
Development of Hairy Root Cultures for Biomass and Triterpenoid Production in Centella asiatica


Abstract
:1. Introduction
2. Results and Discussion
2.1. Induction and Characterization of HRs
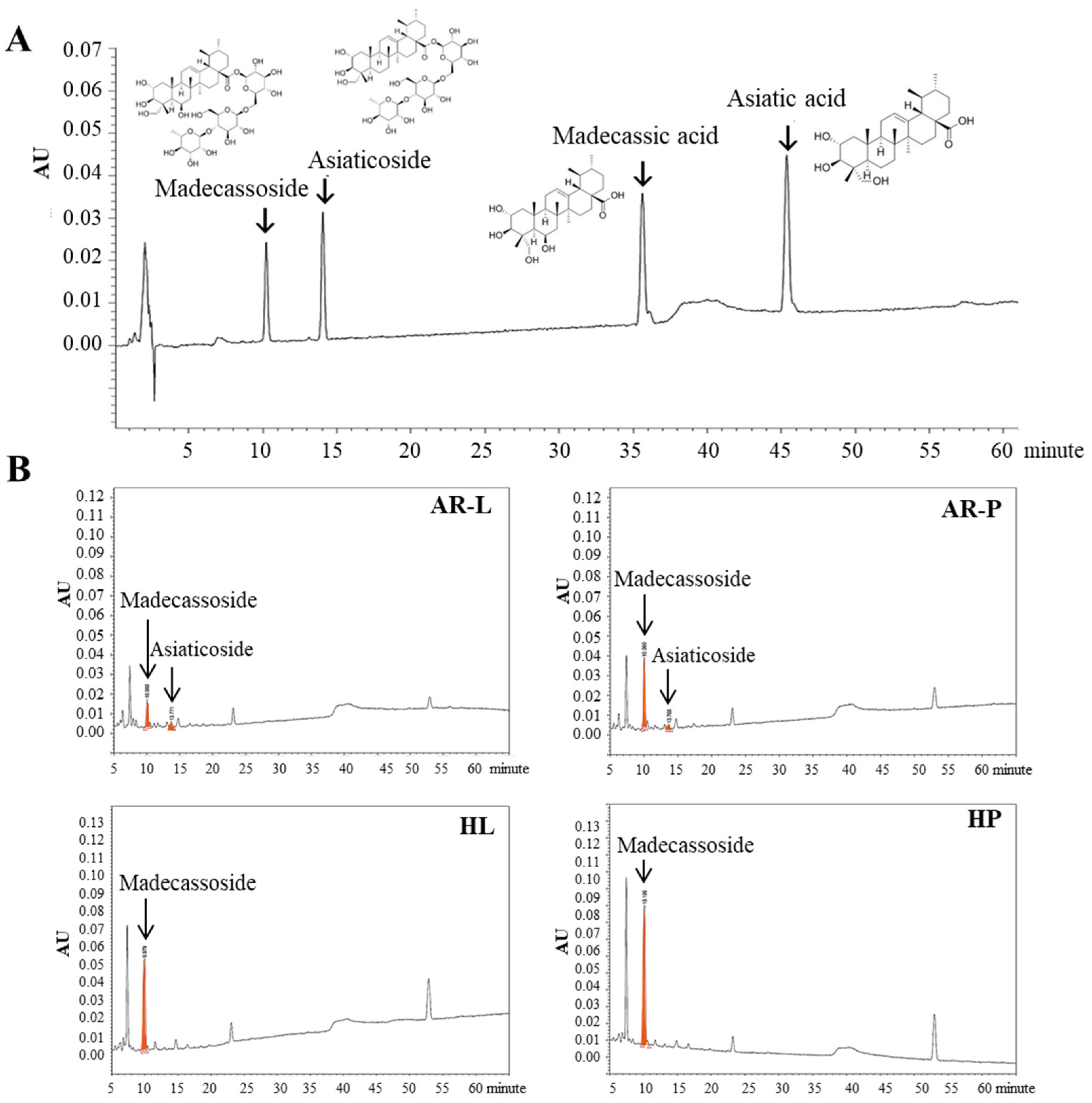
2.2. Confirmation of HR Transformation
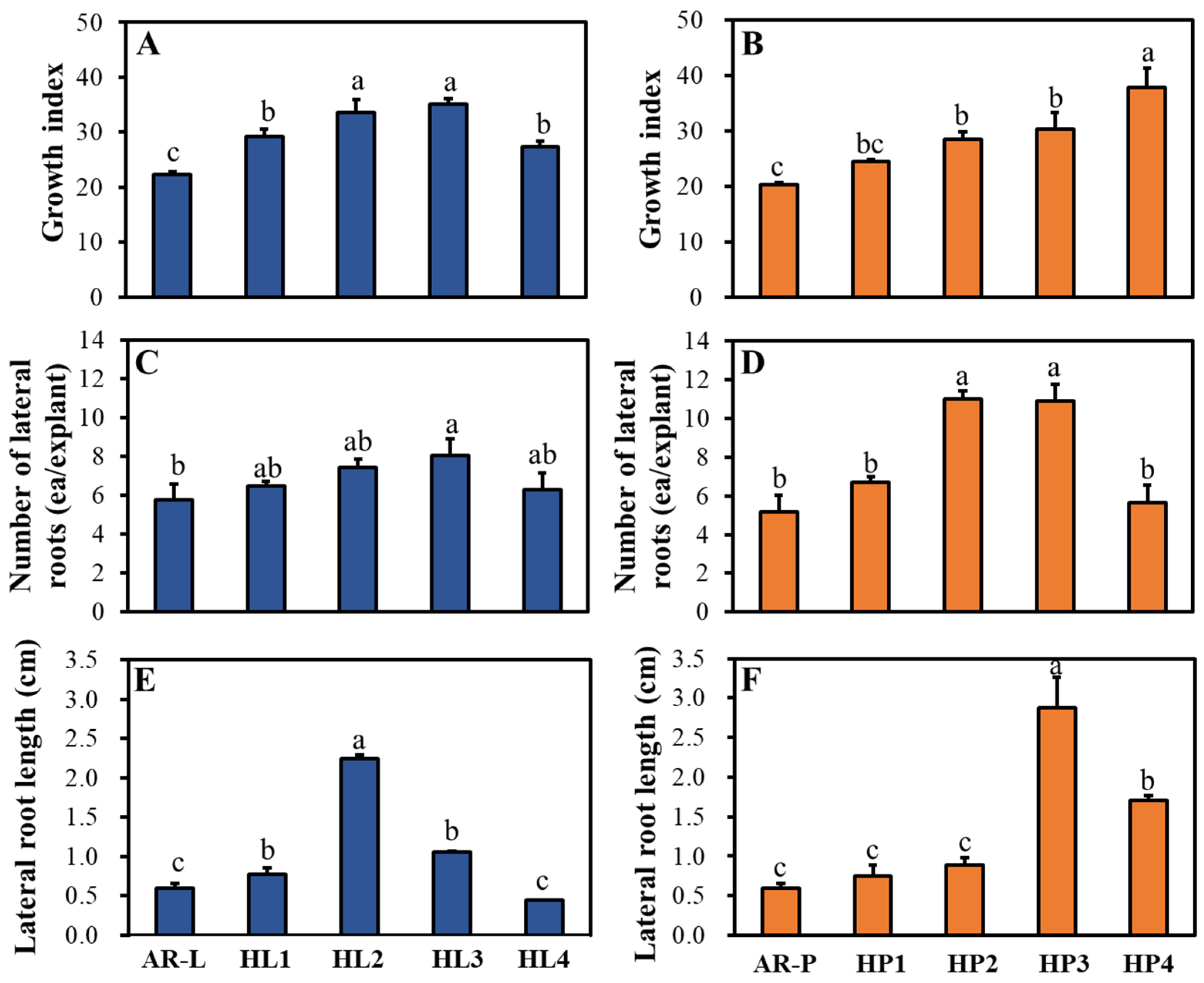
2.3. Growth and Proliferation of HR Lines
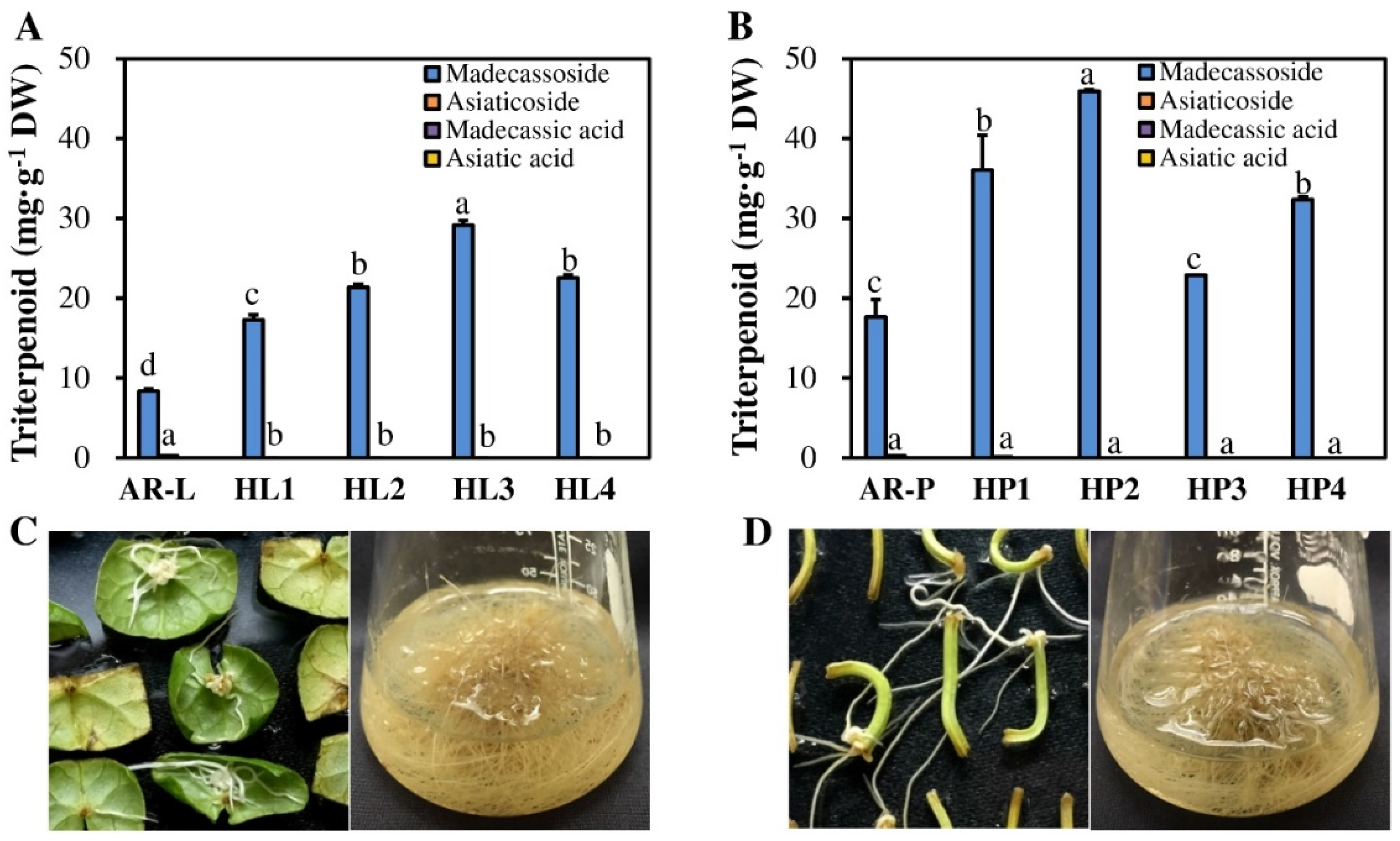
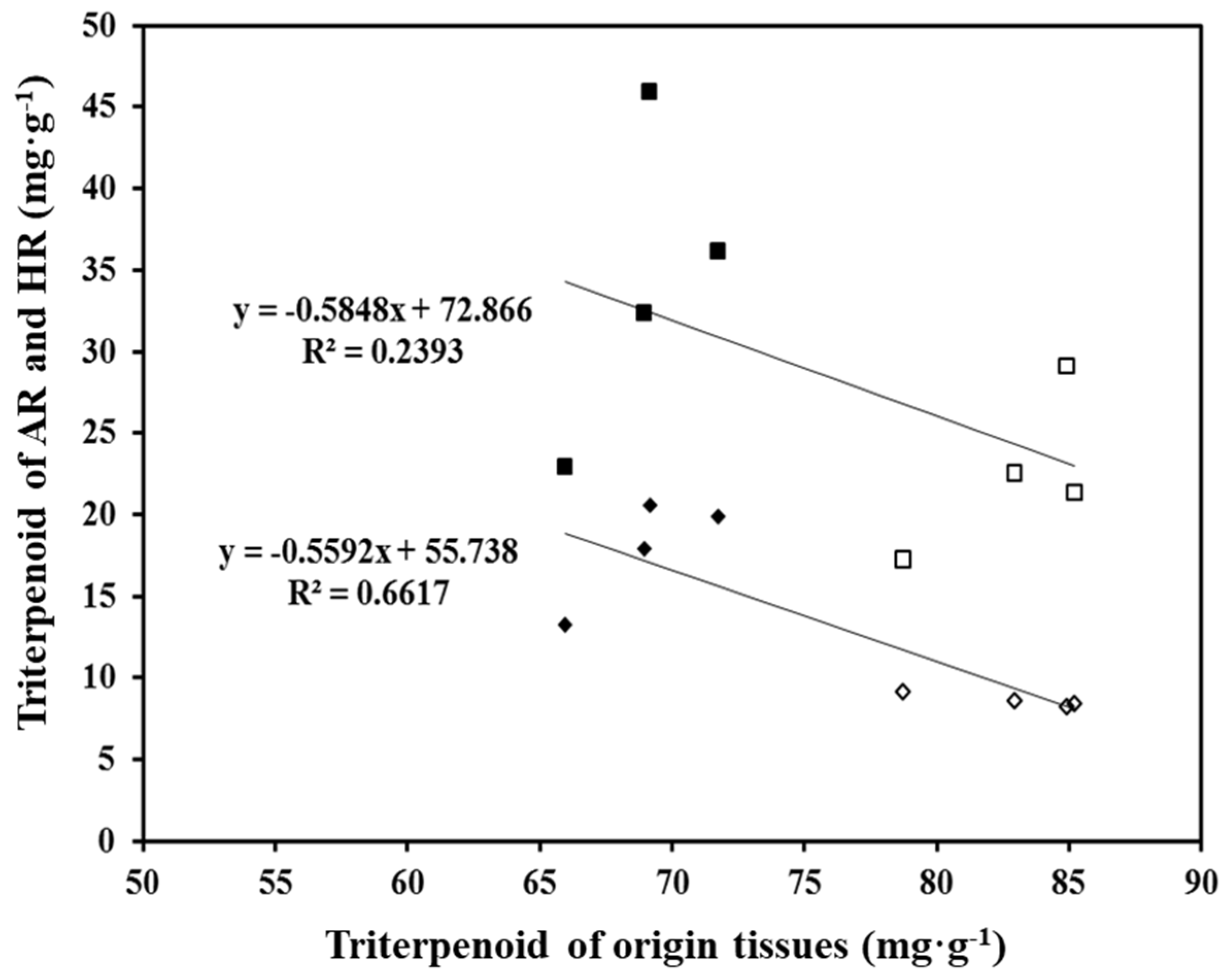
2.4. Comparison of Triterpenoid Accumulation between HR Lines and AR Cultures
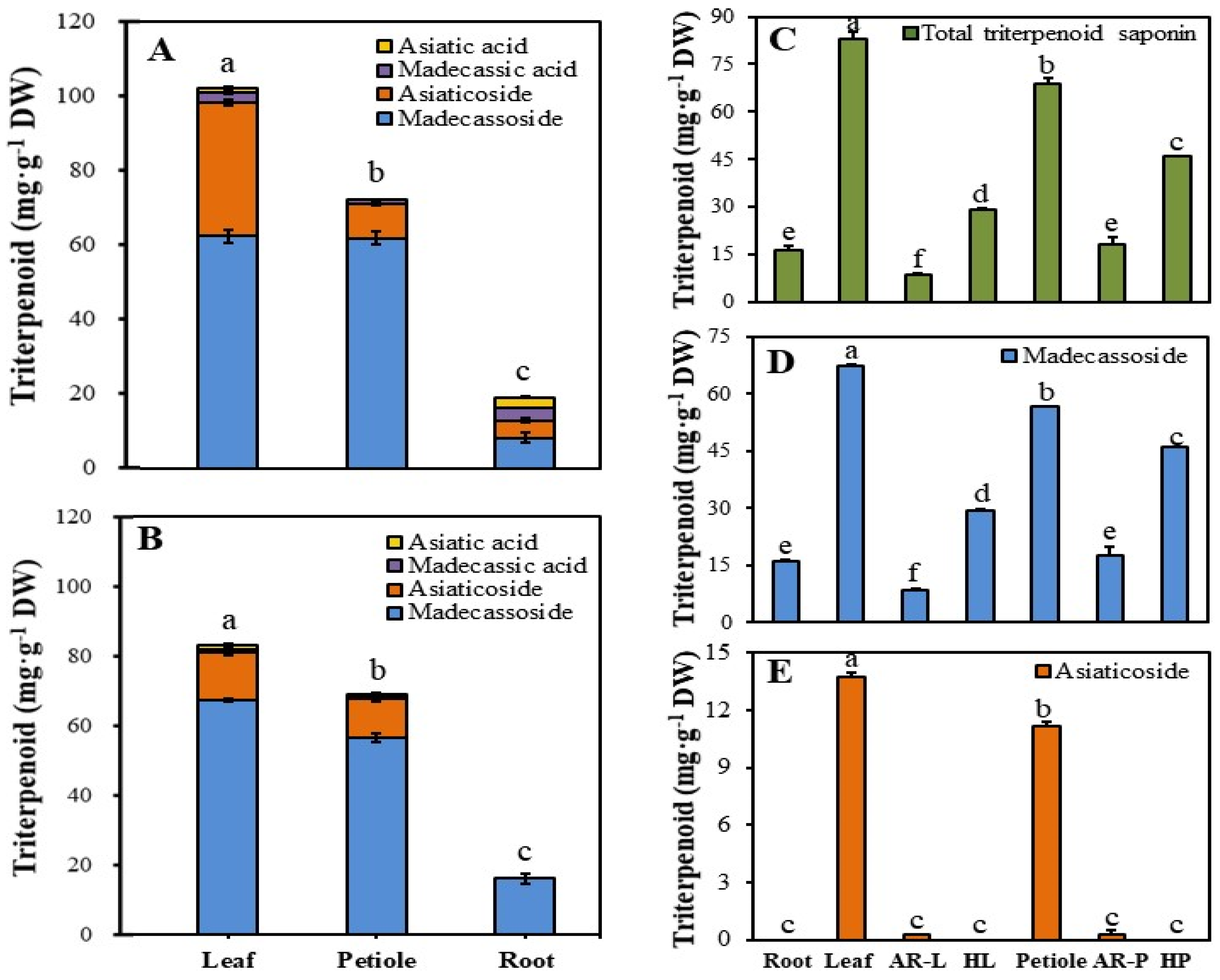
2.5. Comparison of Terpenoid Accumulation among HR Lines, AR Cultures, and Source Explants
3. Materials and Methods
3.1. Plant Materials
3.2. HR Induction
3.3. Genomic DNA Extraction and PCR Analysis
3.4. Characterization of HR Lines
3.5. Analysis of Triterpenoids by HPLC
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singh, G.; Kaur, B.; Sharma, N.; Bano, A.; Kumar, S.; Dhaliwal, H.; Sharma, V. In vitro micropropagation and cytomorphological evaluation of Centella asiatica (L.) Urban (Mandukparni) from Himachal Pradesh, India—An Endemic, Endangered and Threatened Herb. Plant Tissue Cult. Biotechnol. 2014, 24, 155–171. [Google Scholar] [CrossRef] [Green Version]
- Jamil, S.S.; Nizami, Q.; Salam, M. Centella asiatica (Linn.) Urban: A review. Nat. Prod. Radiance 2007, 6, 158–170. [Google Scholar]
- Muller, V.; Lankes, C.; Zimmermann, B.; Noga, G.; Hunsche, M. Centelloside accumulation in leaves of Centella asiatica is determined by resource partitioning between primary and secondary metabolism while influenced by supply levels of either nitrogen, phosphorus or potassium. J. Plant Physiol. 2013, 170, 1165–1175. [Google Scholar] [CrossRef] [PubMed]
- Maquart, F.X.; Chastang, F.; Simeon, A.; Birembaut, P.; Gillery, P.; Wegrowski, Y. Triterpenes from Centella asiatica stimulate extracellular matrix accumulation in rat experimental wounds. Eur. J. Dermatol. 1999, 9, 289–296. [Google Scholar]
- Li, H.Z.; Wan, J.Y.; Zhang, L.; Zhou, Q.X.; Luo, F.L.; Zhang, Z. Inhibitiory action of asiaiticoside on collagen-induced arthritis in mice. Yao Xue Xue Bao 2007, 42, 698–703. [Google Scholar]
- Vasantharuba, S.V.; Banumathi, P.; Premalatha, M.R.; Sundaram, S.P.; Arumugam, T. Functional properties of Centella asiatica (L.): A review. Int. J. Pharm. Pharm. Sci. 2012, 4, 8–14. [Google Scholar]
- James, J.T.; Dubery, I.A. Pentacyclic triterpenoids from the medicinal herb, Centella asiatica (L.) Urban. Molecules 2009, 14, 3922–3941. [Google Scholar] [CrossRef] [Green Version]
- Devkota, A.; Dall’Acqua, S.; Comai, S.; Innocenti, G.; Jha, P.K. Centella asiatica (L.) urban from Nepal: Quali-quantitative analysis of samples from several sites, and selection of high terpene containing populations for cultivation. Biochem. Syst. Ecol. 2010, 38, 12–22. [Google Scholar] [CrossRef]
- Singh, S.; Gautam, A.; Sharma, A.; Batra, A. Centella asiatica (L.): A plant with immense medicinal potential but threatened. Int. J. Pharm. Sci. Rev. Res. 2010, 4, 9–17. [Google Scholar]
- Kim, O.T.; Kim, M.Y.; Hong, M.H.; Ahn, J.C.; Hwang, B. Stimulation of asiaticoside accumulation in the whole plant cultures of Centella asiatica (L.) Urban by elicitors. Plant Cell Rep. 2004, 23, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.T.; Bang, K.H.; Shin, Y.S.; Lee, M.J.; Jung, S.J.; Hyun, D.Y.; Kim, Y.C.; Seong, N.S.; Cha, S.W.; Hwang, B. Enhanced production of asiaticoside from hairy root cultures of Centella asiatica (L.) Urban elicited by methyl jasmonate. Plant Cell Rep. 2007, 26, 1941–1949. [Google Scholar] [CrossRef]
- Ho, T.T.; Lee, J.D.; Ahn, M.S.; Kim, S.W.; Park, S.Y. Enhanced production of phenolic compounds in hairy root cultures of Polygonum multiflorum and its metabolite discrimination using HPLC and FT-IR methods. Appl. Microbiol. Biotechnol. 2018, 102, 9563–9575. [Google Scholar] [CrossRef]
- Shi, H.P.; Kintzios, S. Genetic transformation of Pueraria phaseoloides with Agrobacterium rhizogenes and puerarin production in hairy roots. Plant Cell Rep. 2003, 21, 1103–1107. [Google Scholar] [CrossRef]
- Ono, N.N.; Tain, L. The multiplicity of hairy root cultures: Prolific possibilities. Plant Sci. 2011, 180, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Shanks, J.V.; Morgan, J. Plant ‘hairy root’culture. Curr. Opin. Biotechnol. 1999, 10, 151–155. [Google Scholar] [CrossRef]
- Sevón, N.; Oksman-Caldentey, K.M. Agrobacterium rhizogenes-mediated transformation: Root cultures as a source of alkaloids. Planta Med. 2002, 68, 859–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, Z.A.; Davey, M.R.; Power, J.B.; Anthony, P.; Smith, R.M.; Lowe, K.C. Production of asiaticoside and madecassoside in Centella asiatica in vitro and in vivo. Biol. Plant. 2007, 51, 34–42. [Google Scholar] [CrossRef]
- Nguyen, K.V.; Pongkitwitoon, B.; Pathomwichaiwat, T.; Viboonjun, U.; Prathanturarug, S. Effects of methyl jasmonate on the growth and triterpenoid production of diploid and tetraploid Centella asiatica (L.) Urb. hairy root cultures. Sci. Rep. 2019, 9, 18665. [Google Scholar] [CrossRef]
- Karthikeyan, K.; Chandran, C.; Kulothungan, S. Rapid clonal multiplication through in vitro axillary shoot proliferation of Centella asiatica L. Indian J. Biotechnol. 2009, 8, 232–235. [Google Scholar]
- Nester, E.W. Agrobacterium: Nature’s genetic engineer. Front. Plant Sci. 2014, 5, 730. [Google Scholar] [CrossRef] [Green Version]
- Alpizar, E.; Dechamp, E.; Espeout, S.; Royer, M.; Lecouls, A.C.; Nicole, M.; Bertrand, B.; Lashermes, P.; Etienne, H. Efficient production of Agrobacterium rhizogenes-transformed roots and composite plants for studying gene expression in coffee roots. Plant Cell Rep. 2006, 25, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, K.; Haghbeen, K.; Marefatjo, J.; Jazii, F.R.; Sheikhani, R. Successful production of hairy root of Valeriana sisymbriifolium by Agrobacterium rhizogenes. Biotechnology 2008, 7, 200–204. [Google Scholar] [CrossRef]
- Das, S.; Jha, T.B.; Jha, S. Organogenesis and regeneration from pigmented callus in Camellia sinensis (L.) O. Kuntze cv. Nandadevi, an elite Darjeeling tea clone. Plant Sci. 1996, 121, 207–212. [Google Scholar] [CrossRef]
- Vanegas, P.E.; Cruz-Hernández, A.; Valverde, M.E.; Paredes-López, O. Plant regeneration via organogenesis in marigold. Plant Cell Tissue Organ Cult. 2002, 69, 279–283. [Google Scholar] [CrossRef]
- Hosoki, T.; Asahira, T. In vitro propagation of bromeliads in liquid culture. Hortic. Sci. 1980, 15, 603–604. [Google Scholar]
- Chandra, S. Natural plant genetic engineer Agrobacterium rhizogenes: Role of T-DNA in plant secondary metabolism. Biotechnol. Lett. 2012, 34, 407–415. [Google Scholar] [CrossRef]
- Shkryl, Y.N.; Veremeichik, G.N.; Bulgakov, V.P.; Tchernoded, G.K.; Mischenko, N.P.; Fedoreyev, S.A.; Zhuralev, Y.N. Individual and combined effects of the rol A, B and C genes on anthraquinone production in Rubia cordifolia transformed calli. Biotechnol. Bioeng. 2008, 100, 118–125. [Google Scholar] [CrossRef]
- Ruslan, K.; Selfitri, A.D.; Bulan, S.A.; Rukayadi, Y. Effect of Agrobacterium rhizogenes and elicitation on the asiaticoside production in cell cultures of Centella asiatica. Pharmacogn. Mag. 2012, 8, 111. [Google Scholar]
- Kiselev, K.V.; Dubrovina, A.S.; Veselova, M.V.; Bulgakov, V.P.; Fedoreyev, S.A.; Zhuravlev, Y.N. The rolB gene-induced overproduction of resveratrol in Vitis amurensis transformed cells. J. Biotechnol. 2007, 128, 681–692. [Google Scholar] [CrossRef]
- Palazón, J.; Cusidó, R.; Roig, C.; Piñol, M.T. Effect of rol genes from Agrobacterium rhizogenes TL-DNA on nicotine production in tobacco root cultures. Plant Physiol. Biochem. 1997, 35, 155–162. [Google Scholar]
- Bonhomme, V.; Laurain-Mattar, D.; Fliniaux, M.A. Effects of the rolC gene on hairy root: Induction development and tropane alkaloid production by Atropa belladonna. J. Nat. Prod. 2000, 63, 1249–1252. [Google Scholar] [CrossRef]
- McCoy, E.; O’Connor, S. Natural products from plant cell cultures. Prog. Drug Res. 2008, 65, 330–370. [Google Scholar]
- Kim, K.; Lee, S.C.; Lee, J.; Lee, H.O.; Joh, H.J.; Kim, N.H.; Yang, T.J. Comprehensive survey of genetic diversity in chloroplast genomes and 45S nrDNAs within Panax ginseng species. PLoS ONE 2015, 10, e0117159. [Google Scholar]
- Romani, A.; Vignolini, P.; Galardi, C.; Aroldi, C.; Vazzana, C.; Heimler, D. Polyphenolic content in different plant parts of soy cultivars grown under natural conditions. J. Agric. Food Chem. 2003, 51, 5301–5306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Rogers, S.O.; Bendich, A.J. Extraction of total cellular DNA from plants, algae and fungi. In Plant Molecular Biology Manual; Springer: Amsterdam, The Netherlands, 1988; pp. 183–190. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Explant | Treatment | No. of Explants | Survival Rate (%) | Frequency of Root Formation (%) | No. of Confirmed HR z Lines |
|---|---|---|---|---|---|
| Leaf | Control | 50 | 100 ± 0.0 a y | 4.0 ± 0.1 b | 0.0 ± 0.0 b |
| KCCM 11879 | 250 | 100 ± 0.0 a | 26.8 ± 2.7 a | 19.1 ± 0.7 a | |
| Petiole | Control | 90 | 100 ± 0.0 a | 0.0 ± 0.1 b | 0.0 ± 0.0 b |
| KCCM 11879 | 450 | 100 ± 0.0 a | 12.0 ± 1.6 a | 17.0 ± 1.9 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baek, S.; Han, J.-E.; Ho, T.-T.; Park, S.-Y. Development of Hairy Root Cultures for Biomass and Triterpenoid Production in Centella asiatica. Plants 2022, 11, 148. https://doi.org/10.3390/plants11020148
Baek S, Han J-E, Ho T-T, Park S-Y. Development of Hairy Root Cultures for Biomass and Triterpenoid Production in Centella asiatica. Plants. 2022; 11(2):148. https://doi.org/10.3390/plants11020148
Chicago/Turabian StyleBaek, Seungeun, Jong-Eun Han, Thanh-Tam Ho, and So-Young Park. 2022. "Development of Hairy Root Cultures for Biomass and Triterpenoid Production in Centella asiatica" Plants 11, no. 2: 148. https://doi.org/10.3390/plants11020148