
Extracts of Fruits and Plants Cultivated In Vitro of Aristotelia chilensis (Mol.) Stuntz Show Inhibitory Activity of Aldose Reductase and Pancreatic Alpha-Amylase Enzymes

Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Preparation of Plant Extracts
4.2. Quantification of Total Anthocyanins by the Differential pH Method
4.3. Determination of Total Phenols by the Folin–Ciocalteu Method
4.4. Determination of Antioxidant Activity by DPPH Method
4.5. Determination of the Anthocyanins and Phenolic Compounds Profile of the Extracts by LC-MS/MS
4.6. Homogenization of Bovine Lenses
4.7. Bovine Aldose Reductase Activity
4.8. Pancreatic Alpha-Amylase Activity
4.9. Enzyme Inhibition
4.10. Statistical Treatment
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- De Faria, J. Classification of diabetes. In Diabetes. An Old Disease, a New Insight; Springer Science & Business Media, LLC: Berlin/Heidelberg, Germany, 2013; p. 485. [Google Scholar]
- Patel, D.; Kumar, R.; Kumar, M.; Sairam, K.; Hemalatha, S. Evaluation of in vitro aldose reductase inhibitory potential of different fraction of Hybanthus enneaspermus Linn F. Muell. Asian Pac. J. Trop. Biomed. 2012, 2, 134–139. [Google Scholar] [CrossRef] [Green Version]
- Ali, Y.; Jung, H.A.; Choi, J.S. Anti-diabetic and anti-Alzheimer’s disease activities of Angelica decursiva. Arch. Pharm. Res. 2015, 38, 2216–2227. [Google Scholar] [CrossRef]
- Forbes, J.M.; Cooper, M.E. Mechanisms of Diabetic Complications. Physiol. Rev. 2013, 93, 137–188. [Google Scholar] [CrossRef] [PubMed]
- Kawanishi, K.; Ueda, H.; Moriyasu, M. Aldose reductase inhibitors from the nature. Curr. Med. Chem. 2003, 10, 1353–1374. [Google Scholar] [CrossRef]
- Matsuda, H.; Morikawa, T.; Toguchida, I.; Harima, S.; Yoshikawa, M. Medicinal Flowers. VI. Absolute Stereostructures of Two New Flavanone Glycosides and a Phenylbutanoid Glycoside from the Flowers of Chrysanthemum indicum L.: Their Inhibitory Activities for Rat Lens Aldose Reductase. Chem. Pharm. Bull. 2002, 50, 972–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundarram, A.; Murthy, T. α-Amylase Production and Applications: A Review. J. Appl. Environ. Microbiol. 2014, 2, 166–175. [Google Scholar]
- Akileshwari, C.; Raghu, G.; Muthenna, P.; Mueller, N.H.; Suryanaryana, P.; Petrash, J.M.; Reddy, G.B. Bioflavonoid ellagic acid inhibits aldose reductase: Implications for prevention of diabetic complications. J. Funct. Foods 2014, 6, 374–383. [Google Scholar] [CrossRef]
- Suzen, S. Recent Studies of Aldose Reductase Enzyme Inhibition for Diabetic Complications. Curr. Med. Chem. 2003, 10, 1329–1352. [Google Scholar] [CrossRef]
- Rodriguez, J.P.; Lee, Y.K.; Woo, D.G.; Shim, J.S.; Geraldino, P.J.L.; Jacinto, S.D.; Lee, S. Flavonoids from Cirsium japonicum var. maackii pappus as inhibitors of aldose reductase and their simultaneous determination. Chem. Pap. 2017, 72, 81–88. [Google Scholar] [CrossRef]
- Fredes, C.; Montenegro, G.; Zoffoli, J.P.; Gómez, M.; Robert, P. Polyphenol content and antioxidant activity of maqui (Aristotelia chilensis Molina Stuntz) during fruit development and maturation in Central Chile. Chil. J. Agric. Res. 2012, 72, 582–589. [Google Scholar] [CrossRef] [Green Version]
- Genskowsky, E.; Puente, L.A.; Pérez-Álvarez, J.A.; Fernández-López, J.; Muñoz, L.A.; Viuda-Martos, M. Determination of polyphenolic profile, antioxidant activity and antibacterial properties of maqui [Aristotelia chilensi s (Molina) Stuntz] a Chilean blackberry. J. Sci. Food Agric. 2016, 96, 4235–4242. [Google Scholar] [CrossRef] [PubMed]
- Avello, M.; Valdivia, R.; Sanzana, R.; Mondaca, M.; Mennickent, S.; Aeschlimann, V.; Bittner, M.; Becerra, J. Extractos antioxidantes y antimicrobianos de Aristotelia chilensis y Ugni molinae y sus aplicaciones como preservantes en productos cosméticos. Bol. Latinoam. Caribe Plantas Med. Arom. 2009, 8, 479–486. [Google Scholar]
- Ruiz, A.; Hermosín-Gutiérrez, I.; Vergara, C.; von Baer, D.; Zapata, M.; Hitschfeld, A.; Obando, L.; Mardones, C. Anthocyanin profiles in south Patagonian wild berries by HPLC-DAD-ESI-MS/MS. Food Res. Int. 2013, 51, 706–713. [Google Scholar] [CrossRef]
- Vidal, L. Microencapsulated bioactive components as a source of health. In Encapsulations; Academic Press: Cambridge, MA, USA, 2016; Volume 2, p. 924. [Google Scholar]
- Vogel, H.; Peñailillo, P.; Doll, U.; Contreras, G.; Catenacci, G.; González, B. Maqui (Aristotelia chilensis): Morpho-phenological characterization to design high-yielding cultivation techniques. J. Appl. Res. Med. Aromat. Plants 2014, 1, 123–133. [Google Scholar] [CrossRef]
- Deroles, S. Anthocyanin Biosynthesis in Plant Cell Cultures: A Potential Source of Natural Colourants. In Anthocyanins; Winefield, C., Davies, K., Gould, K., Eds.; Springer: New York, NY, USA, 2008; pp. 108–167. [Google Scholar]
- Céspedes, C.; Mancinelli, P.; Orellana, B.; Silva, M. Cultivo in vitro de Aristotelia chilensis (Mol.) Stuntz Elaeocarpaceae. Gayana Bot. 1995, 52, 77–82. [Google Scholar]
- Díaz, P. Producción de antocianinas de Aristotelia chilensis en biorreactores para uso nutracéutico [Memoria para optar al Título de Ingeniera Civil en Biotecnología]. Bachelor’s Thesis, Facultad de Ciencias Físicas y Matemáticas, Universidad de Chile, Santiago, Chile, 2014. Available online: https://repositorio.uchile.cl/handle/2250/132019 (accessed on 8 September 2022).
- Sadino, M. Estrategias de cultivo e inducción in vitro de células de Aristotelia chilensis (Maqui) para la obtención de antocianinas [Memoria para optar al Título de Ingeniera Civil en Biotecnología. Facultad de Ciencias Físicas y Matemáticas]. Bachelor’s Thesis, Facultad de Ciencias Físicas y Matemáticas, Universidad de Chile, Santiago, Chile, 2015. Available online: https://repositorio.uchile.cl/handle/2250/136347 (accessed on 8 September 2022).
- Brauch, J.; Buchweitz, M.; Schweiggert, R.; Carle, R. Detailed analyses of fresh and dried maqui (Aristotelia chilensis (Mol.) Stuntz) berries and juice. Food Chem. 2016, 190, 308–316. [Google Scholar] [CrossRef]
- Ojo, O.A.; Amanze, J.C.; Oni, A.I.; Grant, S.; Iyobhebhe, M.; Elebiyo, T.C.; Rotimi, D.; Asogwa, N.T.; Oyinloye, B.E.; Ajiboye, B.O.; et al. Antidiabetic activity of avocado seeds (Persea americana Mill.) in diabetic rats via activation of PI3K/AKT signaling pathway. Sci. Rep. 2022, 12, 2919. [Google Scholar] [CrossRef]
- Alexiou, P.; Pegklidou, K.; Chatzopoulou, M.; Nicolaou, I.; Demopoulos, V.J. Aldose reductase enzyme and its implication to major health problems of the 21(st) century. Curr. Med. Chem. 2009, 16, 734–752. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [Green Version]
- Saltveit, M.E. Synthesis and Metabolism of Phenolic Compounds. In Fruit and Vegetable Phytochemicals; Wiley-Blackwell: Hoboken, NJ, USA, 2009; pp. 89–100. [Google Scholar] [CrossRef]
- Weisshaar, B.; Jenkins, G.I. Phenylpropanoid biosynthesis and its regulation. Curr. Opin. Plant Biol. 1998, 1, 251–257. [Google Scholar] [CrossRef]
- Horbowicz, M.; Kosson, R.; Grzesiuk, A.; Dębski, H. Anthocyanins of Fruits and Vegetables—Their Occurrence, Analysis and Role in Human Nutrition. J. Fruit Ornam. Plant Res. 2008, 68, 5–22. [Google Scholar] [CrossRef]
- Garzón, G. Las antocianinas como colorantes naturales y compuestos bioactivos: Review. Acta Biol. Colomb. 2008, 13, 3. [Google Scholar] [CrossRef]
- Gras, C.C.; Nemetz, N.; Carle, R.; Schweiggert, R.M. Anthocyanins from purple sweet potato (Ipomoea batatas (L.) Lam.) and their color modulation by the addition of phenolic acids and food-grade phenolic plant extracts. Food Chem. 2017, 235, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Gironés-Vilaplana, A.; Mena, P.; García-Viguera, C.; Moreno, D.A. A novel beverage rich in antioxidant phenolics: Maqui berry (Aristotelia chilensis) and lemon juice. LWT 2012, 47, 279–286. [Google Scholar] [CrossRef]
- Mazzuca, P.; Ferranti, P.; Picariello, G.; Chianese, L.; Addeo, F. Mass spectrometry in the study of anthocyanins and their derivatives: Differentiation of Vitis vinifera and hybrid grapes by liquid chromatography/electrospray ionization mass spectrometry and tandem mass spectrometry. Biol. Mass Spectrom. 2005, 40, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Aaby, K.; Ekeberg, A.D.; Skrede, G. Characterization of Phenolic Compounds in Strawberry (Fragaria × ananassa) Fruits by Different HPLC Detectors and Contribution of Individual Compounds to Total Antioxidant Capacity. J. Agric. Food Chem. 2007, 55, 4395–4406. [Google Scholar] [CrossRef]
- Trikas, E.D.; Papi, R.M.; Kyriakidis, D.A.; Zachariadis, G.A. A Sensitive LC-MS Method for Anthocyanins and Comparison of Byproducts and Equivalent Wine Content. Separations 2016, 3, 18. [Google Scholar] [CrossRef]
- Zhang, S.-L.; Deng, P.; Xu, Y.-C.; Lü, S.-W.; Wang, J.-J. Quantification and analysis of anthocyanin and flavonoids compositions, and antioxidant activities in onions with three different colors. J. Integr. Agric. 2016, 15, 2175–2181. [Google Scholar] [CrossRef] [Green Version]
- Escribano-Bailon, M.T.; Alcalde-Eon, C.; Muñoz, O.; Rivas-Gonzalo, J.C.; Santos-Buelga, C. Anthocyanins in berries of Maqui [Aristotelia chilensis (Mol.) Stuntz]. Phytochem. Anal. 2005, 17, 8–14. [Google Scholar] [CrossRef]
- Jara, E.; Hidalgo, J.; Flores, C.; Pérez, M.; Yáñez, A.; Hidalgo, A.; Quiñones, L.; Hancke, J.; Burgos, R. Estudio de un extracto estandarizado de maqui rico en delfinidinas en el mantenimiento del balance de glucosa. Rev. Farmacol. Chile 2012, 5, 27–34. [Google Scholar]
- Rojo, L.E.; Ribnicky, D.; Logendra, S.; Poulev, A.; Rojas-Silva, P.; Kuhn, P.; Dorn, R.; Grace, M.H.; Lila, M.A.; Raskin, I. In vitro and in vivo anti-diabetic effects of anthocyanins from Maqui Berry (Aristotelia chilensis). Food Chem. 2012, 131, 387–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, S.; Tanaka, J.; Imada, T.; Shimoda, H.; Tsubota, K. Delphinidin 3,5-O-diglucoside, a constituent of the maqui berry (Aristotelia chilensis) anthocyanin, restores tear secretion in a rat dry eye model. J. Funct. Foods 2014, 10, 346–354. [Google Scholar] [CrossRef]
- Abu-Reidah, I.M.; Ali-Shtayeh, M.S.; Jamous, R.; Arraez-Roman, D.; Segura-Carretero, A. HPLC–DAD–ESI-MS/MS screening of bioactive components from Rhus coriaria L. (Sumac) fruits. Food Chem. 2015, 166, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Karim, A.A.; Azlan, A.; Ismail, A.; Hashim, P.; Gani, S.S.A.; Zainudin, B.H.; Abdullah, N.A. Phenolic composition, antioxidant, anti-wrinkles and tyrosinase inhibitory activities of cocoa pod extract. BMC Complement. Altern. Med. 2014, 14, 381. [Google Scholar] [CrossRef] [Green Version]
- Bochi, V.C.; Godoy, H.T.; Giusti, M.M. Anthocyanin and other phenolic compounds in Ceylon gooseberry (Dovyalis hebecarpa) fruits. Food Chem. 2015, 176, 234–243. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Singh, A.; Kumar, B. Identification and characterization of phenolics and terpenoids from ethanolic extracts of Phyllanthus species by HPLC-ESI-QTOF-MS/MS. J. Pharm. Anal. 2017, 7, 214–222. [Google Scholar] [CrossRef]
- Dartora, N.; de Souza, L.M.; de Santana-Filho, A.P.; Iacomini, M.; Valduga, A.T.; Gorin, P.A.; Sassaki, G.L. UPLC-PDA–MS evaluation of bioactive compounds from leaves of Ilex paraguariensis with different growth conditions, treatments and ageing. Food Chem. 2011, 129, 1453–1461. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, Y.; Garud, M.; Oza, M.; Barve, K.; Gaikwad, A. Diabetes, diabetic complications, and flavonoids. In Fruits, Vegetables, and Herbs; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 77–104. [Google Scholar] [CrossRef]
- Cespedes, C.L.; Balbontin, C.; Avila, J.G.; Dominguez, M.; Alarcon, J.; Paz, C.; Burgos, V.; Ortiz, L.; Peñaloza-Castro, I.; Seigler, D.S.; et al. Inhibition on cholinesterase and tyrosinase by alkaloids and phenolics from Aristotelia chilensis leaves. Food Chem. Toxicol. 2017, 109, 984–995. [Google Scholar] [CrossRef]
- Abu-Reidah, I.M.; Arráez-Román, D.; Lozano-Sánchez, J.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Phytochemical Characterisation of Green Beans (Phaseolus vulgaris L.) by Using High-performance Liquid Chromatography Coupled with Time-of-flight Mass Spectrometry. Phytochem. Anal. 2012, 24, 105–116. [Google Scholar] [CrossRef]
- Cetto, A.A.; Wiedenfeld, H.; Revilla, M.C.; Sergio, I.A. Hypoglycemic effect of Equisetum myriochaetum aerial parts on streptozotocin diabetic rats. J. Ethnopharmacol. 2000, 72, 129–133. [Google Scholar] [CrossRef]
- Sirotkin, A.V.; Hrabovszká, S.; Štochmaľová, A.; Grossmann, R.; Alwasel, S.; Harrath, A.H. Effect of quercetin on ovarian cells of pigs and cattle. Anim. Reprod. Sci. 2019, 205, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.-N.; Kang, M.-J.; Lee, S.-J.; Kim, J.-I. Ameliorative effect of myricetin on insulin resistance in mice fed a high-fat, high-sucrose diet. Nutr. Res. Pract. 2014, 8, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Aslan, H.E.; Beydemir, S. Phenolic compounds: The inhibition effect on polyol pathway enzymes. Chem. Interact. 2017, 266, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Ogiwara, T.; Satoh, K.; Kadoma, Y.; Murakami, Y.; Unten, S.; Atsumi, T.; Sakagami, H.; Fujisawa, S. Radical scavenging activity and cytotoxicity of ferulic acid. Anticancer Res. 2002, 22, 2711–2717. [Google Scholar] [PubMed]
- Gülçin, I. Antioxidant activity of caffeic acid (3,4-dihydroxycinnamic acid). Toxicology 2006, 217, 213–220. [Google Scholar] [CrossRef]
- Calderon-Montaño, J.M.; Burgos-Morón, E.; Perez-Guerrero, C.; Lopez-Lazaro, M. A Review on the Dietary Flavonoid Kaempferol. Mini-Rev. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef]
- Fujita, T.; Funako, T.; Hayashi, H. 8-Hydroxydaidzein, an Aldose Reductase Inhibitor from Okara Fermented with Aspergillus sp. HK-388. Biosci. Biotechnol. Biochem. 2004, 68, 1588–1590. [Google Scholar] [CrossRef] [Green Version]
- Naeem, S.; Hylands, P.; Barlow, D. Construction of an Indonesian herbal constituents database and its use in Random Forest modelling in a search for inhibitors of aldose reductase. Bioorg. Med. Chem. 2012, 20, 1251–1258. [Google Scholar] [CrossRef]
- Kraft, T. Protective Potential of Fruits Against Diabetes and Its Complications. Ph.D. Thesis, University of Illinois at Urbana-Champaign, Champaign, IL, USA, 2010. [Google Scholar]
- Chethan, S.; Dharmesh, S.M.; Malleshi, N.G. Inhibition of aldose reductase from cataracted eye lenses by finger millet (Eleusine coracana) polyphenols. Bioorg. Med. Chem. 2008, 16, 10085–10090. [Google Scholar] [CrossRef]
- Kato, A.; Minoshima, Y.; Yamamoto, J.; Adachi, I.; Watson, A.A.; Nash, R.J. Protective Effects of Dietary Chamomile Tea on Diabetic Complications. J. Agric. Food Chem. 2008, 56, 8206–8211. [Google Scholar] [CrossRef]
- El-Beshbishy, H.; Bahashwan, S. Hypoglycemic effect of basil (Ocimum basilicum) aqueous extract is mediated through inhibition of α-glucosidase and α-amylase activities. Toxicol. Ind. Health 2011, 28, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Ranilla, L.G.; Kwon, Y.-I.; Apostolidis, E.; Shetty, K. Phenolic compounds, antioxidant activity and in vitro inhibitory potential against key enzymes relevant for hyperglycemia and hypertension of commonly used medicinal plants, herbs and spices in Latin America. Bioresour. Technol. 2010, 101, 4676–4689. [Google Scholar] [CrossRef]
- Rubilar, M.; Jara, C.; Poo, Y.; Acevedo, F.; Gutierrez, C.; Sineiro, J.; Shene, C. Extracts of Maqui (Aristotelia chilensis) and Murta (Ugni molinae Turcz.): Sources of Antioxidant Compounds and α-Glucosidase/α-Amylase Inhibitors. J. Agric. Food Chem. 2011, 59, 1630–1637. [Google Scholar] [CrossRef] [PubMed]
- Giusti, M.M.; Wrolstad, R.E. Characterization and Measurement of Anthocyanins by UV-Visible Spectroscopy. Curr. Protoc. Food Anal. Chem. 2001. [Google Scholar]
- Singleton, V.; Rossi, J. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Joyeux, M.; Lobstein, A.; Anton, R.; Mortier, F. Comparative Antilipoperoxidant, Antinecrotic and Scavanging Properties of Terpenes and Biflavones from Ginkgo and some Flavonoids. Planta Med. 1995, 61, 126–129. [Google Scholar] [CrossRef] [PubMed]
- Contreras, R.A.; Köhler, H.; Pizarro, M.; Zúñiga, G.E. In Vitro Cultivars of Vaccinium corymbosum L. (Ericaceae) are a Source of Antioxidant Phenolics. Antioxidants 2015, 4, 281–292. [Google Scholar] [CrossRef]
- Del-Corso, A.; Balestri, F.; Di Bugno, E.; Moschini, R.; Cappiello, M.; Sartini, S.; La-Motta, C.; Da-Settimo, F.; Mura, U. A New Approach to Control the Enigmatic Activity of Aldose Reductase. PLoS ONE 2013, 8, e74076. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Total Anthocyanin | Total Phenol | DPPH (IC50) |
|---|---|---|---|
| mg Anthocyanin/g DW | mg GAE/g DW | mg/g DW | |
| Fruit | 7.63 ± 0.09 a | 79.90 ± 3.77 c | 0.11 ± 0.01 e |
| In vitro plant | 0.40 ± 0.01 b | 19.65 ± 2.06 d | 0.12 ± 0.01 e |
| Peak | RT | Molecule | Chemical Formula | [M-H]+ m/z | Relative Abundance (%) |
|---|---|---|---|---|---|
| 1 | 0.74 | Cyanidin 3-(3″,6″-dimalonylglucoside) | C27H24O17 | 621.1 | 9.71 |
| 2 | 0.86 | Pelargonidin 3-glucoside | C21H20O10 | 429.1 | 1.06 |
| 3 | 1.09 | Delphinidin 3,5-diglucoside | C27H30O17 | 627.1 | 20.53 |
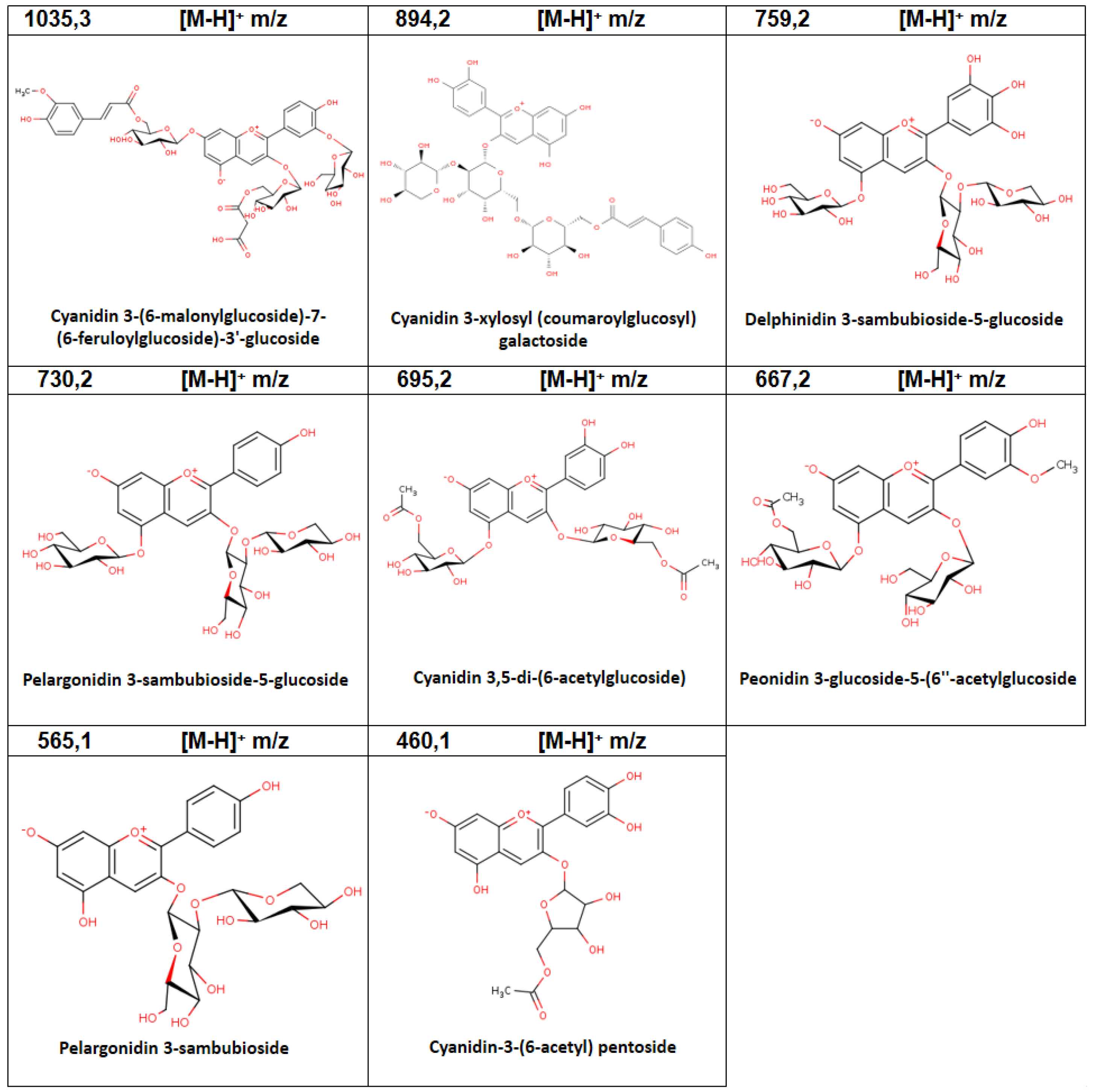
| 4 | 1.39 | Cyanidin-3-(6-acetyl) pentoside | C22H20O11 | 460.1 | 1.20 |
| 5 | 2.86 | Delphinidin 3-(2″-galloylgalactoside) | C28H24O16 | 617.1 | 8.58 |
| 6 | 3.44 | Delphinidin 3,7-diglucoside-3′,5′-di (6-p-coumaroyl-beta-glucoside) | C57H62O31 | 1242.3 | 0.68 |
| 7 | 3.75 | Pelargonidin 3-sambubioside | C26H28O14 | 565.1 | 0.23 |
| 8 | 4.04 | Delphinidin 3-sambubioside-5-glucoside | C32H38O21 | 759.2 | 2.00 |
| 9 | 5.23 | Peonidin 3-glucoside-5- (6″-acetylglucoside) | C30H34O17 | 667.2 | 0.70 |
| 10 | 7.54 | Peonidin 3-rutinoside | C28H32O15 | 607.2 | 3.41 |
| 11 | 8.21 | Cyanidin 3-(6-malonylglucoside)-7- (6-feruloylglucoside)-3′-glucoside | C46H50O27 | 1035.3 | 1.86 |
| 12 | 9.03 | Cyanidin 3-xylosyl (coumaroylglucosyl) galactoside | C41H45O22 | 894.2 | 4.33 |
| 13 | 9.48 | Cyanidin 3-sambubioside | C26H29O15 | 587.2 | 1.71 |
| 14 | 10.06 | Malvidin 3-rutinoside | C29H34O16 | 638.2 | 7.21 |
| 15 | 10.32 | Cyanidin-3-(2′-acetylrutinoside) | C29H32O31 | 635.2 | 5.53 |
| 16 | 11.08 | Cyanidin 3,5-di-(6-acetylglucoside) | C31H34O18 | 695.2 | 0.97 |
| 17 | 11.29 | Cyanidin 3-[6-(6-p-coumarylglucosyl)- 2-xylosylgalactoside] | C41H14O22 | 887.2 | 1.16 |
| 18 | 11.51 | Pelargonidin 3-sambubioside-5-glucoside | C32H38O19 | 730.2 | 0.72 |
| 19 | 11.84 | Pelargonidin 3-(6″-p-coumaryl sambubioside)-5-(6″′-malonylglucoside) | C44H46O24 | 958.2 | 1.45 |
| 20 | 12.10 | Delphinidin-3-(6-feruloyl)-5-diglucoside | C27H50O27 | 803.1 | 26.95 |
| Peak | RT | Molecule | Chemical Formula | [M-H]+ m/z | Relative Abundance (%) |
|---|---|---|---|---|---|
| 1 | 0.83 | Pelargonidin 3-glucoside | C21H20O10 | 429.1 | 10.27 |
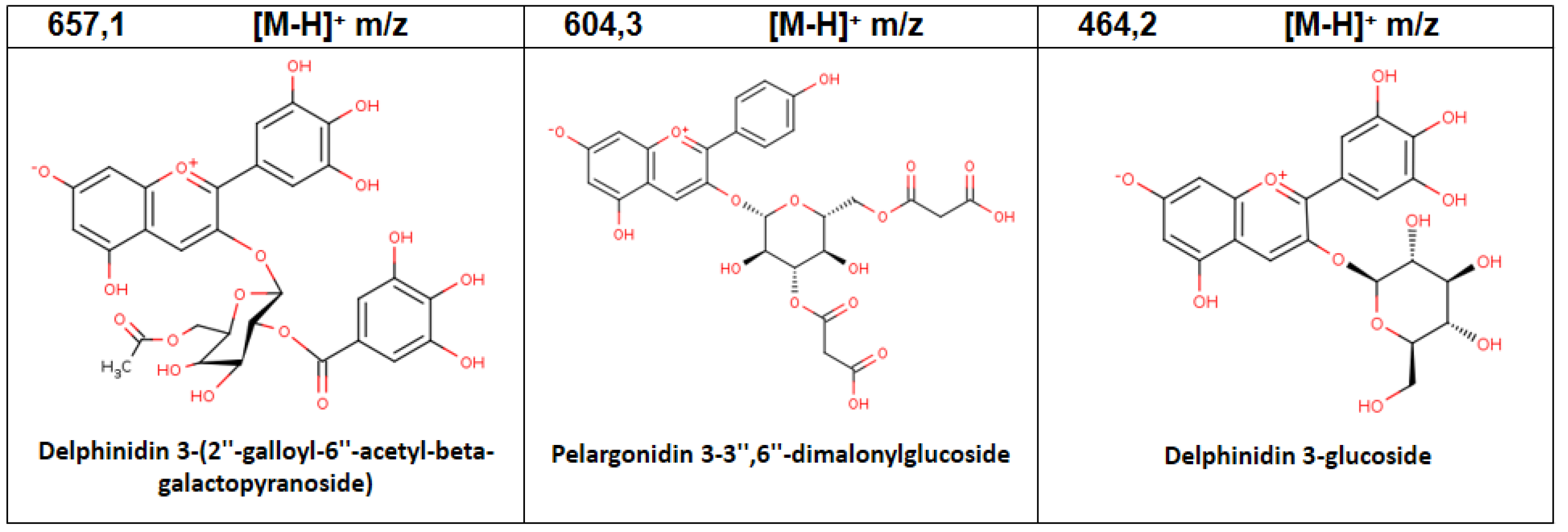
| 2 | 1.01 | Delphinidin 3-glucoside | C21H20O12 | 464.2 | 14.32 |
| 3 | 1.38 | Cyanidin 3-(3″,6″-dimalonylglucoside) | C27H24O17 | 621.1 | 2.84 |
| 4 | 1.66 | Cyanidin 3-sambubioside | C26H29O15 | 587.2 | 2.73 |
| 5 | 2.39 | Delphinidin 3,7-diglucoside-3′,5′- di(6-p-coumaroyl-beta-glucoside) | C57H62O31 | 1242.3 | 0.47 |
| 6 | 2.73 | Pelargonidin 3-3″,6″-dimalonylglucoside | C27H42O16 | 604.3 | 1.31 |
| 7 | 4.17 | Cyanidin-3-(2′-acetylrutinoside) | C29H32O16 | 635.2 | 2.50 |
| 8 | 4.49 | Delphinidin 3,5-diglucoside | C27H30O17 | 627.1 | 0.91 |
| 9 | 6.83 | Delphinidin 3-(2″-galloyl-6″-acetyl-beta-galactopyranoside) | C30H26O17 | 657.1 | 3.04 |
| 10 | 7.32 | Pelargonidin 3-(6″-p-coumaryl sambubioside)-5-(6″′-malonylglucoside) | C44H46O24 | 958.2 | 0.80 |
| 11 | 7.52 | Peonidin 3-rutinoside | C28H32O15 | 607.2 | 5.50 |
| 12 | 8.26 | Malvidin 3-rutinoside | C29H34O16 | 638.2 | 0.51 |
| 13 | 8.88 | Delphinidin 3-(2″-galloylgalactoside) | C28H24O16 | 617.1 | 1.61 |
| 14 | 9.51 | Cyanidin 3-[6-(6-p-coumarylglucosyl)- 2-xylosylgalactoside] | C41H44O22 | 887.2 | 0.94 |
| 15 | 12.09 | Delphinidin-3-(6-feruloyl)-5-diglucoside | C46H50O27 | 803.1 | 52.24 |
| Peak | RT | Molecule | Chemical Formula | [M-H]− m/z | Relative Abundance (%) |
|---|---|---|---|---|---|
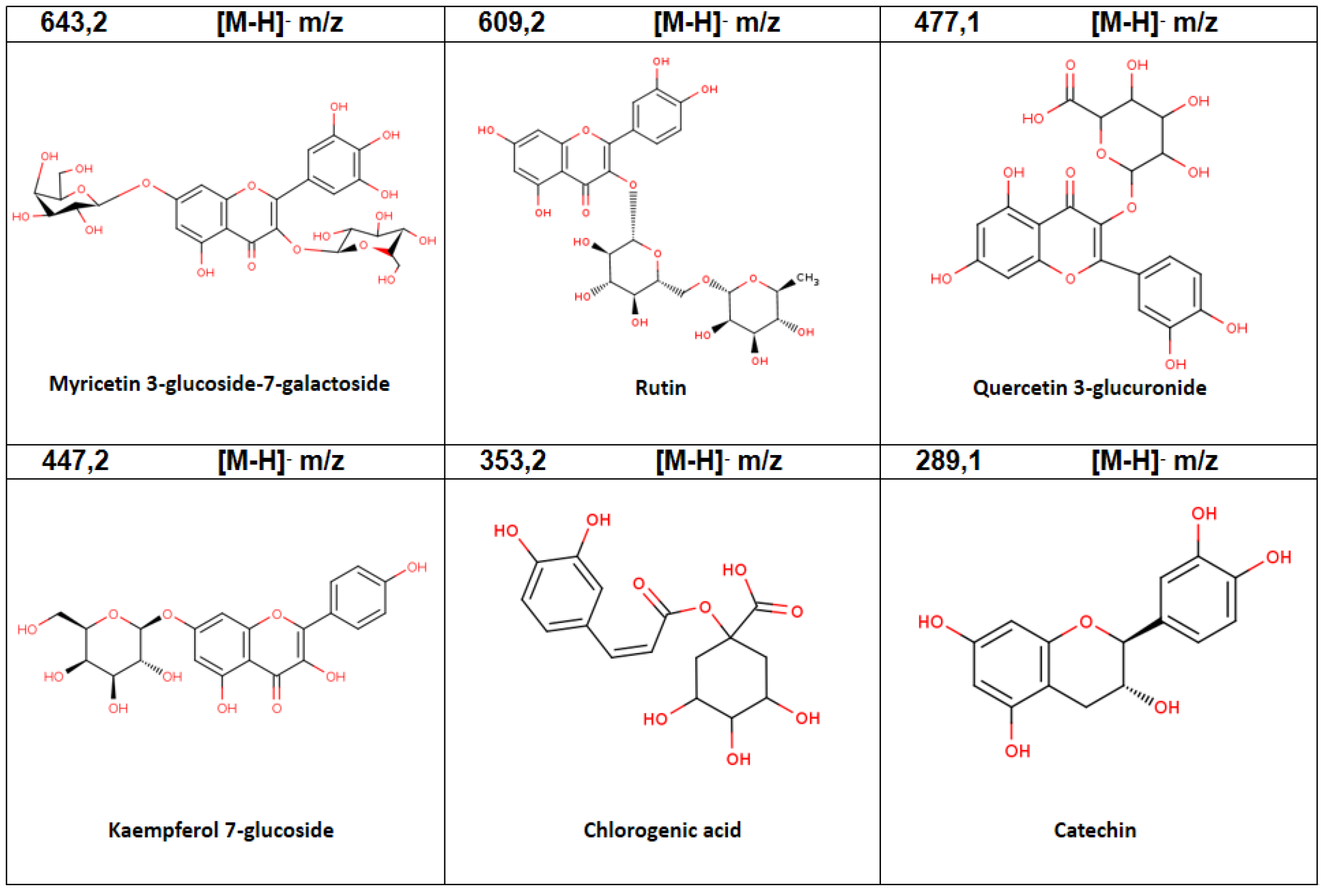
| 1 | 0.74 | Catechin | C15H14O6 | 289.1 | 2.53 |
| 2 | 1.13 | Quercetin 4′-galactoside | C20H18O12 | 451.1 | 35.90 |
| 3 | 1.67 | Chlorogenic acid | C16H18O9 | 353.2 | 2.68 |
| 4 | 2.93 | 3,5-di-chlorogenic acid | C25H24O12 | 515.2 | 22.42 |
| 5 | 3.47 | Myricetin 3-glucoside-7-galactoside | C27H30O18 | 643.2 | 3.68 |
| 6 | 4.05 | Kaempferol 3-[2″-glucosyl-6″-acetyl-galactoside] 7-glucoside | C27H30O19 | 813.1 | 3.51 |
| 7 | 6.18 | Tetramethylquercetin 3-rutinoside | C31H38O16 | 666.2 | 3.25 |
| 8 | 7.67 | Granatin B | C41H28O27 | 951.1 | 1.51 |
| 9 | 8.52 | Rutin | C27H30O16 | 609.2 | 1.14 |
| 10 | 9.06 | Quercetin 3-glucuronide | C21H18O13 | 477.1 | 0.95 |
| 11 | 10.79 | Quercetin 3-(6″″-ferulylsophorotrioside) | C43H48O25 | 965.3 | 1.48 |
| 12 | 11.88 | Kaempferol 3-(4″,6″-diacetylglucoside)- 7-rhamnoside | C31H34O17 | 679.2 | 6.34 |
| 13 | 13.30 | Kaempferol 7-glucoside | C21H20O11 | 447.2 | 14.61 |
| Peak | RT | Molecule | Chemical Formula | [M-H]− m/z | Relative Abundance (%) |
|---|---|---|---|---|---|
| 1 | 0.83 | 3,5-di-chlorogenic acid | C25H24O12 | 515.2 | 6.11 |
| 2 | 1.11 | Quercetin 4′-galactoside | C20H18O12 | 451.1 | 15.94 |
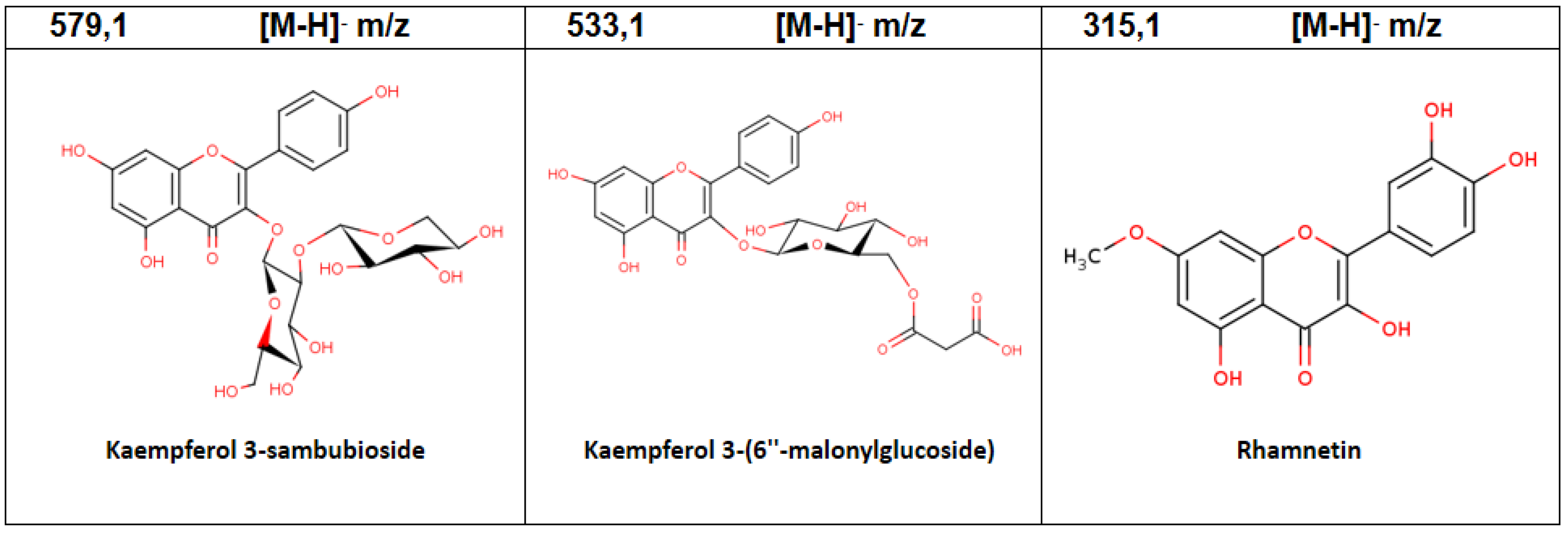
| 3 | 1.36 | Kaempferol 3-(6″-malonylglucoside) | C24H22O14 | 533.1 | 7.65 |
| 4 | 1.69 | Rhamnetin | C16H12O7 | 315.1 | 12.16 |
| 5 | 3.70 | Tetramethylquercetin 3-rutinoside | C31H38O16 | 666.2 | 1.93 |
| 6 | 4.33 | Kaempferol 3-[2″-glucosyl-6″-acetyl-galactoside] 7-glucoside | C27H30O19 | 813.1 | 1.29 |
| 7 | 4.57 | Kaempferol 3-sambubioside | C26H28O15 | 579.1 | 5.59 |
| 8 | 7.33 | Quercetin 3-(6″″-ferulylsophorotrioside) | C43H48O25 | 965.3 | 2.78 |
| 9 | 7.98 | Granatin B | C41H28O27 | 951.1 | 11.17 |
| 10 | 9.90 | Kaempferol 3-(4″,6″-diacetylglucoside)- 7-rhamnoside | C31H34O17 | 679.2 | 35.38 |
| Inhibitor | AR % Inhibition | AA % Inhibition |
|---|---|---|
| Fruit | 85.54 ± 1.86 a | 29.64 ± 0.63 a |
| In vitro plant | 75.67 ± 1.21 b | 47.66 ± 0.66 b |
| Delphinidin-3-O-glucoside | 93.45 ± 5.67 c | 73.01 ± 0.05 c |
| Cyanidin-3-O-glucoside | 83.52 ± 1.98 a | 51.96 ± 0.22 d |
| Pelargonidin-3-O-glucoside | 77.33 ± 1.26 b | 44.56 ± 1.37 b |
| Peonidin-3-O-glucoside | 44.97 ± 1.61 d | 54.92 ± 1.42 d |
| Quercetin | 74.47 ± 1.78 b | 29.96 ± 5.27 a |
| Ellagic acid | 93.97 ± 2.92 c | 15.16 ± 0.27 e |
| Acarbose | NT | 96.70 ± 0.26 f |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pineda, A.; Arenas, A.; Balmaceda, J.; Zúñiga, G.E. Extracts of Fruits and Plants Cultivated In Vitro of Aristotelia chilensis (Mol.) Stuntz Show Inhibitory Activity of Aldose Reductase and Pancreatic Alpha-Amylase Enzymes. Plants 2022, 11, 2772. https://doi.org/10.3390/plants11202772
Pineda A, Arenas A, Balmaceda J, Zúñiga GE. Extracts of Fruits and Plants Cultivated In Vitro of Aristotelia chilensis (Mol.) Stuntz Show Inhibitory Activity of Aldose Reductase and Pancreatic Alpha-Amylase Enzymes. Plants. 2022; 11(20):2772. https://doi.org/10.3390/plants11202772
Chicago/Turabian StylePineda, Adriana, Andrea Arenas, Juan Balmaceda, and Gustavo E. Zúñiga. 2022. "Extracts of Fruits and Plants Cultivated In Vitro of Aristotelia chilensis (Mol.) Stuntz Show Inhibitory Activity of Aldose Reductase and Pancreatic Alpha-Amylase Enzymes" Plants 11, no. 20: 2772. https://doi.org/10.3390/plants11202772