
Agronomic Behavior of Mexican Roselle Cultivars Produced under Protected Agricultural Conditions

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
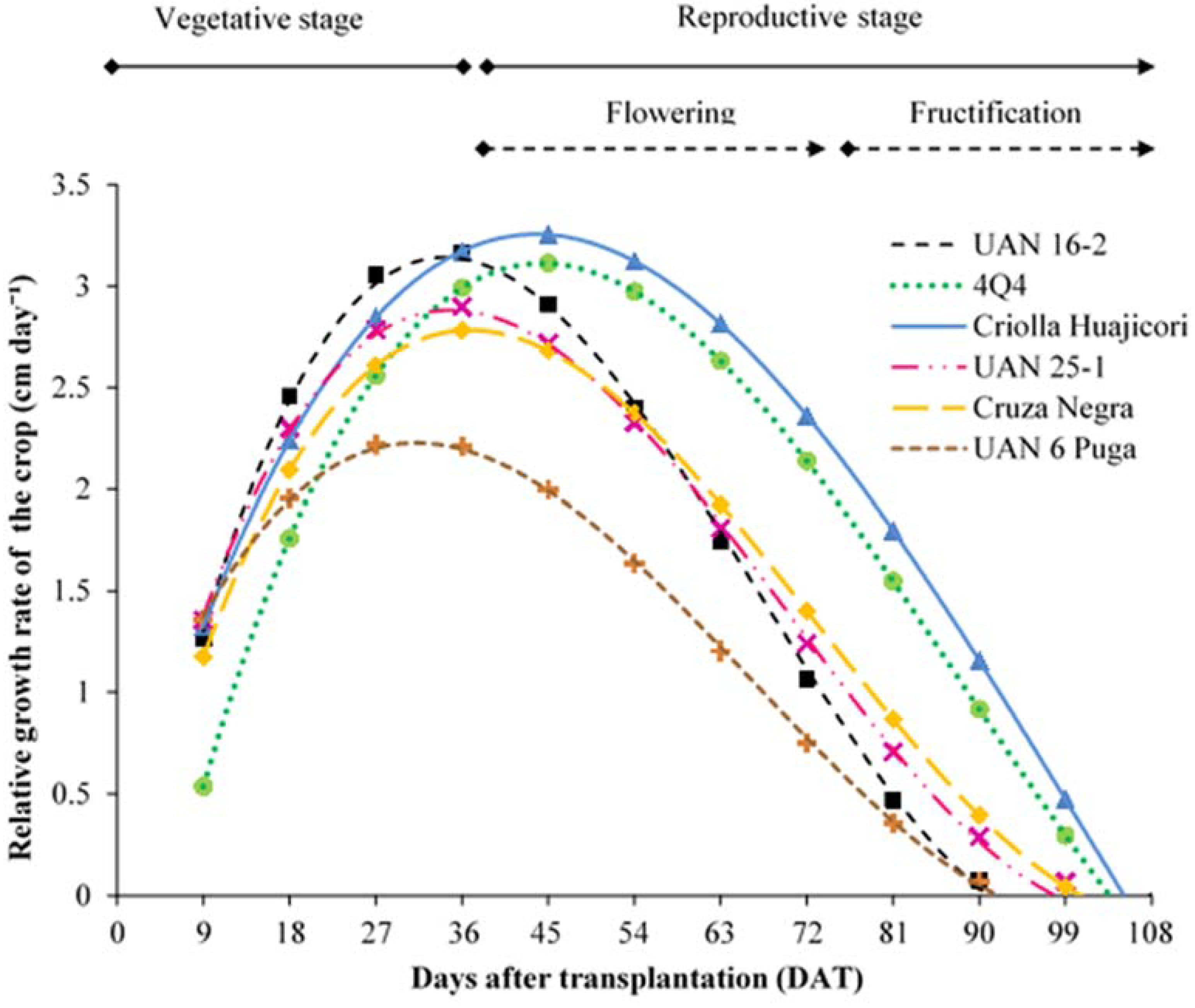
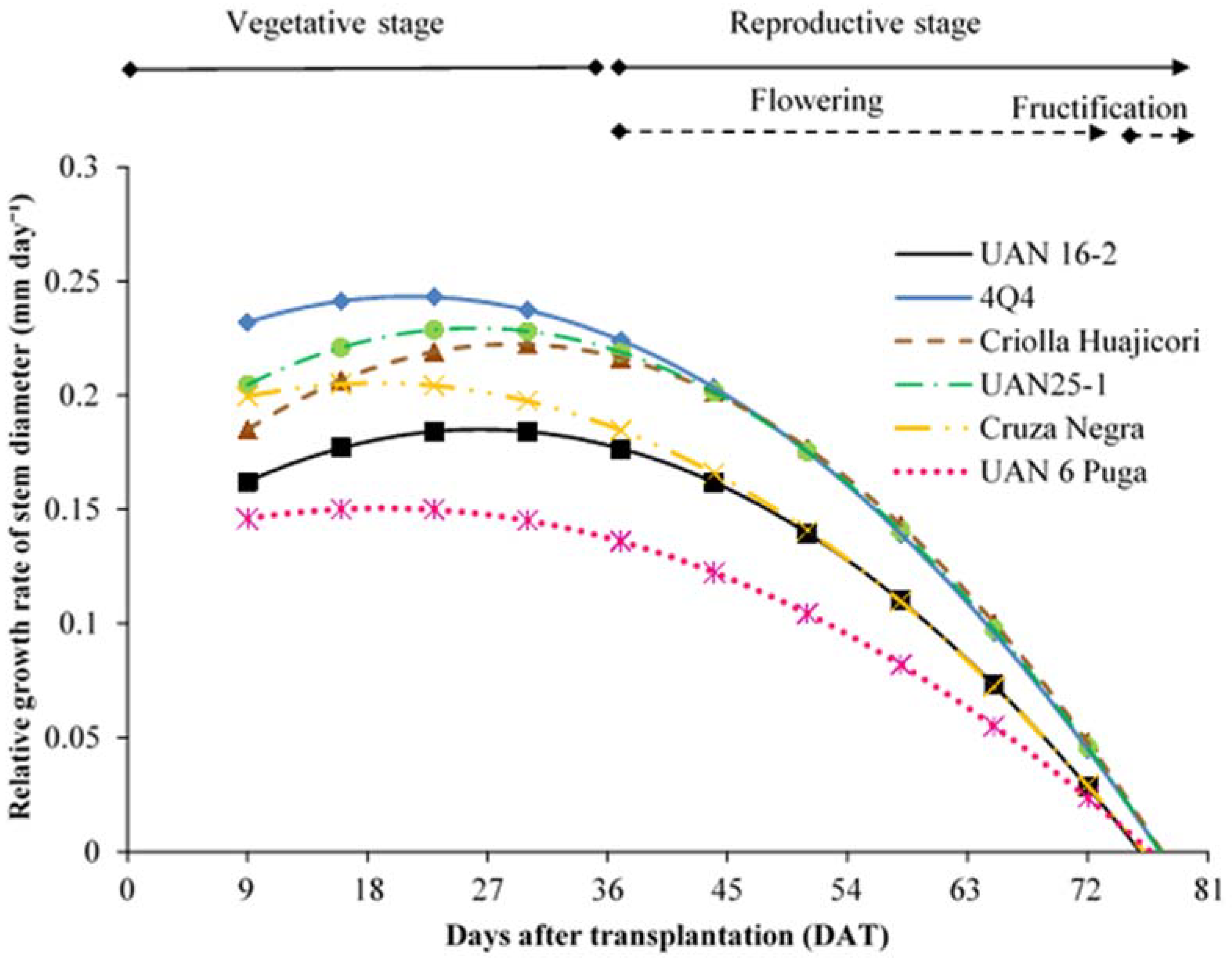
2.1. Growth Rates
2.2. Growth Variables
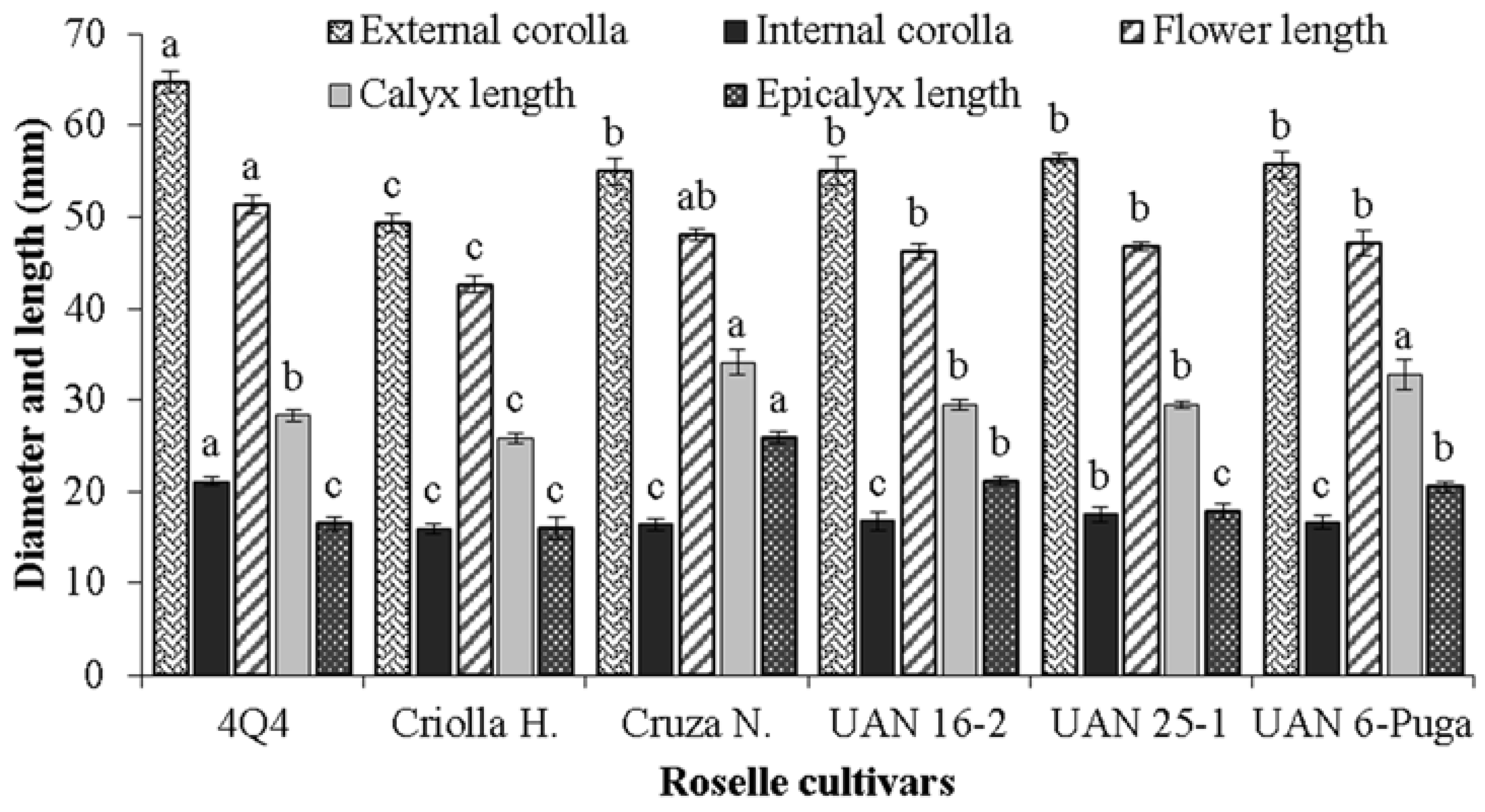
2.3. Roselle Flower Measurements
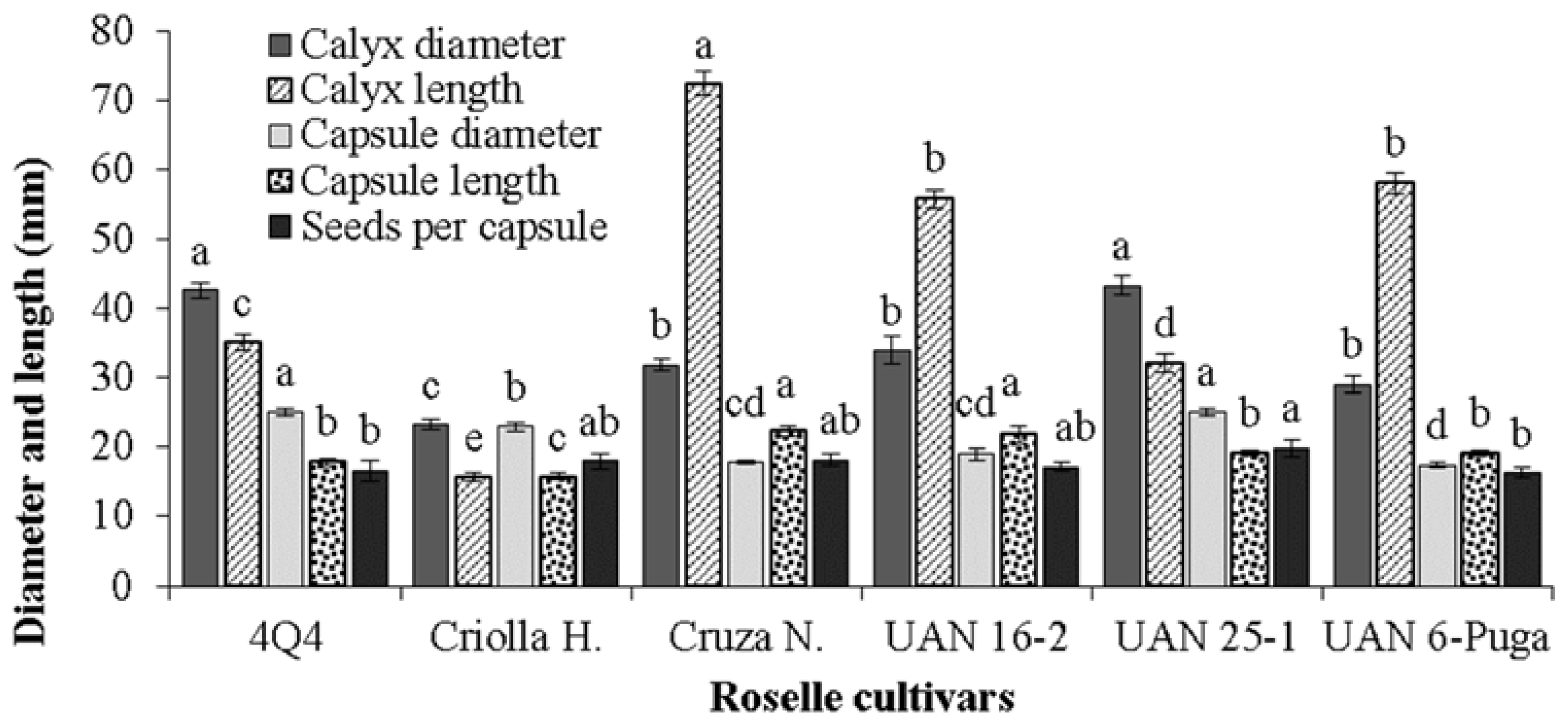
2.4. Roselle Calyx and Capsule Measurements
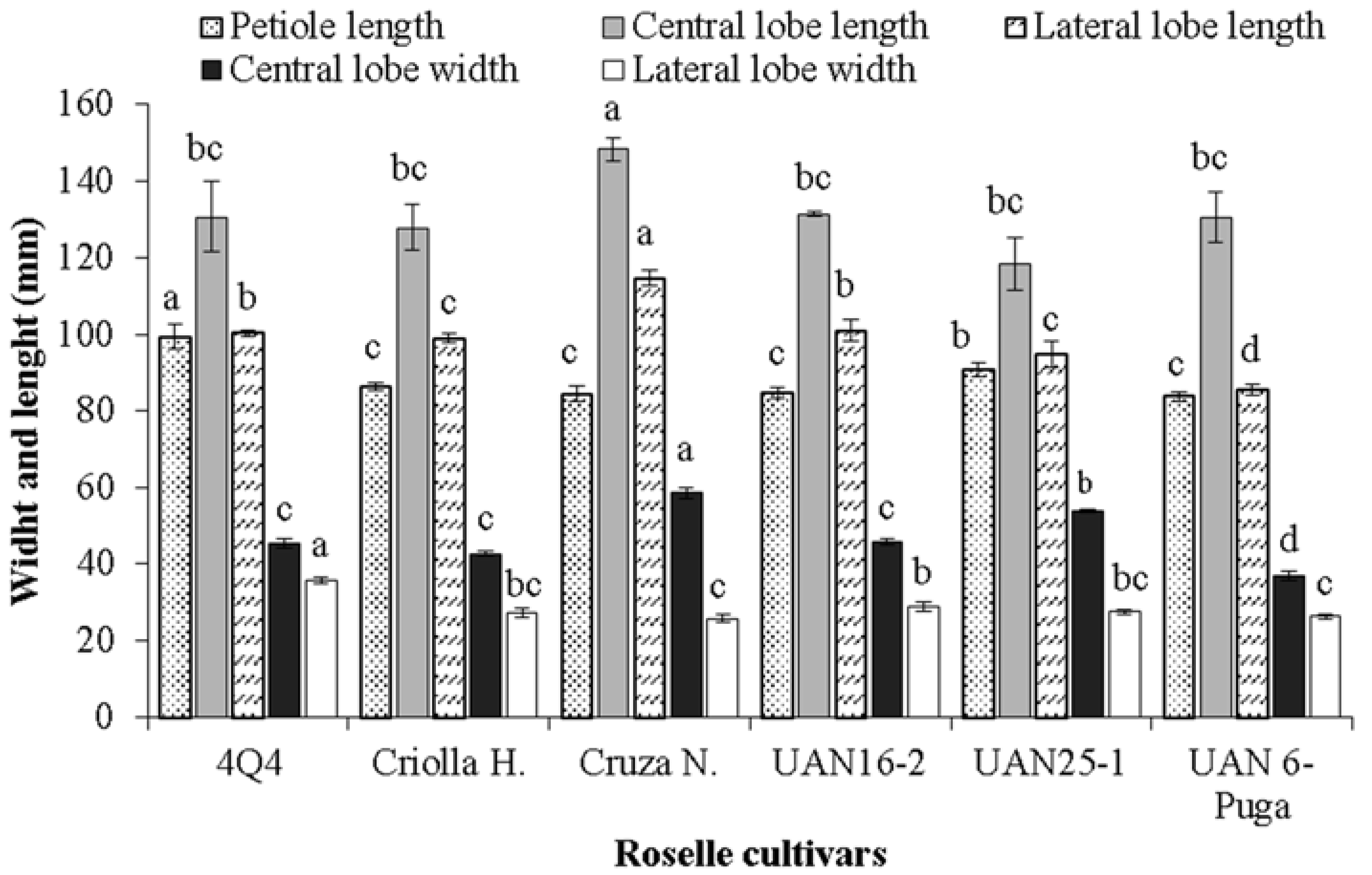
2.5. Leaf Size and Leaf Surface Area
2.6. Yield of Roselle Cultivars
2.7. Number of Days in the Roselle Development Cycle
2.8. Mineral Composition of Roselle Leaves
2.9. Pearson’s Correlation
3. Materials and Methods
3.1. Materials and General Conditions of Cultivation
3.2. Growth Rates
3.3. Morphological Parameters
3.4. Leaf Surface Area
3.5. Leaf Analysis
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sáyago-Ayerdi, S.G.; Velázquez-López, C.; Montalvo-González, E.; Goñi, I. By-product from decoction process of Hibiscus sabdariffa L. calyces as a source of polyphenols and dietary fiber. J. Sci. Food Agric. 2014, 94, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Morton, J.F. Roselle. In Fruits of Warm Climates; Morton Collectanea; University of Miami: Coral Gables, FL, USA, 1987; pp. 281–286. [Google Scholar]
- Ross, I.A. Hibiscus sabdariffa. Medicinal Plants of the World; Humana Press: Passaic, NJ, USA, 2003; pp. 267–275. [Google Scholar]
- Adanlawo, I.G.; Ajibade, V.A. Nutritive Value of the Two Varieties of Roselle (Hibiscus sabdariffa) Calyces Soaked with Wood Ash. Pak. J. Nutr. 2006, 5, 555–557. Available online: https://pdfs.semanticscholar.org/cdbb/c3da21abfadb05f04bcdaf098998fed77f04.pdf (accessed on 11 February 2020).
- Mahadevan, N.; Shivali, A.; Kamboj, P. Hibiscus sabdariffa Linn.—An overview. Nat. Prod. Radiance 2009, 8, 77–83. [Google Scholar]
- Atta, S.; Sarr, B.; Bakasso, Y.; Diallo, A.B.; Lona, I.; Saadou, M.; Glew, R.H. Roselle (Hibiscus sabdariffa L.) yield and yield components in response to nitrogen fertilization in niger. Indian J. Agric. Res. 2010, 44, 96–103. Available online: https://www.arccjournals.com/indianjournals.com (accessed on 11 February 2020).
- Atta, S.; Diallo, A.B.; Bakasso, Y.; Sarr, B.; Saadou, M.; Glew, R.H. Micro-element contents in roselle (Hibiscus sabdariffa L.) at different growth stages. Afr. J. Food Agric. Nutr. Dev. 2010, 10, 5. Available online: https://www.ajol.info/index.php/ajfand/article/view/57898 (accessed on 11 February 2020). [CrossRef] [Green Version]
- Garrow, J.; James, W.; Ralph, A. Human nutrition and dietetics. Clin. Nutr. 2000, 19, 141–142. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 2nd ed.; Sinauer Associates, Inc. Publishers: Sunderland, MA, USA, 1998. [Google Scholar]
- Fraga, C.G. Relevance, essentiality and toxicity of trace elements in human health. Mol. Asp. Med. 2005, 26, 235–244. [Google Scholar] [CrossRef]
- Solon-Biet, S.M.; Mitchell, S.J.; de Cabo, R.; Raubenheimer, D.; Le Couteur, D.; Simpson, S.J. Macronutrients and caloric intake in health and longevity. J. Endocrinol. 2015, 226, R17–R28. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-H.; Wang, C.-J. Chemopreventive Properties and Molecular Mechanisms of the Bioactive Compounds in Hibiscus Sabdariffa Linne. Curr. Med. Chem. 2011, 18, 1245–1254. [Google Scholar] [CrossRef]
- Chiu, C.-T.; Hsuan, S.-W.; Lin, H.-H.; Hsu, C.-C.; Chou, F.-P.; Chen, J.-H. Hibiscus sabdariffa Leaf Polyphenolic Extract Induces Human Melanoma Cell Death, Apoptosis and Autophagy. J. Food Sci. 2015, 80, H649–H658. [Google Scholar] [CrossRef] [PubMed]
- Gosain, S.; Ircchiaya, R.; Sharma, P.C.; Thareja, S.; Kalra, A.; Deep, A.; Bhardwaj, T.R. Hypolipidemic effect of ethanolic extract from the leaves of Hibiscus sabdariffa L. in hyperlipidemic rats. Acta Pol. Pharm. Drug Res. 2010, 67, 179–184. [Google Scholar]
- Bridle, P.; Timberlake, C. Anthocyanins as natural food colours—Selected aspects. Food Chem. 1997, 58, 103–109. [Google Scholar] [CrossRef]
- Ali, B.H.; Al-Wabel, N.; Blunden, G. Phytochemical, pharmacological and toxicological aspects of Hibiscus sabdariffa L.: A review. Phytother. Res. 2005, 19, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Hinojosa-Gómez, J.; San Martín-Hernández, C.; Heredia, J.B.; León-Félix, J.; Osuna-Enciso, T.; y Muy-Rangel, M.D. Anthocyanin induction by drought stress in the calyx of roselle cultivars. Molecules 2020, 25, 1555. [Google Scholar] [CrossRef]
- Fernández-Arroyo, S.; Rodríguez-Medina, I.C.; Beltrán-Debón, R.; Pasini, F.; Joven, J.; Micol, V.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Quantification of the polyphenolic fraction and In Vitro antioxidant and in vivo anti-hyperlipemic activities of Hibiscus sabdariffa aqueous extract. Food Res. Int. 2011, 44, 1490–1495. [Google Scholar] [CrossRef]
- Lin, H.-H.; Chan, K.-C.; Sheu, J.-Y.; Hsuan, S.-W.; Wang, C.-J.; Chen, J.-H. Hibiscus sabdariffa leaf induces apoptosis of human prostate cancer cells in vitro and in vivo. Food Chem. 2012, 132, 880–891. [Google Scholar] [CrossRef]
- Hopkins, A.L.; Lamm, M.G.; Funk, J.L.; Ritenbaugh, C. Hibiscus sabdariffa L. in the treatment of hypertension and hyperlipidemia: A comprehensive review of animal and human studies. Fitoterapia 2013, 85, 84–94. [Google Scholar] [CrossRef] [Green Version]
- Sindi, H.A.; Marshall, L.J.; Morgan, M.R. Comparative chemical and biochemical analysis of extracts of Hibiscus sabdariffa. Food Chem. 2014, 164, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Khare, C. Hibiscus sabdariffa Linn. Indian Med. Plants 2007, 8, 1. [Google Scholar] [CrossRef]
- Lim, S.-H.; Song, J.-H.; Kim, D.-H.; Kim, J.K.; Lee, J.-Y.; Kim, Y.-M.; Ha, S.-H. Activation of anthocyanin biosynthesis by expression of the radish R2R3-MYB transcription factor gene RsMYB. Plant Cell Rep. 2015, 35, 641–653. [Google Scholar] [CrossRef]
- Guha, A.; Sengupta, D.; Rasineni, G.K.; Reddy, A.R. An integrated diagnostic approach to understand drought tolerance in mulberry (Morus indica L.). Flora. Morphol. Distrib. Funct. Ecol. Plants 2009, 205, 144–151. [Google Scholar] [CrossRef]
- Faruq, G.; Rahman, M.; Zabed, H.; Latif, A. Assessment of Genetic Variation in Different Kenaf (Hibiscus cannabinus) Genotypes using Morpho-agronomic Traits and RAPD Markers. Int. J. Agric. Biol. 2015, 17, 507–514. [Google Scholar] [CrossRef]
- San-Martín-Hernández, C.; Trejo-Téllez, L.I.; Gómez-Merino, F.C.; Volke-Haller, V.H.; Escalante-Estrada, J.A.; Prometeo, S.C.; Crescenciano, S.V. Nitrogen and potassium nutrition differentially affect tomato biomass and growth. Interciencia 2016, 41, 60–66. Available online: http://www.redalyc.org/articulo.oa?id=33943362010 (accessed on 12 February 2020).
- Antúnez, I.; Retamosa, E.C.; Villar, R. Relative growth rate in phylogenetically related deciduous and evergreen woody species. Oecologia. Oecologia 2001, 128, 172–180. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Akanbi, W.B.; Olaniyan, A.B.; Togun, A.O.; Olaniran, O.A.; Biology, E. The Effect of Organic and Inorganic Fertilizer on Growth, Calyx Yield And Quality of Roselle (Hibiscus Sabdariffa L.). Am. Eurasian J. Sustain. Agric. 2009, 3, 652–657. [Google Scholar]
- Atta, S.; Diallo, A.; Sarr, B.; Bakasso, Y.; Saadou, M.; Glew, R. Variation in macro-elements and protein contents of roselle (Hibiscus sabdariffa L.) from Niger. Afr. J. Food Agric. Nutr. Dev. 2010, 10, 2707–2718. [Google Scholar] [CrossRef] [Green Version]
- Abo-Baker, A.; Mostafa, G.G. Effect of Bio-and Chemical Fertilizers on Growth, Sepals Yield and Chemical Composition of Hibiscus sabdariffa at New Reclaimed Soil of South Valley Area. Asian J. Crop Sci. 2010, 3, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Abbas, M.K.; Ali, A.S. Effect of Foliar Application of NPK on Some Growth Characters of Two Cultivars of Roselle (Hibiscus sabdariffa L.). Am. J. Plant Physiol. 2011, 6, 220–227. [Google Scholar] [CrossRef] [Green Version]
- Gad, N.; Badran, M. Effect of Cobalt and Nickel on Plant Growth, Yield and Flavonoids Content of Hibiscus sabdariffa L. Aust. J. Basic Appl. Sci. 2007, 1, 73–78. Available online: https://pdfs.semanticscholar.org/6cfd/bdfbae91e2defc5f0677e1b006fa04dd127c.pdf (accessed on 12 February 2020).
- Sánchez-Rodríguez, E.; Moreno, D.A.; Ferreres, F.; Rubio-Wilhelmi, M.D.M.; Ruiz, J.M. Differential responses of five cherry tomato varieties to water stress: Changes on phenolic metabolites and related enzymes. Phytochemistry 2011, 72, 723–729. [Google Scholar] [CrossRef]
- Cruz, N.A.; Forestales, A.Y.P.I.N.D.I.; Solano, J.P.L.; Chapingo, U.A. Morphological Characterization of An Ethnographic Sample of Roselle (Hibiscus sabdariffa L.). Rev. Chapingo Ser. Hortic. 2013, XIX, 85–98. [Google Scholar] [CrossRef]
- Terán, Z.; Soto, F. Evaluación de densidades de plantación en el cultivo de la jamaica (Hibiscus sabdariffa L.). Cultiv. Trop. 2004, 25, 67–69. [Google Scholar] [CrossRef]
- Ramírez-Cortés, B.; De Jesús Caro-Velarde, F.; Valdivia-Reynoso, M.G.; Ramírez-Lozano, M.H.; Machuca-Sánchez, M.L. Cambios en tamaño y características químicas de cálices de jamaica (Hibiscus sabdariffa L.) durante su maduración. Rev. Chapingo Ser. Hortic. 2011, 17, 19–31. [Google Scholar] [CrossRef]
- Villatoro, S.G.H.; Reyes, W.A.D.L.C.; Ruano-Solís, H.H.; Cano-Castillo, L.E. Caracterización de trece genotipos de rosa de jamaica Hibiscus sabdariffa en Guatemala. Agron. Mesoam. 2008, 20, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Wong, P.; Yusof, S.; Ghazali, H.; Man, Y.C. Physico-chemical characteristics of roselle (Hibiscus sabdariffa L.). Nutr. Food Sci. 2002, 32, 68–73. [Google Scholar] [CrossRef]
- Wright, I.J.; Westoby, M. Cross-species relationships between seedling relative growth rate, nitrogen productivity and root v.s. leaf function in 28 Australian woody species. Funct. Ecol. 2000, 14, 97–107. [Google Scholar] [CrossRef]
- Shukla, M.S.; Dube, K.G. Effect of gamma rays and ems on yield parameters in Hibiscus sabdariffa. Int. J. Bot. Stud. 2017, 2, 180–187. Available online: http://www.botanyjournals.com/archives/2017/vol2/issue6/2-6-53 (accessed on 13 February 2020).
- De Castro, N.E.A.; Pinto, J.E.B.P.; Cardoso, M.D.G.; de Morais, A.R.; Bertolucci, S.K.V.; da Silva, F.G.; Filho, N.D. Planting time for maximization of yield of vinegar plant calyx (Hibiscus sabdariffa L.). Ciên. e Agrotecnol. 2004, 28, 542–551. [Google Scholar] [CrossRef] [Green Version]
- Gabarra, A.G. ESPAÑA: Cantidades Diarias Recomendadas (CDR). Nutr Hosp. 2006, 21, 437–447. Available online: http://scielo.isciii.es/pdf/nh/v21n4/revision1.pdf (accessed on 17 March 2021).
- Musa, A. Effect of Plant Leaf Positions on Some Micronutrients, Anti-nutrients and Toxic Substances in Telfairia occidentalis at the Vegetative Phase. Am. J. Exp. Agric. 2012, 2, 219–232. [Google Scholar] [CrossRef]
- Musa, A. Levels of Phytotoxins and Nutrients in Hibiscus sabdariffa as Influenced by Freezing Storage. Br. J. Appl. Sci. Technol. 2013, 3, 799–812. [Google Scholar] [CrossRef]
- Hinojosa-Gómez, J.; Martin-Hernández, C.S.; Heredia, J.B.; León-Félix, J.; Osuna-Enciso, T.; Muy-Rangel, M.D. Roselle (Hibiscus sabdariffa L.) cultivars calyx produced hydroponically: Physicochemical and nutritional quality. Chil. J. Agric. Res. 2018, 78, 478–485. [Google Scholar] [CrossRef]
- Ribas, F.; Cabello, M.; Moreno, M.; Moreno, A.; López-Bellido, L. Influencia del riego y de la aplicación de potasio en la producción del melón (Cucumis melo L.). II: Calidad. Span. J. Agric. Res. 2003, 1, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela, J.; Del Río, A.; Sánchez, A.; Lópezcantarero, Y.; Romero, L. Influencia de la fertirrigación N, P, K sobre diversos nutrientes totales y solubles, su evolución temporal. Suelo y Planta 1994, 2, 735–745. [Google Scholar]
- Ho, L.; Adams, P. Nutrient uptake and distribution in relation to crop quality. Acta Hortic. 1995, 33–44. [Google Scholar] [CrossRef]
- Montoya, R.B.; Spinola, A.G.; García, P.S.; Paredes, D.G. Demanda de potasio del tomate tipo saladette. Terra Latinoam. 2002, 20, 391–399. [Google Scholar]
- Cakmak, I.; Kirkby, E.A. Role of magnesium in carbon partitioning and alleviating photooxidative damage. Physiol. Plant. 2008, 133, 692–704. [Google Scholar] [CrossRef] [Green Version]
- Fischer, S.; Wilckens, R.; Vidal, I.; Astete, P.; Maier, J. Respuesta de la achicoria (Cichorium intybus L.) a la aplicación de magnesio. Chil. J. Agric. Anim. Sci. 2016, 32, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Galvez, J. Reparto de materia seca como factor determinante de la producción de las hortalizas de fruto cultivadas en invernadero. Curr. Agric. Sci. Technol. 2005, 11, 5–11. [Google Scholar]
- Rodríguez-Yzquierdo, G.A.; Pradenas-Aguila, H.E.; Basso-de-Figuera, C.A.; Barrios-García, M.; León-Pacheco, R.I.; Pérez-Macias, M. Effect of doses of nitrogen in the agronomy and physiology of yellow passion fruit. Agron. Mesoam. 2020, 31, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Steiner, A.A. A universal method for preparing nutrient solutions of a certain desired composition. Plant Soil 1961, 15, 134–154. [Google Scholar] [CrossRef]
- Parsons, I.T.; Hunt, R. Plant Growth Analysis: A Program for the Fitting of Lengthy Series of Data by the Method of B-splines*. Ann. Bot. 1981, 48, 341–352. [Google Scholar] [CrossRef]
- Díaz-Pérez, J.C.; Muy-Rangel, M.D.; Mascorro, A.G. Fruit size and stage of ripeness affect postharvest water loss in bell pepper fruit (Capsicum annuum L.). J. Sci. Food Agric. 2006, 87, 68–73. [Google Scholar] [CrossRef]
- Bremner, J.M. Nitrogen-Total. In Methods of Soil Analysis, Part 3 Chemical Methods; Sparks, D.L., Ed.; SSSA: Madison, WI, USA, 1996; pp. 1085–1122. [Google Scholar] [CrossRef]
- AOAC. Official methods of analysis. In Association of Official Analytical Chemists (AOAC), 16th ed.; AOAC: Washington, DC, USA, 1998. [Google Scholar]
- Volke, H.V.; Turrent, F.A.; Castillo, M.A. Diseños de Tratamientos y Estimación de Funciones de Respuesta en la Investigación Agrícola; Colegio de Posgraduados: Montecillos, Mexico, 2005. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Rate | Cultivar | Model | p-Value | |
|---|---|---|---|---|
| RGRC * | UAN 16-2 | Y = −0.64686 + 0.2557DAT − 0.0051 DAT2 + 0.0000260DAT3 | <0.0001 | |
| MSE a = 0.22750 | R2 = 0.8573 | |||
| 4Q4 | Y = −1.16136 + 0.21702DAT − 0.00327DAT2 + 0.00001239DAT3 | <0.0001 | ||
| MSE = 0.12680 | R2 = 0.9148 | |||
| Criolla H. | Y = 0.05443 + 0.16121DAT − 0.00235DAT2 + 0.00000772DAT3 | <0.0001 | ||
| MSE = 0.23711 | R2 = 0.98435 | |||
| UAN 25-1 | Y = −0.12310 + 0.19722DAT − 0.00382DAT2 + 0.00001866DAT3 | <0.0001 | ||
| MSE = 0.25238 | R2 = 0.8141 | |||
| Cruza N. | Y = −0.21955 + 0.18356DAT − 0.00333DAT2 + 0.00001518DAT3 | <0.0001 | ||
| MSE = 0.09309 | R2 = 0.9186 | |||
| UAN 6 PUGA | Y = 0.35909 + 0.13533DAT − 0.00286DAT2 + 0.00001467DAT3 | <0.0001 | ||
| MSE = 0.12374 | R2 = 0.8537 | |||
| RGRSD * | UAN 16-2 | Y = 0.13227 + 0.00399DAT − 0.00007546DAT2 | <0.0001 | |
| MSE = 0.00166 | R2 = 0.7774 | |||
| 4Q4 | Y = 0.20901 + 0.00325DAT − 0.00007679DAT2 | <0.0001 | ||
| MSE = 0.00175 | R2 = 0.8313 | |||
| Criolla H. | Y = 0.14337 + 0.00546DAT − 0.00009423DAT2 | <0.0001 | ||
| MSE = 0.00197 | R2 = 0.7722 | |||
| UAN 25-1 | Y = 0.17113 + 0.00450DAT − 0.00008664DAT2 | <0.0001 | ||
| MSE = 0.00096 | R2 = 0.8884 | |||
| Cruza N. | Y = 0.18355 + 0.00233DAT − 0.00006217DAT2 | <0.0001 | ||
| MSE = 0.00134 | R2 = 0.8419 | |||
| UAN 6 PUGA | Y = 0.13392 + 0.00173DAT − 0.00004531DAT2 | <0.0001 | ||
| MSE = 0.00102 | R2 = 0.7764 | |||
| Cultivar | Num. of Productive Branches | Height (cm) | Stem Diameter (mm) | Foliar Surface Area (cm2) |
|---|---|---|---|---|
| 4Q4 | 8.1 ± 0.5 a b * | 200.7 ± 13.7 b | 17.2 ± 1.0 a | 89.9 ± 1.3 a |
| Criolla H. | 14.9 ± 1.2 a | 245.4 ± 22.4 a | 15.9 ± 1.7 ab | 86.9 ± 1.6 c |
| Cruza N. | 5.9 ± 0.5 c | 176.5 ± 10.8 c | 14.3 ± 1.9 bc | 88 ± 1.5 a |
| UAN 16-2 | 7.7 ± 0.9 b | 173.2 ± 23.8 c | 13.5 ± 1.6 c | 91.3 ± 0.9 a |
| UAN 25-1 | 8.2 ± 0.7 b | 173.8 ± 15.1 c | 15.3 ± 1.2 b | 77.1 ± 1.9 b |
| UAN 6-Puga | 4.8 ± 0.3 c | 129.8 ± 17.1 d | 11.5 ± 1.1 d | 76.4 ± 2.2 b |
| Cultivar | Calyces Harvested per Plant | Fresh Weight Calyx | Fresh Weight Yield Calyx | Dry Weight Calyx Yield |
|---|---|---|---|---|
| kg ha−1 | ||||
| 4Q4 | 96.8 ± 21.1 a bc * | 0.63 ± 0.04 c | 5551 ± 378 c | 971.62 ± 80.7 b |
| Criolla H. | 173.5 ± 39.3 a | 0.91 ± 0.05 b | 7900 ± 410 b | 989.67 ± 50.9 b |
| Cruza N. | 116.4 ± 39.8 ab | 1.47 ± 0.11 a | 12,855 ± 977 a | 1872.43 ± 119.3 a |
| UAN 16-2 | 90.4 ± 29.5 bc | 0.77 ± 0.04 bc | 6720 ± 343 bc | 910.51 ± 29.8 b |
| UAN 25-1 | 120.3 ± 23.9 ab | 0.88 ± 0.07 b | 7713 ± 614 b | 946.19 ± 72.8 b |
| UAN 6-Puga | 49.0 ± 12.13 c | 0.24 ± 0.02 d | 2077.9 ± 137.7 d | 263.9 ± 27.4 c |
| Cultivar | Days to Flowering | Days to Harvest |
|---|---|---|
| 4Q4 | 48.7 ± 1.3 a ab * | 154.1 ± 1.3 bc |
| Criolla H. | 51.2 ± 2.1 a | 156.4 ± 0.4 ab |
| Cruza N. | 46.7 ± 1.5 bc | 148.3 ± 2.2 d |
| UAN 16-2 | 43.1 ± 1.1 d | 151.9 ± 0.5 c |
| UAN 25-1 | 44.9 ± 1.1 cd | 156.9 ± 0.6 a |
| UAN 6-Puga | 42.3 ± 0.7 d | 157.9 ± 1.1 a |
| 4Q4 | Criolla H. | Cruza N. | UAN 16-2 | UAN 25-1 | UAN 6-Puga | RDI a | |
|---|---|---|---|---|---|---|---|
| ------------------------------------------------- mg kg−1 --------------------------------------------------- | mg d−1 | ||||||
| N | 36,594 ± 262 b bc * | 38,502 ± 694 a | 31,158 ± 874 d | 36,856 ± 312 b | 36,808 ± 652 b | 35,320 ± 582 c | - |
| P | 10,663 ± 67.9 c | 11,747 ± 43.3 b | 11,929 ± 91.5 a | 11,832 ± 64.7 ab | 11,702 ± 32.6 b | 10,663 ± 18 c | 664 |
| K | 13,397 ± 104 b | 13,259 ± 233 b | 18,171 ± 546 a | 11,365 ± 542 c | 10,848 ± 94 c | 7554 ± 302 d | 4700 |
| Ca | 18,364 ± 267 a | 10,648 ± 255 d | 10,527 ± 346 d | 13,291 ± 90 c | 12,716 ± 227 c | 14,346 ± 373 b | 900 |
| Mg | 3813 ± 103.8 c | 3819 ± 65.9 c | 4722 ± 110.8 a | 4107 ± 79 b | 4739 ± 183 a | 3820 ± 101 c | 248 |
| Na | 76.7 ± 2.3 c | 86.0 ± 3.0 b | 88.3 ± 1.2 b | 97.6 ± 2.0 a | 75.6 ± 2.0 c | 52.6 ± 3.9 d | 1500 |
| Fe | 130.8 ± 9.3 bc | 108.2 ± 4.3 c | 154.1 ± 14.2 ab | 147.0 ± 17.2 b | 175.5 ± 8.4 a | 150.4 ± 11.7 ab | 17 |
| Mn | 309.4 ± 11.8 bc | 289.1 ± 11.4 c | 355.0 ± 17.6 ab | 283.6 ± 11.5 c | 323.1 ± 16.1 abc | 364.6 ± 39.7 a | 2 |
| Zn | 207.7 ± 8.2 c | 289.6 ± 10.8 b | 205.8 ± 10.8 c | 300.9 ± 16.1 b | 314.3 ± 8.6 b | 363.3 ± 11.3 a | 10 |
| Cu | 8.3 ± 0.6 a | 8.3 ± 1.0 a | 5.5 ± 0.3 b | 9.3 ± 0.8 a | 8.2 ± 1.3 a | 8.3 ± 0.3 a | 1 |
| Total | 83,565.6 | 78,756.3 | 77,316.2 | 78,290.8 | 77,711.4 | 72,643.4 | |
| Flower Length (mm) | Calyx Length (mm) | Central Lobe Length (mm) | Lateral Lobe Length (mm) | Calyces Harvested per Plant | Fresh Weight Yield Calyx (kg ha−1) | Dry Weight Calyx Yield (kg ha−1) | |
|---|---|---|---|---|---|---|---|
| K | 0.121 | 0.407 | 0.942 * | 0.483 | 0.894 * | 0.894 * | 0.96 * |
| Mg | 0.456 | 0.522 | 0.459 | 0.842 * | 0.676 | 0.676 | 0.606 |
| N | 0.836 * | 0.864 * | 0.53 | 0.653 | 0.501 | 0.501 | 0.579 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hinojosa-Gómez, J.; San Martín-Hernández, C.; Osuna-Enciso, T.; Heredia, J.B.; León-Félix, J.; Muy-Rangel, M.D. Agronomic Behavior of Mexican Roselle Cultivars Produced under Protected Agricultural Conditions. Plants 2022, 11, 2767. https://doi.org/10.3390/plants11202767
Hinojosa-Gómez J, San Martín-Hernández C, Osuna-Enciso T, Heredia JB, León-Félix J, Muy-Rangel MD. Agronomic Behavior of Mexican Roselle Cultivars Produced under Protected Agricultural Conditions. Plants. 2022; 11(20):2767. https://doi.org/10.3390/plants11202767
Chicago/Turabian StyleHinojosa-Gómez, Jeny, César San Martín-Hernández, Tomás Osuna-Enciso, José B. Heredia, Josefina León-Félix, and María D. Muy-Rangel. 2022. "Agronomic Behavior of Mexican Roselle Cultivars Produced under Protected Agricultural Conditions" Plants 11, no. 20: 2767. https://doi.org/10.3390/plants11202767