a | 22.5 ± 2.0 a |
| Leaves | ||||
| Dry matter, % | 26.0 ± 2.3 a | 15.7 ± 1.3 b | 22.5 ± 2.1 a | 15.7 ± 1.3 b |
| Nitrates, mg kg−1 d.w. | 170 ± 15 b | 195 ± 16 ab | 179 ± 15 b | 226 ± 18 a |
| Ascorbic acid, mg 100 g−1 d.w. | 255 ± 25 b | 349 ± 30 a | 380 ± 35 a | 376 ± 36 a |
| AOA, mg GAE g−1 d.w. | 42.8 ± 4.0 b | 60.2 ± 5.8 a | 56.8 ± 5.2 a | 63.9 ± 6.0 a |
| TP, mg GAE g−1 d.w. | 19.1 ± 1.6 a | 17.9 ± 1.4 a | 19.4 ± 1.6 a | 17.9 ± 1.4 a |
| Month | 2021 | 2022 | ||
|---|---|---|---|---|
| Mean Temperature, °C | Rainfall, mm | Mean Temperature, °C | Rainfall, mm | |
| January | 0.4 | 16 | 0.2 | 18 |
| February | −1 | 48 | 2.7 | 19 |
| March | 3.7 | 34 | 1.7 | 50 |
| April | 12.3 | 30 | 13 | 21 |
| May | 18.2 | 60 | 14.3 | 98 |
| June | 22.4 | 72 | ||
| July | 24.4 | 88 | ||
| August | 26 | 9 | ||
| September | 16.3 | 90 | ||
| October | 9.8 | 57 | ||
| November | 5.5 | 29 | ||
| December | 1.5 | 33 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amagova, Z.; Matsadze, V.; Kavarnakaeva, Z.; Golubkina, N.; Antoshkina, M.; Sękara, A.; Tallarita, A.; Caruso, G. Joint Cultivation of Allium ursinum and Armoracia rusticana under Foliar Sodium Selenate Supply. Plants 2022, 11, 2778. https://doi.org/10.3390/plants11202778
Amagova Z, Matsadze V, Kavarnakaeva Z, Golubkina N, Antoshkina M, Sękara A, Tallarita A, Caruso G. Joint Cultivation of Allium ursinum and Armoracia rusticana under Foliar Sodium Selenate Supply. Plants. 2022; 11(20):2778. https://doi.org/10.3390/plants11202778
Chicago/Turabian StyleAmagova, Zarema, Visita Matsadze, Zulfia Kavarnakaeva, Nadezhda Golubkina, Marina Antoshkina, Agnieszka Sękara, Alessio Tallarita, and Gianluca Caruso. 2022. "Joint Cultivation of Allium ursinum and Armoracia rusticana under Foliar Sodium Selenate Supply" Plants 11, no. 20: 2778. https://doi.org/10.3390/plants11202778