
Calcium Signaling in Plant-Insect Interactions


Department of Life Sciences and Systems Biology, University of Turin, Via Quarello 15/a, 10135 Turin, Italy
*
Author to whom correspondence should be addressed.
Plants 2022, 11(20), 2689; https://doi.org/10.3390/plants11202689
Submission received: 22 September 2022
/
Revised: 9 October 2022
/
Accepted: 11 October 2022
/
Published: 12 October 2022
(This article belongs to the Special Issue Calcium Signaling in Plants: Roles in Stress Tolerance and Development)
{kind=link}
{kind=link}
{kind=link}
Abstract
:In plant–insect interactions, calcium (Ca2+) variations are among the earliest events associated with the plant perception of biotic stress. Upon herbivory, Ca2+ waves travel long distances to transmit and convert the local signal to a systemic defense program. Reactive oxygen species (ROS), Ca2+ and electrical signaling are interlinked to form a network supporting rapid signal transmission, whereas the Ca2+ message is decoded and relayed by Ca2+-binding proteins (including calmodulin, Ca2+-dependent protein kinases, annexins and calcineurin B-like proteins). Monitoring the generation of Ca2+ signals at the whole plant or cell level and their long-distance propagation during biotic interactions requires innovative imaging techniques based on sensitive sensors and using genetically encoded indicators. This review summarizes the recent advances in Ca2+ signaling upon herbivory and reviews the most recent Ca2+ imaging techniques and methods.
1. Introduction
The plant–insect interaction dates back to prehistorical times [1]. The evolution of plant defense responses against herbivores requires stress response-associated mechanisms, including phytohormones production [2,3,4], host specificity [5], protease inhibitor activities [6,7], plant volatile emission [8], phenotypic plasticity [9], cross-species communication [10], morpho-physiological and chemical reprogramming [11], response to insect-derived elicitors or effectors [12,13], all involving metabolic costs [14].
The recognition and appropriate response to the attacking enemy occur within a few minutes and require the activation of specific pathways most of which finally lead to gene activation (the signaling pathway) [15]. Upon insect feeding on plants, rapid early events eventually lead to the production and release of plant defense molecules [16]. These signaling events also function in propagation of long-distance signals (calcium, reactive oxygen species, and electrical signals), which contribute to rapid, systemic induction of defense responses [17]. Interestingly, the speed of response to biotic damage is proportional to the speed of feeding. For instance, chewing herbivores induce faster responses that sucking herbivores, which in turn are faster in inducing plants responses when compared to microorganisms [18]. However, the term rapid is relative to the communication systems of plants which by no way is comparable or similar the to highly efficient and rapid nervous system of animals, as recently clarified [19]. Therefore, biotic stresses induced by herbivores result in diverse physiological changes in plants.
Generation of Calcium Signals
One of the earliest players in plant–insect interactions is calcium (Ca2+), a universal second messenger the intracellular variation in concentration of which has been associated to the plant perception of biotic stress [20,21]. Herbivory-induced cytosolic Ca2+ concentration ([Ca2+]cyt) elevation involves multiple channels and pathways regulating local and long-distance [Ca2+]cyt signals [22]. The depolarization of the plasma membrane potential (Vm) is another early event induced by insect feeding on leaves, and results from both the damage caused by the insect and the delivery of oral secretions (OS) [23]. Although this herbivore-induced Vm depolarization depends on a Ca2+-dependent opening of potassium ion (K+) channels [24], the attacked leaf remains depolarized for an extended period, which cannot be explained by the sole action of K+ channels. The plant plasma membrane H+-ATPase is strongly inhibited by insect OS, and this inhibition contributes to the log-lasting Vm depolarization [25]. Studies on mechanical and robotic wounding of leaves show that these treatments do not induce a Ca2+ signaling, whereas significant increase in [Ca2+]cyt were observed only after herbivory [26].
Herbivore attacks can trigger Ca2+ waves traveling a long distance to transmit and convert the local signal to a systemic defense program in the whole plant, and recent studies uncovered Ca2+ channels that orchestrate specific Ca2+ signatures [27]. Electrical signals can result in systemic Ca2+ and reactive oxygen species (ROS) waves, as demonstrated in several insect herbivores such as Spodoptera littoralis, Pieris brassicae, Myzus persicae and Tetranychus urticae feeding of model plants like Arabidopsis thaliana; important crops like Hordeum vulgare, Phaseolus lunatus, Nicotiana tabacum and Vicia faba; the living fossil plant Ginkgo biloba and the fern Pteris vittata [18,24,28,29,30,31,32].
Mutations affecting Ca2+ homeostasis interfere with the plant’s leaf-to-leaf electrical signaling capacity as shown when Arabidopsis type IIB auto-inhibited Ca2+-ATPases (ACAs) was used as a candidate gene for a reverse genetic screening [33]. Both root-to-shoot Ca2+ waves and slow wave potentials (SWPs) are triggered by either root wounding or the application of glutamate (Glu) to wounded roots. Ca2+ waves and SWPs are dependent on the activity of glutamate receptor-like proteins (GLRs) and P-type H+-ATPase AHA1. Inhibition of the H+-ATPase activity due to herbivore OS [25] induces the alkalinization of the apoplast that depolarizes Vm. The activation of GLRs, along with the release of Glu from damaged phloem, causes the Vm depolarization in the form of SWPs that together with the [Ca2+]cyt increase propagates the systemic wound signaling [34,35]. Recently, a key functional role for the cyclic nucleotide-gated ion channel CNGC19 in Arabidopsis defense against Spodoptera litura demonstrated the role of Ca2+-mediated defense signaling, with the modulation of late events like the biosynthesis of phytohormones and secondary metabolites [36].
Wounding may also induces a drop in hydraulic pressure (turgor pressure, ΨT) and an increase in apoplastic amino acids (AAapo), including Glu, that are perceived by plasma-membrane located mechanosensitive ion channels (MSC) that depolarize the plant Vm [37]. Finally, herbivore’s OS may contain oligosaccharides that interact with specific plant plasma membrane receptors [24]. For instance, spider mites secrete elicitors (tetranin1 and tetranin2) that trigger [Ca2+]cyt, ROS production and Vm depolarization [12]. In Arabidopsis, plasma membrane localized NADPH oxidases respiratory burst oxidase homolog D (RBOHD) functions as an essential regulator of ROS [38]. Herbivore-produced elicitors like N-linolenoyl-l-glutamine (GLN18:3) induce a RBOHD and RBOHF-dependent ROS burst in several plant species [13,39].
Figure 1 summarizes the early events upon herbivory where Ca2+ plays a central role along with Vm depolarization and ROS production.
2. Calcium and ROS Signaling
The plant shows a consistent number and variability of signaling molecules throughout, but three potentially interacting messengers: ROS, Ca2+ and electrical signaling are interlinked to form a network supporting rapid signal transmission [40,41,42,43].
Several plant species produce hydrogen peroxide (H2O2) in response to herbivory. The expression of the Ca2+-sensing aequorin system in transgenic soybean (Glycine max) suspension cells and the effects of herbivore-wounding in Lima bean (P. lunatus) leaves show increasing amounts of H2O2 that correlate with a higher [Ca2+]cyt [44]. The same responses were found in the living fossil G. biloba leaves damaged by S. littoralis [30] and in the fern P. vittata [29]. Changes of [Ca2+] fluxes and H2O2 were detected in a tobacco variety (G140) infested by the tobacco aphid M. persicae. Even in this case, H2O2 accumulation depended on Ca2+ influx, which were both increased in a long period of aphid feeding [45].
Ca2+ and ROS signaling are induced by herbivore-associated molecular patterns (HAMPs) [46]. As shown in Figure 1, ROS signals could activate the Ca2+ permeable channels in membranes [47]; ROS and Ca2+ signaling are the consequence of the combined effect of wounding and the secretion of salivary elicitors from the feeding insect [48,49,50]. For instance, aphids like Acyrthosiphon pisum produce two types of saliva that mediate their interactions with plants which are based on Ca2+ signaling-related responses [51]. The activity of S. littoralis ventral eversible gland contains elicitors able to trigger both Ca2+ signaling and H2O2 production in Arabidopsis [52]. The piercing-sucking planthopper (Laodelphax striatellus) secretes salivary proteins into its plant host during feeding that enhance plant resistance to insects by inducing Ca2+ and ROS signaling pathways [53]; whereas the tobacco hornworm caterpillar (Manduca sexta) OS induce Ca2+-dependent ROS production in tomato (Solanum lycopersicum) protoplasts [54]. Elicitors produced during the feeding activity of the plant-sucking arthropod herbivore two-spotted spider mite (T. urticae) were found to trigger the [Ca2+]cyt influx and the generation of ROS in kidney bean plants (P. vulgaris) [12].
Besides OS direct activity, several HAMPs released upon cell damage trigger cascades of events, eventually leading to a coordinated response. Among these, extracellular DNA (eDNA), deriving from cell disruption made by herbivory, induces an increase in [Ca2+]cyt; this event is associated to the opening of K+ channels, with particular action on ligand-gated rectified K+ channels, which in turn are correlated to ROS production [55,56]. It has been suggested that fragmented DNA might play a crucial role as an important and powerful elicitor involved in early and late responses to biotic stress [56]. Extracellular ATP (eATP) is a constitutive HAMP that is released by wounding and herbivory. eATP can increase [Ca2+]cyt, and Arabidopsis loss of function mutants were found to have an impaired increase in [Ca2+]cyt in response to eATP, thus demonstrating a link between eATP and Ca2+ signaling [57].
Calcium and ROS signaling are also induced by volatile organic compounds (VOCs) [15]. Plants respond to herbivory by emitting VOCs composed of both green leaf volatiles (GLVs) and terpenoids, which are released into the surrounding atmosphere and are perceived by receiving plants [58]. GLVs emitted by herbivore-wounded tomato (including (Z)-3-hexenyl acetate) were found to induce a strong [Ca2+]cyt increase in receiving plants [59]. The emission of β-ocimene induced by (Z)-3-hexenol, linalool, α-farnesene and (E)-4,8–dimethyl–1,3,7-nonatriene (DMNT) (which are typical VOCs emitted upon herbivory) were dependent on Ca2+ signaling [60]. H2O2 and [Ca2+]cyt increased in Arabidopsis mesophyll cells after being treated with the volatile monoterpene alcohol linalool in the attempt to increase plant resistance to diamondback moth (Plutella xylostella) [61].
Other interesting effects of plant–insect interactions on Ca2+ and ROS signaling were found when (Z)-11-hexadecenal, the main component of P. xylostella female sex pheromone, was tested in Brassica nigra. The sex pheromone induced depolarization of the Vm, an increase in [Ca2+]cyt and production of H2O2, demonstrating the ability of plants to detect and respond to volatiles emitted by non-plant organisms [62].
3. Calcium Binding Proteins (CBPs)
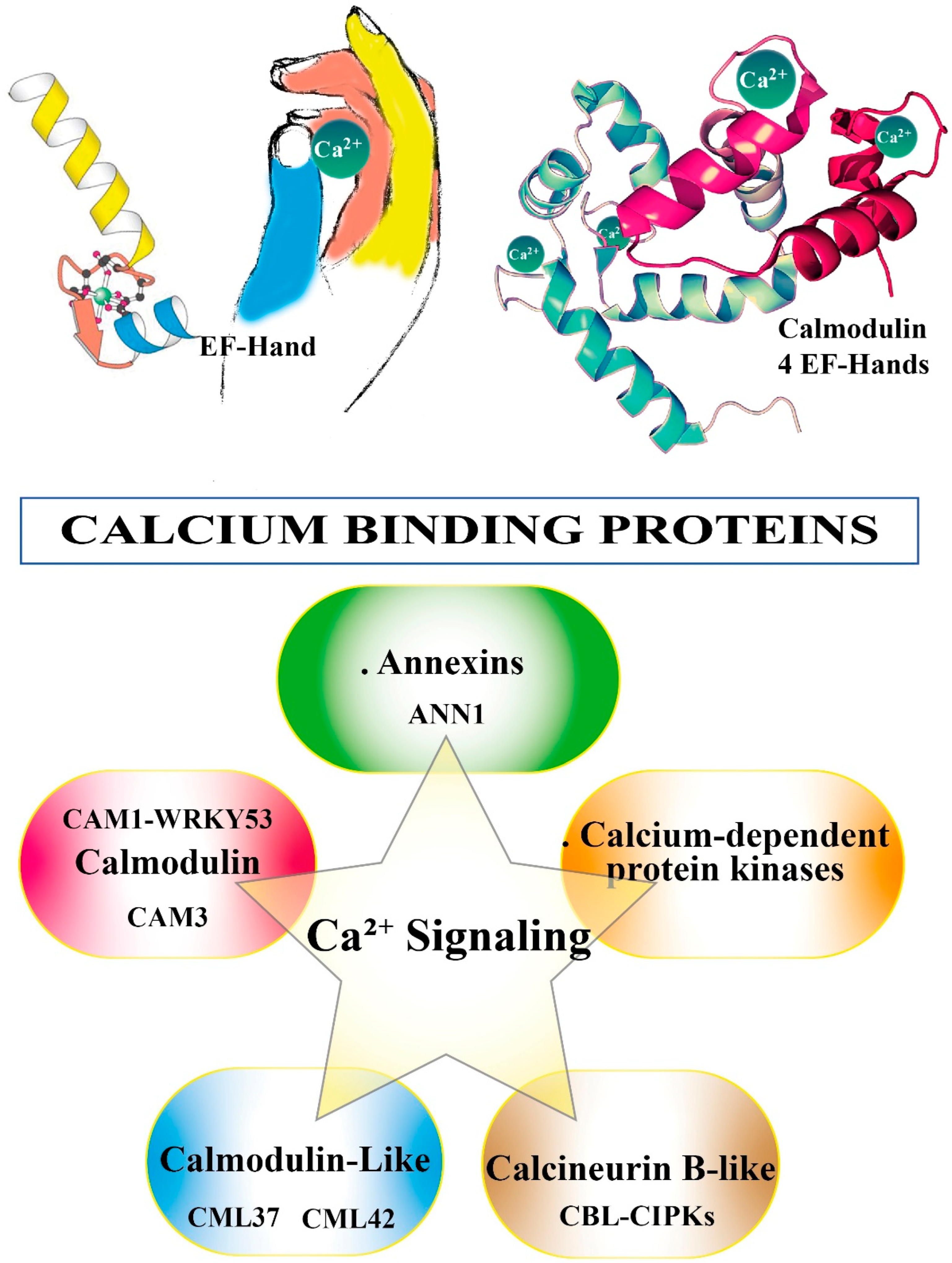
To activate the appropriate cell response and in order to become informative, the Ca2+ message needs to be decoded and relayed. This task is brought by Ca2+-binding proteins (CBPs) [63]. Most CBPs are characterized by the presence in their sequence of the canonical Ca2+-binding motif called the EF-hand [64] (see Figure 2). CBPs in the saliva of herbivorous insects function as effectors to attenuate host plant defenses and thus improve insect feeding performance [65]. A putative EF-hand Ca2+-binding protein (LsECP1) in the small brown planthopper Laodelphax striatellus exhibits Ca2+-binding activity. Knocking down LsECP1 in L. striatellus prompted a higher level of [Ca2+]cyt in rice fed plants, whereas overexpression suppressed wound-induced H2O2 accumulation [66].
CBPs are also involved in the transmission of devastating viruses into plant phloem. Viral infection may inhibit CBP expression and this event facilitates filament-mediated viral secretion first into salivary cavities and then into plant phloem which in rice causes an increased [Ca2+]cyt, followed by a substantial H2O2 production [67].
The NlSEF1 protein is a CBP highly expressed in the salivary glands of the brown planthopper Nilaparvata lugens. It has an EF-hand Ca2+-binding activity and can be secreted into rice plants when infested by the planthopper. Knocking down NISEF1 in the planthopper elicited higher levels of [Ca2+]cyt and H2O2 in rice, indicating that NlSEF1 functions as an effector and plays important roles in interactions between the planthopper and rice by mediating the plant’s defense responses [68,69].
3.1. Calmodulin (CaM) and Calmodulin-like (CML)
Calmodulin (CaM) is present in all eukaryotic cells and is the prototype of a CBP [70]. Being involved in the Ca2+ signaling pathway, CaM acts by modifying its interactions with various CaM-binding proteins [20]. A divergent form of CaMs (CaM-like proteins, CMLs) is also present in plants [71,72].
The salivary glands of planthoppers N. lugens and Laodelphax striatellus show high expression of CaMs that are secreted into the rice plants during feeding and CaM-silenced planthopper nymphs elicited relatively high levels of H2O2 and callose accumulation in rice plants [73]. The transcriptome of spinach leaves exposed to beet armyworm (Spodoptera exigua) larvae identified four CAMs (CAM3, CAM5-1, CAM6-1, and CAM6-2) which were associated with Ca2+ signaling. The genes coding for these CAMs were highly co-overexpressed with several endoplasmic reticulum-type Ca2+-transporting-ATPase genes, known to be involved in Ca2+ transport, confirming also the important role of the Ca2+ signaling cascade in the defense against insect attack [74]. In Arabidopsis, calmodulin 1 (CAM1) cam1 mutants were more resistant to S. littoralis than in the wild-type Arabidopsis group [75]; whereas, calmodulin 3 (CAM3) participated in Ca2+-ATPase activation [61]. CAM3 was also found to interact with the N-terminus of Ca2+-ATPase isoform 8 (ACA8), and treatment with the monoterpene linalool of Arabidopsis leaves showed the enhancement of some JA-related genes and defense genes expressions [61]. cam1 and wrky53 mutants are more resistant to S. littoralis than the wild-type Arabidopsis. The high [Ca2+]cyt causes the breaking down of the CAM1-WRKY53 complex, then the detachment of WRKY53 reduces the JA content by downregulating the LOXs gene expression [75]. CaM can both positively and negatively regulate cyclic nucleotide-gated channels (CNGCs) [76], which are involved in jasmonate-induced Ca2+ mobilization [77], a typical response to herbivore attack [78].
Plants contain unique Ca2+-sensing proteins, the calmodulin-like-proteins (CML), which are involved in many stresses and developmental responses [72]. In soybean, feeding by S. litura, treatment with signaling compounds and wounding induce the differential expression patterns of CMLs, indicating their involvement in Ca2+ signaling and plant defense during herbivory [79]. In defense against herbivory, Arabidopsis CML37 positively regulates the plants’ defense against S. littoralis [80], whereas CML42 shows a downregulating activity and might play a role as a Ca2+ sensor by displaying diverse functions in responses to both abiotic stress and insect herbivory [81]. Thus, during herbivory, plants may use CML37 and CML42 as Ca2+ sensor proteins to maintain equilibrium and adjust the signaling and downstream responses [82]. S. littoralis feeding induces Arabidopsis CML9 upon wounding and feeding; however, CML9 loss-of-function mutant lines were not affected by herbivory and the same occurred in overexpressing Arabidopsis lines. This indicates that in Arabidopsis CML9 acts more like a specialized rather than a general regulator of stress responses [78].
3.2. Calcium-Dependent Protein Kinases (CPKs)
In contrast to CaMs and CMLs, Ca2+-dependent protein kinases (CPKs or CDPKs) are distinctive Ca2+ decoders because of their characteristic of having both downstream signal propagation capabilities and a Ca2+ sensing component. They comprise a variable N-terminal part, a kinase domain and an activation domain [83]. The Ca2+ binding of the CaM-like domain prompts a conformational change that dislocates the pseudo-substrate from the kinase; this allows the occurrence of downstream phosphorylation events [84]. Ca2+-binding sensory proteins such as CPKs are involved in responses to herbivory following [Ca2+]cyt variations [15,85,86], and the involvement of CPKs in response to herbivore feeding has been reviewed [87]. Transgenic N. attenuata plants, in which two CPKs were silenced, when attacked by M. sexta larvae induced high levels of defense metabolites that slowed the insect growth, demonstrated the critical roles of CPKs in modulating phytohormone (jasmonic acid) homeostasis and highlighted the complex coupling between phytohormones (e.g., jasmonate) and MAPK signaling [88]. In apple (Malus × domestica), 30 CPK genes (MdCPK) were identified and correlated to the Ca2+ signaling of resistant cultivars as compared to susceptible cultivars [89]. In soybean, CPKs’ transcript levels changed after wounding and exhibited specific expression patterns upon simulated S. exigua feeding or soybean aphid (Aphis glycines) herbivory, by revealing an interesting role of CPKs in soybean-insect interactions [90].
3.3. Calcineurin B-like (CBL)
Calcineurin B-like proteins (CBLs) are a family of Ca2+ sensor proteins present in plants that bind Ca2+ through EF-hand motifs (see Figure 2) [91,92,93]. The CBL family is unique as well. CBL-interacting protein kinase (CIPK) are downstream kinases that activate specific targets and transduce signals [94]. Changes in Ca2+ levels are detected by CBLs that, upon Ca2+ binding, modify their conformation with the subsequent interaction and activation of downstream CIPKs and CBL [95]. CBL-CIPKs play a major role in plant responses to abiotic stress [96] (e.g., salt stress) and are involved in phytohormone-, ion homeostasis- and sucrose homeostasis-related crosstalks between plant development and stress adaptation [97]. CBLs are involved also in biotic stress. Cells of N. benthamiana leaves overexpressing sugarcane CBL genes after inoculation with the tobacco pathogen Ralstonia solanacearum show that overexpression of CBL genes can effectively promote resistance to infections in tobacco plants [98]. After exposure to the pathogenic fungus Sclerotinia sclerotiorum, CBL1 overexpressing plants display better performance under unfavorable stress conditions and ectopic expression of CBL1 enhanced biotic stress tolerance by facilitating scavenging of the Na+ and ROS from the cell [99]. CBL-CIPKs have been demonstrated to play a role also in plant responses to herbivory [100].
3.4. Annexins (ANNs)
Annexins are proteins widely present in living organisms that bind to membrane phospholipids in a Ca2+-dependent manner and are involved in the regulation of plant growth and response to environmental stimuli [47]. Evidence has been provided for a contribution of ANNs in Ca2+ transport [22] and increasing [Ca2+]cyt enhances the binding of annexin to plasma membrane [101], thus identifying ANNs as Ca2+ sensors or effectors in [Ca2+]cyt-dependent processes [102]. In pepper (Capsicum annuum) and tomato (Solanum lycopersicum), the expression of ANNs was found to be induced by biotic stress, including thrip infestation [103]. Resistant pepper plants attacked by the insect herbivore Bemisia tabaci were characterized by an increased expression of annexin D4-like (ANN4) [104]. In Arabidopsis, overexpression of ANN1 and ANN4 were found to increase the plant resistance to root knot nematodes [105], while ANN1 overexpressing plants induced S. littoralis larvae to gain less weight, confirming the importance of ANN1 in plant–insect interactions with a specific role in systemic rather than local defense responses [22]. Besides their role as CBP, ANNs may be able to act as peroxidase [106] with a phosphorylation/dephosphorylation regulation [107], in agreement with the suggestion that ANNs might provide a molecular link between ROS and [Ca2+]cyt in the systemic defense-related signaling in plants [22]. Indeed, AtAnn1 mutants have reduced Ca2+ signature upon H2O2 treatment, suggesting an important role of ANNs in ROS-induced Ca2+ signatures [108].
Figure 2 shows the Ca2+ binding to EF-hand motif and summarizes the CBPs involved in Ca2+ signaling.
4. Mitogen-Activated Protein Kinases (MAPKs)
Elicitor perception that induces Ca2+ influx, ROS and nitrogen reactive species (NRS) is followed by the activation of other major players in plant–insect interaction: mitogen-activated protein kinases (MAPKs) [109,110]. In Nicotiana attenuata, MAPKs play central roles in modulating herbivory-induced phytohormone and anti-herbivore secondary metabolites, thus regulating wounding- and herbivory-induced responses [111,112,113]. Several MAPK genes have been identified from chickpea (Cicer arietinum) in response to infestation by Helicoverpa armigera [114,115]; whereas two MAPKs were identified in soybean (Glycine max) attacked by the green stink bug (Nezara viridula) [116]. The expression of a MAPK in rice (OsMKK3) was induced by both mechanical wounding and the infestation of brown planthopper N. lugens [117], whereas RNA-Seq analyses of Balsas teosinte (Zea mays ssp. parviglumis) attacked by the armyworm (Mythimna separata) revealed up-regulation of genes markedly enriched in MAPK cascade-mediated signaling pathway [118]. In the date palm (Phoenix dactylifera) infested by the Dubas bug (Ommatissus lybicus), transcriptome analysis revealed the differentially expressed genes mostly belonged to calcium and MAPKs signaling pathways [119]. However, activation of MAPKs has been demonstrated also with the sole mechanical wounding [28,120].
5. Calcium Imaging
A breakthrough in the field of Ca2+ imaging was the development of sensitive fluorescent Ca2+ indicators, dyes and buffers by Tsien in 1980 [121]. Reporter dyes (small organic molecule-based sensors) are useful tools to measure Ca2+ levels without the need to genetically transform plants. This allows Ca2+ imaging also in those plants for which it is difficult obtaining transgenic lines [122]. In the last paragraph of this review, we focus on the development of new sensors used for Ca2+ imaging in plant–insect interactions.
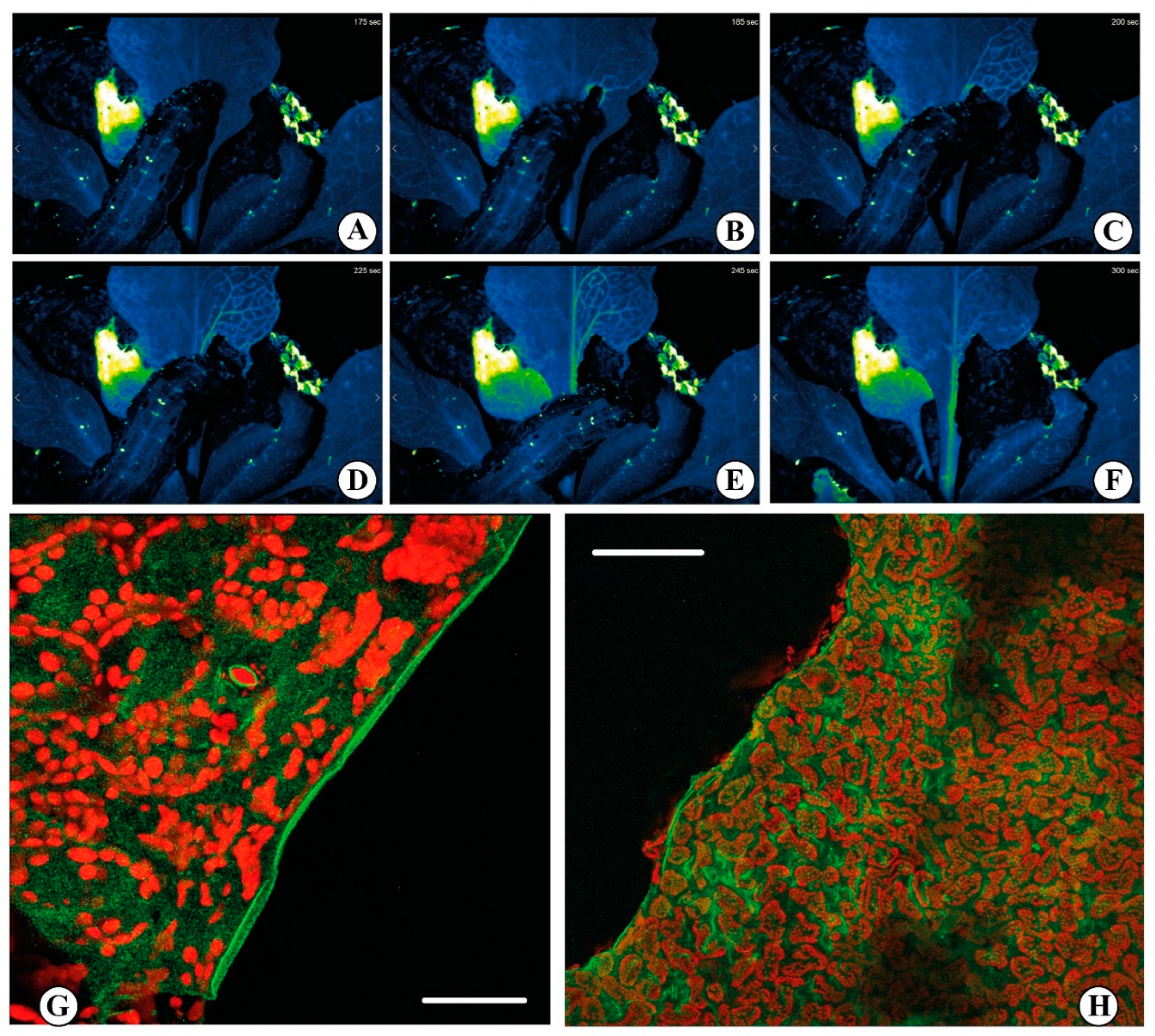
The use of synthetic dyes (e.g., Calcium Orange) allowed the study the effect of insect chewing on leaves, allowing to observe that Ca2+ release was localized at the very edge of the bite (see Figure 3) [28]. Although reporter dyes have been foundational in developing the field of Ca2+ imaging, the advent of fluorescent protein (FP) technologies, and the associated possibility of engineering genetically encoded indicators, led to new interesting results [122,123]. In biology, a myriad of signaling processes is quantified by the use of fluorescent biosensors, which are fluorescent molecules introduced into an organism to monitor some parameters of some biological activity [124]. The development of different fluorescent sensors enabled the single cell, tissue, organ and even whole plant visualization of Ca2+ signals [125]. In plant studies, the bioluminescent protein aequorin (derived from the jellyfish Aequorea victoria) was first used in 1967 as a genetically encoded Ca2+ biosensor [126]. However, despite the use of aequorin to detect Ca2+ changes in response to various stresses in plants [127], including temperature [128], pathogens [129], salt stress [127] and wounding [130] was successful, this system it is not well-suited for real-time imaging due to the extremely low luminescent signal it produces [131]. Anyway, aequorin-expressing transgenic plants have been used in the study of diverse microbe- or damage-associated molecular patterns (MAMPs/DAMPs) where comparative analyses between the elicitors flg22 and elf18 revealed differences in Ca2+ signaling and cellobiose exposure which generated an intracellular Ca2+ elevation [129]. They have also been instrumental to quantify the response to insect OS [28]. Indeed, the study of these Ca2+ elevations has been largely restricted to the use of elicitors as opposed to living organisms [132].
We can devise the fluorescence-based genetically encoded indicators into two classes: (i) intensiometric single-FP biosensors and (ii) ratiometric FRET-based biosensors [125]. Although ratiometric FRET-based sensors are quantitatively accurate [133], intensiometric Ca2+ indicators, including GCaMP3 [134] and R-GECO1 [135], provide both higher temporal resolution and ease of use due to their generally brighter Ca2+-responsive signal and their simpler microscope requirements [122,125]. Comparing the ratiometric fluorescence-based Ca2+ sensor Yellow Cameleon NES-YC3.6 [136] and the intensity-based sensor R-GECO1, it has been demonstrated that the latter shows a significantly enhanced signal change in response to several stimuli [135]. Examples of the greater sensitivity of R-GECO1 have been provided int the study of Ca2+ signals in flg22- and chitin-induced experiments on a cellular scale. Here the use of GECO1 allowed to visualize well defined [Ca2+]cyt oscillations in both epidermal and guard cells [135].
In the last decade, the approach to Ca2+ imaging using genetically encoded indicators had a great impact in the study of Ca2+ signaling, and a good amount of protocols and review papers described how to perform plant Ca2+ imaging in vitro but especially in vivo [20,133,134,137,138,139,140,141,142,143,144,145]. These fluorescent-based genetically encoded probes have been successfully used to study Ca2+ signaling in the detection of abiotic stresses [35]. In fact, the Ca2+ involvement in response to abiotic stimuli such as wounding is well documented [46], but its role in plant defense strategies to insect herbivory and other pathogen attacks is less understood [146]. S. littoralis feeding on an Arabidopsis plants expressing the GCaMP3 fluorescent protein–based [Ca2+]cyt sensor showed a rise in [Ca2+]cyt at the wounding position within 2 sec [134]. Furthermore, the signal spread to distal leaves in 1 to 2 min with a more pronounced signature in the vasculature, especially when the feeding activity was affecting the major vein. The propagation of this [Ca2+]cyt signal was observed moving from younger to older leaves and vice versa [35]. By following the protocol of Toyota [35], we observed in Arabidopsis expressing the genetically encoded R-GECO1 sensor an increase in the [Ca2+]cyt upon S. littoralis feeding on a rosette leaf. The signal propagates quickly through the vasculature (Figure 3 and Video S1). Thus, it is possible to investigate the complexity of Ca2+ signaling with the use of reporter dyes and especially with the more recent genetically encoded Ca2+ indicators, which can be very helpful to decode both abiotic and biotic responses in plants.
6. Conclusions
Ca2+ variations are among the earliest events during plant–insect interactions. Ca2+ waves arise by the wounded plant to transmit at long distance and to convert local signals into a systemic defense program. In this review, we showed that the Ca2+ signaling is accompanied by other important rapid signals, with particular reference to ROS and electrical signaling. The interdependence of these three networked events is clear in plant-biotic interactions and have been demonstrated several times in plant–insect interactions. In order to be decoded and relayed, the Ca2+ message requires proteins able to bind the ion and trigger the signal transduction pathways that lead to plant responses to biotic attack. We described the role of calmodulin, calmodulin-like proteins, Ca2+-dependent protein kinases, annexins and calcineurin B-like proteins in plant–insect interactions. An important role in the study of Ca2+ signaling is the ability to visualize in real-time the generation of Ca2+ signals both at the single cell and whole plant level. Innovative imaging techniques have been developed based on sensitive sensors and using genetically encoded indicators that open new ways in the deciphering of the Ca2+ signaling in plant–insect interactions.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants11202689/s1, Video S1: Rapid Ca2+ signals generated when Spodoptera littoralis feeds on Arabidopsis thaliana expressing the genetically encoded R-GECO1 sensor.
Author Contributions
Conceptualization, M.E.M.; methodology, M.E.M.; investigation, A.S.P. and M.E.M.; resources, M.E.M.; writing—original draft preparation, A.S.P. and M.E.M.; writing—review and editing, A.S.P. and M.E.M.; funding acquisition, M.E.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the University of Turin Local Research grants to M.E.M.
Acknowledgments
The authors whish to thank A. Costa, M. Grenzi and NOLIMITS, an advanced imaging facility established by the University of Milan, for providing the video and images of the fast calcium imaging analyses upon S. littoralis herbivory.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cariglino, B.; Moisan, P.; Lara, M.B. The fossil record of plant-insect interactions and associated entomofaunas in Permian and Triassic floras from southwestern Gondwana: A review and future prospects. J. S. Am. Earth Sci. 2021, 111, 103512. [Google Scholar] [CrossRef]
- Arimura, G.-I.; Garms, S.; Maffei, M.; Bossi, S.; Schulze, B.; Leitner, M.; Mithöfer, A.; Boland, W. Herbivore-induced terpenoid emission in Medicago truncatula: Concerted action of jasmonate, ethylene and calcium signaling. Planta 2007, 227, 453–464. [Google Scholar] [CrossRef] [Green Version]
- Pan, L.L.; Miao, H.Y.; Wang, Q.M.; Walling, L.L.; Liu, S.S. Virus-induced phytohormone dynamics and their effects on plant-insect interactions. New Phytol. 2021, 230, 1305–1320. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.; Yamasaki, Y.; Shinya, T.; Galis, I.; Arimura, G.-I. Phytohormone elicitation in maize by oral secretions of specialist Mythimna separata and generalist Spodoptera litura. J. Plant Interact. 2021, 16, 587–590. [Google Scholar] [CrossRef]
- Sharma, A.; Raman, A. Gall-inducing psylloidea (insecta: Hemiptera)—Plant interactions. J. Plant Interact. 2022, 17, 580–594. [Google Scholar] [CrossRef]
- Mithöfer, A. Carnivorous plants and their biotic interactions. J. Plant Interact. 2022, 17, 333–343. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Huang, P.-H.; Yeh, K.-W.; Wang, S.-J. Colonization of Piriformospora indica enhances insect herbivore resistance of rice plants through jasmonic acid- and antioxidant-mediated defense mechanisms. J. Plant Interact. 2022, 17, 9–18. [Google Scholar] [CrossRef]
- Howe, G.A.; Jander, G. Plant immunity to insect herbivores. Annu. Rev. Plant Biol. 2008, 59, 41–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashra, H.; Nair, S. Review: Trait plasticity during plant-insect interactions: From molecular mechanisms to impact on community dynamics. Plant Sci. 2022, 317, 13. [Google Scholar] [CrossRef]
- Zu, P.J.; García-García, R.; Schuman, M.C.; Saavedra, S.; Melián, C.J. Plant—Insect chemical communication in ecological communities: An information theory perspective. J. Syst. Evol. 2022, 9. [Google Scholar] [CrossRef]
- Noman, A.; Aqeel, M.; Qasim, M.; Haider, I.; Lou, Y.G. Plant-insect-microbe interaction: A love triangle between enemies in ecosystem. Sci. Total Environ. 2020, 699, 11. [Google Scholar] [CrossRef] [PubMed]
- Iida, J.; Desaki, Y.; Hata, K.; Uemura, T.; Yasuno, A.; Islam, M.; Maffei, M.E.; Ozawa, R.; Nakajima, T.; Galis, I.; et al. Tetranins: New putative spider mite elicitors of host plant defense. New Phytol. 2019, 224, 875–885. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.E.; Arimura, G.; Mithofer, A. Natural elicitors, effectors and modulators of plant responses. Nat. Prod. Rep. 2012, 29, 1288–1303. [Google Scholar] [CrossRef] [PubMed]
- Furstenberg-Hagg, J.; Zagrobelny, M.; Bak, S. Plant defense against insect herbivores. Int. J. Mol. Sci. 2013, 14, 10242–10297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanchiswamy, C.N.; Maffei, M.E. Calcium signaling preceding the emission of plant volatiles in plant-insect interactions. J. Indian Inst. Sci. 2015, 95, 15–23. [Google Scholar]
- Maffei, M.E.; Mithofer, A.; Boland, W. Insects feeding on plants: Rapid signals and responses preceding the induction of phytochemical release. Phytochemistry 2007, 68, 2946–2959. [Google Scholar] [CrossRef]
- Luo, S.T.; Zhang, X.; Wang, J.F.; Jiao, C.Y.; Chen, Y.Y.; Shen, Y.B. Plant ion channels and transporters in herbivory-induced signalling. Funct. Plant Biol. 2018, 45, 111–131. [Google Scholar] [CrossRef]
- Bricchi, I.; Bertea, C.M.; Occhipinti, A.; Paponov, I.A.; Maffei, M.E. Dynamics of membrane potential variation and gene expression induced by Spodoptera littoralis, Myzus persicae, and Pseudomonas syringae in Arabidopsis. PLoS ONE 2012, 7, e46673. [Google Scholar]
- Robinson, D.G.; Draguhn, A. Plants have neither synapses nor a nervous system. J. Plant Physiol. 2021, 263, 153467. [Google Scholar] [CrossRef]
- Aldon, D.; Mbengue, M.; Mazars, C.; Galaud, J.P. Calcium signalling in plant biotic interactions. Int. J. Mol. Sci. 2018, 19, 19. [Google Scholar]
- Kloth, K.J.; Dicke, M. Rapid systemic responses to herbivory. Curr. Opin. Plant Biol. 2022, 68, 7. [Google Scholar] [CrossRef] [PubMed]
- Malabarba, J.; Meents, A.K.; Reichelt, M.; Scholz, S.S.; Peiter, E.; Rachowka, J.; Konopka-Postupolska, D.; Wilkins, K.A.; Davies, J.M.; Oelmuller, R.; et al. ANNEXIN1 mediates calcium-dependent systemic defense in Arabidopsis plants upon herbivory and wounding. N. Phytol. 2021, 231, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Zebelo, S.A.; Maffei, M.E. Role of early signalling events in plant-insect interactions. J. Exp. Bot. 2015, 66, 435–448. [Google Scholar] [PubMed]
- Bricchi, I.; Occhipinti, A.; Bertea, C.M.; Zebelo, S.A.; Brillada, C.; Verrillo, F.; De Castro, C.; Molinaro, A.; Faulkner, C.; Maule, A.J.; et al. Separation of early and late responses to herbivory in Arabidopsis by changing plasmodesmal function. Plant J. 2013, 73, 14–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camoni, L.; Barbero, F.; Aducci, P.; Maffei, M.E. Spodoptera littoralis oral secretions inhibit the activity of Phaseolus lunatus plasma membrane H+-ATPase. PLoS ONE 2018, 13, e0202142. [Google Scholar] [CrossRef] [PubMed]
- Bricchi, I.; Leitner, M.; Foti, M.; Mithofer, A.; Boland, W.; Maffei, M.E. Robotic mechanical wounding (MecWorm) versus herbivore-induced responses: Early signaling and volatile emission in Lima bean (Phaseolus lunatus L.). Planta 2010, 232, 719–729. [Google Scholar]
- Tian, W.; Wang, C.; Gao, Q.; Li, L.; Luan, S. Calcium spikes, waves and oscillations in plant development and biotic interactions. Nat. Plants 2020, 6, 750–759. [Google Scholar]
- Maffei, M.; Bossi, S.; Spiteller, D.; Mithofer, A.; Boland, W. Effects of feeding Spodoptera littoralis on lima bean leaves. I. Membrane potentials, intracellular calcium variations, oral secretions, and regurgitate components. Plant Physiol. 2004, 134, 1752–1762. [Google Scholar]
- Imbiscuso, G.; Trotta, A.; Maffei, M.; Bossi, S. Herbivory induces a ROS burst and the release of volatile organic compounds in the fern Pteris vittata L. J. Plant Interact. 2009, 4, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Mohanta, T.K.; Occhipinti, A.; Zebelo, S.A.; Foti, M.; Fliegmann, J.; Bossi, S.; Maffei, M.E.; Bertea, C.M. Ginkgo biloba responds to herbivory by activating early signaling and direct defenses. PLoS ONE 2012, 7, e32822. [Google Scholar]
- Zimmermann, M.R.; Mithofer, A.; Will, T.; Felle, H.H.; Furch, A.C.U. Herbivore-triggered electrophysiological reactions: Candidates for systemic signals in higher plants and the challenge of their identification. Plant Physiol. 2016, 170, 2407–2419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najdenovska, E.; Dutoit, F.; Tran, D.; Plummer, C.; Wallbridge, N.; Camps, C.; Raileanu, L.E. Classification of plant electrophysiology signals for detection of spider mites infestation in tomatoes. Appl. Sci. 2021, 11, 1414. [Google Scholar] [CrossRef]
- Fotouhi, N.; Fischer-Stettler, M.; Lenzoni, G.; Stolz, S.; Glauser, G.; Zeeman, S.C.; Farmer, E.E. ACA pumps maintain leaf excitability during herbivore onslaught. Curr. Biol. 2022, 32, 2517–2528.e6. [Google Scholar] [CrossRef] [PubMed]
- Shao, Q.; Gao, Q.; Lhamo, D.; Zhang, H.; Luan, S. Two glutamate- and pH-regulated Ca2+ channels are required for systemic wound signaling in Arabidopsis. Sci. Signal. 2020, 13, eaba1453. [Google Scholar] [CrossRef] [PubMed]
- Toyota, M.; Spencer, D.; Sawai-Toyota, S.; Jiaqi, W.; Zhang, T.; Koo, A.J.; Howe, G.A.; Gilroy, S. Glutamate triggers long-distance, calcium-based plant defense signaling. Science 2018, 361, 1112–1115. [Google Scholar] [CrossRef] [PubMed]
- Meena, M.K.; Prajapati, R.; Krishna, D.; Divakaran, K.; Pandey, Y.; Reichelt, M.; Mathew, M.K.; Boland, W.; Mithofer, A.; Vadassery, J. The Ca2+ channel CNGC19 regulates arabidopsis defense against spodoptera herbivory. Plant Cell 2019, 31, 1539–1562. [Google Scholar] [CrossRef]
- Moe-Lange, J.; Gappel, N.M.; Machado, M.; Wudick, M.M.; Sies, C.S.A.; Schott-Verdugo, S.N.; Bonus, M.; Mishra, S.; Hartwig, T.; Bezrutczyk, M.; et al. Interdependence of a mechanosensitive anion channel and glutamate receptors in distal wound signaling. Sci. Adv. 2021, 7, eabg4298. [Google Scholar] [CrossRef]
- Miller, G.; Schlauch, K.; Tam, R.; Cortes, D.; Torres, M.A.; Shulaev, V.; Dangl, J.L.; Mittler, R. The plant NADPH oxidase RBOHD mediates rapid systemic signaling in response to diverse stimuli. Sci. Signal. 2009, 2, ra45. [Google Scholar] [CrossRef] [Green Version]
- Block, A.; Christensen, S.A.; Hunter, C.T.; Alborn, H.T. Herbivore-derived fatty-acid amides elicit reactive oxygen species burst in plants. J. Exp. Bot. 2017, 69, 1235–1245. [Google Scholar] [CrossRef]
- Fichman, Y.; Mittler, R. Rapid systemic signaling during abiotic and biotic stresses: Is the ROS wave master of all trades? Plant J. 2020, 102, 887–896. [Google Scholar] [CrossRef] [Green Version]
- Gilroy, S.; Białasek, M.; Suzuki, N.; Górecka, M.; Devireddy, A.R.; Karpiński, S.; Mittler, R. ROS, calcium, and electric signals: Key mediators of rapid systemic signaling in plants. Plant Physiol. 2016, 171, 1606–1615. [Google Scholar] [CrossRef]
- Choi, W.G.; Miller, G.; Wallace, I.; Harper, J.; Mittler, R.; Gilroy, S. Orchestrating rapid long-distance signaling in plants with Ca2+, ROS and electrical signals. Plant J. Cell Mol. Biol. 2017, 90, 698–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandhi, A.; Kariyat, R.; Harikishore, A.; Ayati, M.; Bhunia, A.; Sahoo, N. Deciphering the role of ion channels in early defense signaling against herbivorous insects. Cells 2021, 10, 23. [Google Scholar] [CrossRef]
- Maffei, M.E.; Mithofer, A.; Arimura, G.I.; Uchtenhagen, H.; Bossi, S.; Bertea, C.M.; Cucuzza, L.S.; Novero, M.; Volpe, V.; Quadro, S.; et al. Effects of feeding Spodoptera littoralis on lima bean leaves. III. Membrane depolarization and involvement of hydrogen peroxide. Plant Physiol. 2006, 140, 1022–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, G.W.; Wang, X.F.; Chen, D.; Wang, X.W.; Liu, X.D. Effects of aphids Myzus persicae on the changes of Ca2+ and H2O2 flux and enzyme activities in tobacco. J. Plant Interact. 2014, 9, 883–888. [Google Scholar] [CrossRef]
- Vega-Munoz, I.; Duran-Flores, D.; Fernandez-Fernandez, A.D.; Heyman, J.; Ritter, A.; Stael, S. Breaking bad news: Dynamic molecular mechanisms of wound response in plants. Front. Plant Sci. 2020, 11, 26. [Google Scholar] [CrossRef]
- Wu, X.; Wang, Y.; Bian, Y.; Ren, Y.; Xu, X.; Zhou, F.; Ding, H. A critical review on plant annexin: Structure, function, and mechanism. Plant Physiol. Biochem. 2022, 190, 81–89. [Google Scholar] [PubMed]
- Lippert, D.N.; Ralph, S.G.; Phillips, M.; White, R.; Smith, D.; Hardie, D.; Gershenzon, J.; Ritland, K.; Borchers, C.H.; Bohlmann, J. Quantitative iTRAQ proteome and comparative transcriptome analysis of elicitor-induced Norway spruce (Picea abies) cells reveals elements of calcium signaling in the early conifer defense response. Proteomics 2009, 9, 350–367. [Google Scholar]
- Huang, H.J.; Zhang, C.X.; Hong, X.Y. How does saliva function in planthopper-host interactions? Arch. Insect Biochem. Physiol. 2019, 100, 7. [Google Scholar]
- Will, T.; Furch, A.C.U.; Zimmermann, M.R. How phloem-feeding insects face the challenge of phloem-located defenses. Front. Plant Sci. 2013, 4, 12. [Google Scholar] [CrossRef] [Green Version]
- Medina-Ortega, K.J.; Walker, G.P. Faba bean forisomes can function in defence against generalist aphids. Plant Cell Environ. 2015, 38, 1167–1177. [Google Scholar] [CrossRef] [Green Version]
- Zebelo, S.A.; Maffei, M.E. The ventral eversible gland (VEG) of Spodoptera littoralis triggers early responses to herbivory in Arabidopsis thaliana. Arthropod-Plant Interact. 2012, 6, 543–551. [Google Scholar] [CrossRef]
- Fu, J.M.; Shi, Y.; Wang, L.; Zhang, H.; Li, J.; Fang, J.C.; Ji, R. Planthopper-secreted salivary disulfide isomerase activates immune responses in plants. Front. Plant Sci. 2021, 11, 13. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, A.; Kariyat, R.R.; Chappa, C.; Tayal, M.; Sahoo, N. Tobacco hornworm (manduca sexta) oral secretion elicits reactive oxygen species in isolated tomato protoplasts. Int. J. Mol. Sci. 2020, 21, 14. [Google Scholar] [CrossRef] [PubMed]
- Barbero, F.; Guglielmotto, M.; Capuzzo, A.; Maffei, M.E. Extracellular self-DNA (esDNA), but not heterologous plant or insect DNA (etDNA), induces plasma membrane depolarization and calcium signaling in lima bean (Phaseolus lunatus) and maize (Zea mays). Int. J. Mol. Sci. 2016, 17, 1659. [Google Scholar] [CrossRef] [Green Version]
- Barbero, F.; Guglielmotto, M.; Islam, M.; Maffei, M.E. Extracellular fragmented self-DNA is involved in plant responses to biotic stress. Front. Plant Sci. 2021, 12, 17. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Ning, Y.Z.; Wang, L.M.; Wilkins, K.A.; Davies, J.M. Damage signaling by extracellular nucleotides: A role for cyclic nucleotides in elevating cytosolic free calcium? Front. Plant Sci. 2021, 12, 8. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.E.; Gertsch, J.; Appendino, G. Plant volatiles: Production, function and pharmacology. Nat. Prod. Rep. 2011, 28, 1359–1380. [Google Scholar] [CrossRef]
- Zebelo, S.A.; Matsui, K.; Ozawa, R.; Maffei, M.E. Plasma membrane potential depolarization and cytosolic calcium flux are early events involved in tomato (Solanum lycopersicon) plant-to-plant communication. Plant Sci. 2012, 196, 93–100. [Google Scholar] [CrossRef]
- Jing, T.T.; Qian, X.N.; Du, W.K.; Gao, T.; Li, D.F.; Guo, D.Y.; He, F.; Yu, G.M.; Li, S.P.; Schwab, W.; et al. Herbivore-induced volatiles influence moth preference by increasing the beta-ocimene emission of neighbouring tea plants. Plant Cell Environ. 2021, 44, 3667–3680. [Google Scholar] [CrossRef]
- Jiao, C.; Gong, J.; Guo, Z.; Li, S.; Zuo, Y.; Shen, Y. Linalool activates oxidative and calcium burst and CAM3-ACA8 participates in calcium recovery in arabidopsis leaves. Int. J. Mol. Sci. 2022, 23, 5357. [Google Scholar] [CrossRef]
- Brosset, A.; Islam, M.; Bonzano, S.; Maffei, M.E.; Blande, J.D. Exposure to (Z)-11-hexadecenal (Z)-11-16:Ald increases Brassica nigra susceptibility to subsequent herbivory. Sci. Rep. 2021, 11, 12. [Google Scholar]
- Steinhorst, L.; Kudla, J. Signaling in cells and organisms—Calcium holds the line. Curr. Opin. Plant Biol. 2014, 22, 14–21. [Google Scholar] [CrossRef]
- Yap, K.L.; Ames, J.B.; Swindells, M.B.; Ikura, M. Diversity of conformational states and changes within the EF-hand protein superfamily. Proteins 1999, 37, 499–507. [Google Scholar] [CrossRef]
- Garzo, E.; Fernandez-Pascual, M.; Morcillo, C.; Fereres, A.; Gomez-Guillamon, M.L.; Tjallingii, W.F. Ultrastructure of compatible and incompatible interactions in phloem sieve elements during the stylet penetration by cotton aphids in melon. Insect Sci. 2018, 25, 631–642. [Google Scholar] [CrossRef]
- Tian, T.; Ji, R.; Fu, J.M.; Li, J.; Wang, L.; Zhang, H.; Yang, S.Y.; Ye, W.F.; Fang, J.C.; Zhu-Salzman, K. A salivary calcium-binding protein from Laodelphax striatellus acts as an effector that suppresses defense in rice. Pest Manag. Sci. 2021, 77, 2272–2281. [Google Scholar] [CrossRef]
- Wu, W.; Yi, G.; Lv, X.W.; Mao, Q.Z.; Wei, T.Y. A leafhopper saliva protein mediates horizontal transmission of viral pathogens from insect vectors into rice phloem. Commun. Biol. 2022, 5, 13. [Google Scholar] [CrossRef]
- Liu, X.Q.; Zhou, H.Y.; Zhao, J.; Hua, H.X.; He, Y.P. Identification of the secreted watery saliva proteins of the rice brown planthopper, Nilaparvata lugens (Stal) by transcriptome and Shotgun LC-MS/MS approach. J. Insect Physiol. 2016, 89, 60–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, W.F.; Yu, H.X.; Jian, Y.K.; Zeng, J.M.; Ji, R.; Chen, H.D.; Lou, Y.G. A salivary EF-hand calcium-binding protein of the brown planthopper Nilaparvata lugens functions as an effector for defense responses in rice. Sci. Rep. 2017, 7, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Wang, L.; Li, J.; Yang, W.; Ci, J.; Ren, X.; Wang, W.; Wang, Y.; Jiang, L.; Yang, W. Identification and expression analysis revealed drought stress-responsive Calmodulin and Calmodulin-like genes in maize. J. Plant Interact. 2022, 17, 450–461. [Google Scholar] [CrossRef]
- Zhu, X.; Dunand, C.; Snedden, W.; Galaud, J.P. CaM and CML emergence in the green lineage. Trends Plant Sci. 2015, 20, 483–489. [Google Scholar] [CrossRef]
- McCormack, E.; Tsai, Y.C.; Braam, J. Handling calcium signaling: Arabidopsis CaMs and CMLs. Trends Plant Sci. 2005, 10, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.M.; Shi, Y.; Wang, L.H.; Tian, T.; Li, J.; Gong, L.; Zheng, Z.T.; Jing, M.F.; Fang, J.C.; Ji, R. Planthopper-secreted salivary calmodulin acts as an effector for defense responses in rice. Front. Plant Sci. 2022, 13, 14. [Google Scholar] [CrossRef] [PubMed]
- Pamplona, R.S.; Kim, J.; Lee, J.W.; Kim, C.S.; Boo, K.H. Comparative transcriptome analysis of spinach in response to insect herbivory. Plant Biotechnol. Rep. 2022, 16, 43–55. [Google Scholar] [CrossRef]
- Jiao, C.; Li, K.; Zuo, Y.; Gong, J.; Guo, Z.; Shen, Y. CALMODULIN1 and WRKY53 function in plant defense by negatively regulating the jasmonic acid biosynthesis pathway in Arabidopsis. Int. J. Mol. Sci. 2022, 23, 7718. [Google Scholar] [CrossRef]
- DeFalco, T.A.; Marshall, C.B.; Munro, K.; Kang, H.G.; Moeder, W.; Ikura, M.; Snedden, W.A.; Yoshioka, K. Multiple calmodulin-binding sites positively and negatively regulate arabidopsis CYCLIC NUCLEOTIDE-GATED CHANNEL12. Plant Cell 2016, 28, 1738–1751. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Zhang, Y.; Tang, S.; Pan, J.; Yu, Y.; Han, J.; Li, Y.; Du, X.; Nan, Z.; Sun, Q. AtCNGC2 is involved in jasmonic acid-induced calcium mobilization. J. Exp. Bot. 2016, 67, 809–819. [Google Scholar] [CrossRef] [Green Version]
- Heyer, M.; Scholz, S.S.; Voigt, D.; Reichelt, M.; Aldon, D.; Oelmuller, R.; Boland, W.; Mithofer, A. Herbivory-responsive calmodulin-like protein CML9 does not guide jasmonate-mediated defenses in Arabidopsis thaliana. PLoS ONE 2018, 13, e0197633. [Google Scholar] [CrossRef] [Green Version]
- Yadav, M.; Pandey, J.; Chakraborty, A.; Hassan, M.I.; Kundu, J.K.; Roy, A.; Singh, I.K.; Singh, A. A comprehensive analysis of calmodulin-like proteins of glycine max indicates their role in calcium signaling and plant defense against insect attack. Front. Plant Sci. 2022, 13, 18. [Google Scholar] [CrossRef]
- Scholz, S.S.; Reichelt, M.; Vadassery, J.; Mithöfer, A. Calmodulin-like protein CML37 is a positive regulator of ABA during drought stress in Arabidopsis. Plant Signal. Behav. 2015, 10, e1011951. [Google Scholar] [CrossRef]
- Vadassery, J.; Reichelt, M.; Hause, B.; Gershenzon, J.; Boland, W.; Mithofer, A. CML42-mediated calcium signaling coordinates responses to Spodoptera herbivory and abiotic stresses in Arabidopsis. Plant Physiol. 2012, 159, 1159–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyer, M.; Scholz, S.S.; Reichelt, M.; Kunert, G.; Oelmuller, R.; Mithofer, A. The Ca2+ sensor proteins CML37 and CML42 antagonistically regulate plant stress responses by altering phytohormone signals. Plant Mol. Biol. 2022, 109, 611–625. [Google Scholar] [CrossRef]
- Harper, J.F.; Sussman, M.R.; Schaller, G.E.; Putnam-Evans, C.; Charbonneau, H.; Harmon, A.C. A calcium-dependent protein kinase with a regulatory domain similar to calmodulin. Science 1991, 252, 951–954. [Google Scholar] [CrossRef]
- Boudsocq, M.; Droillard, M.J.; Regad, L.; Laurière, C. Characterization of arabidopsis calcium-dependent protein kinases: Activated or not by calcium? Biochem. J. 2012, 447, 291–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanchiswamy, C.N.; Muroi, A.; Maffei, M.E.; Yoshioka, H.; Sawasaki, T.; Arimura, G. Ca2+-dependent protein kinases and their substrate HsfB2a are differently involved in the heat response signaling pathway in Arabidopsis. Plant Biotechnol. 2010, 27, 469–473. [Google Scholar] [CrossRef] [Green Version]
- Kanchiswamy, C.N.; Takahashi, H.; Quadro, S.; Maffei, M.E.; Bossi, S.; Bertea, C.; Zebelo, S.A.; Muroi, A.; Ishihama, N.; Yoshioka, H.; et al. Regulation of Arabidopsis defense responses against Spodoptera littoralis by CPK-mediated calcium signaling. BMC Plant Biol. 2010, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Arimura, G.; Maffei, M.E. Calcium and secondary CPK signaling in plants in response to herbivore attack. Biochem. Biophys. Res. Commun. 2010, 400, 455–460. [Google Scholar] [CrossRef]
- Yang, D.H.; Hettenhausen, C.; Baldwin, I.T.; Wu, J.Q. Silencing Nicotiana attenuata calcium-dependent protein kinases, CDPK4 and CDPK5, strongly up-regulates wound- and herbivory-induced jasmonic acid accumulations. Plant Physiol. 2012, 159, 1591–1607. [Google Scholar] [CrossRef] [Green Version]
- Kanchiswamy, C.N.; Mohanta, T.K.; Capuzzo, A.; Occhipinti, A.; Verrillo, F.; Maffei, M.E.; Malnoy, M. Differential expression of CPKs and cytosolic Ca2+ variation in resistant and susceptible apple cultivars (Malus × domestica) in response to the pathogen Erwinia amylovora and mechanical wounding. BMC Genom. 2013, 14, 14. [Google Scholar] [CrossRef] [Green Version]
- Hettenhausen, C.; Sun, G.L.; He, Y.B.; Zhuang, H.F.; Sun, T.; Qi, J.F.; Wu, J.Q. Genome-wide identification of calcium-dependent protein kinases in soybean and analyses of their transcriptional responses to insect herbivory and drought stress. Sci. Rep. 2016, 6, 14. [Google Scholar] [CrossRef]
- Sánchez-Barrena, M.J.; Martínez-Ripoll, M.; Albert, A. Structural biology of a major signaling network that regulates plant abiotic stress: The CBL-CIPK mediated pathway. Int. J. Mol. Sci. 2013, 14, 5734–5749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kundu, P.; Nehra, A.; Gill, R.; Tuteja, N.; Gill, S.S. Unraveling the importance of EF-hand mediated calcium signaling in plants. S. Afr. J. Bot. 2022, 148, 615–633. [Google Scholar] [CrossRef]
- Kim, K.N.; Cheong, Y.H.; Gupta, R.; Luan, S. Interaction specificity of Arabidopsis calcineurin B-like calcium sensors and their target kinases. Plant Physiol. 2000, 124, 1844–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanyal, S.K.; Pandey, A.; Pandey, G.K. The CBL—CIPK signaling module in plants: A mechanistic perspective. Physiol. Plant. 2015, 155, 89–108. [Google Scholar] [CrossRef] [PubMed]
- Luan, S. The CBL—CIPK network in plant calcium signaling. Trends Plant Sci. 2009, 14, 37–42. [Google Scholar] [CrossRef]
- Ma, X.; Li, Q.-H.; Yu, Y.-N.; Qiao, Y.-M.; Haq, S.; Gong, Z.-H. The CBL–CIPK Pathway in Plant Response to Stress Signals. Int. J. Mol. Sci. 2020, 21, 5668. [Google Scholar] [CrossRef]
- Mao, J.; Mo, Z.; Yuan, G.; Xiang, H.; Visser, R.G.F.; Bai, Y.; Liu, H.; Wang, Q.; van der Linden, C.G. The CBL-CIPK network is involved in the physiological crosstalk between plant growth and stress adaptation. Plant Cell Environ. 2022, 11. [Google Scholar] [CrossRef]
- Su, W.H.; Huang, L.; Ling, H.; Mao, H.Y.; Huang, N.; Su, Y.C.; Ren, Y.J.; Wang, D.J.; Xu, L.P.; Muhammad, K.; et al. Sugarcane calcineurin B-like (CBL) genes play important but versatile roles in regulation of responses to biotic and abiotic stresses. Sci. Rep. 2020, 10, 13. [Google Scholar] [CrossRef] [Green Version]
- Aslam, M.; Fakher, B.; Jakada, B.H.; Zhao, L.H.; Cao, S.J.; Cheng, Y.; Qin, Y. Genome-wide identification and expression profiling of CBL-CIPK gene family in pineapple (Ananas comosus) and the role of AcCBL1 in abiotic and biotic stress response. Biomolecules 2019, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.M.; Xie, S.X.; Wang, L.; Jing, S.L.; Zhu, X.P.; Li, X.W.; Zeng, W.; Yuan, H.Y. Identification of up-regulated genes in tea leaves under mild infestation of green leafhopper. Sci. Hortic. 2011, 130, 476–481. [Google Scholar] [CrossRef]
- Gerke, V.; Moss, S.E. Annexins: From structure to function. Physiol. Rev. 2002, 82, 331–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laohavisit, A.; Davies, J.M. Multifunctional annexins. Plant Sci. 2009, 177, 532–539. [Google Scholar] [CrossRef]
- Deka, S.; Barthakur, S. Impact of biorational pesticides and response of annexin gene expression in tomato (Solanum lycopersicum) and capsicum (Capsicum annum L.) in biotic stress management under greenhouse condition. Ann. Agric. Res. N. Ser. 2015, 36, 233–242. [Google Scholar]
- Wu, X.; Yan, J.; Wu, Y.; Zhang, H.; Mo, S.; Xu, X.; Zhou, F.; Ding, H. Proteomic analysis by iTRAQ-PRM provides integrated insight into mechanisms of resistance in pepper to Bemisia tabaci (Gennadius). BMC Plant Biol. 2019, 19, 270. [Google Scholar] [CrossRef]
- Zhao, J.; Li, L.; Liu, Q.; Liu, P.; Li, S.; Yang, D.; Chen, Y.; Pagnotta, S.; Favery, B.; Abad, P.; et al. A MIF-like effector suppresses plant immunity and facilitates nematode parasitism by interacting with plant annexins. J. Exp. Bot. 2019, 70, 5943–5958. [Google Scholar] [CrossRef]
- Mortimer, J.C.; Laohavisit, A.; Macpherson, N.; Webb, A.; Brownlee, C.; Battey, N.H.; Davies, J.M. Annexins: Multifunctional components of growth and adaptation. J. Exp. Bot. 2008, 59, 533–544. [Google Scholar] [CrossRef] [Green Version]
- Konopka-Postupolska, D.; Clark, G.; Hofmann, A. Structure, function and membrane interactions of plant annexins: An update. Plant Sci. 2011, 181, 230–241. [Google Scholar] [CrossRef] [Green Version]
- Demidchik, V. ROS-activated ion channels in plants: Biophysical characteristics, physiological functions and molecular nature. Int. J. Mol. Sci. 2018, 19, 270. [Google Scholar] [CrossRef] [Green Version]
- Hettenhausen, C.; Heinrich, M.; Baldwin, I.T.; Wu, J.Q. Fatty acid-amino acid conjugates are essential for systemic activation of salicylic acid-induced protein kinase and accumulation of jasmonic acid in Nicotiana attenuata. BMC Plant Biol. 2014, 14, 12. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.Q.; Baldwin, I.T. Herbivory-induced signalling in plants: Perception and action. Plant Cell Environ. 2009, 32, 1161–1174. [Google Scholar] [CrossRef]
- Heinrich, M.; Baldwin, I.T.; Wu, J.Q. Two mitogen-activated protein kinase kinases, MKK1 and MEK2, are involved in wounding- and specialist lepidopteran herbivore Manduca sexta-induced responses in Nicotiana attenuata. J. Exp. Bot. 2011, 62, 4355–4365. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, M.; Baldwin, I.T.; Wu, J.Q. Three MAPK kinases, MEK1, SIPKK, and NPK2, are not involved in activation of SIPK after wounding and herbivore feeding but important for accumulation of trypsin proteinase inhibitors. Plant Mol. Biol. Rep. 2012, 30, 731–740. [Google Scholar] [CrossRef]
- Meldau, S.; Ullman-Zeunert, L.; Govind, G.; Bartram, S.; Baldwin, I.T. MAPK-dependent JA and SA signalling in Nicotiana attenuata affects plant growth and fitness during competition with conspecifics. BMC Plant Biol. 2012, 12, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, I.K.; Kumar, S.; Singh, S.; Singh, A. Expression profiling of mitogen-activated protein kinase genes from chickpea (Cicer arietinum L.) in response to Helicoverpa armigera, wounding and signaling compounds. J. Asia-Pac. Entomol. 2017, 20, 942–948. [Google Scholar] [CrossRef]
- Radhika Keshan; Ragini; Singh, I.K.; Singh, A. Genome wide investigation of MAPKKKs from Cicer arietinum and their involvement in plant defense against Helicoverpa armigera. Physiol. Mol. Plant Pathol. 2021, 115, 13. [Google Scholar] [CrossRef]
- Giacometti, R.; Ilina, N.; Eduardo, P.A.; Zavala, J.A. Stink bug nezara viridula sustains late MAPKs phosphorylation status and induces expression of genes related with cell wall rearrangement in developing soybean seeds. Arthropod-Plant Interact. 2018, 12, 531–541. [Google Scholar] [CrossRef]
- Zhou, S.X.; Chen, M.T.; Zhang, Y.B.; Gao, Q.; Noman, A.; Wang, Q.; Li, H.; Chen, L.; Zhou, P.Y.; Lu, J.; et al. OsMKK3, a stress-responsive protein kinase, positively regulates rice resistance to Nilaparvata lugens via phytohormone dynamics. Int. J. Mol. Sci. 2019, 20, 15. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.Y.; Gao, J.; Zhang, Y.R.; Tian, J.Y.; Sun, Y.; Wang, C.Y. RNA-Seq identification of candidate defense genes by analyzing Mythimna separata feeding-damage induced systemic resistance in balsas teosinte. Pest Manag. Sci. 2020, 76, 333–342. [Google Scholar] [CrossRef]
- Khan, A.L.; Asaf, S.; Khan, A.; Khan, A.; Imran, M.; Al-Harrasi, A.; Lee, I.J.; Al-Rawahi, A. Transcriptomic analysis of dubas bug (Ommatissus lybicus bergevin) infestation to date palm. Sci. Rep. 2020, 10, 15. [Google Scholar] [CrossRef]
- Sozen, C.; Schenk, S.T.; Boudsocq, M.; Chardin, C.; Almeida-Trapp, M.; Krapp, A.; Hirt, H.; Mithofer, A.; Colcombet, J. Wounding and insect feeding trigger two independent MAPK pathways with distinct regulation and kinetics. Plant Cell 2020, 32, 1988–2003. [Google Scholar] [CrossRef] [Green Version]
- Tsien, R.Y. New calcium indicators and buffers with high selectivity against magnesium and protons: Design, synthesis, and properties of prototype structures. Biochemistry 1980, 19, 2396–2404. [Google Scholar] [CrossRef] [PubMed]
- Grenzi, M.; Resentini, F.; Vanneste, S.; Zottini, M.; Bassi, A.; Costa, A. Illuminating the hidden world of calcium ions in plants with a universe of indicators. Plant Physiol. 2021, 187, 550–571. [Google Scholar] [CrossRef] [PubMed]
- Wiederschain, G.Y. The molecular probes handbook. A guide to fluorescent probes and labeling technologies. Biochemistry 2011, 76, 1276–1277. [Google Scholar] [CrossRef] [Green Version]
- Hilleary, R.; Choi, W.-G.; Kim, S.-H.; Lim, S.D.; Gilroy, S. Sense and sensibility: The use of fluorescent protein-based genetically encoded biosensors in plants. Curr. Opin. Plant Biol. 2018, 46, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.E.; Qin, Y.; Park, J.G.; McCombs, J.E. Design and application of genetically encoded biosensors. Trends Biotechnol. 2011, 29, 144–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridgway, E.B.; Ashley, C.C. Calcium transients in single muscle fibers. Biochem. Biophys. Res Commun. 1967, 29, 229–234. [Google Scholar] [CrossRef]
- Zhu, X.; Feng, Y.; Liang, G.; Liu, N.; Zhu, J.-K. Aequorin-based luminescence imaging reveals stimulus- and tissue-specific Ca2+ dynamics in arabidopsis plants. Mol. Plant 2013, 6, 444–455. [Google Scholar] [CrossRef] [Green Version]
- Vatsa, P.; Chiltz, A.; Bourque, S.; Wendehenne, D.; Garcia-Brugger, A.; Pugin, A. Involvement of putative glutamate receptors in plant defence signaling and NO production. Biochimie 2011, 93, 2095–2101. [Google Scholar] [CrossRef]
- Souza, C.D.A.; Li, S.; Lin, A.Z.; Boutrot, F.; Grossmann, G.; Zipfel, C.; Somerville, S.C. Cellulose-derived oligomers act as damage-associated molecular patterns and trigger defense-like responses. Plant Physiol. 2017, 173, 2383–2398. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, C.T.; Kurenda, A.; Stolz, S.; Chételat, A.; Farmer, E.E. Identification of cell populations necessary for leaf-to-leaf electrical signaling in a wounded plant. Proc. Natl. Acad. Sci. USA 2018, 115, 10178–10183. [Google Scholar] [CrossRef] [Green Version]
- Ruberti, C.; Feitosa-Araujo, E.; Xu, Z.; Wagner, S.; Grenzi, M.; Darwish, E.; Lichtenauer, S.; Fuchs, P.; Parmagnani, A.S.; Balcerowicz, D.; et al. MCU proteins dominate in vivo mitochondrial Ca2+ uptake in Arabidopsis roots. Plant Cell 2022. Online ahead of print. [Google Scholar] [CrossRef]
- Lavanya, S.N.; Niranjan-Raj, S.; Jadimurthy, R.; Sudarsan, S.; Srivastava, R.; Tarasatyavati, C.; Rajashekara, H.; Gupta, V.K.; Nayaka, S.C. Immunity elicitors for induced resistance against the downy mildew pathogen in pearl millet. Sci. Rep. 2022, 12, 4078. [Google Scholar] [CrossRef]
- Verrillo, F.; Occhipinti, A.; Kanchiswamy, C.N.; Maffei, M.E. Quantitative analysis of herbivore-induced cytosolic calcium by using a Cameleon (YC 3.6) calcium sensor in Arabidopsis thaliana. J. Plant Physiol. 2014, 171, 136–139. [Google Scholar] [CrossRef] [PubMed]
- Krogman, W.; Sparks, J.A.; Blancaflor, E.B. Cell type-specific imaging of calcium signaling in Arabidopsis thaliana seedling roots using GCaMP3. Int. J. Mol. Sci. 2020, 21, 6385. [Google Scholar] [CrossRef]
- Keinath, N.F.; Waadt, R.; Brugman, R.; Schroeder, J.I.; Grossmann, G.; Schumacher, K.; Krebs, M. Live cell imaging with R-GECO1 sheds light on flg22- and chitin-induced transient [Ca2+]cyt patterns in Arabidopsis. Mol. Plant 2015, 8, 1188–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagai, T.; Yamada, S.; Tominaga, T.; Ichikawa, M.; Miyawaki, A. Expanded dynamic range of fluorescent indicators for Ca2+ by circularly permuted yellow fluorescent proteins. Proc. Natl. Acad. Sci. USA 2004, 101, 10554–10559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanchiswamy, C.N.; Malnoy, M.; Occhipinti, A.; Maffei, M.E. Calcium imaging perspectives in plants. Int. J. Mol. Sci. 2014, 15, 3842–3859. [Google Scholar] [CrossRef] [Green Version]
- Read, N.D.; Allan, W.T.G.; Knight, H.; Knight, M.R.; Malho, R.; Russell, A.; Shacklock, P.S.; Trewavas, A.J. Imaging and measurement of cytosolic free calcium in plant and fungal cells. J. Microsc. 1992, 166, 57–86. [Google Scholar] [CrossRef]
- Allan, C.; Morris, R.J.; Meisrimler, C.N. Encoding, transmission, decoding, and specificity of calcium signals in plants. J. Exp. Bot. 2022, 73, 3372–3385. [Google Scholar] [CrossRef] [PubMed]
- Grossmann, G.; Krebs, M.; Maizel, A.; Stahl, Y.; Vermeer, J.E.M.; Ott, T. Green light for quantitative live-cell imaging in plants. J. Cell Sci. 2018, 131, 14. [Google Scholar] [CrossRef] [Green Version]
- Webb, S.E.; Karplus, E.; Miller, A.L. Retrospective on the development of aequorin and aequorin-based imaging to visualize changes in intracellular free Ca2+. Mol. Reprod. Dev. 2015, 82, 563–586. [Google Scholar] [CrossRef]
- Mithöfer, A.; Mazars, C.; Maffei, M.E. Probing spatio-temporal intracellular calcium variations in plants. Methods Mol. Biol. 2009, 479, 79–92. [Google Scholar] [PubMed]
- Uemura, T.; Wang, J.Q.; Aratani, Y.; Gilroy, S.; Toyota, M. Wide-field, real-time imaging of local and systemic wound signals in Arabidopsis. J. Vis. Exp. 2021, 172, e62114. [Google Scholar] [CrossRef] [PubMed]
- Vincent, T.R.; Avramova, M.; Canham, J.; Higgins, P.; Bilkey, N.; Mugford, S.T.; Pitino, M.; Toyota, M.; Gilroy, S.; Miller, A.J.; et al. Interplay of plasma membrane and vacuolar ion channels, together with BAK1, elicits rapid cytosolic calcium elevations in Arabidopsis during aphid feeding. Plant Cell 2017, 29, 1460–1479. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gong, Q.; Huang, F.; He, L.; Liu, Y. Live imaging and quantitation of insect feeding-induced Ca2+ signal using GCaMP3-based system in Nicotiana benthamiana. STAR Protoc. 2022, 3, 101040. [Google Scholar] [CrossRef]
- Mousavi, S.A.R.; Chauvin, A.; Pascaud, F.; Kellenberger, S.; Farmer, E.E. GLUTAMATE RECEPTOR-LIKE genes mediate leaf-to-leaf wound signalling. Nature 2013, 500, 422–426. [Google Scholar] [CrossRef]
Figure 1.
Framework of early events induced in responses to herbivory within the first minutes after infestation. Herbivory is the combination of tissue damage (wounding) and the delivery of insect oral secretions. Both herbivory and mechanical damage induce the emission of volatile organic compounds (VOCs), which depolarize the plasma membrane potential (Vm). Wounding induces variations in the water potential that affect the turgor pressure (ΨT), which is perceived by plasma-membrane located mechanosensitive ion channels (MSC) resulting in Vm depolarization. Type IIB auto-inhibited Ca2+-ATPases (ACAs) play a role in recovery of excitability after long-term herbivory by interfering with both the Vm and [Ca2+]cyt. Mechanical wounding does not exert a direct effect on Vm, Ca2+ and K+ channel activity (red crosses). Herbivory delivers oral secretions that exert different effects. Upon herbivory receptors activate a plasma-membrane–localized cyclic nucleotide-gated ion channel (CNGC) which interacts with the Ca2+-sensor calmodulin (CaM). Oral secretions inhibit the H+-ATPase activity by hampering the association between 14-3-3 protein and H+-ATPase proteins and cause a reduced phosphohydrolitic activity of the proton pump with a reduced extrusion of H+ from the cytosol and an alkalinization of the apoplast (pHAPO), both concurring to the Vm depolarization. Elicitors from oral secretions like N-linolenoyl-l-glutamine (GLN18:3) induce a RBOHD and RBOHF-dependent ROS burst. Specific oligosaccharides present in the oral secretions interact with plasma membrane receptors that also trigger ROS and Ca2+ signaling. Extracellular DNA (eDNA), deriving from cell disruption made by herbivory, and extracellular ATP (eATP) released by wounding and herbivory increase [Ca2+]cyt. In summary, oral secretions, wounding, ΨT reduction, pHAPO alkalinization, an increase in apoplastic amino acids (AAapo) and glutamate (Glu) and the activity of the glutamate receptor-like cation channels (GLRs) depolarize the Vm. Oral secretions and ROS/NRS activate the Ca2+ channel that in turn opens the inward rectifying K+ channel that reduces the cytosolic pH causing Vm depolarization. Connections supported by empirical studies in planta are represented by solid lines, hypothetical connections are shown in dashed lines.
Figure 1.
Framework of early events induced in responses to herbivory within the first minutes after infestation. Herbivory is the combination of tissue damage (wounding) and the delivery of insect oral secretions. Both herbivory and mechanical damage induce the emission of volatile organic compounds (VOCs), which depolarize the plasma membrane potential (Vm). Wounding induces variations in the water potential that affect the turgor pressure (ΨT), which is perceived by plasma-membrane located mechanosensitive ion channels (MSC) resulting in Vm depolarization. Type IIB auto-inhibited Ca2+-ATPases (ACAs) play a role in recovery of excitability after long-term herbivory by interfering with both the Vm and [Ca2+]cyt. Mechanical wounding does not exert a direct effect on Vm, Ca2+ and K+ channel activity (red crosses). Herbivory delivers oral secretions that exert different effects. Upon herbivory receptors activate a plasma-membrane–localized cyclic nucleotide-gated ion channel (CNGC) which interacts with the Ca2+-sensor calmodulin (CaM). Oral secretions inhibit the H+-ATPase activity by hampering the association between 14-3-3 protein and H+-ATPase proteins and cause a reduced phosphohydrolitic activity of the proton pump with a reduced extrusion of H+ from the cytosol and an alkalinization of the apoplast (pHAPO), both concurring to the Vm depolarization. Elicitors from oral secretions like N-linolenoyl-l-glutamine (GLN18:3) induce a RBOHD and RBOHF-dependent ROS burst. Specific oligosaccharides present in the oral secretions interact with plasma membrane receptors that also trigger ROS and Ca2+ signaling. Extracellular DNA (eDNA), deriving from cell disruption made by herbivory, and extracellular ATP (eATP) released by wounding and herbivory increase [Ca2+]cyt. In summary, oral secretions, wounding, ΨT reduction, pHAPO alkalinization, an increase in apoplastic amino acids (AAapo) and glutamate (Glu) and the activity of the glutamate receptor-like cation channels (GLRs) depolarize the Vm. Oral secretions and ROS/NRS activate the Ca2+ channel that in turn opens the inward rectifying K+ channel that reduces the cytosolic pH causing Vm depolarization. Connections supported by empirical studies in planta are represented by solid lines, hypothetical connections are shown in dashed lines.

Figure 2.
Calcium binding proteins. The top cartoon illustrates the canonical EF-hand helix–loop–helix. EF-hand Ca2+ binding motif contains a 29-residue helix–loop–helix topology, much like the spread thumb and forefinger of the human hand. Calmodulin is shown with its four calcium sites (shown as green balls) occupied. Calcium binding proteins are major players in Ca2+ signaling. See text for more details.
Figure 2.
Calcium binding proteins. The top cartoon illustrates the canonical EF-hand helix–loop–helix. EF-hand Ca2+ binding motif contains a 29-residue helix–loop–helix topology, much like the spread thumb and forefinger of the human hand. Calmodulin is shown with its four calcium sites (shown as green balls) occupied. Calcium binding proteins are major players in Ca2+ signaling. See text for more details.

Figure 3.
Calcium signaling upon herbivore feeding. (A–F) false colors time-course imaging of Ca2+ release upon feeding with the insect Spodoptera littoralis on a rosette leaf of Arabidopsis thaliana expressing the biosensor R-GECO1. The rapid fluorescence variation (blue to green) indicates the increase in the cytosolic Ca2+ concentration following the insect feeding. A wave of Ca2+ is generated at the site of feeding and rapidly spreads through the vascular system. The acquisitions have been obtained by a fluorescence stereo microscope Nikon SMZ18 with a SHR PLAN APO 0.5X WD:71. Excitation light was produced by a fluorescent CoolLED pE-300 ultra at 580 nm. Images were collected with a Mono Camera Nikon DS-Fi3 camera. Exposure time was set to 1 sec with a resolution of 8 bit 1440 × 1024. Images were acquired every 5 sec (See also Video S1). (G,H) Confocal laser scanning micrographs showing the increased [Ca2+]cyt in Lima bean (Phaseolus lunatus) leaves upon feeding by S. littoralis. The red fluorescence is associated to the chlorophyll present in the chloroplasts, whereas the green fluorescence indicates the cytosolic localization of Ca2+ by the indicator Calcium OrangeTM. Scale bars: G = 100 µm; H = 200 µm (Figures (G,H) by Massimo Maffei).
Figure 3.
Calcium signaling upon herbivore feeding. (A–F) false colors time-course imaging of Ca2+ release upon feeding with the insect Spodoptera littoralis on a rosette leaf of Arabidopsis thaliana expressing the biosensor R-GECO1. The rapid fluorescence variation (blue to green) indicates the increase in the cytosolic Ca2+ concentration following the insect feeding. A wave of Ca2+ is generated at the site of feeding and rapidly spreads through the vascular system. The acquisitions have been obtained by a fluorescence stereo microscope Nikon SMZ18 with a SHR PLAN APO 0.5X WD:71. Excitation light was produced by a fluorescent CoolLED pE-300 ultra at 580 nm. Images were collected with a Mono Camera Nikon DS-Fi3 camera. Exposure time was set to 1 sec with a resolution of 8 bit 1440 × 1024. Images were acquired every 5 sec (See also Video S1). (G,H) Confocal laser scanning micrographs showing the increased [Ca2+]cyt in Lima bean (Phaseolus lunatus) leaves upon feeding by S. littoralis. The red fluorescence is associated to the chlorophyll present in the chloroplasts, whereas the green fluorescence indicates the cytosolic localization of Ca2+ by the indicator Calcium OrangeTM. Scale bars: G = 100 µm; H = 200 µm (Figures (G,H) by Massimo Maffei).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Parmagnani, A.S.; Maffei, M.E. Calcium Signaling in Plant-Insect Interactions. Plants 2022, 11, 2689. https://doi.org/10.3390/plants11202689
AMA Style
Parmagnani AS, Maffei ME. Calcium Signaling in Plant-Insect Interactions. Plants. 2022; 11(20):2689. https://doi.org/10.3390/plants11202689
Chicago/Turabian StyleParmagnani, Ambra S., and Massimo E. Maffei. 2022. "Calcium Signaling in Plant-Insect Interactions" Plants 11, no. 20: 2689. https://doi.org/10.3390/plants11202689
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.