


Edible Wild Vegetables Urtica dioica L. and Aegopodium podagraria L.–Antioxidants Affected by Processing

Abstract
:1. Introduction
2. Results
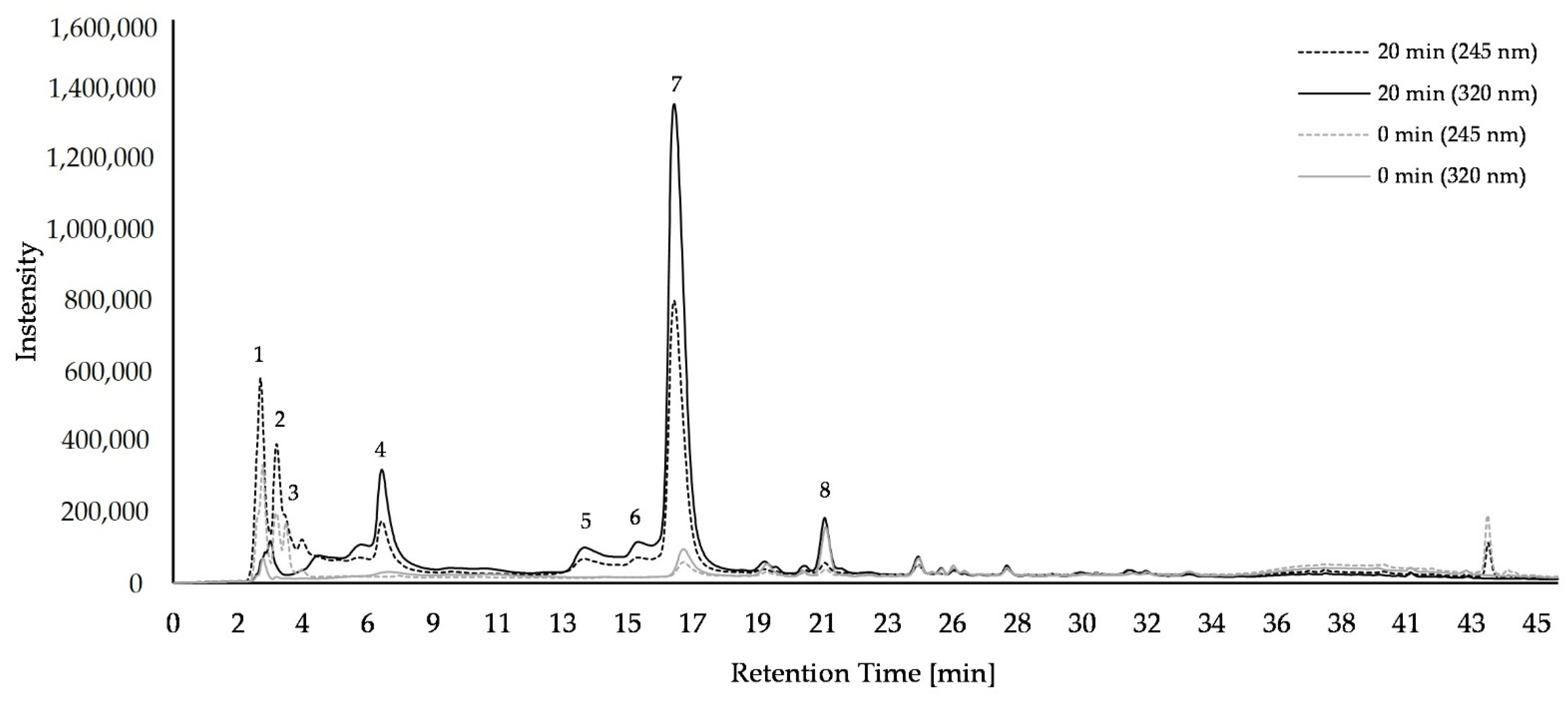
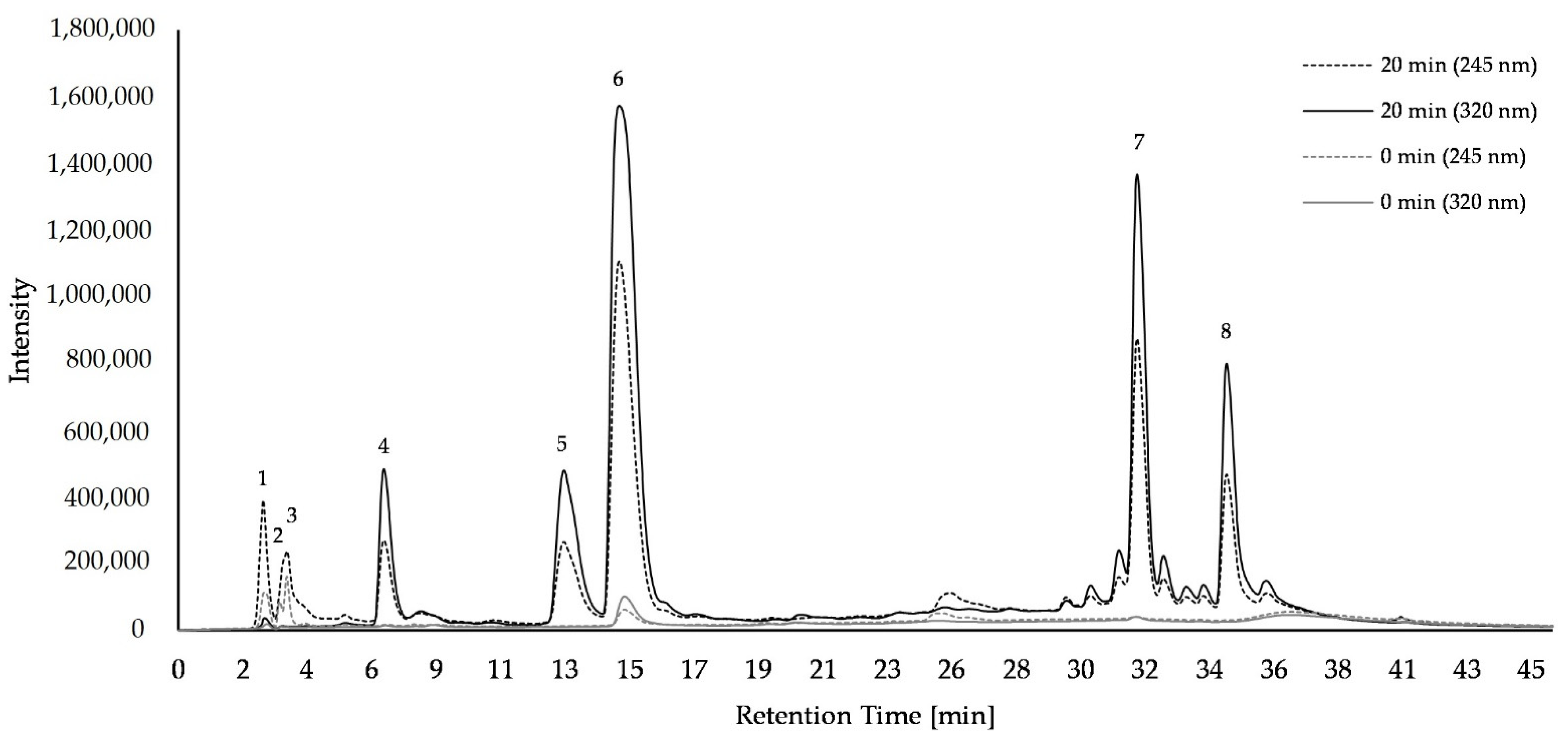
2.1. Composition of Bioactice Compounds
2.2. Minerals
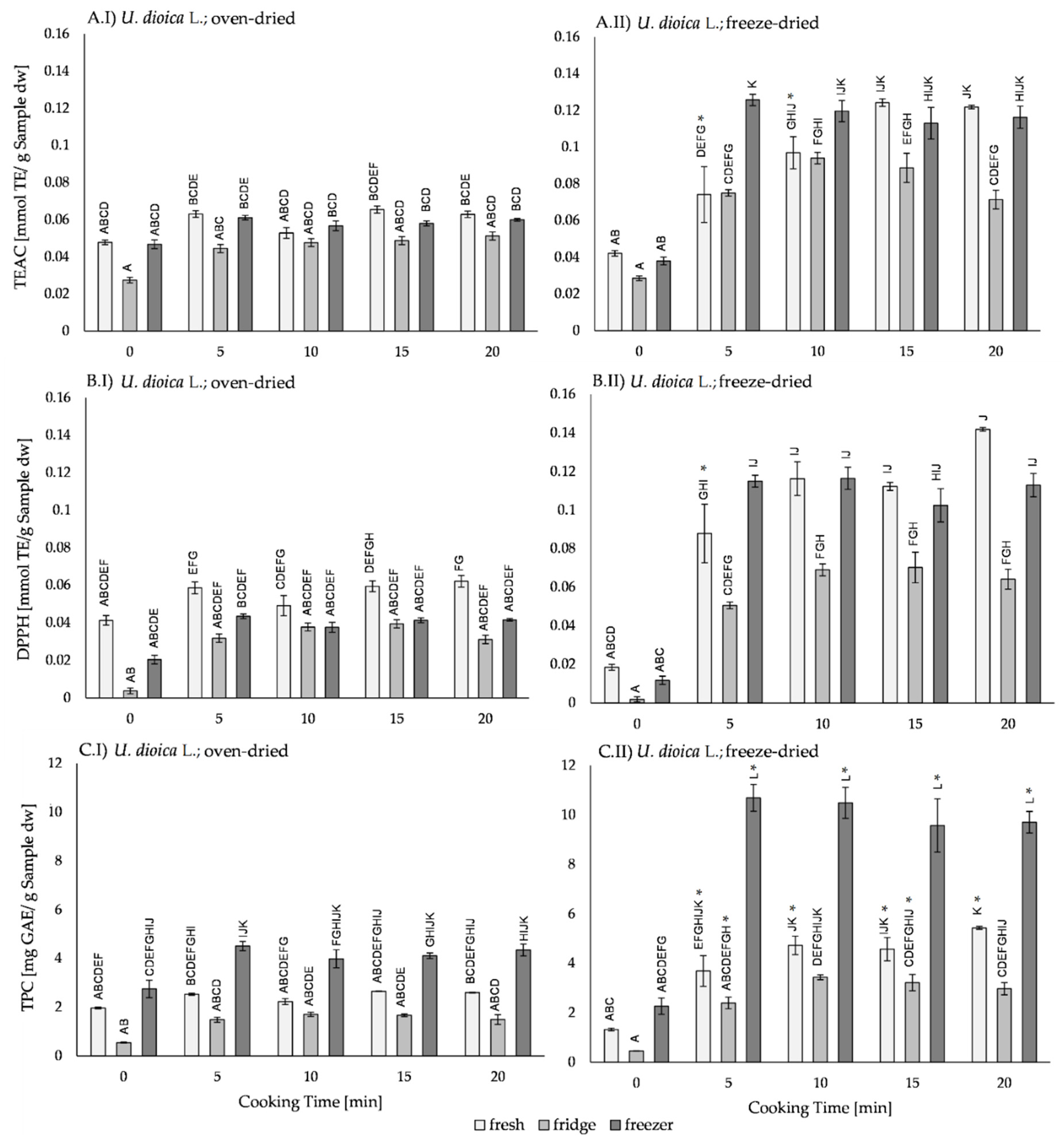
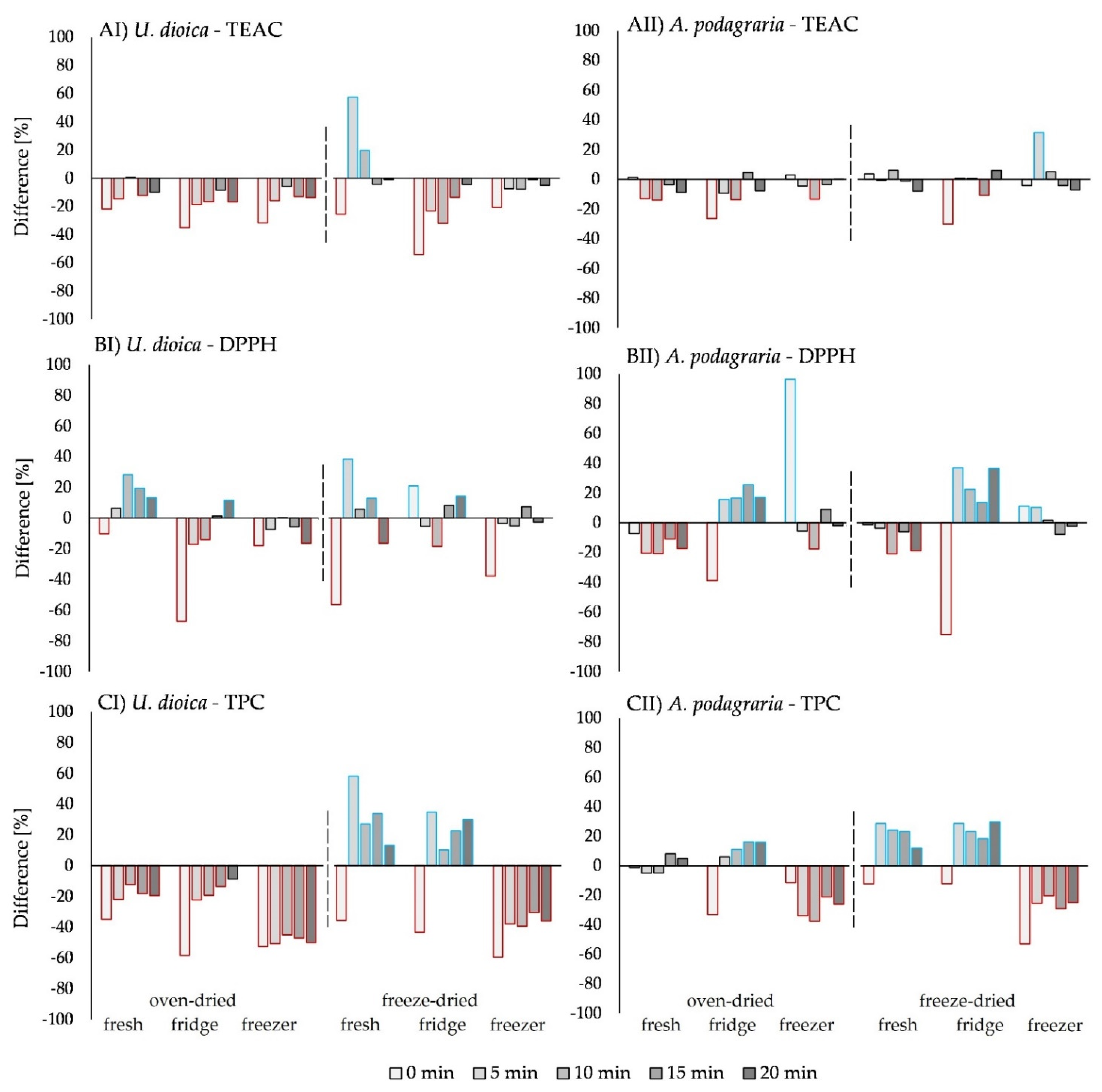
2.3. Effects of Processing on The Antioxidant Activity in U. dioica L.
2.3.1. Effect of The Drying Method on the Antioxidant Activity of U. dioica L.
2.3.2. Effect of Thermal Processing on The Antioxidant Activity of U. dioica L.
2.3.3. Effect of Storage on the Antioxidant Activity of U. dioica L.
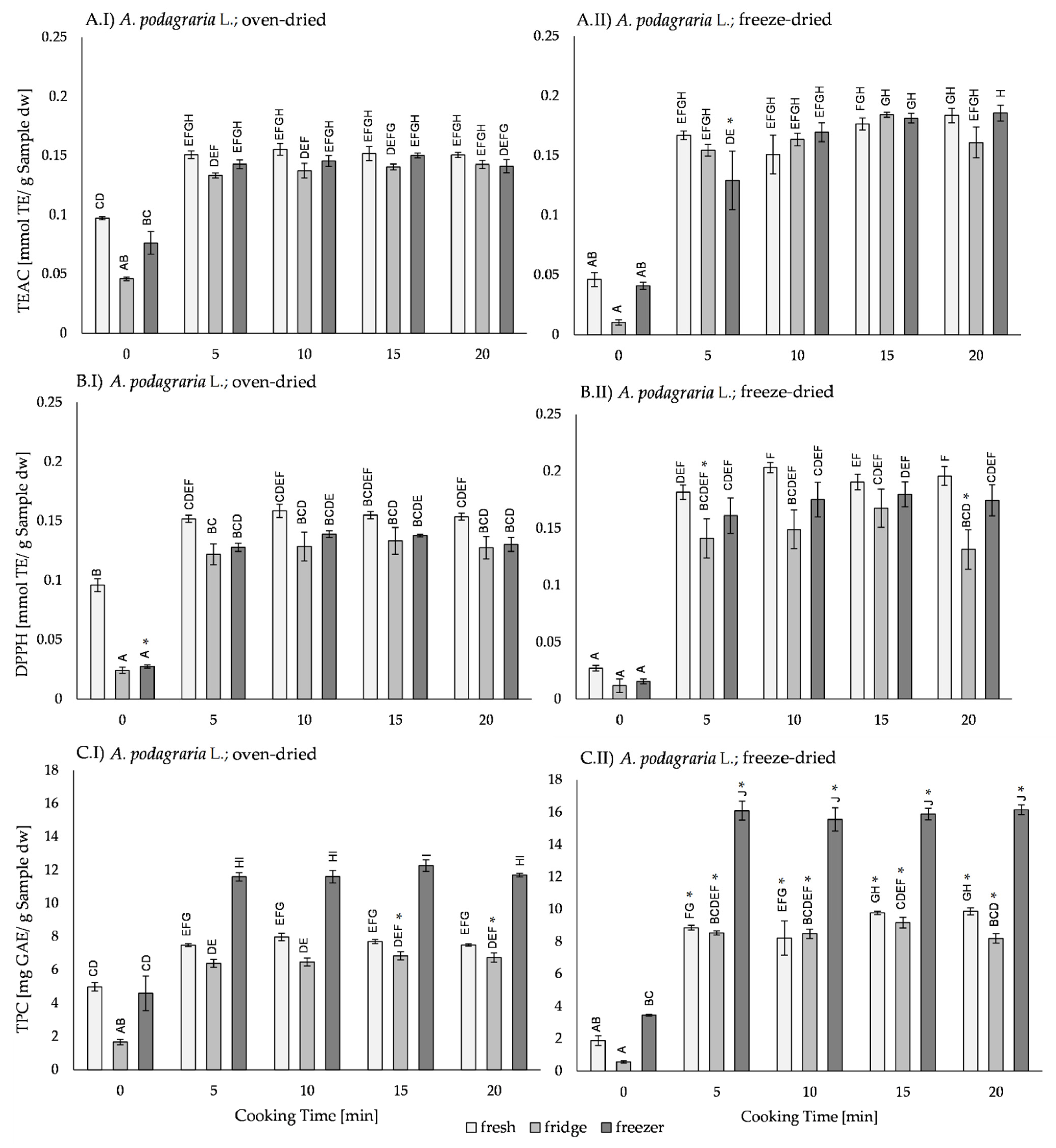
2.4. Effects of Processing on the Antioxidant Activity in A. podagraria L.
2.4.1. Effect of Drying on the Antioxidant Activity of A. podagraria L.
2.4.2. Effect of Thermal Processing on the Antioxidant Activity of A. podagraria L.
2.4.3. Effect of Storage on the Antioxidant Activity of A. podagraria L.
2.5. Synergistic and Antagonistic Effects on the Antioxidant Activity by Exogenous 5-Caffeoylquinic Addition
3. Discussion
3.1. Influence of Drying Methods on Antioxidant Activity
3.2. Thermal Processing
3.3. Storage
3.4. Addition of Exogenous 5-caffeoylquinic acid
3.5. Interactions of Minerals Contained in the Plant Matrix
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material
4.3. Procesing and Storage
4.4. Photometric Measurements
4.4.1. Total Phenolic Content (TPC)
4.4.2. Trolox Equivalent Antioxidant Capacity (TEAC)
4.4.3. 2,2-diphenyl-1-picrylhydrazyl (DPPH)
4.5. HPLC
4.6. Mineral Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olsen, C. The Ecology of Urtica dioica. J. Ecol. 1921, 9, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Jakubczyk, K.; Janda, K.; Styburski, D.; Łukomska, A. Goutweed (Aegopodium podagraria L.)—Botanical Characteristics and Prohealthy Properties. Postepy Hig. Med. Dosw. 2020, 74, 28–35. [Google Scholar] [CrossRef]
- Sarkar, P.; Bosneaga, E.; Auer, M. Plant Cell Walls throughout Evolution: Towards a Molecular Understanding of Their Design Principles. J. Exp. Bot. 2009, 60, 3615–3635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, L.E.; Cook, M.E.; Busse, J.S. The Origin of Plants: Body Plan Changes Contributing to a Major Evolutionary Radiation. Proc. Natl. Acad. Sci. USA 2000, 97, 4535–4540. [Google Scholar] [CrossRef] [Green Version]
- Falster, D.S.; Westoby, M. Plant Height and Evolutionary Games. Trends Ecol. Evol. 2003, 18, 337–343. [Google Scholar] [CrossRef]
- Chon, S.-U. Total Polyphenols and Bioactivity of Seeds and Sprouts in Several Legumes. CPD 2013, 19, 6112–6124. [Google Scholar] [CrossRef]
- Weidinger, A.; Kozlov, A. Biological Activities of Reactive Oxygen and Nitrogen Species: Oxidative Stress versus Signal Transduction. Biomolecules 2015, 5, 472–484. [Google Scholar] [CrossRef] [Green Version]
- Banjarnahor, S.D.S.; Artanti, N. Antioxidant Properties of Flavonoids. Med. J. Indones. 2015, 23, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Hartl, A.; Vogl, C.R. Dry Matter and Fiber Yields, and the Fiber Characteristics of Five Nettle Clones (Urtica dioica L.) Organically Grown in Austria for Potential Textile Use. Am. J. Altern. Agric. 2002, 17, 195–200. [Google Scholar] [CrossRef]
- Vogl, C.R.; Hartl, A. Production and Processing of Organically Grown Fiber Nettle (Urtica dioica L.) and Its Potential Use in the Natural Textile Industry: A Review. Am. J. Altern. Agric. 2003, 18, 119–128. [Google Scholar] [CrossRef]
- Stoner, G.D.; Wang, L.-S.; Casto, B.C. Laboratory and Clinical Studies of Cancer Chemoprevention by Antioxidants in Berries. Carcinogenesis 2008, 29, 1665–1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollman, P.C.H.; Cassidy, A.; Comte, B.; Heinonen, M.; Richelle, M.; Richling, E.; Serafini, M.; Scalbert, A.; Sies, H.; Vidry, S. The Biological Relevance of Direct Antioxidant Effects of Polyphenols for Cardiovascular Health in Humans Is Not Established. J. Nutr. 2011, 141, 989S–1009S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahidi, F.; Yeo, J. Insoluble-Bound Phenolics in Food. Molecules 2016, 21, 1216. [Google Scholar] [CrossRef] [PubMed]
- Chua, L.Y.W.; Chong, C.H.; Chua, B.L.; Figiel, A. Influence of Drying Methods on the Antibacterial, Antioxidant and Essential Oil Volatile Composition of Herbs: A Review. Food Bioprocess Technol. 2019, 12, 450–476. [Google Scholar] [CrossRef]
- Ciurzyńska, A.; Lenart, A. Freeze-Drying—Application in Food Processing and Biotechnology—A Review. Pol. J. Food Nutr. Sci. 2011, 61, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Kontoghiorghes, G.J.; Kolnagou, A.; Kontoghiorghe, C.N.; Mourouzidis, L.; Timoshnikov, V.A.; Polyakov, N.E. Trying to Solve the Puzzle of the Interaction of Ascorbic Acid and Iron: Redox, Chelation and Therapeutic Implications. Medicines 2020, 7, 45. [Google Scholar] [CrossRef]
- Engelhardt, L.; Pöhnl, T.; Neugart, S. Interactions of Ascorbic Acid, 5-Caffeoylquinic Acid, and Quercetin-3-Rutinoside in the Presence and Absence of Iron during Thermal Processing and the Influence on Antioxidant Activity. Molecules 2021, 26, 7698. [Google Scholar] [CrossRef]
- Bourdoux, S.; Li, D.; Rajkovic, A.; Devlieghere, F.; Uyttendaele, M. Performance of Drying Technologies to Ensure Microbial Safety of Dried Fruits and Vegetables. Compr. Rev. Food Sci. Food Saf. 2016, 15, 1056–1066. [Google Scholar] [CrossRef]
- Garcìa, L.M.; Ceccanti, C.; Negro, C.; De Bellis, L.; Incrocci, L.; Pardossi, A.; Guidi, L. Effect of Drying Methods on Phenolic Compounds and Antioxidant Activity of Urtica dioica L. Leaves. Horticulturae 2021, 7, 10. [Google Scholar] [CrossRef]
- Chávez-Reyes, Y.; Dorantes-Alvarez, L.; Arrieta-Baez, D.; Osorio-Esquivel, O.; Ortiz-Moreno, A. Polyphenol Oxidase Inactivation by Microwave Oven and Its Effect on Phenolic Profile of Loquat (Eriobotrya Japonica) Fruit. FNS 2013, 04, 87–94. [Google Scholar] [CrossRef]
- Duan, X.; Yang, X.; Ren, G.; Pang, Y.; Liu, L.; Liu, Y. Technical Aspects in Freeze-Drying of Foods. Dry. Technol. 2016, 34, 1271–1285. [Google Scholar] [CrossRef]
- Śledź, M.; Nowacka, M.; Wiktor, A.; Witrowa-Rajchert, D. Selected Chemical and Physico-Chemical Properties of Microwave-Convective Dried Herbs. Food Bioprod. Processing 2013, 91, 421–428. [Google Scholar] [CrossRef]
- Mirhosseini, F.; Rahimmalek, M.; Pirbalouti, A.G.; Taghipoor, M. Effect of Different Drying Treatments on Essential Oil Yield, Composition and Color Characteristics of Kelussia Odoratissima Mozaff. J. Essent. Oil Res. 2015, 27, 204–211. [Google Scholar] [CrossRef]
- Sulaiman, A.; Silva, F.V.M. High Pressure Processing, Thermal Processing and Freezing of ‘Camarosa’ Strawberry for the Inactivation of Polyphenoloxidase and Control of Browning. Food Control 2013, 33, 424–428. [Google Scholar] [CrossRef]
- Hossain, M.B.; Barry-Ryan, C.; Martin-Diana, A.B.; Brunton, N.P. Effect of Drying Method on the Antioxidant Capacity of Six Lamiaceae Herbs. Food Chem. 2010, 123, 85–91. [Google Scholar] [CrossRef]
- Yi, W.; Wetzstein, H.Y. Effects of Drying and Extraction Conditions on the Biochemical Activity of Selected Herbs. HortScience 2011, 46, 70–73. [Google Scholar] [CrossRef]
- Chong, K.L.; Lim, Y.Y. Effects of Drying on the Antioxidant Properties of Herbal Tea from Selected Vitex Species: Antioxidant Properties of Dried Vitex Leaves. J. Food Qual. 2012, 35, 51–59. [Google Scholar] [CrossRef]
- Grauso, L.; de Falco, B.; Lanzotti, V.; Motti, R. Stinging Nettle, Urtica dioica L.: Botanical, Phytochemical and Pharmacological Overview. Phytochem. Rev. 2020, 19, 1341–1377. [Google Scholar] [CrossRef]
- Pogozhikh, N.I.; Tovchiga, O.V.; Evlash, V.V.; Stepanova, S.I.; Koyro, O.O. Substantiation of the Rational Drying Conditions for the Herbal Raw Material of Goutweed (Aegopodium podagraria L.) Aerial Part. South Asian Res. J. Nat. Prod. 2018, 1, 1–11. [Google Scholar] [CrossRef]
- Orav, A.; Viitak, A.; Vaher, M. Identification of Bioactive Compounds in the Leaves and Stems of Aegopodium podagraria by Various Analytical Techniques. Procedia Chem. 2010, 2, 152–160. [Google Scholar] [CrossRef]
- Ebun, O.; Santosh, K. Effect of Domestic Cooking on the Polyphenolic Content and Antioxid. World J. Dairy Food Sci. 2011, 6, 189–194. [Google Scholar]
- Chumyam, A.; Whangchai, K.; Jungklang, J.; Faiyue, B.; Saengnil, K. Effects of Heat Treatments on Antioxidant Capacity and Total Phenolic Content of Four Cultivars of Purple Skin Eggplants. ScienceAsia 2013, 39, 246–251. [Google Scholar] [CrossRef]
- Burgos, G.; Amoros, W.; Muñoa, L.; Sosa, P.; Cayhualla, E.; Sanchez, C.; Díaz, C.; Bonierbale, M. Total Phenolic, Total Anthocyanin and Phenolic Acid Concentrations and Antioxidant Activity of Purple-Fleshed Potatoes as Affected by Boiling. J. Food Compos. Anal. 2013, 30, 6–12. [Google Scholar] [CrossRef]
- Bellail, A.A.; Shaltout, O.E.; Youssef, M.M.; Gamal, A.M.A.E. Effect of Home-Cooking Methods on Phenolic Composition and Antioxidant Activity of Sweetpotato (Ipomoea batatas (L.) Lam.) Cultivars Grown in Egypt. FNS 2012, 3, 490–499. [Google Scholar] [CrossRef] [Green Version]
- McLusky, S.R.; Bennett, M.H.; Beale, M.H.; Lewis, M.J.; Gaskin, P.; Mansfiels, J.W. Cell Wall Alterations and Localized Accumulation of Feruloyl-39-Methoxytyramine in Onion Epidermis at Sites of Attempted Penetration by Botrytis Allii Are Associated with Actin Polarisation, Peroxidase Activity and Suppression of Flavonoid Biosynthesis. Plant J. 1999, 17, 523–534. [Google Scholar] [CrossRef]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal Processing Enhances the Nutritional Value of Tomatoes by Increasing Total Antioxidant Activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef]
- Hailemariam, G.A.; Wudineh, T.A. Effect of Cooking Methods on Ascorbic Acid Destruction of Green Leafy Vegetables. J. Food Qual. 2020, 2020, 8908670. [Google Scholar] [CrossRef]
- Klopsch, R.; Baldermann, S.; Voss, A.; Rohn, S.; Schreiner, M.; Neugart, S. Bread Enriched With Legume Microgreens and Leaves—Ontogenetic and Baking-Driven Changes in the Profile of Secondary Plant Metabolites. Front. Chem. 2018, 6, 322. [Google Scholar] [CrossRef]
- Fiol, M.; Weckmüller, A.; Neugart, S.; Schreiner, M.; Rohn, S.; Krumbein, A.; Kroh, L.W. Thermal-Induced Changes of Kale’s Antioxidant Activity Analyzed by HPLC–UV/Vis-Online-TEAC Detection. Food Chem. 2013, 138, 857–865. [Google Scholar] [CrossRef]
- Nayak, B.; Liu, R.H.; Berrios, J.D.J.; Tang, J.; Derito, C. Bioactivity of Antioxidants in Extruded Products Prepared from Purple Potato and Dry Pea Flours. J. Agric. Food Chem. 2011, 59, 8233–8243. [Google Scholar] [CrossRef]
- Li, D.; Zhu, Z.; Sun, D.-W. Effects of Freezing on Cell Structure of Fresh Cellular Food Materials: A Review. Trends Food Sci. Technol. 2018, 75, 46–55. [Google Scholar] [CrossRef]
- Nicoli, M.C.; Anese, M.; Parpinel, M. Infuence of Processing on the Antioxidant Properties of Fruit and Vegetables. Trends Food Sci. Technol. 1999, 10, 94–100. [Google Scholar] [CrossRef]
- Song, H.-P.; Kim, D.-H.; Jo, C.; Lee, C.-H.; Kim, K.-S.; Byun, M.-W. Effect of Gamma Irradiation on the Microbiological Quality and Antioxidant Activity of Fresh Vegetable Juice. Food Microbiol. 2006, 23, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Gil, M.I.; Ferreres, F.; Tomás-Barberán, F.A. Effect of Postharvest Storage and Processing on the Antioxidant Constituents (Flavonoids and Vitamin C) of Fresh-Cut Spinach. J. Agric. Food Chem. 1999, 47, 2213–2217. [Google Scholar] [CrossRef]
- Häkkinen, S.H.; Kärenlampi, S.O.; Mykkänen, H.M.; Törrönen, A.R. Influence of Domestic Processing and Storage on Flavonol Contents in Berries. J. Agric. Food Chem. 2000, 48, 2960–2965. [Google Scholar] [CrossRef]
- Mitra, S.; Tareq, A.M.; Das, R.; Emran, T.B.; Nainu, F.; Chakraborty, A.J.; Ahmad, I.; Tallei, T.E.; Idris, A.M.; Simal-Gandara, J. Polyphenols: A First Evidence in the Synergism and Bioactivities. Food Rev. Int. 2022. [Google Scholar] [CrossRef]
- Platzer, M.; Kiese, S.; Herfellner, T.; Schweiggert-Weisz, U.; Eisner, P. How Does the Phenol Structure Influence the Results of the Folin-Ciocalteu Assay? Antioxidants 2021, 10, 811. [Google Scholar] [CrossRef]
- Souci, S.W.; Fachmann, W.; Kraut, H. Food Composition and Nutrition Tables, 8th ed.; German Research Centre of Food Chemistry, Ed.; MedPharm Scientific Publishers: Berlin/Bonn, Germany, 2016; ISBN 978-3-8047-5072-2. [Google Scholar]
- Ma, J.F.; Tsay, Y.-F. Transport Systems of Mineral Elements in Plants: Transporters, Regulation and Utilization. Plant Cell Physiol. 2021, 62, 539–540. [Google Scholar] [CrossRef]
- Améziane, J.; Aplincourt, M.; Dupont, L.; Heirman, F.; Pierrard, J.-C. Thermodynamic Stability of Copper (II), Manganese (II), Zinc (II) and Iron (II) Complexes with Chlorogenic Acid. Bull. Soc. Chim. Fr. 1996, 133, 243–249. [Google Scholar]
- de Souza, R.F.V.; De Giovani, W.F. Antioxidant Properties of Complexes of Flavonoids with Metal Ions. Redox Rep. 2004, 9, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.V.; Bone, D.E.; Carrington, M.F. Antioxidant Activity of Dulse (Palmaria palmata) Extract Evaluated in Vitro. Food Chem. 2005, 91, 485–494. [Google Scholar] [CrossRef]
- Zhou, K.; Yu, L. Total Phenolic Contents and Antioxidant Properties of Commonly Consumed Vegetables Grown in Colorado. LWT Food Sci. Technol. 2006, 39, 1155–1162. [Google Scholar] [CrossRef]
- Lynch, S.R. Interaction of Iron with Other Nutrients. Nutr. Rev. 2009, 55, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Schmeil, O.; Fitschen, J. Schmeil-Fitschen: Die Flora Deutschlands Und Angrenzender Länder: Ein Buch Zum Bestimmen Aller Wildwachsenden Und Häufig Kultivierten Gefäßpflanzen; Parolly, G., Rohwer, J.G., Eds.; 97.; Quelle & Meyer: Wiebelsheim, Germany, 2016; ISBN 978-3-494-01700-6. [Google Scholar]
- Lenth, R.V. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. 2020. Available online: https://cran.r-project.org/web/packages/emmeans/index.html (accessed on 9 August 2022).
- Hothorn, T.; Brenz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Wu, H.; Ponnampalam, E.N.; Cottrell, J.J.; Dunshea, F.R.; Suleria, H.A.R. Comprehensive Profiling of Most Widely Used Spices for Their Phenolic Compounds through LC-ESI-QTOF-MS2 and Their Antioxidant Potential. Antioxidants 2021, 10, 721. [Google Scholar] [CrossRef] [PubMed]
- Pinelli, P.; Ieri, F.; Vignolini, P.; Bacci, L.; Baronti, S.; Romani, A. Extraction HPLC Analysis of Phenolic Compounds in Leaves Stalks, Textile Fibers of Urtica Dioica L. J. Agric. Food Chem. 2008, 56, 9127–9132. [Google Scholar] [CrossRef] [PubMed]
- Orčić, D.; Francišković, M.; Bekvalac, K.; Svirčev, E.; Beara, I.; Lesjak, M.; Mimica-Dukić, N. Quantitative Determination of Plant Phenolics in Urtica Dioica Extracts by High-Performance Liquid Chromatography Coupled with Tandem Mass Spectrometric Detection. Food Chem. 2014, 143, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Poyatos, M.D.; Ruiz-Medina, A.; Zengin, G.; Llorent-Martínez, E.J. Phenolic Characterization, Antioxidant Activity, and Enzyme Inhibitory Properties of Berberis Thunbergii DC. Leaves: A Valuable Source of Phenolic Acids. Molecules 2019, 24, 4171. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Minerals | U. dioica L. [mg/100g fw] | A. podagraria L. [mg/ 100g fw] | Difference * |
|---|---|---|---|
| Al | 0.867 ± 0.022 | 0.629 ± 0.032 | + 1.4 |
| Ca | 396.546 ± 1.586 | 144.217 ± 1.293 | + 2.7 |
| Cu | 0.379 ± 0.038 | 0.317 ± 0.052 | + 1.2 |
| Fe | 2.084 ± 0.013 | 1.370 ± 0.017 | + 1.5 |
| K | 490.703 ± 3.407 | 584.154 ± 4.047 | - 1.2 |
| Mg | 50.041 ± 0.357 | 34.927 ± 0.253 | + 1.4 |
| Mn | 0.842 ± 0.006 | 0.692 ± 0.009 | + 1.2 |
| Na | 1.797 ± 0.478 | 1.427 ± 0.374 | + 1.3 |
| P | 107.967 ± 0.441 | 48.638 ± 0.451 | + 2.2 |
| S | 87.197 ± 0.480 | 28.878 ± 0.074 | + 3.0 |
| Zn | 0.721 ± 0.013 | 0.657 ± 0.131 | + 1.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Engelhardt, L.; Pöhnl, T.; Neugart, S. Edible Wild Vegetables Urtica dioica L. and Aegopodium podagraria L.–Antioxidants Affected by Processing. Plants 2022, 11, 2710. https://doi.org/10.3390/plants11202710
Engelhardt L, Pöhnl T, Neugart S. Edible Wild Vegetables Urtica dioica L. and Aegopodium podagraria L.–Antioxidants Affected by Processing. Plants. 2022; 11(20):2710. https://doi.org/10.3390/plants11202710
Chicago/Turabian StyleEngelhardt, Layla, Tobias Pöhnl, and Susanne Neugart. 2022. "Edible Wild Vegetables Urtica dioica L. and Aegopodium podagraria L.–Antioxidants Affected by Processing" Plants 11, no. 20: 2710. https://doi.org/10.3390/plants11202710