

Chromosome Numbers and Genome Sizes of All 36 Duckweed Species (Lemnaceae)

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
3. Results and Discussion
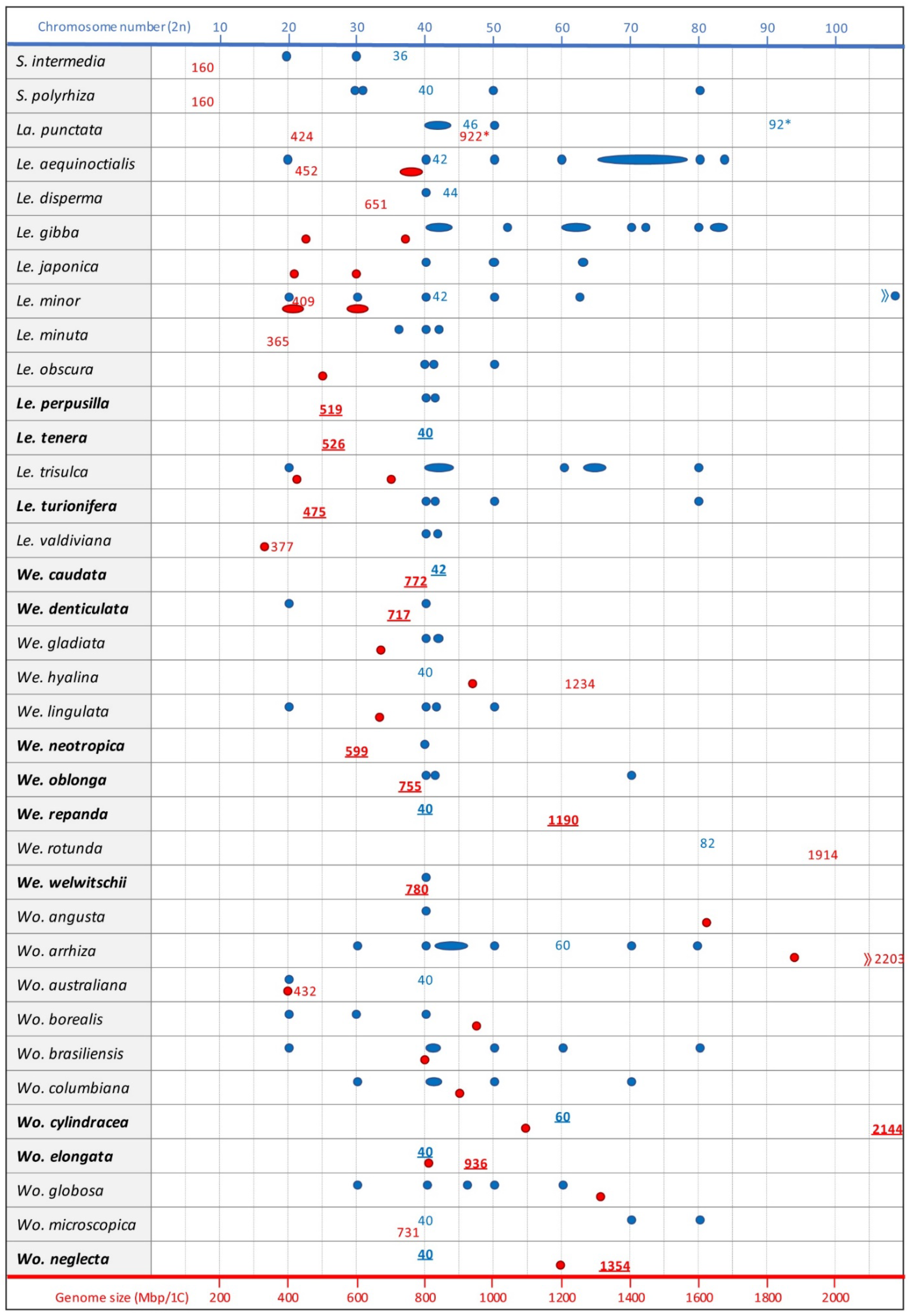
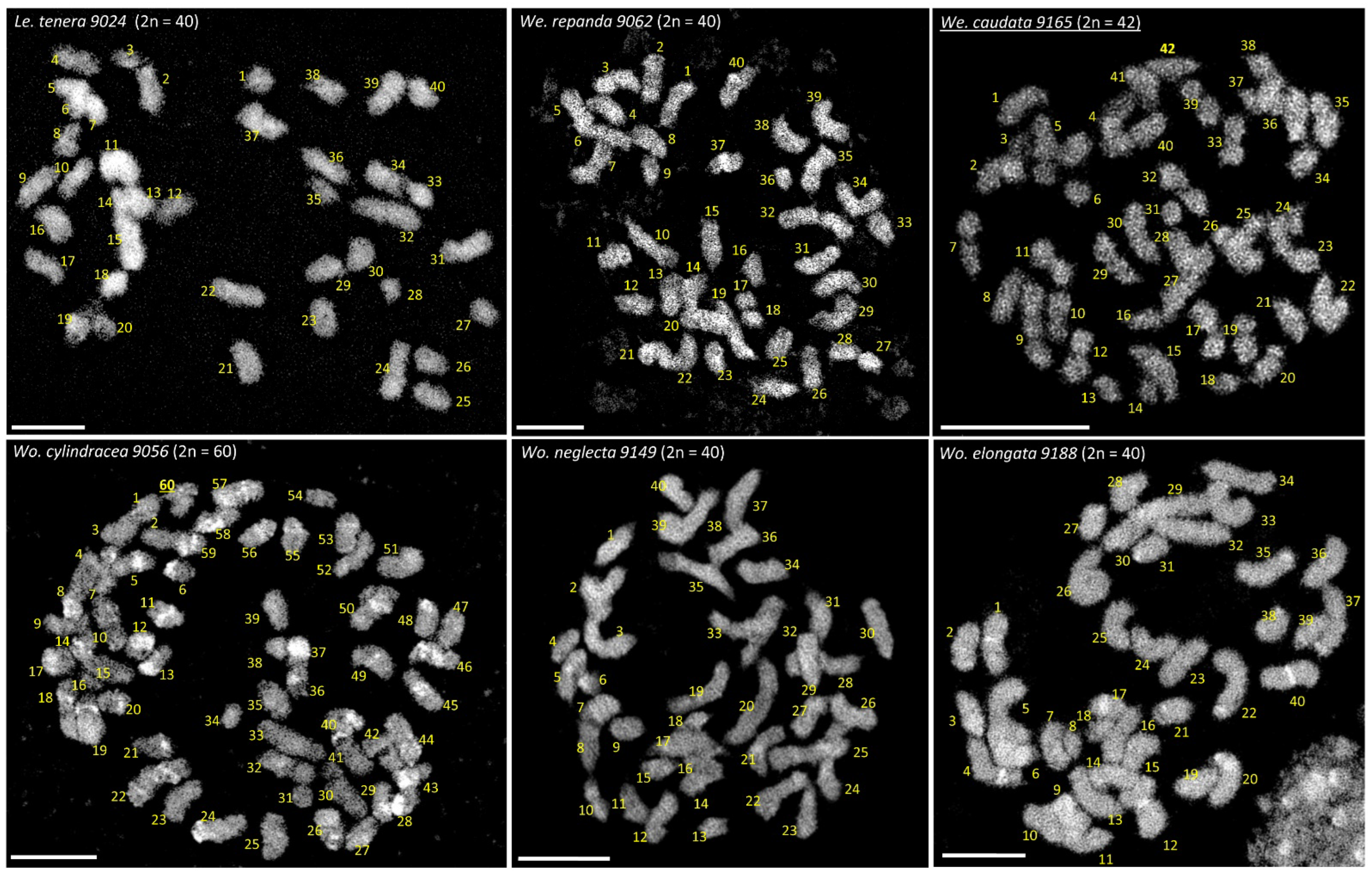
3.1. The Chromosomes
3.2. The Genome Sizes
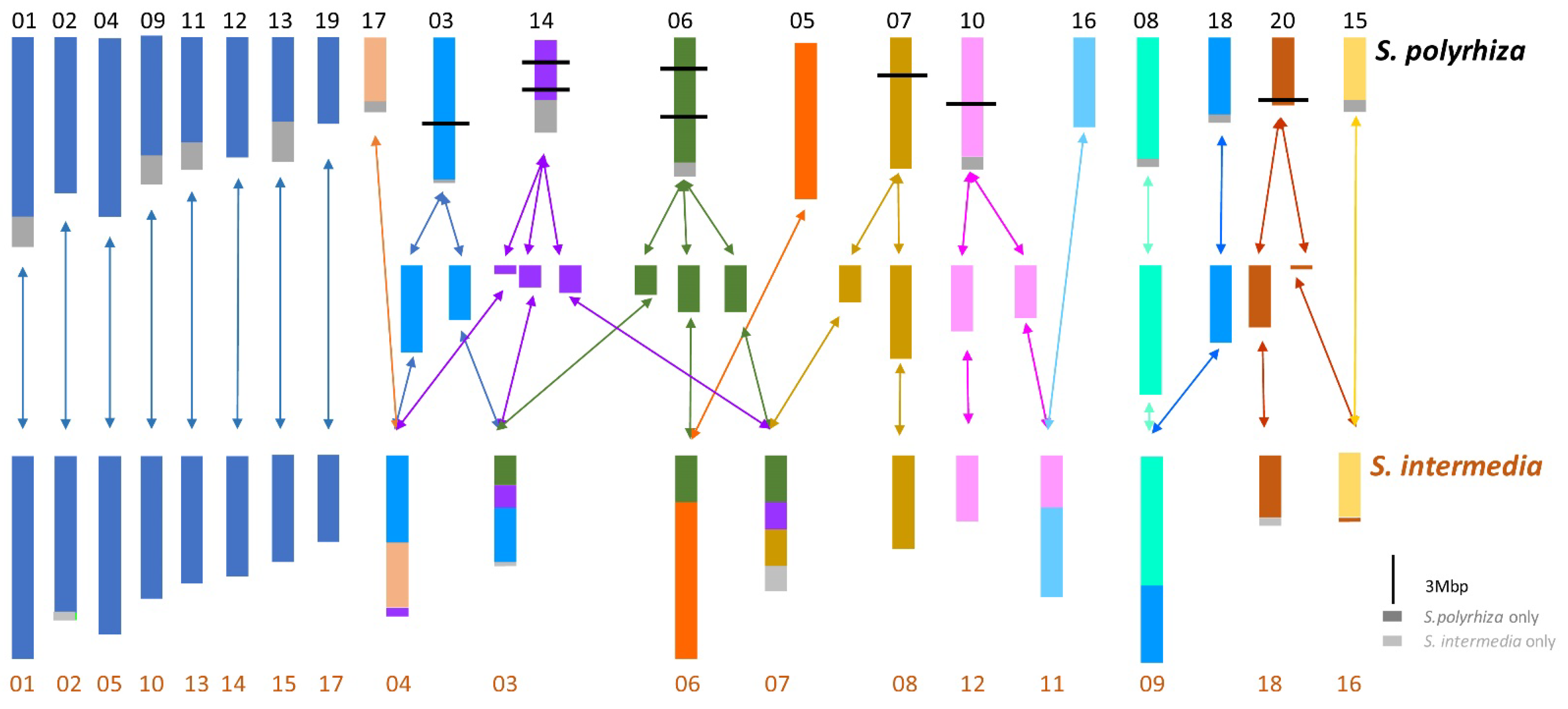
3.3. Evolutionary Impact of Genome Size and Karyotype Studies
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Acosta, K.; Appenroth, K.-J.; Borisjuk, L.; Edelman, M.; Heinig, U.; Jansen, M.; Oyama, T.; Pasaribu, B.; Schubert, I.; Sorrels, S.; et al. Return of the Lemnaceae: Duckweed as a model plant system in the genomics and post-genomics era. Plant Cell 2021, 33, 3207–3234. [Google Scholar] [CrossRef] [PubMed]
- Hoang, P.T.N.; Schubert, V.; Meister, A.; Fuchs, J.; Schubert, I. Variation in genome size, cell and nucleus volume, chromosome number and rDNA loci among duckweeds. Sci. Rep. 2019, 9, 3234. [Google Scholar] [CrossRef] [PubMed]
- Bog, M.; Sree, K.S.; Xu, S.; Himmelbach, A.; Brandt, R.; Fuchs, J.; Hoang, P.T.N.; Schubert, I.; Kuever, J.; Rabenstein, A.; et al. A taxonomic revision of the section Uninerves, genus Lemna. Taxon 2020, 69, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Kerstetter, R.A.; Michael, T.P. Evolution of genome size in duckweeds (Lemnaceae). J. Bot. 2011, 2011, 570319. [Google Scholar] [CrossRef] [Green Version]
- Urbanska, W.K. Cytological variation within the family of Lemnaceae. In Veröffentlichungen des Geobotanischen Institutes der Eidg. Tech. Hochschule, Stiftung Rübel, in Zürich; ETH Library: Zürich, Switzerland, 1980. [Google Scholar] [CrossRef]
- Geber, G. Zur Karyosystematik der Lemnaceae. Ph.D. Thesis, University of Vienna, Vienna, Austria, 1989. [Google Scholar]
- Beppu, T.; Takimoto, A. Geographical distribution and cytological variation of Lemna paucicostata Hegelm. in Japan. Bot. Mag. 1981, 94, 11–20. [Google Scholar] [CrossRef]
- Vunsh, R.; Heinig, U.; Malitsky, S.; Aharoni, A.; Lerner, A.; Edelman, M. Manipulating duckweed through genome duplication. Plant Biol. 2014, 17, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Dolezel, J.; Greilhuber, J.; Suda, J. Estimation of nuclear DNA content in plants using flow cytometry. Nat. Protoc. 2007, 2, 2233–2244. [Google Scholar] [CrossRef]
- Sun, H.; Ding, J.; Piednoel, M.; Schneeberger, K. findGSE: Estimating genome size variation within human and Arabidopsis using k-mer frequencies. Bioinformatics 2018, 34, 550–557. [Google Scholar] [CrossRef] [Green Version]
- Pflug, J.M.; Holmes, V.R.; Burrus, C.; Johnston, J.S.; Maddison, D.R. Measuring genome sizes using read-depth, k-mers, and flow cytometry: Methodological comparisons in beetles (Coleoptera). G3 2020, 10, 3047–3060. [Google Scholar] [CrossRef]
- Bog, M.; Lautenschläger, U.; Landrock, M.F.; Landolt, E.; Fuchs, J.; Sree, K.S.; Oberprieler, C.; Appenroth, K.J. Genetic characterization and barcoding of taxa in the genera Landoltia and Spirodela (Lemnaceae) by three plastidic markers and amplified fragment length polymorphism (AFLP). Hydrobiologia 2015, 749, 169–182. [Google Scholar] [CrossRef]
- Michael, T.P.; Bryant, D.; Gutierrez, R.; Borisjuk, N.; Chu, P.; Zhang, H.; Xia, J.; Zhou, J.; Peng, H.; El Baidouri, M.; et al. Comprehensive definition of genome features in Spirodela polyrhiza by high-depth physical mapping and short-read DNA sequencing strategies. Plant J. 2017, 89, 617–635. [Google Scholar] [CrossRef] [Green Version]
- Michael, T.P.; Ernst, E.; Hartwick, N.; Chu, P.; Bryant, D.; Gilbert, S.; Ortleb, S.; Baggs, E.L.; Sree, S.K.; Appenroth, K.J.; et al. Genome and time-of-day transcriptome of Wolffia australiana link morphological extreme minimization with gene loss and less growth control. Genome Res. 2021, 31, 225–238. [Google Scholar] [CrossRef]
- Schubert, I.; Vu, G.T.H. Genome stability and evolution: Attempting a holistic view. Trends Plant Sci. 2016, 21, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Van Hoeck, A.; Horemans, N.; Monsieurs, P.; Cao, H.X.; Vandenhove, H.; Blust, R. The first draft genome of the aquatic model plant Lemna minor opens the route for future stress physiology research and biotechnological applications. Biotechnol. Biofuels 2015, 8, 188. [Google Scholar] [CrossRef] [Green Version]
- Braglia, L.; Lauria, M.; Appenroth, K.-J.; Bog, M.; Breviario, D.; Grasso, A.; Gavazzi, F.; Morello, L.; Giani, S. Duckweed species genotyping and interspecific hybrid discovery by tubulin-based polymorphism fingerprinting. Front. Plant Sci. 2021, 12, 625670. [Google Scholar] [CrossRef]
- Mandakova, T.; Joly, S.; Krzywinski, M.; Mummenhoff, K.; Lysak, M.A. Fast diploidization in close mesopolyploid relatives of Arabidopsis. Plant Cell 2010, 22, 2277–2290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, H.X.; Vu, G.T.H.; Wang, W.; Appenroth, K.J.; Messing, J.; Schubert, I. The map-based genome sequence of Spirodela polyrhiza aligned with its chromosomes, a reference for karyotype evolution. New Phytol. 2016, 209, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Hoang, P.N.T.; Michael, T.P.; Gilbert, S.; Chu, P.; Motley, T.S.; Appenroth, K.J.; Schubert, I.; Lam, E. Generating a high-confidence reference genome map of the Greater Duckweed by integration of cytogenomic, optical mapping and Oxford Nanopore technologies. Plant J. 2018, 96, 670–684. [Google Scholar] [CrossRef] [Green Version]
- Hoang, P.T.N.; Fiebig, A.; Novak, P.; Macas, J.; Cao, H.X.; Stepaneko, A.; Chen, G.; Borisjuk, N.; Scholz, U.; Schubert, I. Chromosome-scale genome assembly for the duckweed Spirodela intermedia, integrating cytogenetic maps, PacBio and Oxford Nanopore libraries. Sci. Rep. 2020, 10, 19230. [Google Scholar] [CrossRef]
- Hoang, P.T.N.; Schubert, I. Reconstruction of chromosome rearrangements between the two most ancestral duckweed species Spirodela polyrhiza and S. intermedia. Chromosoma 2017, 126, 729–739. [Google Scholar] [CrossRef]
- Lysak, M.A.; Berr, A.; Pecinka, A.; Schmidt, R.; McBreen, K.; Schubert, I. Mechanisms of chromosome number reduction in Arabidopsis thaliana and related Brassicaceae species. Proc. Natl. Acad. Sci. USA 2006, 103, 5224–5229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, P.T.N. Comparative Cytology and Cytogenomics for Representative Species of the Five Duckweed Genera. Ph.D. Thesis, Martin-Luther-Universität Halle-Wittenberg, Halle, Germany, 2019. [Google Scholar]
- Hoang, P.T.N.; Macas, J.; Rouillard, J.-M.; Kubalová, I.; Schubert, V.; Schubert, I. Limitation of current probe design for oligo-cross FISH, exemplified by chromosome evolution studies in duckweeds. Chromosoma 2021, 130, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Weisshart, K.; Fuchs, J.; Schubert, V. Structured illumination microscopy(SIM) and photoactivated localization microscopy (PALM) to analyze the abundance and distribution of RNA polymerase II molecules on flow-sorted Arabidopsis nuclei. Bio-Protocol 2016, 6, e1725. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoang, P.T.N.; Fuchs, J.; Schubert, V.; Tran, T.B.N.; Schubert, I. Chromosome Numbers and Genome Sizes of All 36 Duckweed Species (Lemnaceae). Plants 2022, 11, 2674. https://doi.org/10.3390/plants11202674
Hoang PTN, Fuchs J, Schubert V, Tran TBN, Schubert I. Chromosome Numbers and Genome Sizes of All 36 Duckweed Species (Lemnaceae). Plants. 2022; 11(20):2674. https://doi.org/10.3390/plants11202674
Chicago/Turabian StyleHoang, Phuong T. N., Jörg Fuchs, Veit Schubert, Tram B. N. Tran, and Ingo Schubert. 2022. "Chromosome Numbers and Genome Sizes of All 36 Duckweed Species (Lemnaceae)" Plants 11, no. 20: 2674. https://doi.org/10.3390/plants11202674