
Mineral Nutrition of Naturally Growing Scots Pine and Norway Spruce under Limited Water Supply
 , ,
, , 
Abstract
:1. Introduction
- -
- the influence of water supply on the dynamics of available pools of mineral elements in soils and on their contents in plants;
- -
- the interrelations of nutrient dynamics between wood, needles, and bark under short- and long-term water shortages;
- -
- physiological effects of observed changes in element concentrations in needles.
2. Results
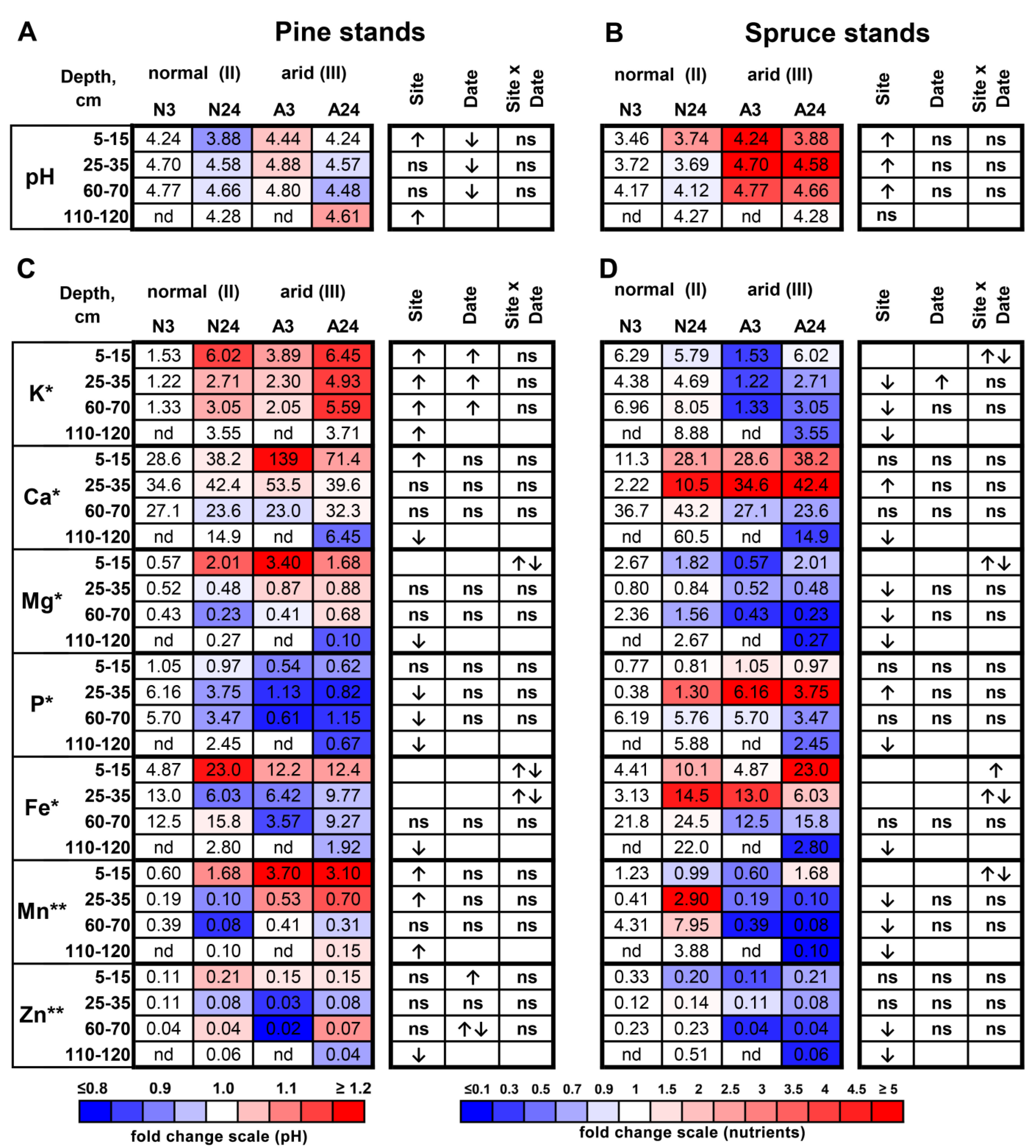
2.1. pH and Nutrient Dynamics in Soil
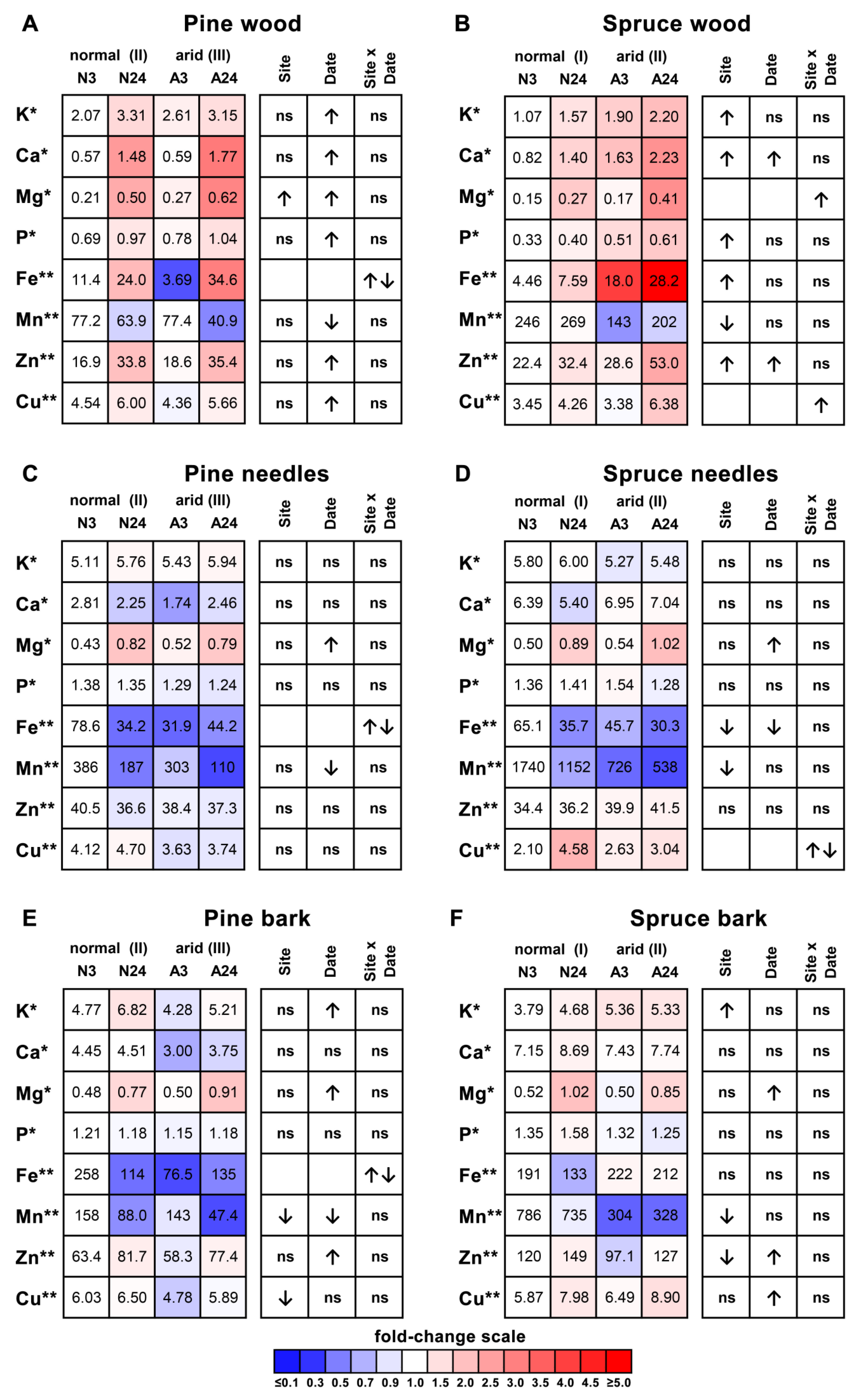
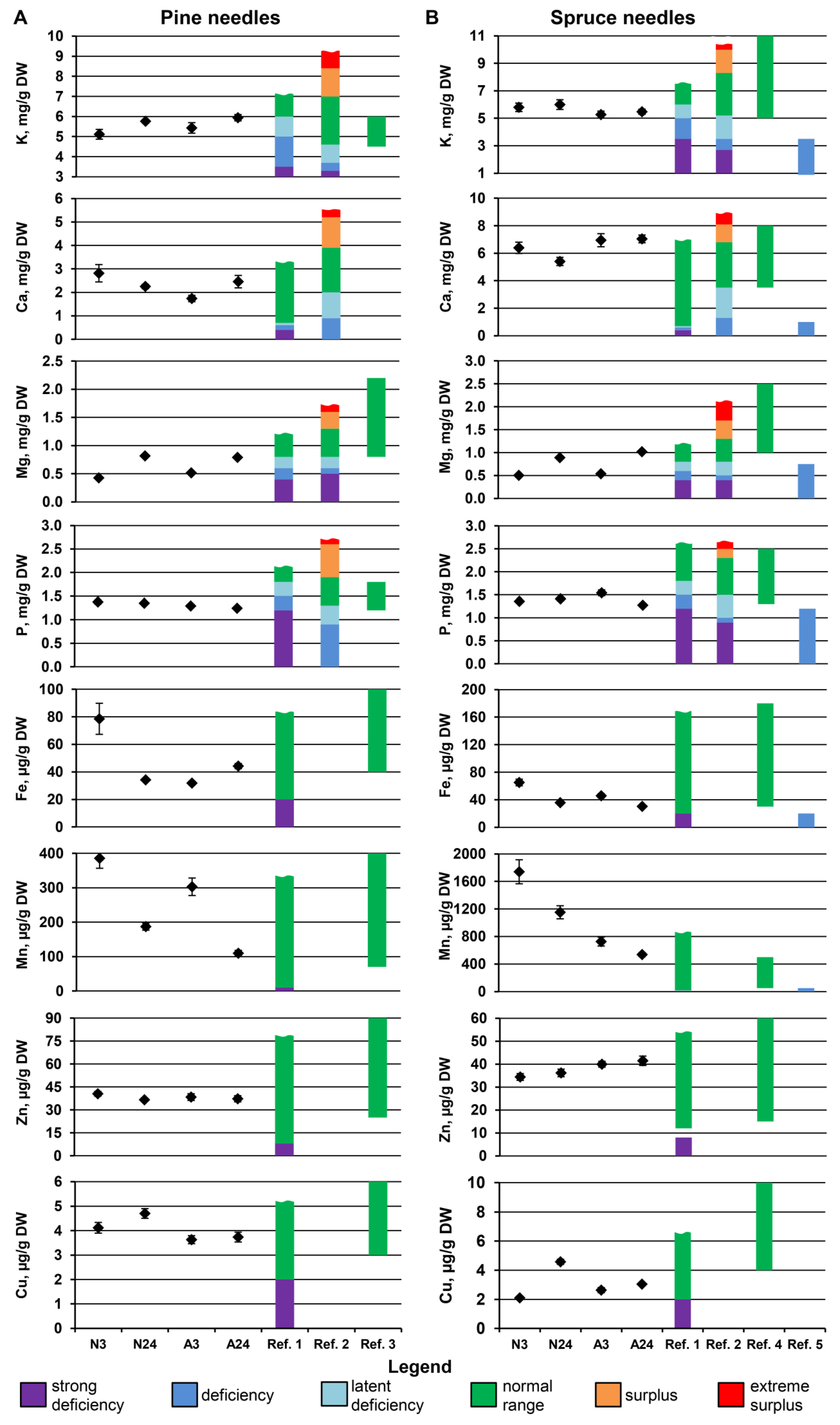
2.2. Nutrient Dynamics in Plant Organs
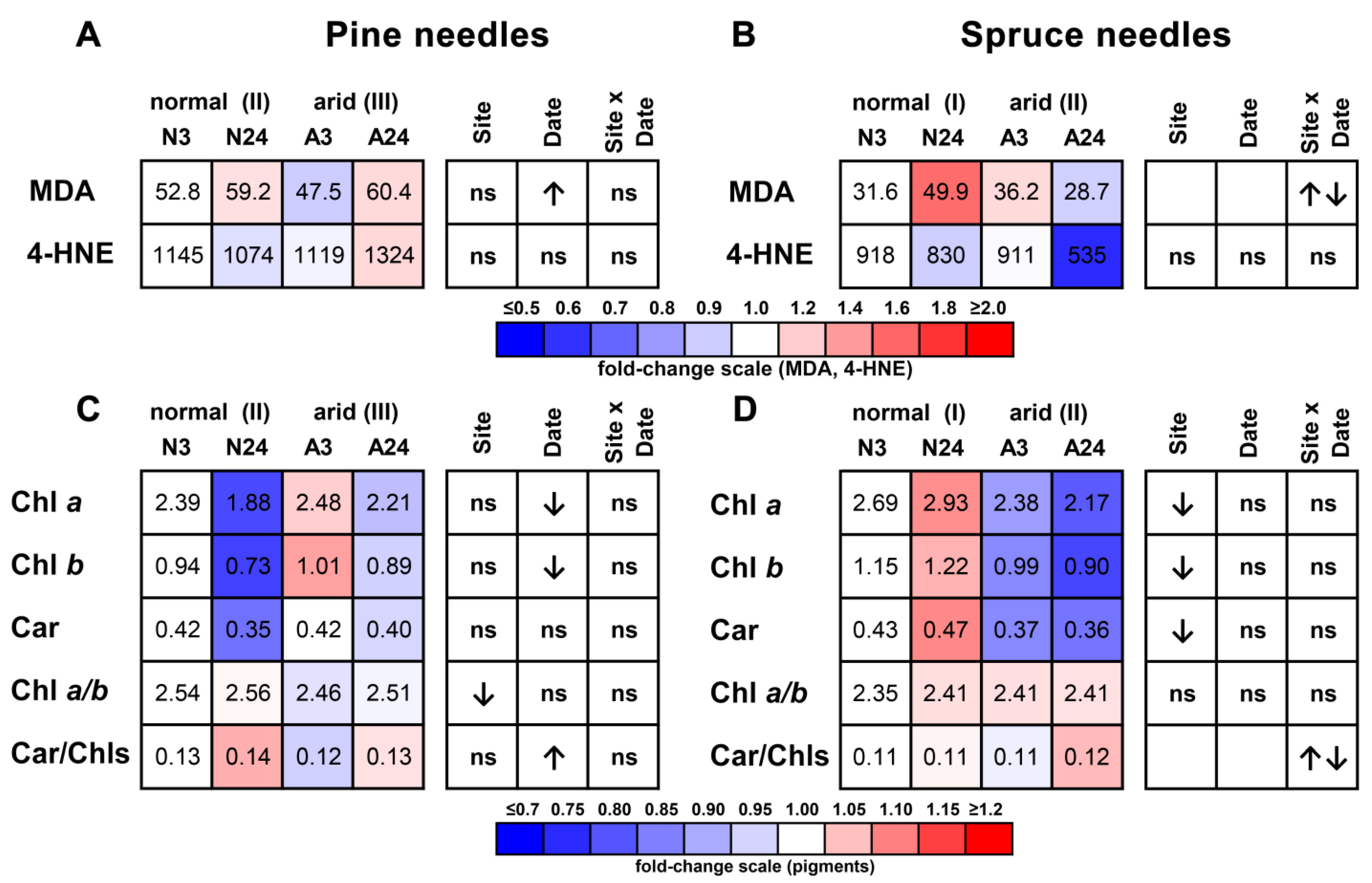
2.3. Concentrations of Lipid Peroxidation Products and Photosynthetic Pigments
3. Discussion
4. Materials and Methods
4.1. Selection of Experimental Sites
- -
- Site I (53 10.923, 34 34.334). Mean age of adult trees, 70 years; productivity class, I; relative density, 0.8; stand composition, 10 spruce; forest site type, B3; forest type, bilberry-spruce forest; and soil type, slightly soddy sandy mesopodzol with a ferruginous illuvial horizon on fluvioglacial sand. Based on the living ground cover species, this site is mesohydrophilous [36], and it was therefore considered a site with a normal water regime for spruce (designated N).
- -
- Site II (53 11.525, 34 34.760). Mean age of adult trees, 70 years; productivity class, I; relative density, 0.9; stand composition, 8.7 pine, 1.3 spruce + birch; forest site type, A2; forest type, cowberry pine forest; and soil type, medium soddy sandy mesopodzol on fluvioglacial sand, with quartz-glauconitic sand containing phosphorites underneath. Based on the living ground cover species, this site is mesophilic [36], and it was therefore considered arid (A) for spruce and normal (N) for pine.
- -
- Site III (53 13.785, 34 35.352). Mean age of adult trees, 60 years; productivity class, I; relative density, 0.5; stand composition, 10 pine; forest site type, A1; forest type, cowberry-pine forest; and soil type, slightly soddy sandy cryptopodzol on fluvioglacial sands. Based on the living ground cover species, this site is mesoxerophilous [36], and it was therefore considered arid (A) for pine.
4.2. Collection of Plant and Soil Samples
4.3. Analysis of Soil pH and Available Pools of Mineral Nutrients
4.4. Analysis of the Elemental Composition of Wood, Needles, and Bark of Current-Year Twigs
4.5. Evaluating the Level of Lipid Peroxidation in the Needles
4.6. Determination of Photosynthetic Pigments in the Needles
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- DeSoto, L.; Cailleret, M.; Sterck, F.; Jansen, S.; Kramer, K.; Robert, E.M.; Aakala, T.; Amoroso, M.M.; Bigler, C.; Camarero, J.J.; et al. Low growth resilience to drought is related to future mortality risk in trees. Nat Commun. 2020, 11, 545. [Google Scholar] [CrossRef] [Green Version]
- Gessler, A.; Schaub, M.; McDowell, N.G. The role of nutrients in drought-induced tree mortality and recovery. New Phytol. 2017, 214, 513–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlesinger, W.H.; Dietze, M.C.; Jackson, R.B.; Phillips, R.P.; Rhoades, C.C.; Rustad, L.E.; Vose, J.M. Forest biogeochemistry in response to drought. Global Change Biol. 2016, 22, 2318–2328. [Google Scholar] [CrossRef] [PubMed]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology, 3rd ed.; Springer: New York, NY, USA, 2008; pp. 11–99. [Google Scholar]
- Peuke, A.D.; Rennenberg, H. Impacts of drought on mineral macro- and microelements in provenances of beech (Fagus sylvatica L.) seedlings. Tree Physiol. 2011, 31, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Reich, P.B.; Chen, H.Y.; Xiang, Y.; Luo, Y.; Shen, Y.; Meng, C.; Han, W.; Niu, S. Global changes alter plant multi-element stoichiometric coupling. New Phytol. 2019, 221, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Querejeta, J.I.; Ren, W.; Prieto, I. Vertical decoupling of soil nutrients and water under climate warming reduces plant cumulative nutrient uptake, water-use efficiency and productivity. New Phytol. 2021, 230, 1378–1393. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Tortosa, D.; Castro, J.; Villar-Salvador, P.; Viñegla, B.; Matías, L.; Michelsen, A.; Rubio de Casas, R.; Querejeta, J.I. The “isohydric trap”: A proposed feedback between water shortage, stomatal regulation, and nutrient acquisition drives differential growth and survival of European pines under climatic dryness. Global Change Biol. 2018, 24, 4069–4083. [Google Scholar] [CrossRef] [PubMed]
- González de Andrés, E.; Suárez, M.L.; Querejeta, J.I.; Camarero, J.J. Chronically low nutrient concentrations in tree rings are linked to greater tree vulnerability to drought in Nothofagus dombeyi. Forests. 2021, 12, 1180. [Google Scholar] [CrossRef]
- González de Andrés, E.; Gazol, A.; Querejeta, J.I.; Igual, J.M.; Colangelo, M.; Sánchez-Salguero, R.; Linares, J.C.; Camarero, J.J. The role of nutritional impairment in carbon-water balance of silver fir drought-induced dieback. Glob. Change Biol. 2022, 28, 4439–4458. [Google Scholar] [CrossRef] [PubMed]
- León-Sánchez, L.; Nicolas, E.; Prieto, I.; Nortes, P.; Maestre, F.T.; Querejeta, J.I. Altered leaf elemental composition with climate change is linked to reductions in photosynthesis, growth and survival in a semi-arid shrubland. J. Ecol. 2020, 108, 47–60. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Jiménez, J.J.; Moret-Fernández, D.; López, M.V.; Sangüesa-Barreda, G.; Igual, J.M. Beneath the canopy: Linking drought-induced forest die off and changes in soil properties. Forest Ecol Manag. 2018, 422, 294–302. [Google Scholar] [CrossRef]
- Hevia, A.; Sánchez-Salguero, R.; Camarero, J.J.; Querejeta, J.I.; Sangüesa-Barreda, G.; Gazol, A. Long-term nutrient imbalances linked to drought-triggered forest dieback. Sci. Total Environ. 2019, 690, 1254–1267. [Google Scholar] [CrossRef]
- Buras, A.; Hirsch, F.; Schneider, A.; Scharnweber, T.; van der Maaten, E.; Cruz-García, R.; Raab, T.; Wilmking, M. Reduced above-ground growth and wood density but increased wood chemical concentrations of Scots pine on relict charcoal hearths. Sci. Total Environ. 2020, 717, 137189. [Google Scholar] [CrossRef] [PubMed]
- Hevia, A.; Sánchez-Salguero, R.; Camarero, J.J.; Buras, A.; Sangüesa-Barreda, G.; Galván, J.D.; Gutiérrez, E. Towards a better understanding of long-term wood-chemistry variations in old-growth forests: A case study on ancient Pinus uncinata trees from the Pyrenees. Sci. Total Environ. 2018, 625, 220–232. [Google Scholar] [CrossRef]
- Liu, Y.; Ta, W.; Cherubini, P.; Liu, R.; Wang, Y.; Sun, C. Elements content in tree rings from Xi’an, China and environmental variations in the past 30 years. Sci. Total Environ. 2018, 619, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Kreuzwieser, J.; Gessler, A. Global climate change and tree nutrition: Influence of water availability. Tree Physiol. 2010, 30, 1221–1234. [Google Scholar] [CrossRef] [PubMed]
- Pashkovskiy, P.P.; Vankova, R.; Zlobin, I.E.; Dobrev, P.; Kartashov, A.V.; Ivanova, A.I.; Ivanov, V.P.; Marchenko, S.I.; Nartov, D.I.; Ivanov, Y.V.; et al. Hormonal responses to short-term and long-term water deficit in native Scots pine and Norway spruce trees. Environ. Exp. Bot. 2022, 195, 104789. [Google Scholar] [CrossRef]
- Han, W.X.; Fang, J.Y.; Reich, P.B.; Woodward, F.I.; Wang, Z.H. Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China. Ecol. Lett. 2011, 14, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants., 2nd ed.; London Academic Press: London, UK, 2011. [Google Scholar]
- Ivanov, Y.V.; Kartashov, A.V.; Ivanova, A.I.; Ivanov, V.P.; Marchenko, S.I.; Nartov, D.I.; Kuznetsov, V.V. Long-term impact of cement plant emissions on the elemental composition of both soils and pine stands and on the formation of Scots pine seeds. Environ. Pollut. 2018, 243, 1383–1393. [Google Scholar] [CrossRef]
- Blanco, J.A.; Imbert, J.B.; Castillo, F.J. Thinning affects nutrient resorption and nutrient-use efficiency in two Pinus sylvestris stands in the Pyrenees. Ecol Appl. 2009, 19, 682–698. [Google Scholar] [CrossRef]
- Juranović Cindrić, I.; Zeiner, M.; Starčević, A.; Stingeder, G. Metals in pine needles: Characterisation of bio-indicators depending on species. Int. J. Environ. Sci. Technol. 2019, 16, 4339–4346. [Google Scholar] [CrossRef] [Green Version]
- Kavvadias, V.A.; Miller, H.G. Manganese and calcium nutrition of Pinus sylvestris and Pinus nigra from two different origins. I. Manganese. Forestry. 1999, 72, 35–46. [Google Scholar] [CrossRef]
- Schleppi, P.; Tobler, L.; Bucher, J.B.; Wyttenbach, A. Multivariate interpretation of the foliar chemical composition of Norway spruce (Picea abies). Plant and Soil. 2000, 219, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Dučić, T.; Leinemann, L.; Finkeldey, R.; Polle, A. Uptake and translocation of manganese in seedlings of two varieties of Douglas fir (Pseudotsuga menziesii var. viridis and glauca). New Phytol. 2006, 170, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Brække, F.H.; Salih, N. Reliability of foliar analyses of Norway spruce stands in a Nordic gradient. Silva Fenn. 2002, 36, 489–504. [Google Scholar] [CrossRef] [Green Version]
- Mellert, K.H.; Göttlein, A. Comparison of new foliar nutrient thresholds derived from van den Burg’s literature compilation with established central European references. Eur. J. Forest Res. 2012, 131, 1461–1472. [Google Scholar] [CrossRef]
- Oleksyn, J.; Reich, P.B.; Zytkowiak, R.; Karolewski, P.; Tjoelker, M.G. Needle nutrients in geographically diverse Pinus sylvestris L. populations. Ann. Forest Sci. 2002, 59, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kulhavý, J.; Marková, I.; Drápelová, I.; Truparová, S. The effect of liming on the mineral nutrition of the mountain Norway spruce (Picea abies L.) forest. J. For. Sci. 2009, 55, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ewald, J. Ecological background of crown condition, growth and nutritional status of Picea abies (L.) Karst. in the Bavarian Alps. Eur J Forest Res. 2005, 124, 9–18. [Google Scholar] [CrossRef]
- Eroglu, S.; Meier, B.; von Wirén, N.; Peiter, E. The vacuolar manganese transporter MTP8 determines tolerance to iron deficiency-induced chlorosis in Arabidopsis. Plant Physiol. 2016, 170, 1030–1045. [Google Scholar] [CrossRef] [PubMed]
- Lanquar, V.; Ramos, M.S.; Lelièvre, F.; Barbier-Brygoo, H.; Krieger-Liszkay, A.; Krämer, U.; Thomine, S. Export of vacuolar manganese by AtNRAMP3 and AtNRAMP4 is required for optimal photosynthesis and growth under manganese deficiency. Plant Physiol. 2010, 152, 1986–1999. [Google Scholar] [CrossRef] [Green Version]
- Lynch, J.P.; Clair, S.B.S. Mineral stress: The missing link in understanding how global climate change will affect plants in real world soils. Field Crop Res. 2004, 90, 101–115. [Google Scholar] [CrossRef]
- Andresen, E.; Peiter, E.; Küpper, H. Trace metal metabolism in plants. J Exp Bot. 2018, 69, 909–954. [Google Scholar] [CrossRef] [Green Version]
- Fomin, V.V.; Zalesov, S.V.; Popov, A.S.; Mikhailovich, A.P. Historical avenues of research in Russian forest typology: Ecological, phytocoenotic, genetic, and dynamic classifications. Can. J. Forest Res. 2017, 47, 849–860. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, S.M.; Cruz, N.; Coelho, C.; Henriques, B.; Carvalho, L.; Duarte, A.C.; Pereira, E.; Römkens, P.F. Risk assessment for Cd, Cu, Pb and Zn in urban soils: Chemical availability as the central concept. Environ Pollut. 2013, 183, 234–242. [Google Scholar] [CrossRef] [PubMed]
- ISO 10390:2005; Soil Quality, Determination of pH. International Standards Organization: Geneve, Switzerland, 2005.
- Mamo, T.; Richter, C.; Heiligtag, B. Comparison of extractants for the determination of available phosphorus, potassium, calcium, magnesium and sodium in some Ethiopian and German soils. Commun. Soil Sci. Plant. 1996, 27, 2197–2212. [Google Scholar] [CrossRef]
- Nagul, E.A.; McKelvie, I.D.; Worsfold, P.; Kolev, S.D. The molybdenum blue reaction for the determination of orthophosphate revisited: Opening the black box. Anal. Chim. Acta. 2015, 890, 60–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gérard-Monnier, D.; Erdelmeier, I.; Régnard, K.; Moze-Henry, N.; Yadan, J.C.; Chaudiere, J. Reactions of 1-methyl-2-phenylindole with malondialdehyde and 4-hydroxyalkenals. Analytical applications to a colorimetric assay of lipid peroxidation. Chem. Res. Toxicol. 1998, 11, 1176–1183. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. [34] Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology; Packer, L., Douce, R., Eds.; Academic Press: Cambridge, MA, USA, 1987; Volume 148, pp. 350–382. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ca | Mg | P | Fe | Mn | Zn | Cu | Ca | Mg | P | Fe | Mn | Zn | Cu | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pine wood | Spruce wood | |||||||||||||
| K | 0.36 * | 0.63 *** | 0.89 *** | 0.30 | 0.05 | 0.66 *** | 0.40 * | 0.68 *** | 0.66 *** | 0.88 *** | 0.71 *** | 0.04 | 0.53 ** | 0.53 ** |
| Ca | 0.69 *** | 0.48 ** | 0.60 *** | −0.46 ** | 0.53 ** | 0.34 | 0.72 *** | 0.81 *** | 0.76 *** | 0.07 | 0.90 *** | 0.47 ** | ||
| Mg | 0.71 *** | 0.72 *** | −0.53 ** | 0.82 *** | 0.61 *** | 0.73 *** | 0.63 *** | 0.24 | 0.75 *** | 0.81 *** | ||||
| P | 0.29 | −0.07 | 0.65 *** | 0.45 ** | 0.73 *** | 0.07 | 0.68 *** | 0.58 *** | ||||||
| Fe | −0.54 ** | 0.69 *** | 0.43 * | 0.01 | 0.73 *** | 0.60 *** | ||||||||
| Mn | −0.16 | −0.19 | 0.21 | 0.21 | ||||||||||
| Zn | 0.47 ** | 0.52 ** | ||||||||||||
| Pine needles | Spruce needles | |||||||||||||
| K | −0.50 ** | 0.27 | 0.48 ** | −0.53 ** | −0.43 * | −0.38 * | 0.50 ** | −0.39 * | 0.14 | 0.42 * | −0.26 | 0.11 | 0.28 | 0.03 |
| Ca | −0.02 | −0.41 * | 0.80 *** | 0.45 ** | 0.55 *** | −0.40 ** | 0.07 | −0.42 * | 0.26 | 0.17 | 0.36 * | −0.38 * | ||
| Mg | −0.05 | −0.31 | −0.51 ** | 0.04 | 0.19 | −0.08 | −0.60 *** | −0.18 | 0.33 | 0.54 ** | ||||
| P | −0.27 | −0.11 | −0.28 | 0.73 *** | −0.20 | 0.00 | 0.10 | 0.30 | ||||||
| Fe | 0.66 *** | 0.40 * | −0.26 | 0.48 ** | −0.18 | −0.44 * | ||||||||
| Mn | 0.55 ** | −0.25 | 0.05 | −0.20 | ||||||||||
| Zn | −0.22 | −0.12 | ||||||||||||
| Pine bark | Spruce bark | |||||||||||||
| K | 0.05 | 0.39 * | 0.29 | −0.31 | −0.31 | 0.37 * | 0.48 ** | −0.11 | 0.19 | 0.34 | −0.32 | −0.13 | −0.26 | 0.26 |
| Ca | 0.30 | −0.07 | 0.67 *** | 0.09 | 0.13 | 0.21 | 0.27 | 0.20 | −0.12 | 0.17 | 0.59 *** | −0.03 | ||
| Mg | 0.15 | 0.02 | −0.59 *** | 0.67 *** | 0.48 ** | 0.44 * | −0.50 ** | 0.21 | 0.35 * | 0.59 *** | ||||
| P | −0.23 | −0.11 | 0.24 | 0.22 | −0.69 *** | 0.34 | 0.12 | 0.11 | ||||||
| Fe | 0.23 | −0.00 | 0.17 | −0.30 | −0.14 | −0.36 * | ||||||||
| Mn | −0.16 | −0.15 | 0.29 | −0.08 | ||||||||||
| Zn | 0.49 ** | 0.02 | ||||||||||||
| Nutrients | Scots Pine | Norway Spruce | ||||
|---|---|---|---|---|---|---|
| Wood–Needles | Wood–Bark | Needles–Bark | Wood–Needles | Wood–bark | Needles–Bark | |
| K | 0.50 ** | 0.44 * | 0.58 *** | 0.34 | 0.61 *** | 0.48 ** |
| Ca | 0.31 | 0.30 | 0.81 *** | 0.06 | −0.20 | 0.08 |
| Mg | 0.78 *** | 0.73 *** | 0.76 *** | 0.68 *** | 0.53 ** | 0.60 *** |
| P | 0.29 | 0.17 | 0.33 | 0.18 | −0.25 | 0.22 |
| Fe | 0.06 | 0.17 | 0.87 *** | −0.48 ** | 0.40 * | 0.17 |
| Mn | 0.59 *** | 0.71 *** | 0.82 *** | 0.62 *** | 0.69 *** | 0.86 *** |
| Zn | −0.11 | 0.66 *** | 0.15 | 0.24 | 0.24 | 0.04 |
| Cu | 0.49 ** | 0.71 *** | 0.49 ** | 0.15 | 0.62 *** | 0.40 * |
| Elements | Pine Needles | Spruce Needles | ||
|---|---|---|---|---|
| MDA | 4-HNE | MDA | 4-HNE | |
| Mn | −0.47 ** | −0.10 | 0.09 | 0.05 |
| Fe | 0.02 | −0.04 | −0.23 | 0.10 |
| Elements | Chl a | Chl b | Car | Chl a/b | Car/Chls |
|---|---|---|---|---|---|
| Pine needles | |||||
| Mg | −0.25 | −0.21 | −0.14 | −0.08 | 0.36 * |
| Fe | 0.29 | 0.23 | 0.34 | 0.26 | 0.01 |
| Spruce needles | |||||
| Mg | −0.05 | −0.06 | 0.05 | 0.15 | 0.56 *** |
| Fe | 0.17 | 0.20 | 0.11 | −0.29 | −0.44 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivanov, Y.V.; Zlobin, I.E.; Kartashov, A.V.; Ivanova, A.I.; Ivanov, V.P.; Marchenko, S.I.; Nartov, D.I.; Kuznetsov, V.V. Mineral Nutrition of Naturally Growing Scots Pine and Norway Spruce under Limited Water Supply. Plants 2022, 11, 2652. https://doi.org/10.3390/plants11192652
Ivanov YV, Zlobin IE, Kartashov AV, Ivanova AI, Ivanov VP, Marchenko SI, Nartov DI, Kuznetsov VV. Mineral Nutrition of Naturally Growing Scots Pine and Norway Spruce under Limited Water Supply. Plants. 2022; 11(19):2652. https://doi.org/10.3390/plants11192652
Chicago/Turabian StyleIvanov, Yury V., Ilya E. Zlobin, Alexander V. Kartashov, Alexandra I. Ivanova, Valery P. Ivanov, Sergey I. Marchenko, Dmitry I. Nartov, and Vladimir V. Kuznetsov. 2022. "Mineral Nutrition of Naturally Growing Scots Pine and Norway Spruce under Limited Water Supply" Plants 11, no. 19: 2652. https://doi.org/10.3390/plants11192652