
Rosmarinic Acid Production from Origanum dictamnus L. Root Liquid Cultures In Vitro
 ,
,
Abstract
:1. Introduction
2. Results
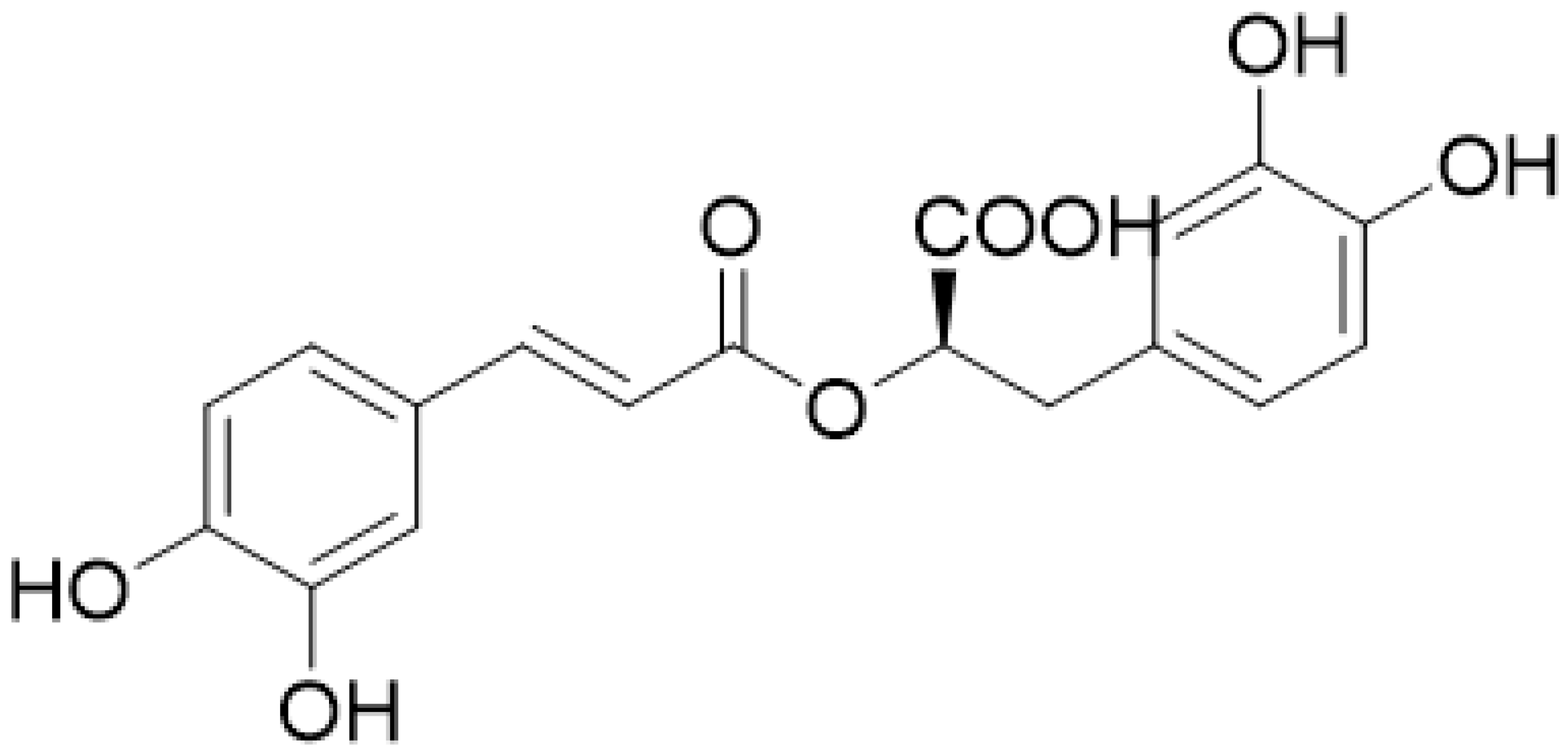
2.1. Evaluation of Rosmarinic Acid Production by HPLC-UV and NMR
2.2. In vitro Adventitious Root Culture Establishment
2.3. In vitro Adventitious Root Culture
2.3.1. Adventitious Roots from Leaf Callus
2.3.2. Adventitious Roots from Petiole Callus
2.3.3. Adventitious Roots from Root Callus
2.3.4. Combined Effect of Explant Type (Leaf, Petiole and Root Callus Derived Roots) and Medium Composition under 240 Days of Total Incubation
3. Discussion
4. Materials and Methods
4.1. In vitro Adventitious Root Culture Establishment
4.2. In vitro Adventitious Root Culture
4.3. Sample Preparation for HPLC Analysis
4.4. Chemicals for the HPLC Analysis
4.5. HPLC-PDA-MS Analysis Instrumentation and Quantitative Determination of Rosmarinic Acid
4.6. Sample Preparation for NMR Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guan, H.; Luo, W.; Bao, B.; Cao, Y.; Cheng, F.; Yu, S.; Fan, Q.; Zhang, L.; Wu, Q.; Shan, M. A comprehensive review of rosmarinic acid: From phytochemistry to pharmacology and its new insight. Molecules 2022, 27, 3292. [Google Scholar] [CrossRef] [PubMed]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouységu, L. Plant polyphenols: Chemical properties, biological activities, and synthesis. Angew. Chem. 2011, 50, 586–621. [Google Scholar] [CrossRef] [PubMed]
- Lešnik, S.; Furlan, V.; Bren, U. Rosemary (Rosmarinus officinalis L.): Extraction techniques, analytical methods and health-promoting biological effects. Phytochem Rev. 2021, 20, 1273–1328. [Google Scholar] [CrossRef]
- Park, S.U.; Uddin, M.R.; Xu, H.; Kim, Y.K.; Lee, S.Y. Biotechnological applications for rosmarinic acid production in plant. Afr. J. Biotechnol. 2008, 7, 4959–4965. [Google Scholar]
- Hitl, M.; Kladar, N.; Gavarić, N.; Božin, B. Rosmarinic acid-human pharmacokinetics and health benefits. Planta Med. 2021, 87, 273–282. [Google Scholar] [CrossRef]
- Nadeem, M.; Imran, M.; Aslam Gondal, T.; Imran, A.; Shahbaz, M.; Muhammad Amir, R.; Wasim Sajid, M.; Batool Qaisrani, T.; Atif, M.; Hussain, G.; et al. Therapeutic potential of rosmarinic acid: A comprehensive review. Appl. Sci 2019, 9, 3139. [Google Scholar] [CrossRef] [Green Version]
- Georgiev, M.I.; Kuzeva, S.L.; Pavlov, A.I.; Kovacheva, E.G.; Ilieva, M.P. Elicitation of rosmarinic acid by Lavandula vera MM cell suspension culture with abiotic elicitors. World J. Microbiol. Biotechnol. 2007, 23, 301–304. [Google Scholar] [CrossRef]
- Sanchez-Medina, A.; Etheridge, C.J.; Hawkes, G.E.; Hylands, P.J.; Pendry, B.A.; Hughes, M.J.; Corcoran, O. Comparison of rosmarinic acid content in commercial tinctures produced from fresh and dried lemon balm (Melissa officinalis). J. Pharm. Pharm. Sci. 2007, 10, 455–463. [Google Scholar] [CrossRef] [Green Version]
- Sahraroo, A.; Babalar, M.; Mirjalili, M.H.; Moghaddam, M.R.F.; Ebrahimic, S.N. In vitro callus induction and rosmarinic acid quantification in callus culture of Satureja khuzistanica Jamzad (Lamiaceae). Iran. J. Pharm. Res. 2014, 13, 1447–1456. [Google Scholar] [CrossRef]
- Mitropoulou, G.; Fitsiou, E.; Stavropoulou, E.; Papavassilopoulou, E.; Vamvakias, M.; Pappa, A.; Oreopoulou, A.; Kourkoutas, Y. Composition, antimicrobial, antioxidant, and antiproliferative activity of Origanum dictamnus (dittany) essential oil. Microb Ecol. Health Dis. 2015, 26, 26543. [Google Scholar] [CrossRef]
- Triantaphyllou, K.; Blekas, G.; Boskou, D. Antioxidative properties of water extracts obtained from herbs of the species Lamiaceae. Int. J. Food Sci. Nutr. 2001, 52, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Couladis, M.; Tzakou, O.; Verykokidou, E.; Harvala, C. Screening of some Greek aromatic plants for antioxidant activity. Phytother Res. 2003, 17, 194–195. [Google Scholar] [CrossRef] [PubMed]
- Kouri, G.; Tsimogiannis, D.; Bardouki, H.; Oreopoulou, V. Extraction and analysis of antioxidant components from Origanum dictamnus. Innov. Food Sci. Emerg. Technol. 2007, 8, 155–162. [Google Scholar] [CrossRef]
- Lagouri, V.; Boskou, D. Nutrient antioxidants in oregano. Int. J. Food Sci. Nutr. 1996, 47, 493–497. [Google Scholar] [CrossRef]
- Karanika, M.S.; Komaitis, M.; Aggelis, G. Effect of aqueous extracts of some plants of Lamiaceae family on the growth of Yarrowiali polytica. Int. J. Food Microbiol. 2001, 64, 175–181. [Google Scholar] [CrossRef]
- Chorianopoulos, N.; Kalpoutzakis, E.; Aligiannis, N.; Mitaku, S.; Nychas, G.J.; Haroutounian, S.A. Essential oils of Satureja, Origanum, and Thymus species: Chemical composition and antibacterial activities against food borne pathogens. J. Agric. Food Chem. 2004, 52, 8261–8267. [Google Scholar] [CrossRef]
- Fokialakis, N.; Kalpoutzakis, E.; Tekwani, B.L.; Khan, S.I.; Kobaisy, M.; Skaltsounis, A.L.; Duke, S.O. Evaluation of the antimalarial and antileishmanial activity of plants from the Greek island of Crete. J. Nat. Med. 2007, 61, 38–45. [Google Scholar] [CrossRef]
- Liolios, C.C.; Gortzi, O.; Lalas, S.; Tsaknis, J.; Chinou, I. Liposomal incorporation of carvacrol and thymol isolated from the essential oil of Origanum dictamnus L. and in vitro antimicrobial activity. Food Chem. 2009, 112, 77–83. [Google Scholar] [CrossRef]
- Chatzopoulou, A.; Karioti, A.; Gousiadou, C.; Vivancos, V.L.; Kyriazopoulos, P.; Golegou, S.; Skaltsa, H. Depsides and other polar constituents from Origanum dictamnus L. and their in vitro antimicrobial activity in clinical strains. J. Agric. Food Chem. 2010, 58, 6064–6068. [Google Scholar] [CrossRef]
- Chinou, I.; Liolios, C.; Moreau, D.; Roussakis, C. Cytotoxic activity of Origanum dictamnus. Fitoterapia 2007, 78, 342–344. [Google Scholar] [CrossRef]
- Community Herbal Monograph on Origanum dictamnus L., Herba. European Medicines Agency, Committee on Herbal Medicinal Products (HMPC). EMA/HMPC/200429/2012. Available online: https://www.ema.europa.eu/en/documents/herbal-monograph/final-community-herbal-monograph-origanum-dictamnus-l-herba-first-version_en.pdf (accessed on 28 January 2014).
- Assessment Report on Origanum dictamnus L., Herba. European Medicines Agency, Committee on Herbal Medicinal Products (HMPC). EMA/HMPC/200431/2012. Available online: https://www.ema.europa.eu/en/documents/herbal-report/final-assessment-report-origanum-dictamnus-l-herba-first-version_en.pdf (accessed on 9 July 2013).
- Bauer, N.; Leljak-Levanić, D.; Jelaska, S. Rosmarinic acid synthesis in transformed callus culture of Coleus blumei Benth. Z Nat. C 2004, 59, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Zhang, L.; Gao, S.; Saechao, S.; Di, P.; Chen, J.; Chen, W. The c4h, tat, hppr and hppd genes prompted engineering of rosmarinic acid biosynthetic pathway in Salvia miltiorrhiza hairy root cultures. PLoS ONE 2011, 6, e29713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, M.; Simmonds, M.S. Rosmarinic acid. Phytochemistry 2003, 62, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Hakkim, F.L.; Kalyani, S.; Essa, M.; Girija, S.; Song, H. Production of rosmarinic acid in Ocimum sanctum (L) cell suspension cultures by the influence of sucrose, phenylalanine, yeast extract, and methyl jasmonate. Int. J. Biol. Med. Res. 2011, 2, 1070–1074. [Google Scholar]
- Qian, J.; Guiping, L.; Xiujun, L.; Xincai, H.; Hongmei, L. Influence of growth regulators and sucrose concentrations on growth and rosmarinic acid production in calli and suspension cultures of Coleus blumei. Nat. Prod. Res. 2009, 23, 127–137. [Google Scholar] [CrossRef]
- Wu, C.F.; Karioti, A.; Rohr, D.; Bilia, A.R.; Efferh, T. Production of rosmarinic acid and salvianolic acid B from callus culture of Salvia miltiorrhiza with cytotoxicity towards acute lymphoblastic leukemia cells. Food Chem. 2016, 201, 292–297. [Google Scholar] [CrossRef]
- Grzegorczyk, I.; Bilichowski, I.; Mikiciuk-Olasik, E.; Wysokińska, H. In vitro cultures of Salvia officinalis L. as a source of antioxidant compounds. Acta Soc. Bot. Pol. 2005, 74, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Karam, N.S.; Jawad, F.M.; Arikat, N.A.; Shibli, R.A. Growth and rosmarinic acid accumulation in callus, cell suspension, and root cultures of wild Salvia fruticosa. Plant Cell Tiss. Organ Cult. 2003, 73, 117–121. [Google Scholar] [CrossRef]
- Pavlov, A.I.; Ilieva, M.P.; Panchev, I.N. Nutrient medium optimization for rosmarinic acid production by Lavandula vera MM cell suspension. Biotechnol. Prog. 2000, 16, 668–670. [Google Scholar] [CrossRef]
- Biswas, T. Elicitor induced increased rosmarinic acid content of in vitro root cultures of Ocimum basilicum L. (sweet basil). Plant Sci. Today 2020, 7, 157–163. [Google Scholar] [CrossRef]
- Sivakumar, G. Bioreactor technology: A novel industrial tool for high-tech production of bioactive molecules and biopharmaceuticals from plant root. Biotechnol. J. 2006, 12, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Paek, K.Y.; Murthy, H.N.; Hahn, E.J.; Zhong, J.J. Large-scale culture of ginseng adventitious roots for production of ginsenosides. Adv. Biochem. Eng. Biotechnol. 2009, 113, 151–176. [Google Scholar] [CrossRef]
- Sudha, C.G.; Seeni, S. Establishment and analysis of fast-grown normal root culture of Decalepis arayalpathra, a rare endemic medicinal plant. Curr. Sci. 2001, 81, 371–374. [Google Scholar]
- Govari, S.; Paloukopoulou, C.; Soulioti, A.; Tasi, G.; Karioti, A. Origanum dictamnus, an Important Greek Species with Long Ethnomedicinal History as Anti-Inflammatory Agent, is Rich in Phenolic Content as Confirmed by HPLC-PDA-MS and Phytochemical Analyses. In Proceedings of the 19th International Congress of International Society for Ethnopharmacology (ISE), Dresden, Germany, 12–14 June 2019. [Google Scholar]
- Fattahi, M.; Nazeri, V.; Torras-Claveria, L.; Sefidkon, F.; Cusido, R.M.; Zamani, Z.; Palazon, J. A new biotechnological source of rosmarinic acid and surface flavonoids: Hairy root cultures of Dracocephalum kotschyi Boiss. Ind. Crop Prod. 2013, 50, 256–263. [Google Scholar] [CrossRef]
- Bais, H.P.; Walker, T.S.; Schweizer, H.P.; Vivanco, J.M. Root specific elicitation and antimicrobial activity of rosmarinic acid in hairy root cultures of Ocimum basilicum. Plant Physiol. Biochem. 2002, 40, 983–995. [Google Scholar] [CrossRef]
- Gerdakaneh, M.; Badakhshan, H.; Mohamadi, M.; Arji, I. Effect of different media and growth regulators on micropropagation of GF677. J. Plant Prod. 2020, 43, 241–254. [Google Scholar] [CrossRef]
- Chimdessa, E. Composition and preparation of plant tissue culture medium. J. Tissue Cult. Bioengin. 2020, 3, 120. [Google Scholar] [CrossRef]
- Minutolo, M.; Chiaiese, P.; Di Matteo, A.; Errico, A.; Corrado, G. Accumulation of ascorbic acid in tomato cell culture: Influence of the genotype, source explant and time of in vitro cultivation. Antioxidants 2020, 9, 222. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.R.; Ravishankar, G.A. Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv. 2002, 20, 101–153. [Google Scholar] [CrossRef]
- Ramawat, K.G.; Mathur, M. Factors affecting the production of secondary metabolites. In Biotechnology: Secondary Metabolites, Plants and Microbes; Ramawat, K.G., Merillon, J.M., Eds.; Science Publ Inc.: Jersey/Plymouth, UK, 2007; pp. 59–102. [Google Scholar]
- Sharafzadeh, S.; Zare, M. Influence of growth regulators on growth and secondary metabolites of some medicinal plants from Lamiaceae family. Adv. Env. Biol. 2001, 5, 2296–2302. [Google Scholar]
- Scravoni, J.; Vasconcellos, M.C.; Valmorbida, J.; Ferri, A.F.; Marques, M.O.M.; Ono, E.O.; Rodrigues, J.D. Rendimento e composicao química do oleo essencial de Mentha piperita L. submetida a aplicacoes de giberelina e citocinina. Rev. Bras. Pl. Med. 2006, 8, 40–43. [Google Scholar]
- Mehrotra, S.; Goel, M.K.; Kukreja, A.K.; Mishra, B.N. Efficiency of liquid culture systems over conventional micropropagation: A progress towards commercialization. Afr. J. Biotechnol. 2007, 6, 1484–1492. [Google Scholar] [CrossRef]
- Sandal, I.; Bhattacharya, A.; Ahuja, P.S. An efficient liquid culture system for tea shoot proliferation. Plant Cell Tiss. Organ Cult. 2001, 65, 75–80. [Google Scholar] [CrossRef]
- Korkor, A.M.; Mohamed, S.A.; Abd El-kafie, O.M.; Gohar, A.A. Adaptation of the in vitro culture of Origanum majorana L. for production of phenolic acids. IOSR J. Pharm. Biol. Sci. 2017, 12, 30–38. [Google Scholar] [CrossRef]
- Hartmann, H.T.; Kester, D.E.; Davies, J.F.T.; Geneve, R.L. Plant Propagation: Principles and Practices, 7th ed.; Prentice-Hall: New Jersey, NY, USA, 2007. [Google Scholar]
- Ling, A.P.K.; Chin, M.F.; Hussein, S. Adventitious root production of Centella asiatica in response to plant growth regulators and sucrose concentration. Med. Aromat. Plant Sci. Biotechnol. 2009, 3, 36–41. [Google Scholar]
- Hasan, N.A.; Hussein, S.; Ibrahim, R. Plant growth regulator effect on adventitious roots induction of Labisia pumila. Mal. J. Fun Appl. Sci. 2014, 10, 49–52. [Google Scholar] [CrossRef]
- Nagarajan, A.; Arivalagan, U.; Rajagru, P. In vitro root induction and studies on antibacterial activity of root extract of Costus igneus on clinically important human pathogens. J. Microbiol. Biotech Res. 2011, 1, 67–76. [Google Scholar]
- Mansouri, A.; Fadavi, A.; Mortazavian, S.M.M. Effects of length and position of hypocotyl explants on Cuminum cyminum L. callogensis by image processing analysis. Plant Cell Tiss. Organ Cult. 2015, 121, 657–666. [Google Scholar] [CrossRef]
- Saeedpour, A.; Jahanbakhsh Godehkahriz, S.; Lohrasebi, T.; Esfahani, K.; Salmanian, A.H.; Razavi, K. The effect of endogenous hormones, total antioxidant and total phenol changes on regeneration of barley cultivars. Iran. J. Biotechnol. 2021, 19, 30–39. [Google Scholar] [CrossRef]
- Shetty, K. Rosmarinic acid biosynthesis and mechanism of action. In Functional Foods and Biotechnology; Shetty, K., Paliyath, G., Pometto, A.L., Levin, R.E., Eds.; CRC Taylor & Francis Group: Boca Raton, NY, USA, 2007. [Google Scholar]
- Benedec, D.; Hanganu, D.; Oniga, I.; Tiperciuc, B.; Olah, N.K.; Raita, O.; Bischin, C.; Silaghi-Dumitrescu, R.; Vlase, L. Assessment of rosmarinic acid content in six Lamiaceae species extracts and their antioxidant and antimicrobial potential. Pak. J. Pharm. Sci. 2015, 28, 2297–2303. [Google Scholar]
- Kintzios, S.; Makri, O.; Panagiotopoulos, E.; Scapeti, M. In vitro rosmarinic acid accumulation in sweet basil (Ocimum basilicum L.). Biotechnol. Lett. 2003, 25, 405–408. [Google Scholar] [CrossRef]
- Del Baño, M.J.; Lorente, J.; Castillo, J.; Benavente-García, O.; del Río, J.A.; Ortuño, A.; Quirin, K.W.; Gerard, D. Phenolic diterpenes, flavones, and rosmarinic acid distribution during the development of leaves, flowers, stems, and roots of Rosmarinus officinalis. antioxidant activity. J. Agric. Food Chem. 2003, 51, 4247–4253. [Google Scholar] [CrossRef]
- Bauer, N.; Fulgosi, H.; Jelasaka, S. Overexpression of phenylalanine ammonia-lyase in transgenic roots of Coleus blumei alerts growth and rosmarinic acid synthesis. Food Technol. Biotech 2011, 49, 24–31. [Google Scholar]
- Stalman, M.; Koskamp, A.M.; Luderer, R.; Vernooy, J.H.J.; Wind, J.C.; Wullmes, G.J.; Craes, A.F. Regulation of anthraquinone biosynthesis in cell cultures of Morinda citrifolia. J. Plant Physiol. 2003, 160, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Pépin, M.; Archambault, J.; Chavarie, C.; Cormier, F. Growth kinetics of Vitis vinifera cell suspension cultures: I. Shake flask cultures. Biotechnol. Bioeng. 2004, 47, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Sarropoulou, V.; Maloupa, E.; Grigoriadou, K. In vitro direct organogenesis of the Cretan dittany (Origanum dictamnus L.), an important threatened Greek endemic species. Not. Bot. Horti. Agrobot. Cluj-Napoca 2022, 50, 12715. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised method for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 472–497. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Explant Type | Solid Medium | Liquid Medium | In Vitro Culture Duration (in Days) | F.W. (g) (=Fresh Biomass Growth Rate) | D.W. (g) | F.W./D.W. Ratio | RA (% w/w) | Mean Yield Index * | Yield Index/Explant ** | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Solid Medium | Liquid Medium | Total | |||||||||
| Leaf callus derived roots | ODK3 | ODY1 | 40 | 40 | 80 | 2.8 ± 1.0 e | 0.4 ± 0.1 e | 7.4 ± 0.0 ef | 1.5 ± 0.5 de | 5.8 ± 0.9 e | 0.23 ± 0.04 d |
| 120 | 40 | 160 | 2.7 ± 0.1 e | 0.2 ± 0.0 e | 6.5 ± 0.2 h | 2.2 ± 0.0 cd | 4.6 ± 0.0 f | 0.18 ± 0.00 d | |||
| 200 | 40 | 240 | 15.2 ± 0.0 b | 1.5 ± 0.0 b | 10.1 ± 0.0 b | 5.8 ± 0.6 a | 86.7 ± 8.7 a | 3.47 ± 0.35 a | |||
| ODK4 | ODY1 | 40 | 40 | 80 | 2.1 ± 0.5 e | 0.3 ± 0.1 e | 7.3 ± 0.0 fg | 0.5 ± 0.0 e | 1.5 ± 0.3 g | 0.06 ± 0.01 e | |
| 120 | 40 | 160 | 3.8 ± 0.5 de | 0.5 ± 0.1 e | 7.8 ± 0.2 e | 4.1 ± 0.0 b | 19.9 ± 2.5 cd | 0.80 ± 0.10 bcd | |||
| 200 | 40 | 240 | 19.0 ± 0.2 a | 1.8 ± 0.0 a | 10.3 ± 0.0 b | 5.1 ± 0.6 a | 93.7 ± 12.2 a | 3.75 ± 0.49 a | |||
| ODK3 | ODY2 | 40 | 40 | 80 | 4.4 ± 2.4 de | 0.6 ± 0.2 e | 11.3 ± 0.0 a | 3.8 ± 0.1 b | 22.3 ± 7.9 c | 0.89 ± 0.32 bcd | |
| 120 | 40 | 160 | 4.9 ± 0.3 de | 0.5 ± 0.0 e | 8.9 ± 0.0 c | 5.2 ± 0.1 a | 28.2 ± 1.9 c | 1.13 ± 0.07 bc | |||
| 200 | 40 | 240 | 6.6 ± 0.0 cd | 0.7 ± 0.0 d | 8.9 ± 0.0 c | 1.7 ± 0.4 d | 12.3 ± 2.6 d | 0.49 ± 0.11 cd | |||
| ODK4 | ODY2 | 40 | 40 | 80 | 2.5 ± 1.4 e | 0.4 ± 0.2 e | 6.3 ± 0.0 h | 3.1 ± 0.1 bc | 12.1 ± 5.8 de | 0.48 ± 0.31 cd | |
| 120 | 40 | 160 | 3.1 ± 0.7 e | 0.4 ± 0.1 e | 7.2 ± 0.2 g | 3.9 ± 0.5 b | 16.9 ± 5.2 cd | 0.68 ± 0.21 cd | |||
| 200 | 40 | 240 | 8.3 ± 0.0 c | 1.0 ± 0.0 c | 8.5 ± 0.0 d | 3.7 ± 0.4 b | 36.4 ± 4.3 b | 1.46 ± 0.17 b | |||
| Three-way ANOVA/General Linear Model | |||||||||||
| Solid culture medium composition [ODK3 vs. ODK4] (A) | 0.529 ns | 0.256 ns | 0.000 *** | 0.792 ns | 0.165 ns | 0.165 ns | |||||
| Liquid culture medium composition [ODY1 vs. ODY2] (B) | 0.000 *** | 0.001 ** | 0.000 *** | 0.091 ns | 0.000 *** | 0.000 *** | |||||
| In vitro culture duration in solid medium [40, 120, 200 days] (C) | 0.000 *** | 0.000 *** | 0.000 *** | 0.000 *** | 0.000 *** | 0.000 *** | |||||
| (A) × (B) | 0.074 ns | 0.642 ns | 0.000 *** | 0.904 ns | 0.931 ns | 0.931 ns | |||||
| (A) × (C) | 0.012 * | 0.069 ns | 0.000 *** | 0.014 * | 0.070 ns | 0.070 ns | |||||
| (B) × (C) | 0.000 *** | 0.000 *** | 0.000 *** | 0.000 *** | 0.000 *** | 0.000 *** | |||||
| (A) × (B) × (C) | 0.821 ns | 0.641 ns | 0.000 *** | 0.000 *** | 0.078 ns | 0.078 ns | |||||
| Explant Type | Solid Medium | Liquid Medium | In Vitro Culture Duration (in Days) | F.W. (g) (=Fresh Biomass Growth Rate) | D.W. (g) | F.W./D.W. Ratio | RA (% w/w) | Mean Yield Index * | Yield Index/Explant ** | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Solid Medium | Liquid Medium | Total | |||||||||
| Petiole callus derived roots | ODK3 | ODY1 | 120 | 40 | 160 | 3.1 ± 0.0 d | 0.4 ± 0.0 d | 8.4 ± 0.2 b | 1.8 ± 0.0 d | 6.8 ± 0.1 f | 0.27 ± 0.01 e |
| 200 | 40 | 240 | 18.8 ± 1.0 a | 1.8 ± 0.1 a | 10.7 ± 0.0 a | 6.6 ± 0.0 a | 115.3 ± 5.8 a | 4.61 ± 0.23 a | |||
| ODK4 | ODY1 | 120 | 40 | 160 | 4.2 ± 0.7 d | 0.5 ± 0.1 d | 7.8 ± 0.1 bc | 3.5 ± 0.0 c | 18.8 ± 3.1 e | 0.75 ± 0.12 d | |
| 200 | 40 | 240 | 16.6 ± 0.4 b | 1.6 ± 0.2 a | 10.4 ± 0.9 a | 5.6 ± 0.4 b | 89.9 ± 15.3 b | 3.60 ± 0.61 b | |||
| ODK3 | ODY2 | 120 | 40 | 160 | 3.8 ± 0.6 d | 0.5 ± 0.1 d | 8.3 ± 0.2 bc | 3.6 ± 0.0 c | 16.6 ± 3.3 e | 0.66 ± 0.13 d | |
| 200 | 40 | 240 | 7.2 ± 0.1 c | 0.8 ± 0.0 c | 8.6 ± 0.0 b | 3.7 ± 0.1 c | 31.1 ± 1.2 d | 1.25 ± 0.05 c | |||
| ODK4 | ODY2 | 120 | 40 | 160 | 3.3 ± 0.2 d | 0.5 ± 0.0 d | 7.2 ± 0.2 c | 1.7 ± 0.0 d | 7.8 ± 0.7 f | 0.31 ± 0.03 e | |
| 200 | 40 | 240 | 8.8 ± 0.5 c | 1.1 ± 0.1 b | 8.0 ± 0.0 bc | 5.6 ± 0.3 b | 61.1 ± 6.7 c | 2.44 ± 0.70 b | |||
| Three-way ANOVA/General Linear Model | |||||||||||
| Solid culture medium composition [ODK3 vs ODK4] (A) | 0.979 ns | 0.233 ns | 0.023 * | 0.225 ns | 0.703 ns | 0.000 *** | |||||
| Liquid culture medium composition [ODY1 vs. ODY2] (B) | 0.000 *** | 0.000 *** | 0.000 *** | 0.000 *** | 0.000 *** | 0.490 ns | |||||
| In vitro culture duration in solid medium [120, 200 days] (C) | 0.000 *** | 0.000 *** | 0.000 *** | 0.000 *** | 0.000 *** | 0.000 *** | |||||
| (A) × (B) | 0.143 ns | 0.287 ns | 0.436 ns | 0.104 ns | 0.066 ns | 0.000 *** | |||||
| (A) × (C) | 0.417 ns | 0.794 ns | 0.362 ns | 0.029 * | 0.879 ns | 0.000 *** | |||||
| (B) × (C) | 0.000 *** | 0.000 *** | 0.001 ** | 0.000 *** | 0.000 *** | 0.433 ns | |||||
| (A) × (B) × (C) | 0.003 ** | 0.028 * | 0.935 ns | 0.000 *** | 0.001 ** | 0.000 *** | |||||
| Explant Type | Solid Medium | Liquid Medium | In Vitro Culture Duration (in Days) | F.W. (g) (=Fresh Biomass Growth Rate) | D.W. (g) | F.W./D.W. Ratio | RA (% w/w) | Mean Yield Index * | Yield Index/ Explant ** | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Solid Medium | Liquid Medium | Total | |||||||||
| Root callus derived roots | ODK3 | ODY1 | 40 | 40 | 80 | 4.1 ± 0.0 d | 0.5 ± 0.0 e | 9.0 ± 0.5 bcd | 1.1 ± 0.0 i | 5.0 ± 0.4 e | 0.20 ± 0.01 e |
| 120 | 40 | 160 | 4.3 ± 0.3 d | 0.4 ± 0.0 ef | 12.1 ± 1.6 a | 2.6 ± 0.0 f | 9.3 ± 0.9 d | 0.37 ± 0.03 d | |||
| 200 | 40 | 240 | 16.2 ± 0.6 a | 1.7 ± 0.1 a | 9.8 ± 0.0 bc | 2.9 ± 0.0 d | 47.4 ± 2.2 a | 1.90 ± 0.09 a | |||
| ODK4 | ODY1 | 40 | 40 | 80 | 1.1 ± 0.0 g | 0.1 ± 0.0 g | 7.8 ± 0.6 cde | 1.1 ± 0.0 i | 1.5 ± 0.4 f | 0.06 ± 0.02 f | |
| 120 | 40 | 160 | 2.4 ± 0.3 e | 0.4 ± 0.0 ef | 6.8 ± 0.1 e | 1.6 ± 0.0 h | 5.5 ± 0.6 e | 0.22 ± 0.03 e | |||
| 200 | 40 | 240 | 11.6 ± 0.4 b | 1.1 ± 0.0 b | 10.7 ± 0.3 ab | 4.7 ± 0.0 a | 51.4 ± 1.4 a | 2.06 ± 0.01 a | |||
| ODK3 | ODY2 | 40 | 40 | 80 | 1.1 ± 0.1 g | 0.1 ± 0.0 g | 8.7 ± 0.6 bcd | 1.1 ± 0.0 i | 1.4 ± 0.4 f | 0.06 ± 0.01 g | |
| 120 | 40 | 160 | 3.0 ± 0.6 e | 0.4 ± 0.1 ef | 7.6 ± 0.1 de | 3.2 ± 0.0 c | 12.4 ± 1.8 c | 0.50 ± 0.07 c | |||
| 200 | 40 | 240 | 6.6 ± 0.3 c | 0.8 ± 0.0 c | 8.2 ± 0.3 cde | 2.7 ± 0.0 e | 21.9 ± 1.8 b | 0.88 ± 0.07 b | |||
| ODK4 | ODY2 | 40 | 40 | 80 | 1.5 ± 0.0 fg | 0.2 ± 0.0 g | 8.4 ± 0.3 cde | 0.8 ± 0.0 j | 1.5 ± 0.0 f | 0.06 ± 0.00 f | |
| 120 | 40 | 160 | 2.4 ± 0.0 ef | 0.3 ± 0.0 f | 7.2 ± 0.3 de | 2.0 ± 0.0 g | 6.6 ± 0.7 e | 0.26 ± 0.03 e | |||
| 200 | 40 | 240 | 5.8 ± 0.1 c | 0.6 ± 0.0 d | 9.5 ± 0.0 bc | 3.3 ± 0.0 b | 20.4 ± 0.2 b | 0.82 ± 0.01 b | |||
| Three-way ANOVA/General Linear Model | |||||||||||
| Solid culture medium composition [ODK3 vs. ODK4] (A) | 0.000 *** | 0.000 *** | 0.013 * | 0.608 ns | 0.003 ** | 0.003 ** | |||||
| Liquid culture medium composition [ODY1 vs. ODY2] (B) | 0.000 *** | 0.000 *** | 0.002 ** | 0.000 *** | 0.000 *** | 0.000 *** | |||||
| In vitro culture duration in solid medium [40, 120, 200 days] (C) | 0.000 *** | 0.000 *** | 0.023 * | 0.000 *** | 0.000 *** | 0.000 *** | |||||
| (A) × (B) | 0.000 *** | 0.000 *** | 0.004 ** | 0.000 *** | 0.321 ns | 0.321 ns | |||||
| (A) × (C) | 0.004 ** | 0.000 *** | 0.000 *** | 0.000 *** | 0.004 ** | 0.004 ** | |||||
| (B) × (C) | 0.000 *** | 0.000 *** | 0.022 * | 0.000 *** | 0.000 *** | 0.000 *** | |||||
| (A) × (B) × (C) | 0.015 * | 0.000 *** | 0.011 * | 0.000 *** | 0.019 * | 0.019 * | |||||
| Explant Type | Solid Medium | Liquid Medium | In Vitro Culture Duration (in Days) | F.W. (g) (=Fresh Biomass Growth Rate) | D.W. (g) | F.W./D.W. Ratio | RA (% w/w) | Mean Yield Index * | Yield Index/Explant ** | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Solid Medium | Liquid Medium | Total | |||||||||
| Leaf callus derived roots | ODK3 | ODY1 | 200 | 40 | 240 | 15.2 ± 0.0 b | 1.5 ± 0.0 b | 10.1 ± 0.0 a | 5.8 ± 0.6 ab | 86.7 ± 8.7 b | 3.47 ± 0.35 b |
| ODK4 | 200 | 40 | 240 | 19.0 ± 0.2 a | 1.8 ± 0.0 a | 10.3 ± 0.0 a | 5.1 ± 0.6 b | 93.7 ± 12.2 b | 3.75 ± 0.49 b | ||
| Petiole callus derived roots | ODK3 | ODY1 | 200 | 40 | 240 | 18.8 ± 1.0 a | 1.8 ± 0.1 ab | 10.7 ± 0.0 a | 6.6 ± 0.0 a | 115.3 ± 5.8 a | 4.61 ± 0.23 a |
| ODK4 | 200 | 40 | 240 | 16.6 ± 0.4 b | 1.6 ± 0.2 ab | 10.4 ± 1.0 a | 5.6 ± 0.4 ab | 89.9 ± 15.3 b | 3.60 ± 0.61 b | ||
| Root callus derived roots | ODK3 | ODY1 | 200 | 40 | 240 | 16.2 ± 0.6 b | 1.7 ± 0.1 ab | 9.8 ± 0.0 a | 2.9 ± 0.0 c | 47.4 ± 2.2 c | 1.90 ± 0.09 c |
| ODK4 | 200 | 40 | 240 | 11.6 ± 0.4 c | 1.1 ±0.0 c | 10.7 ± 0.3 a | 4.7 ± 0.0 b | 51.4 ± 0.4 c | 2.06 ± 0.01 c | ||
| Three-way ANOVA/General Linear Model | |||||||||||
| Explant type [Roots derived from leaf callus, petiole callus, root callus] (A) | 0.000 *** | 0.005 ** | 0.501 ns | 0.000 *** | 0.000 *** | 0.000 *** | |||||
| Solid culture medium composition [ODK3 vs. ODK4] (B) | 0.031 * | 0.081 ns | 0.350 ns | 0.792 ns | 0.495 ns | 0.495 ns | |||||
| (A) × (B) | 0.000 *** | 0.001 ** | 0.412 ns | 0.004 ** | 0.186 ns | 0.186 ns | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarropoulou, V.; Paloukopoulou, C.; Karioti, A.; Maloupa, E.; Grigoriadou, K. Rosmarinic Acid Production from Origanum dictamnus L. Root Liquid Cultures In Vitro. Plants 2023, 12, 299. https://doi.org/10.3390/plants12020299
Sarropoulou V, Paloukopoulou C, Karioti A, Maloupa E, Grigoriadou K. Rosmarinic Acid Production from Origanum dictamnus L. Root Liquid Cultures In Vitro. Plants. 2023; 12(2):299. https://doi.org/10.3390/plants12020299
Chicago/Turabian StyleSarropoulou, Virginia, Charikleia Paloukopoulou, Anastasia Karioti, Eleni Maloupa, and Katerina Grigoriadou. 2023. "Rosmarinic Acid Production from Origanum dictamnus L. Root Liquid Cultures In Vitro" Plants 12, no. 2: 299. https://doi.org/10.3390/plants12020299