
Heterologous Expression of Three Transcription Factors Differently Regulated Astragalosides Metabolic Biosynthesis in Astragalus membranaceus Hairy Roots


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
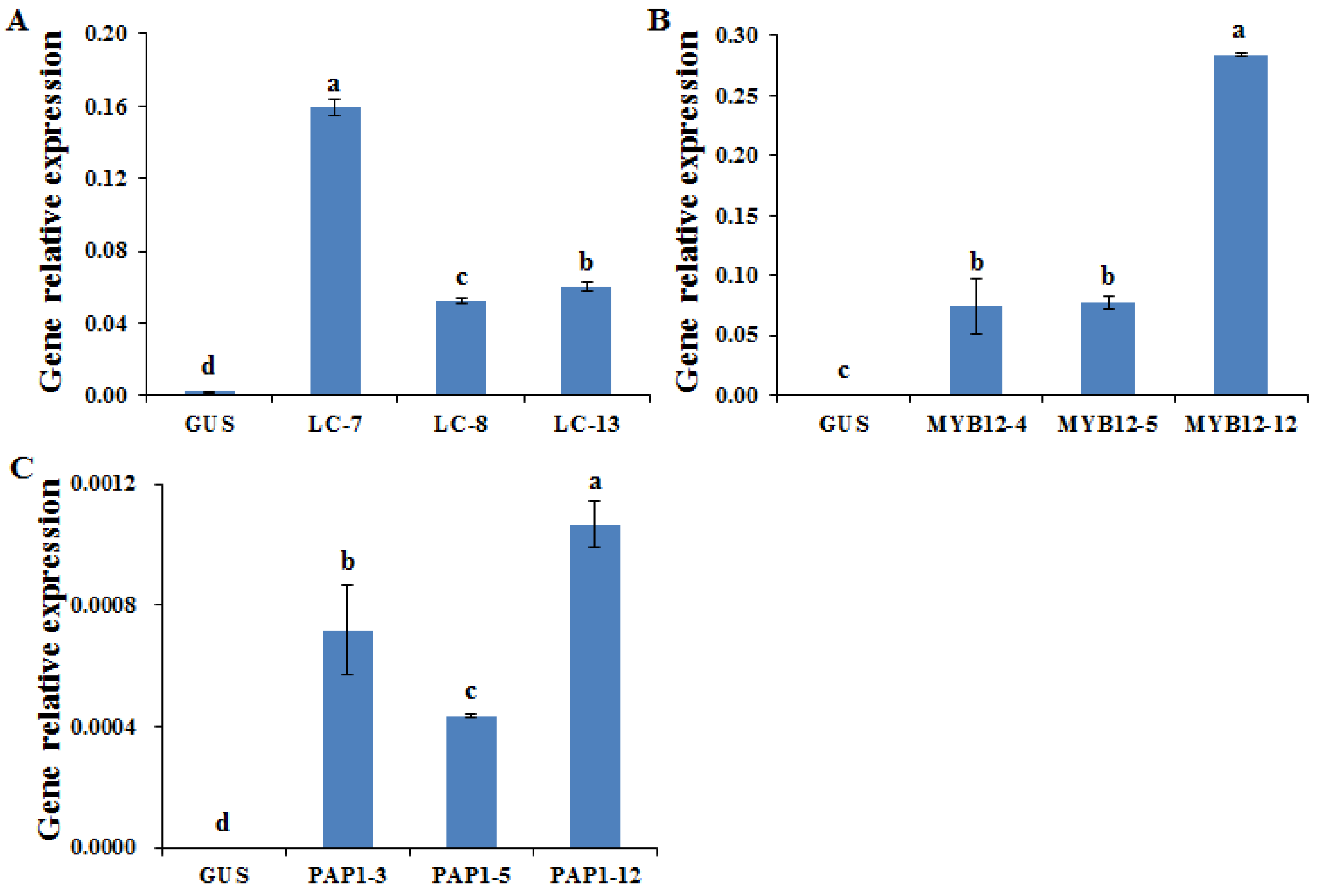
2.1. Identification of A. membranaceus Hairy Root Overexpression Lines
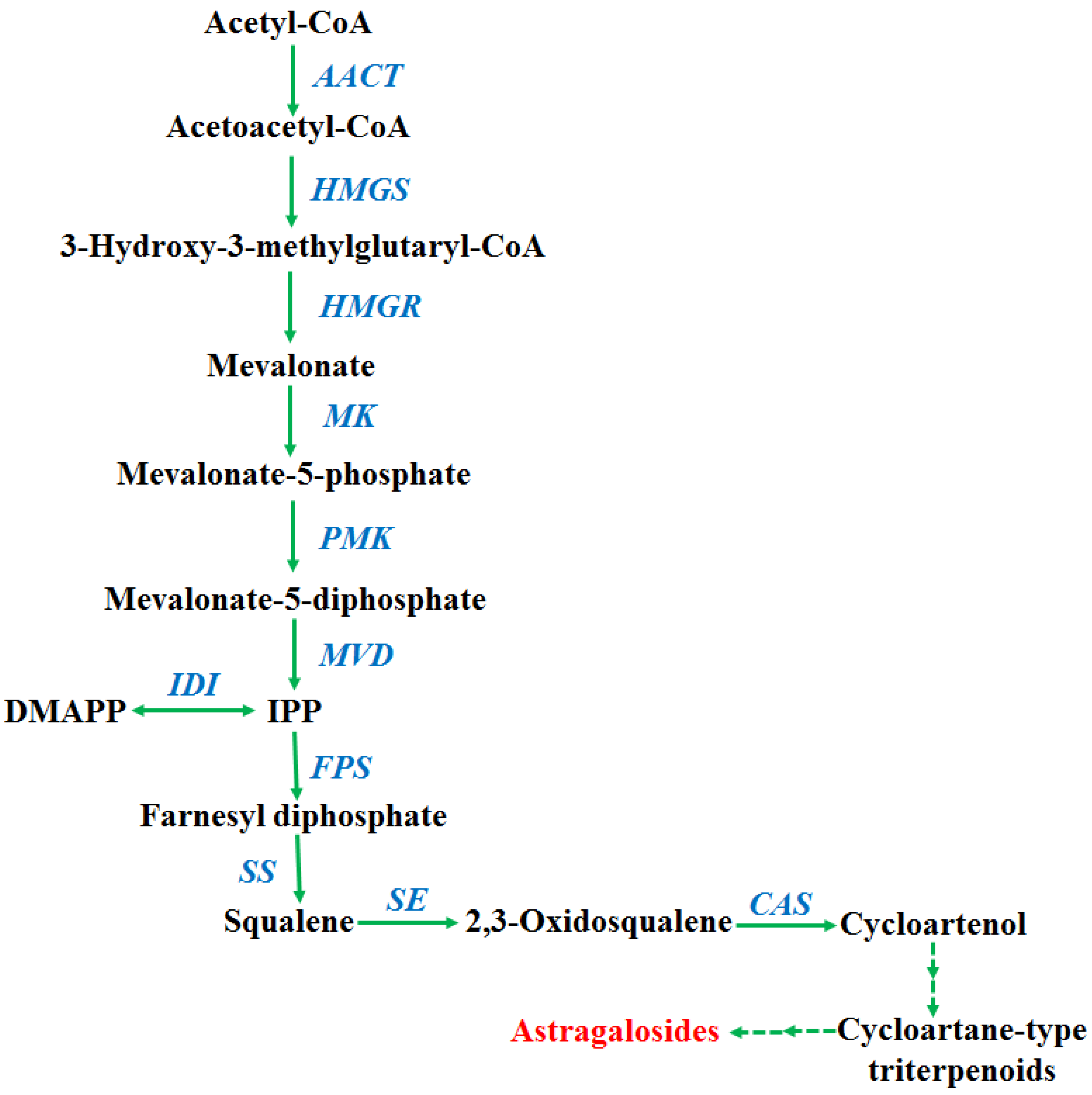
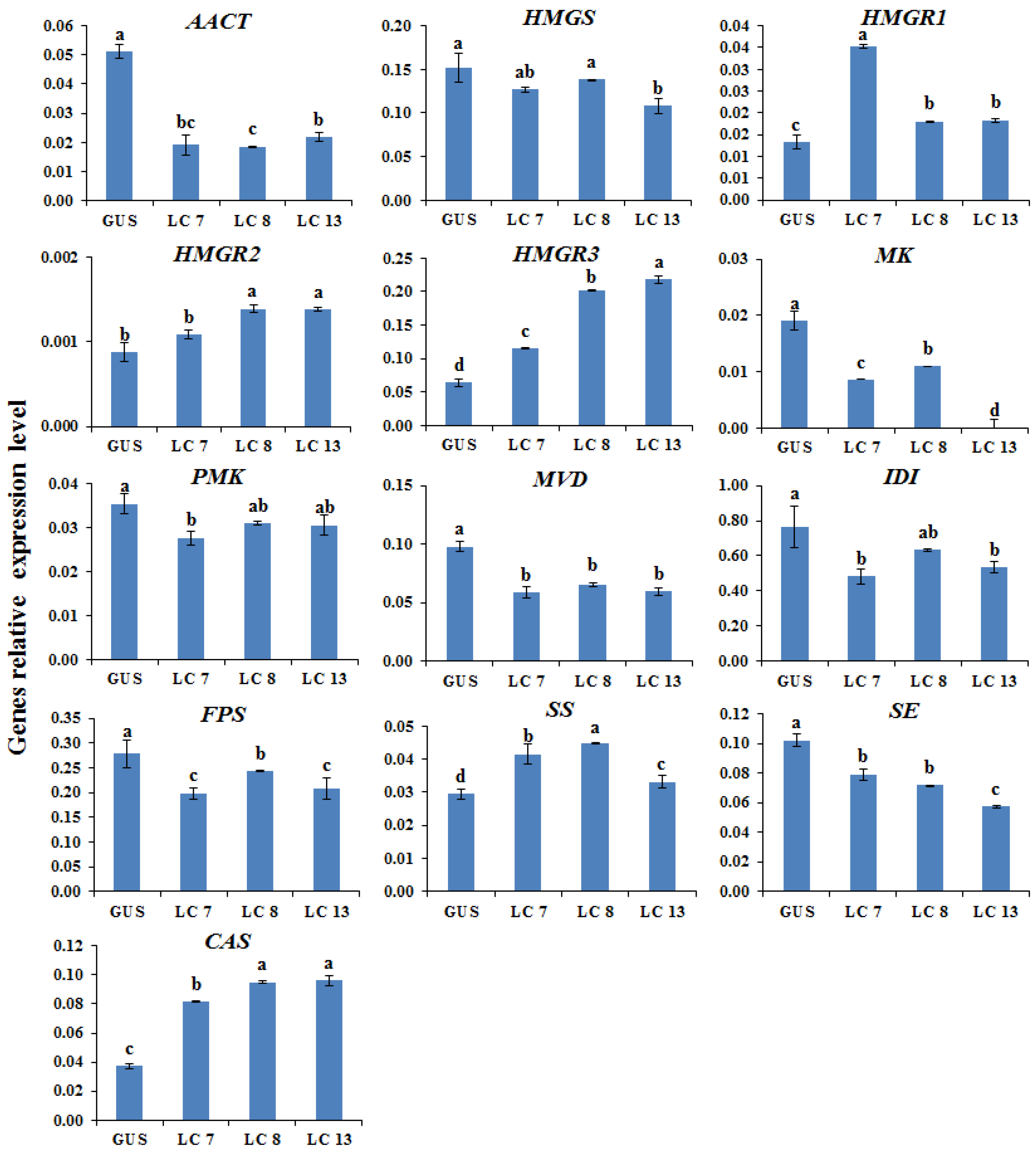
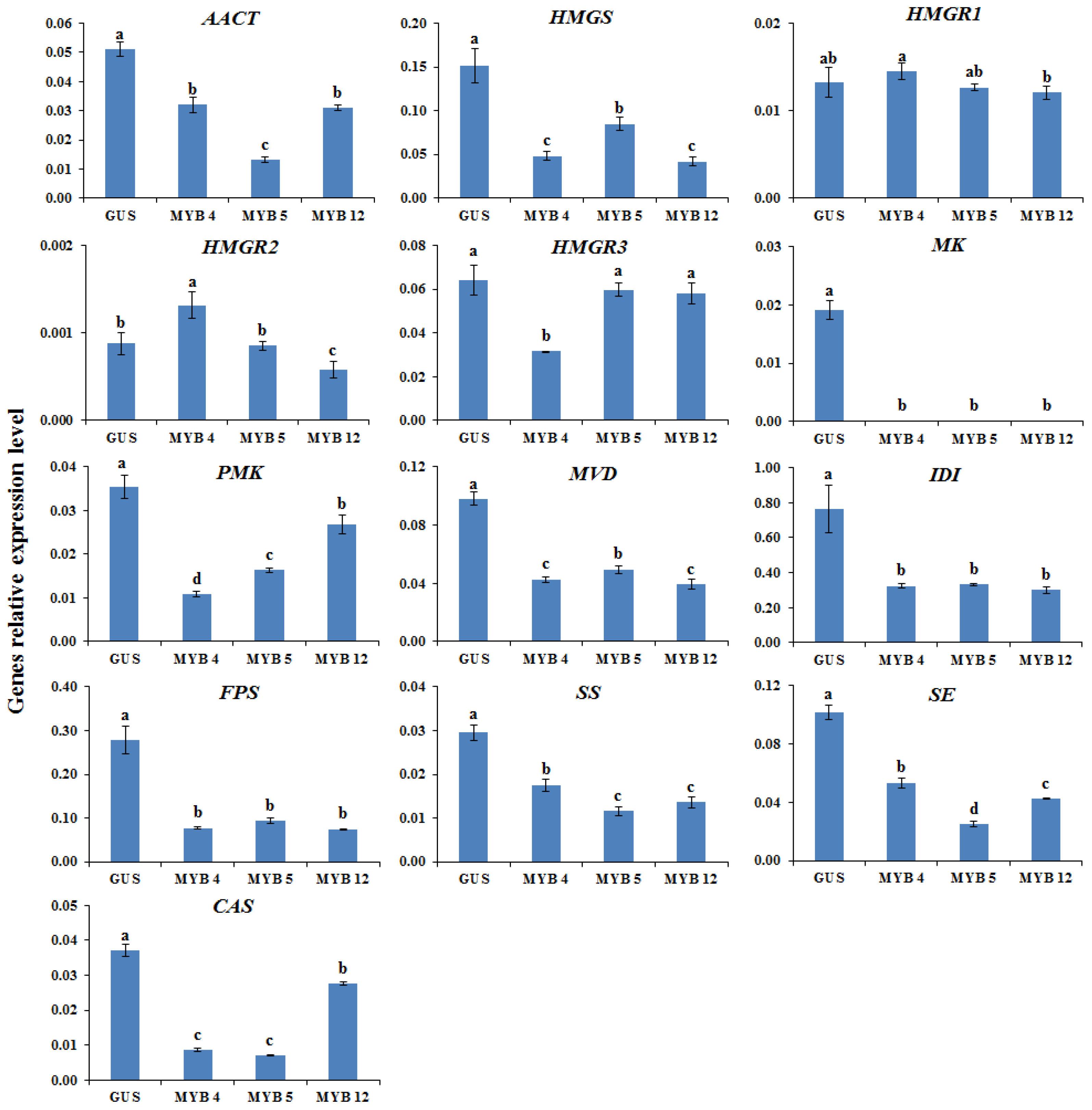
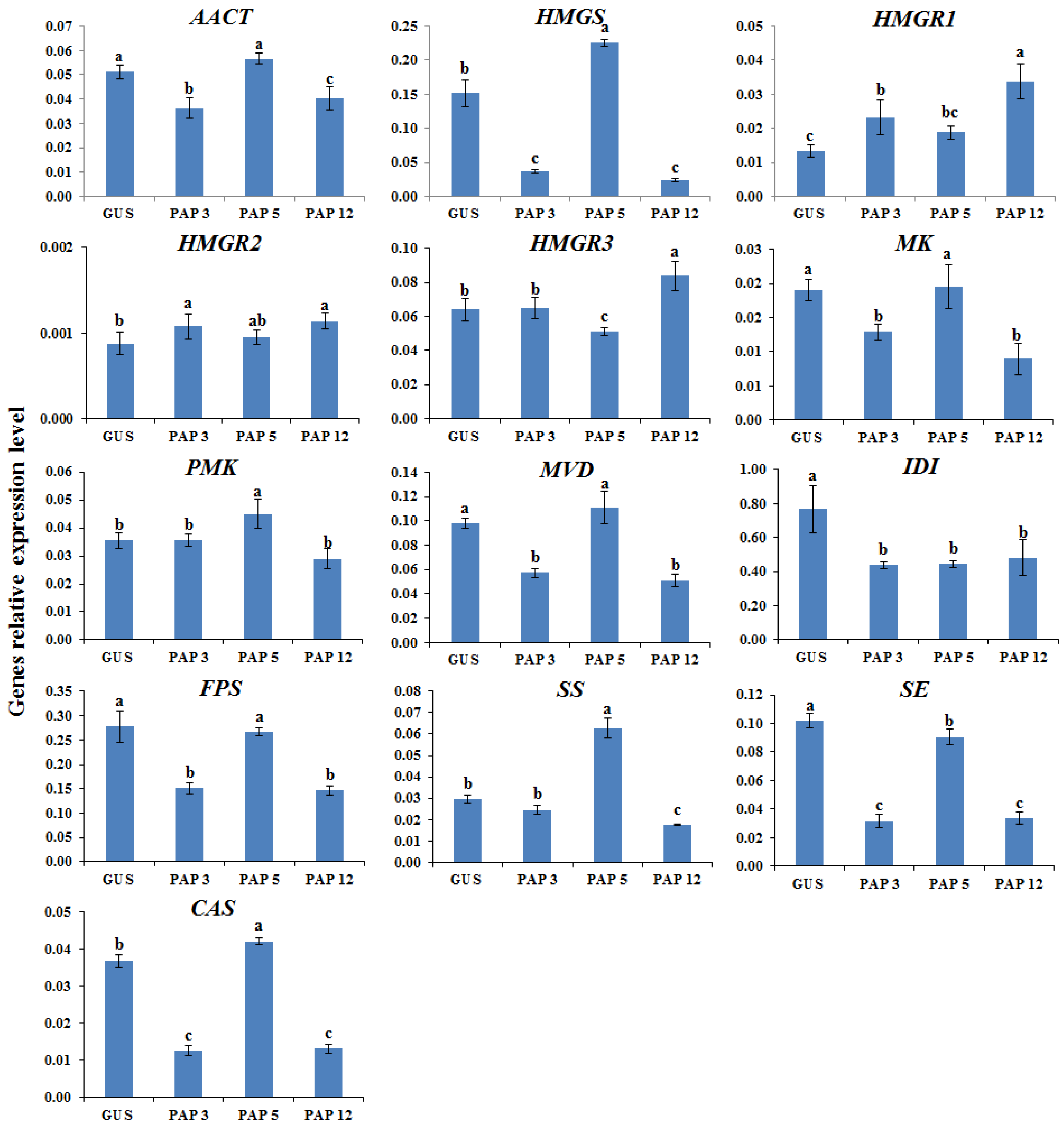
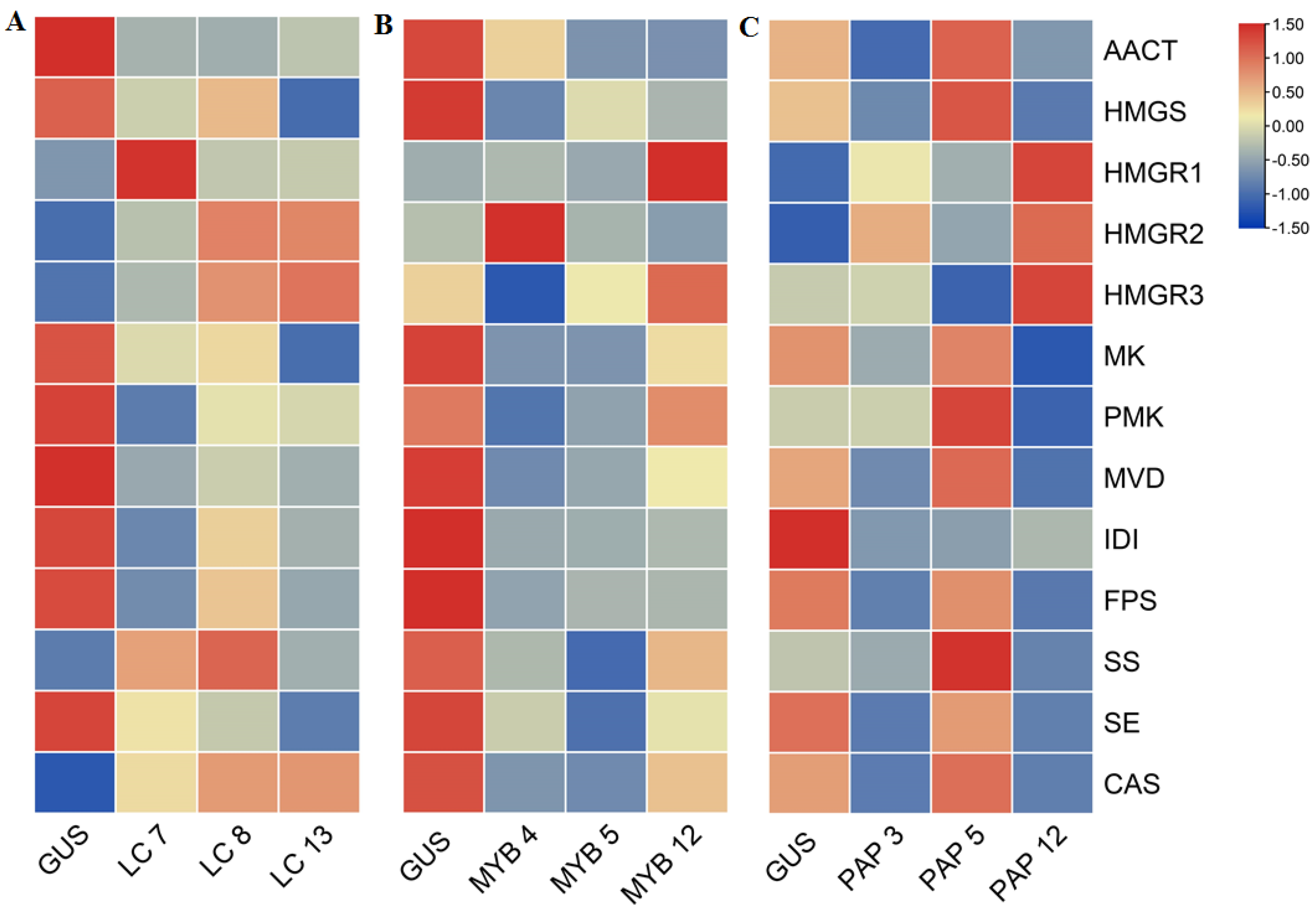
2.2. Transcription Levels of Biosynthetic Genes in AGs Biosynthetic Pathways
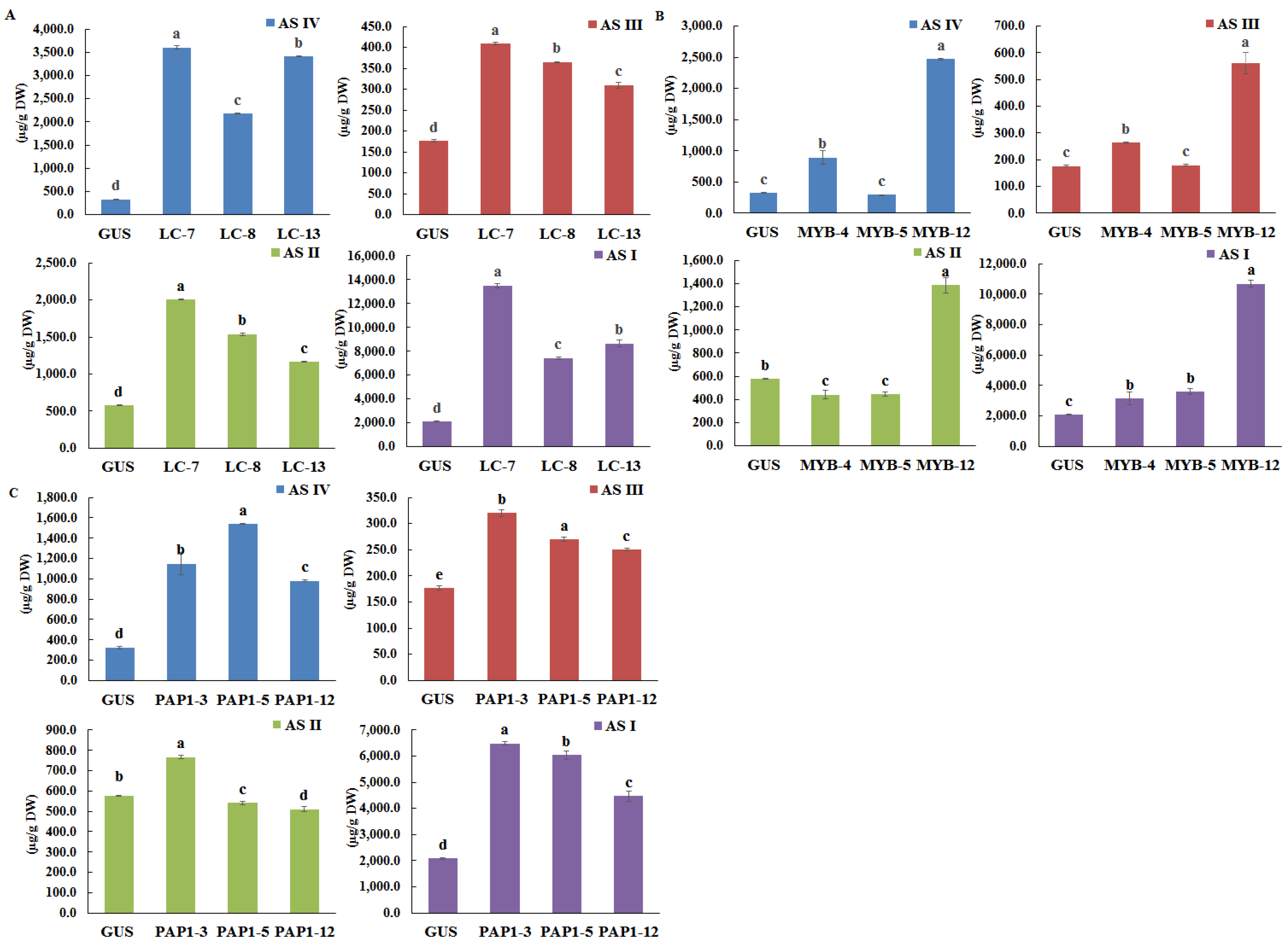
2.3. Synthesis of AGs in Transgenic Hairy Roots of A. membranaceus
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Vector Construction and Generation of Transgenic Hairy Roots
4.3. Quantitative Real-Time PCR
4.4. HPLC Analysis for AGs
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chu, C.; Qi, L.-W.; Liu, E.-H.; Li, B.; Gao, W.; Li, P. Radix Astragali (Astragalus): Latest advancements and trends in chemistry, analysis, pharmacology and pharmacokinetics. Curr. Org. Chem. 2010, 14, 1792–1807. [Google Scholar] [CrossRef]
- Kathy, K.A.; Han, Q.-B.; Joshua, K.K. Astragalus membranaceus: A review of its protection against inflammation and gastrointestinal cancers. Am. J. Chin. Med. 2016, 44, 1–22. [Google Scholar]
- Sheik, A.; Kwanwoo, K.; Ganji Lakshmi, V.; Lee, H.; Kim, S.; Kim, E.; Shin, J.-Y.; Oh, S.Y.; Huh, Y.S. The anti-cancerous activity of adaptogenic herb Astragalus membranaceus. Phytomedicine 2021, 91, 153698. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Yu, J.; Ji, H.; Zhang, H.; Zhang, Y.; Liu, H. Extraction of a novel cold-water-soluble polysaccharide from Astragalus membranaceus and its antitumor and immunological activities. Molecules 2018, 23, 62. [Google Scholar] [CrossRef] [Green Version]
- Shan, H.; Zheng, X.; Li, M. The effects of Astragalus membranaceus active extracts on autophagy-related diseases. Int. J. Mol. Sci. 2019, 20, 1904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elabd, H.; Wang, H.-P.; Shaheen, A.; Yao, H.; Abbass, A. Astragalus membranaceus (AM) enhances growth performance and antioxidant stress profiles in bluegill sunfish (Lepomis macrochirus). Fish Physiol. Biochem. 2016, 42, 955–966. [Google Scholar] [CrossRef]
- Zheng, Y.; Ren, W.; Zhang, L.; Zhang, Y.; Liu, D.; Liu, Y. A review of the pharmacological action of Astragalus Polysaccharide. Front. Pharmacol. 2020, 11, 349. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Guo, S.; Zhu, Y.; Yan, H.; Qian, D.-W.; Wang, H.-Q.; Yu, J.-Q.; Duan, J.-A. Comparative analysis of twenty-five compounds in different parts of Astragalus membranaceus var. mongholicus and Astragalus membranaceus by UPLC-MS/MS. J. Pharm. Anal. 2019, 9, 392–399. [Google Scholar]
- Napolitano, A.; Akay, S.; Mari, A.; Bedir, E.; Pizza, C.; Piacente, S. An analytical approach based on ESI-MS, LC–MS and PCA for the quali–quantitative analysis of cycloartane derivatives in Astragalus spp. J. Pharm. Biomed. Anal. 2013, 85, 46–54. [Google Scholar] [CrossRef]
- Guo, T.; Liu, Z.-L.; Zhao, Q.; Zhao, Z.-M.; Liu, C.-H. A combination of astragaloside I, levistilide A and calycosin exerts anti-liver fibrosis effects in vitro and in vivo. Acta Pharmacol. Sin. 2018, 39, 1483–1492. [Google Scholar]
- Wang, Z.-F.; Ma, D.-G.; Zhu, Z.; Mu, Y.-P.; Yang, Y.-Y.; Feng, L.; Yang, H.; Liang, J.-Q.; Liu, Y.-Y.; Liu, L.; et al. Astragaloside IV inhibits pathological functions of gastric cancer-associated fibroblasts. World J. Gastroenterol. 2018, 23, 8512–8525. [Google Scholar]
- Fu, Y.; Cai, J.; Xi, M.; He, Y.; Zhao, Y.; Zheng, Y.; Zhang, Y.; Xi, J.; He, Y. Neuroprotection effect of Astragaloside IV from 2-DG-induced endoplasmic reticulum stress. Oxid. Med. Cell. Longev. 2020, 2020, 9782062. [Google Scholar]
- Hao, M.; Liu, Y.; Chen, P.; Jiang, H.; Kuang, H.-Y. Astragaloside IV protects RGC-5 cells against oxidative stress. Neural Regen. Res. 2018, 13, 1081–1086. [Google Scholar]
- Indu, P.; Arunagirinathan, N.; Rameshkumar Marimuthu, R.; Sangeetha, K.; Divyadarshini, A.; Rajarajan, S. Antiviral activity of astragaloside II, astragaloside III and astragaloside IV compounds against dengue virus: Computational docking and in vitro studies. Microb. Pathog. 2020, 152, 104563. [Google Scholar] [CrossRef]
- Song, J.-Z.; Mo, S.-F.; Yip, Y.-K.; Qiao, C.-F.; Han, Q.-B.; Xu, H.-X. Development of microwave assisted extraction for the simultaneous determination of isoflavonoids and saponins in Radix Astragali by high performance liquid chromatography. J. Sep. Sci. 2007, 30, 819–824. [Google Scholar] [CrossRef]
- Jiao, J.; Gai, Q.-Y.; Wang, W.; Luo, M.; Zu, Y.-G.; Fu, Y.-J.; Ma, W. Enhanced astragaloside production and transcriptional responses of biosynthetic genes in Astragalus membranaceus hairy root cultures by elicitation with methyl jasmonate. Biochem. Eng. J. 2016, 105, 339–346. [Google Scholar] [CrossRef]
- Tuan, P.A.; Chung, E.; Thwe, A.A.; Li, X.; Kim, Y.B.; Mariadhas, V.A.; Al-Dhabi, N.A.; Lee, J.H.; Park, S.U. Transcriptional profiling and molecular characterization of astragalosides, calycosin, and calycosin-7-O-β-D-glucoside biosynthesis in the hairy roots of Astragalus membranaceus in response to methyl jasmonate. J. Agric. Food Chem. 2015, 63, 6231–6240. [Google Scholar] [CrossRef]
- Park, Y.J.; Kim, J.K.; Park, S.U. Yeast extract improved biosynthesis of astragalosides in hairy root cultures of Astragalus membranaceus. Prep. Biochem. Biotechnol. 2021, 51, 467–474. [Google Scholar] [CrossRef]
- Gai, Q.-Y.; Jiao, J.; Luo, M.; Wang, W.; Zhao, C.-J.; Fu, Y.-J.; Ma, W. UV elicitation for promoting astragaloside production in Astragalus membranaceus hairy root cultures with transcriptional expression of biosynthetic genes. Ind. Crops Prod. 2016, 84, 350–357. [Google Scholar] [CrossRef]
- Naing, A.H.; Kim, C.K. Roles of R2R3-MYB transcription factors in transcriptional regulation of anthocyanin biosynthesis in horticultural plants. Plant Mol. Biol. 2018, 98, 1–18. [Google Scholar] [CrossRef]
- Allan, A.C.; Hellens, R.P.; Laing, W.A. MYB transcription factors that colour our fruit. Trends Plant Sci. 2008, 13, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Deng, L.; Du, M.; Zhao, J.; Chen, Q.; Huang, T.; Jiang, H.; Li, C.-B.; Li, C. A transcriptional network promotes anthocyanin biosynthesis in tomato flesh. Mol. Plant 2020, 13, 42–58. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yang, B.; Liu, J.; Guo, D.; Hou, J.; Chen, S.; Song, B.; Xie, C. Analysis of structural genes and key transcription factors related to anthocyanin biosynthesis in potato tubers. Sci. Hortic. 2017, 225, 310–316. [Google Scholar]
- Katja, K.; Declan, J.L.; Nick, W.A.; Nelli, M.; Tony, M.; Andrew, C.A.; Bilal, M.A.; Hely, H.; Richard, V.E.; Laura, J. MYBA and MYBPA transcription factors co-regulate anthocyanin biosynthesis in blue-coloured berries. New Phytol. 2021, 232, 1350–1367. [Google Scholar]
- Ban, Y.; Honda, C.; Hatsuyama, Y.; Igarashi, M.; Bessho, H.; Moriguchi, T. Isolation and functional analysis of a MYB transcription factor gene that is a key regulator for the development of red coloration in apple skin. Plant Cell Physiol. 2007, 48, 958–970. [Google Scholar] [CrossRef]
- Zhang, Z.; Shi, Y.; Ma, Y.; Yang, X.; Yin, X.; Zhang, Y.; Xiao, Y.; Liu, W.; Li, Y.; Li, S.; et al. The strawberry transcription factor FaRAV1 positively regulates anthocyanin accumulation by activation of FaMYB10 and anthocyanin pathway genes. Plant Biotechnol. J. 2020, 18, 2267–2279. [Google Scholar]
- Tian, J.; Zhang, J.; Han, Z.-Y.; Song, T.-T.; Li, J.-Y.; Wang, Y.-R.; Yao, Y.-C. McMYB12 Transcription factors co-regulate proanthocyanidin and anthocyanin biosynthesis in Malus crabapple. Sci. Rep. 2017, 7, 43715. [Google Scholar]
- Deng, B.; Huang, Z.; Ge, F.; Liu, D.; Lu, R.; Chen, C. An AP2/ERF Family Transcription factor PnERF1 raised the biosynthesis of saponins in Panax notoginseng. J. Plant Growth Regul. 2017, 36, 691–701. [Google Scholar]
- Chen, Q.; Yu, Y.; Zhang, X.; Zhao, R.; Zhang, J.; Liu, D.; Cui, X.; Ge, F. The transcription factor PjERF1 enhances the biosynthesis of triterpenoid saponins in Panax japonicus. Plant Biotechnol. Rep. 2021, 15, 597–607. [Google Scholar] [CrossRef]
- Sun, M.; Shi, M.; Wang, Y.; Huang, Q.; Yuan, T.; Wang, Q.; Wang, C.; Zhou, W.; Kai, G. The biosynthesis of phenolic acids is positively regulated by the JA-responsive transcription factor ERF115 in Salvia miltiorrhiza. J. Exp. Bot. 2019, 70, 243–254. [Google Scholar]
- Zhou, W.; Shi, M.; Deng, C.; Lu, S.; Huang, F.; Wang, Y.; Kai, G. The methyl jasmonate-responsive transcription factor SmMYB1 promotes phenolic acid biosynthesis in Salvia miltiorrhiza. Hortic. Res. 2021, 8, 10. [Google Scholar]
- Li, L.; Wang, D.; Zhou, L.; Yu, X.; Yan, X.; Zhang, Q.; Li, B.; Liu, Y.; Zhou, W.; Cao, X.; et al. JA-responsive transcription factor SmMYB97 promotes phenolic acid and tanshinone accumulation in Salvia miltiorrhiza. J. Agric. Food Chem. 2020, 68, 14850–14862. [Google Scholar]
- Lichtenthaler, H.K.; Rohmer, M.; Schwender, J. Two independent biochemical pathways for isopentenyl diphosphate and isoprenoid biosynthesis in higher plants. Physiol. Plant. 1997, 101, 643–652. [Google Scholar] [CrossRef]
- Kim, Y.B.; Thwe, A.A.; Li, X.; Tuan, P.A.; Lee, S.; Lee, J.W.; Arasu, M.V.; Al-Dhabi, N.A.; Park, S.U. Accumulation of astragalosides and related gene expression in different organs of Astragalus membranaceus Bge. var mongholicus (Bge.). Molecules 2014, 19, 10922–10935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, M.; Liao, P.; Nile, S.H.; Georgiev, M.I.; Kai, G. Biotechnological Exploration of Transformed Root Culture for Value-Added Products. Trends Biotechnol. 2021, 39, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.J.; Thwe, A.A.; Li, X.; Kim, Y.J.; Kim, J.K.; Arasu, M.V.; Al-Dhabi, N.A.; Park, S.U. Triterpene and flavonoid biosynthesis and metabolic profiling of hairy roots, adventitious roots, and seedling roots of Astragalus membranaceus. J. Agric. Food Chem. 2015, 63, 8862–8869. [Google Scholar] [CrossRef] [PubMed]
- Hirotani, M.; Zhou, Y.; Lui, H.; Furuya, T. Astragalosides from hairy root cultures of Astragalus membranaceus. Phytochemistry 1994, 36, 665–670. [Google Scholar] [CrossRef]
- Jiao, J.; Gai, Q.-Y.; Fu, Y.-J.; Ma, W.; Yao, L.-P.; Feng, C.; Xia, X.-X. Optimization of Astragalus membranaceus hairy roots induction and culture conditions for augmentation production of astragalosides. Plant Cell Tissue Organ Cult. 2015, 120, 1117–1130. [Google Scholar] [CrossRef]
- Gai, Q.-Y.; Jiao, J.; Luo, M.; Wang, W.; Yao, L.-P.; Fu, Y.-J. Deacetylation biocatalysis and elicitation by immobilized Penicillium canescens in Astragalus membranaceus hairy root cultures: Towards the enhanced and sustainable production of astragaloside IV. Plant Biotechnol. J. 2017, 15, 297–305. [Google Scholar] [CrossRef]
- Park, C.H.; Xu, H.; Yeo, H.J.; Park, Y.E.; Hwang, G.-S.; Park, N.I.; Park, S.U. Enhancement of the flavone contents of Scutellaria baicalensis hairy roots via metabolic engineering using maize Lc and Arabidopsis PAP1 transcription factors. Metab. Eng. 2021, 64, 64–73. [Google Scholar] [CrossRef]
- Jin, S.; Hyun, T.K. Ectopic Expression of production of Anthocyanin Pigment 1 (PAP1) improves the antioxidant and anti-melanogenic properties of Ginseng (Panax ginseng C.A. Meyer) hairy roots. Antioxidants 2020, 9, 922. [Google Scholar] [CrossRef] [PubMed]
- Mehrtens, F.; Kranz, H.; Bednarek, P.; Weisshaar, B. The Arabidopsis transcription factor MYB12 is a flavonol-specific regulator of phenylpropanoid biosynthesis. Plant Physiol. 2005, 138, 1083–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd, A.M.; Walbot, V.; Davis, R.W. Arabidopsis and Nicotiana anthocyanin production activated by maize regulators R and C1. Science 1992, 258, 1773–1775. [Google Scholar] [CrossRef] [PubMed]
- Borevitz, J.O.; Xia, Y.; Blount, J.; Dixon, R.A.; Lamb, C. Activation tagging identifies a conserved MYB regulator of phenylpropanoid biosynthesis. Plant Cell 2000, 12, 2383–2393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatica-Arias, A.; Farag, M.A.; Stanke, M.; Matoušek, J.; Wessjohann, L.; Weber, G. Flavonoid production in transgenic hop (Humulus lupulus L.) altered by PAP1/MYB75 from Arabidopsis thaliana L. Plant Cell Rep. 2012, 31, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Gao, M.-J.; Pan, H.-Y.; Cui, D.-J.; Gruber, M.Y. Purple Canola: Arabidopsis PAP1 increases antioxidants and phenolics in Brassica napus leaves. J. Agric. Food Chem. 2010, 58, 1639–1645. [Google Scholar] [CrossRef]
- Ludwig, S.R.; Habera, L.F.; Dellaporta, S.L.; Wessler, S.R. Lc, a member of the maize R gene family responsible for tissue-specific anthocyanin production, encodes a protein similar to transcriptional activators and contains the myc-homology region. Proc. Natl. Acad. Sci. USA 1989, 86, 7092–7096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.-A.; Zhao, T.; Wang, N.; Zheng, S.-s. Ectopic expression of Lc differentially regulated anthocyanin biosynthesis in the floral parts of tobacco (Nicotiana tobacum L.) plants. Bot. Stud. 2016, 57, 24. [Google Scholar] [CrossRef] [Green Version]
- Bradley, J.M.; Deroles, S.C.; Boase, M.R.; Bloor, S.; Swinny, E.; Davies, K.M. Variation in the ability of the maize Lc regulatory gene to upregulate flavonoid biosynthesis in heterologous systems. Plant Sci. 1999, 140, 31–39. [Google Scholar] [CrossRef]
- Song, Y.E.; Wang, X.; Shen, Z.W.; Xu, Y.; Li, J.Y. Expressing the maize anthocyanin regulatory gene Lc increased flavonoid content in the seed of white pericarp rice and purple pericarp rice. Russ. J. Genet. 2013, 49, 1127–1133. [Google Scholar] [CrossRef]
- Wang, H.; Yang, J.; Zhang, M.; Fan, W.; Firon, N.; Pattanaik, S.; Yuan, L.; Zhang, P. Altered phenylpropanoid metabolism in the maize Lc-expressed sweet potato (Ipomoea batatas) affects storage root development. Sci. Rep. 2016, 6, 18645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, C.H.; Lee, K.W.; Lee, S.H.; Jeong, Y.J.; Woo, S.G.; Chun, H.; Park, Y.I.; Kwak, S.S.; Kim, C.Y. Heterologous expression of IbMYB1a by different promoters exhibits different patterns of anthocyanin accumulation in tobacco. Plant Physiol. Biochem. 2015, 89, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Zhao, X.; Ding, X.; Li, Y.; Cao, G.; Chu, Z.; Su, X.; Liu, Y.; Chen, X.; Guo, J.; et al. Integrated functional omics analysis of flavonoid-related metabolism in AtMYB12 transcript factor overexpressed tomato. J. Agric. Food Chem. 2020, 68, 6776–6787. [Google Scholar] [CrossRef] [PubMed]
- Nam Il, P.; Xiaohua, L.; Aye Aye, T.; Sook Young, L.; Su Gwan, K.; Qi, W.; Sang Un, P. Enhancement of rutin in Fagopyrum esculentum hairy root cultures by the Arabidopsis transcription factor AtMYB12. Biotechnol. Lett. 2011, 34, 577–583. [Google Scholar]
- Jun, L.; Yuka, H.-L.; Matthew, D.D.; Qianjin, F.; Judith, R.R.; Zhipeng, Q.; David, L.A. Long read reference genome-free reconstruction of a full-length transcriptome from Astragalus membranaceus reveals transcript variants involved in bioactive compound biosynthesis. Cell Discov. 2017, 3, 17031. [Google Scholar]
- Chappell, J.; Wolf, F.; Proulx, J.; Cuellar, R.; Saunders, C. Is the reaction catalyzed by 3-hydroxy-3-methylglutaryl coenzyme A reductase a rate-limiting step for isoprenoid biosynthesis in plants? Plant Physiol. 1995, 109, 1337–1343. [Google Scholar] [CrossRef] [Green Version]
- Hey, S.J.; Powers, S.J.; Beale, M.H.; Hawkins, N.D.; Ward, J.L.; Halford, N.G. Enhanced seed phytosterol accumulation through expression of a modified HMG-CoA reductase. Plant Biotechnol. J. 2006, 4, 219–229. [Google Scholar] [CrossRef]
- Kim, Y.-K.; Kim, J.K.; Kim, Y.B.; Lee, S.; Kim, S.-U.; Park, S.U. Enhanced accumulation of phytosterol and triterpene in hairy root cultures of Platycodon grandiflorum by overexpression of Panax ginseng 3-hydroxy-3-methylglutaryl-coenzyme A reductase. J. Agric. Food Chem. 2013, 61, 1928–1934. [Google Scholar] [CrossRef]
- Bach, T.J. Some new aspects of isoprenoid biosynthesis in plants—A review. Lipids 1995, 30, 191–202. [Google Scholar] [CrossRef]
- Karel, M.; Jacob, P.; Dieter, B.; Philipp, A.; René, C.; Sven, S.; Tessa, M.; Jan, M.; Prashant, D.S.; Laurens, P.; et al. The ancient CYP716 family is a major contributor to the diversification of eudicot triterpenoid biosynthesis. Nat. Commun. 2017, 8, 14153. [Google Scholar]
- Satoru, S.; Kazuki, S. Triterpenoid biosynthesis and engineering in plants. Front. Plant Sci. 2011, 2, 25. [Google Scholar]
- Harrison, D.M. The biosynthesis of triterpenoids and steroids. Nat. Prod. Rep. 1985, 2, 525–560. [Google Scholar] [CrossRef] [PubMed]
- Suzanne, P.; Birger Lindberg, M.; Søren, B. On the origin of family 1 plant glycosyltransferases. Phytochemistry 2003, 62, 399–413. [Google Scholar]
- Xu, W.; Bak, S.; Decker, A.; Paquette, S.M.; Feyereisen, R.; Galbraith, D.W. Microarray-based analysis of gene expression in very large gene families: The cytochrome P450 gene superfamily of Arabidopsis thaliana. Gene 2001, 272, 61–74. [Google Scholar] [CrossRef]
- Yao, C.; Hanbing, L.; Xuejiao, T.; Zaimin, L.; Xin, Z.; Dalong, L.; Xinmei, J.; Xihong, Y. Identification and analysis of CYP450 and UGT supergene family members from the transcriptome of Aralia elata (Miq.) seem reveal candidate genes for triterpenoid saponin biosynthesis. BMC Plant Biol. 2020, 20, 214. [Google Scholar]
- Li, X.; Thwe, A.A.; Park, N.I.; Suzuki, T.; Kim, S.J.; Park, S.U. Accumulation of phenylpropanoids and correlated gene expression during the development of tartary buckwheat sprouts. J. Agric. Food Chem. 2012, 60, 5629–5635. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.H.; Kim, J.K.; Park, S.U. Heterologous Expression of Three Transcription Factors Differently Regulated Astragalosides Metabolic Biosynthesis in Astragalus membranaceus Hairy Roots. Plants 2022, 11, 1897. https://doi.org/10.3390/plants11141897
Li XH, Kim JK, Park SU. Heterologous Expression of Three Transcription Factors Differently Regulated Astragalosides Metabolic Biosynthesis in Astragalus membranaceus Hairy Roots. Plants. 2022; 11(14):1897. https://doi.org/10.3390/plants11141897
Chicago/Turabian StyleLi, Xiao Hua, Jae Kwang Kim, and Sang Un Park. 2022. "Heterologous Expression of Three Transcription Factors Differently Regulated Astragalosides Metabolic Biosynthesis in Astragalus membranaceus Hairy Roots" Plants 11, no. 14: 1897. https://doi.org/10.3390/plants11141897