
Ground Vegetation in Pinus sylvestris Forests at Different Successional Stages following Clear Cuttings: A Case Study

Abstract
:1. Introduction
2. Results
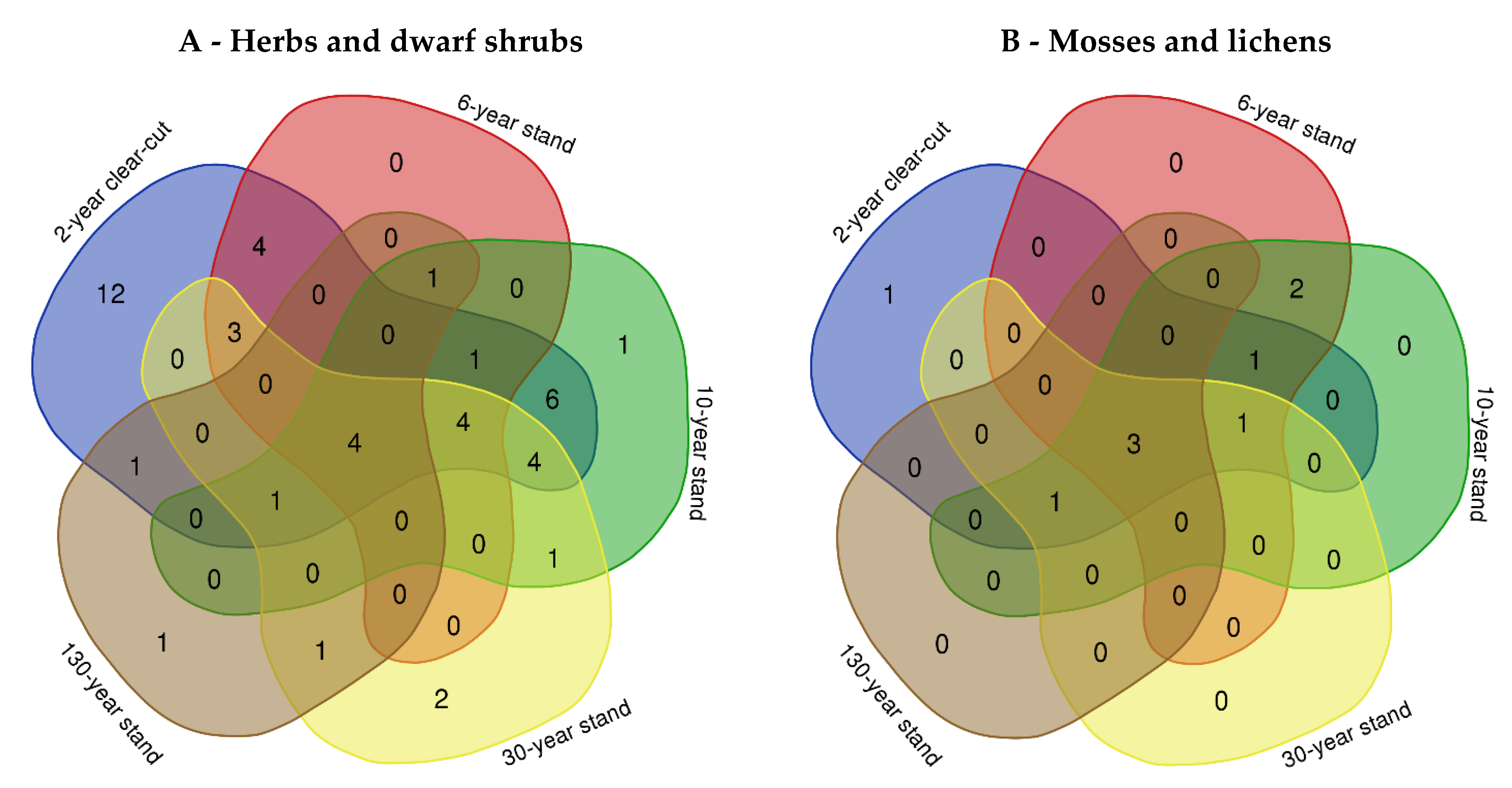
2.1. Ground Vegetation Species Composition at Different Forest Succession Stages
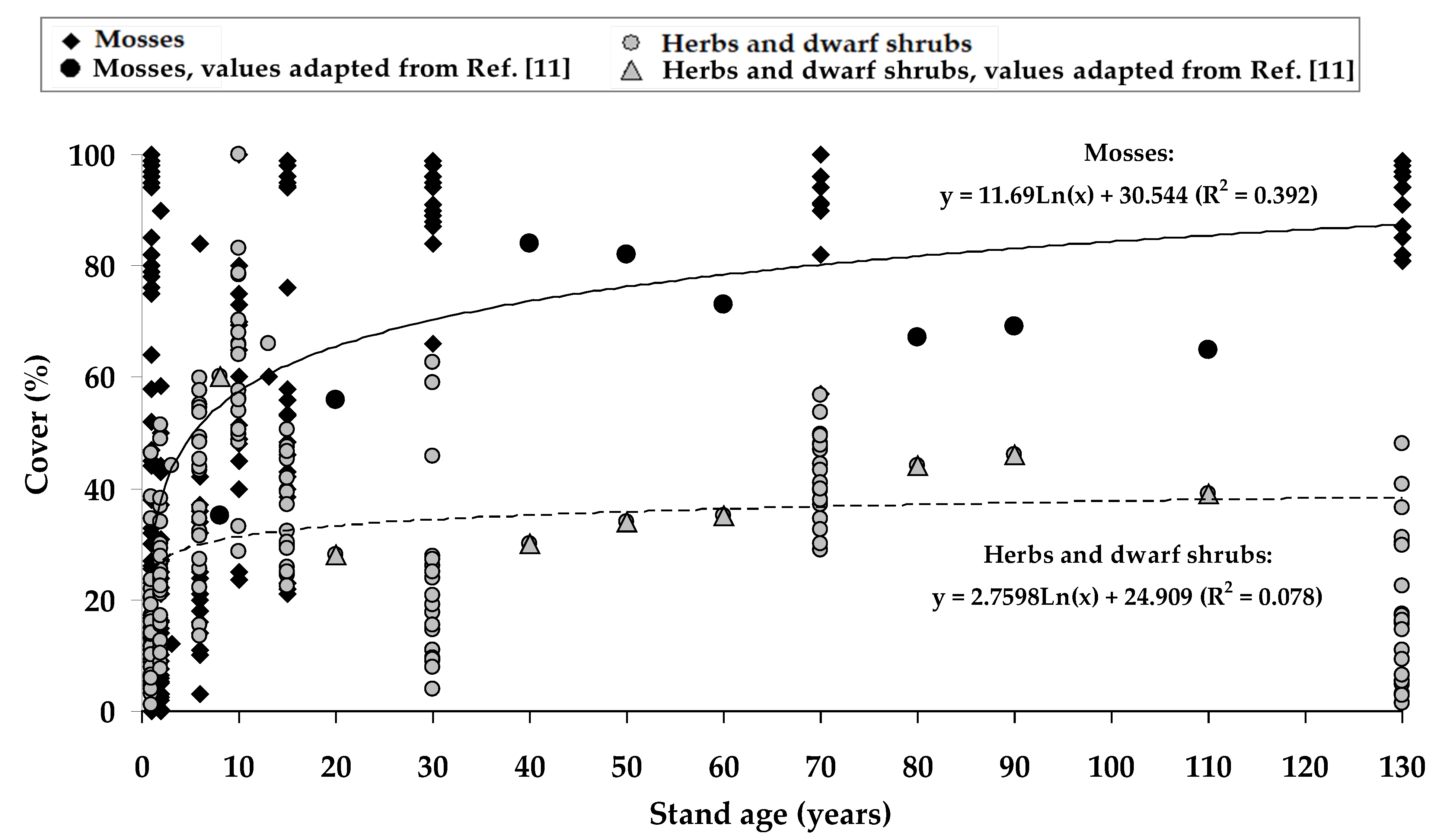
2.2. Ground Vegetation Cover at Different Forest Succession Stages
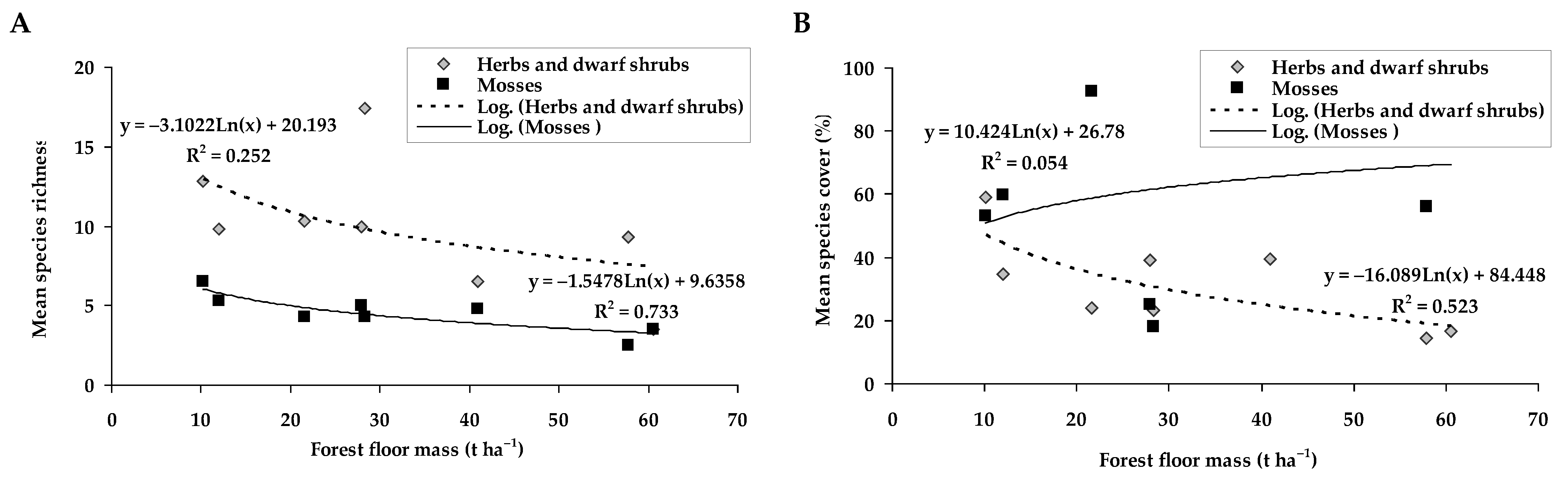
2.3. The Ground Vegetation Relations with the Forest Floor and Mineral Soil Parameters
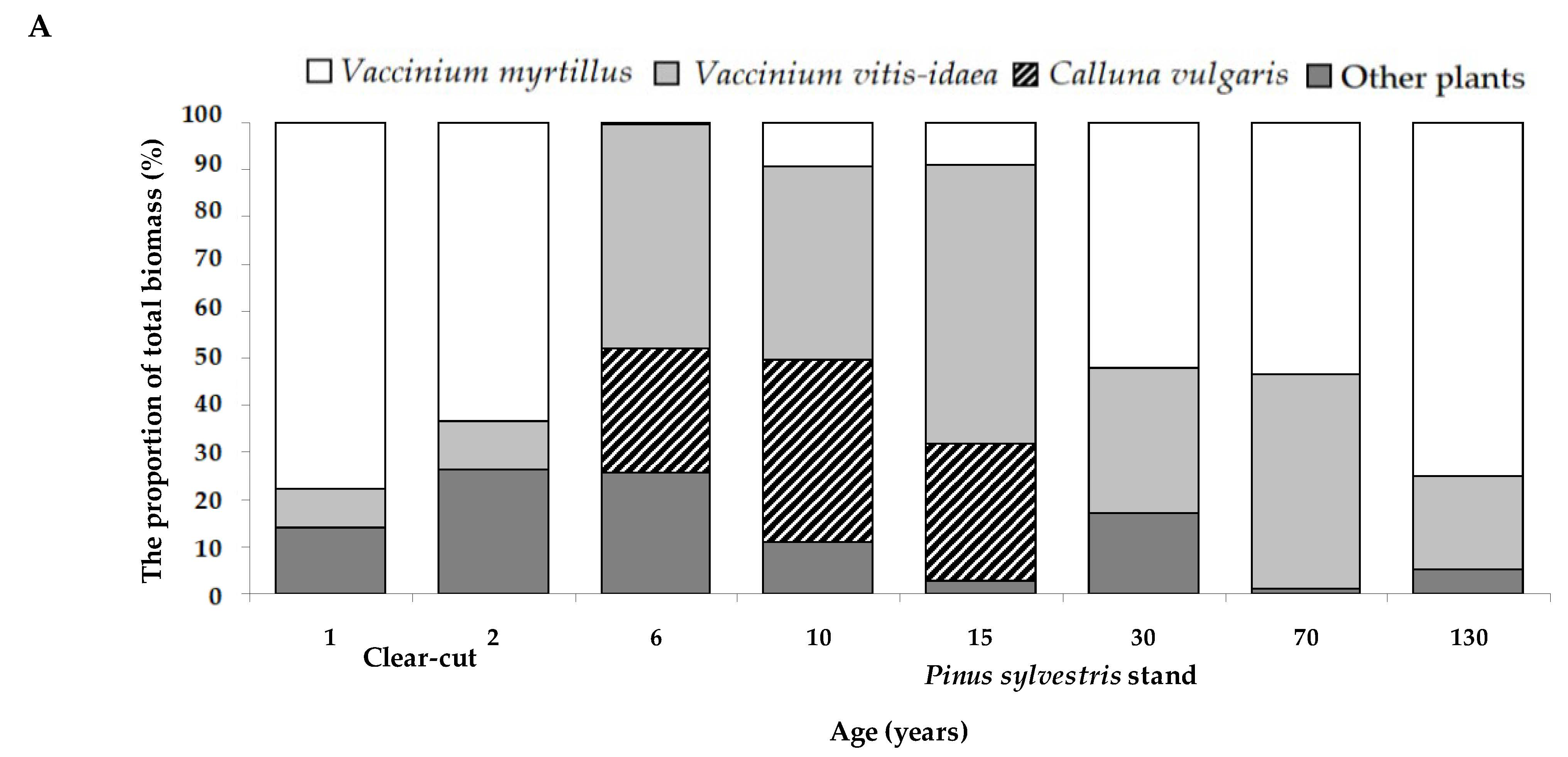
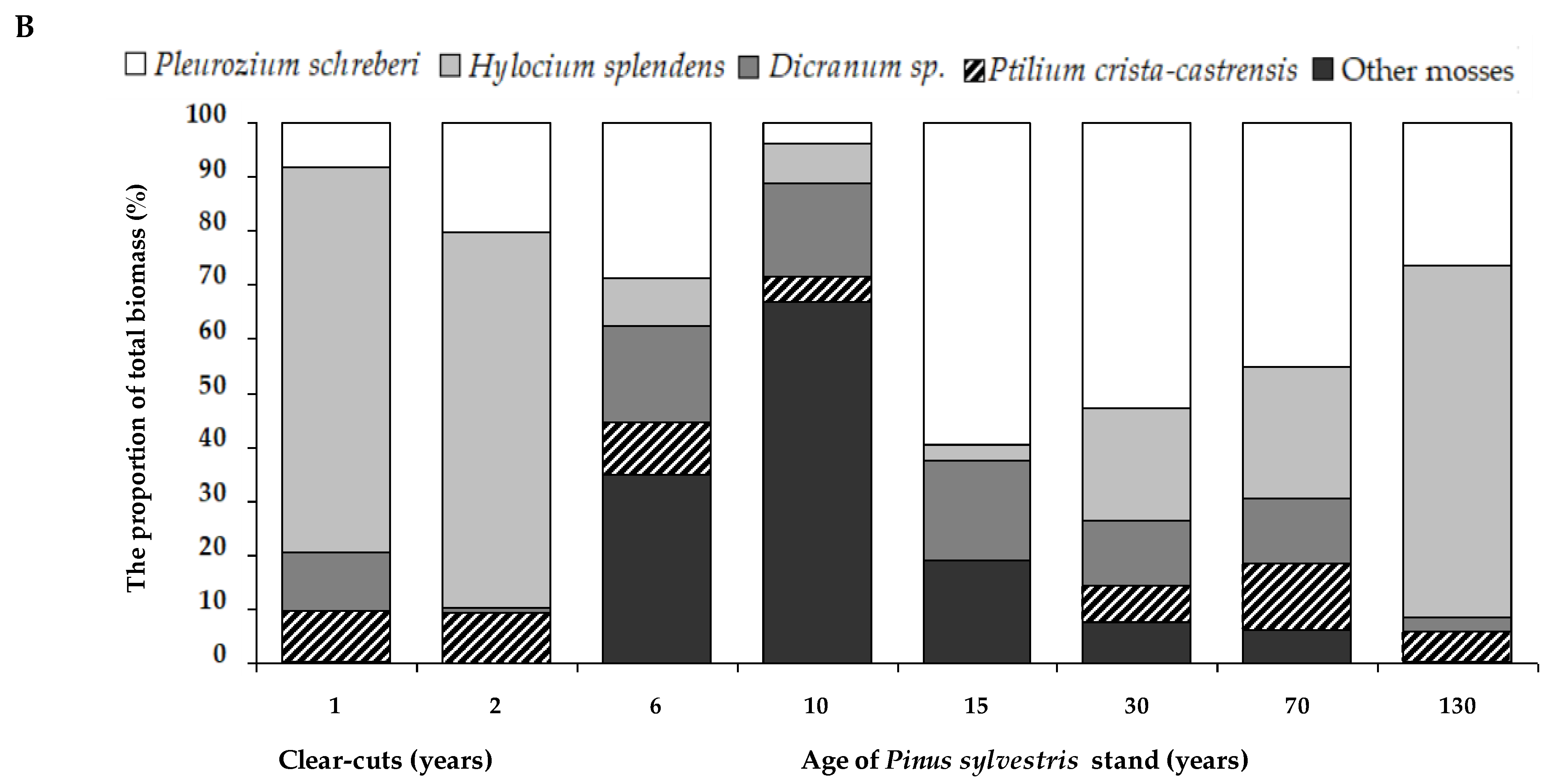
2.4. The Ground Vegetation Biomass
3. Discussion
4. Materials and Methods
4.1. Study Sites
4.2. Ground Vegetation Sampling and Analyses
4.3. Soil Sampling and Analyses
4.4. Calculations and Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dieler, J.; Uhl, E.; Biber, P.; Müller, J.; Rötzer, T.; Pretzsch, H. Effect of forest stand management on species composition, structural diversity, and productivity in the temperate zone of Europe. Eur. J. For. Res. 2017, 136, 739–766. [Google Scholar] [CrossRef]
- Forestry and Wood Products in Lithuania; GAIN Report; Global Agricultural International Network: Warsaw, Poland, 2017; p. 12.
- State Forest Service. Forest Statistic 2021. Available online: https://amvmt.lrv.lt/lt/atviri-duomenys-1/misku-statistikos-leidiniai/misku-ukio-statistika/2021-m-1 (accessed on 2 September 2022).
- Kuuluvainen, T.; Gauthier, S. Young and old forest in the boreal: Critical stages of ecosystem dynamics and management under global change. For. Ecosyst. 2018, 5, 26. [Google Scholar] [CrossRef]
- Decocq, G.; Aubert, M.; Dupont, F.; Alard, D.; Saguez, R.; Wattez-Franger, A.; de Foucault, B.; Delelis-Dusollier, A.; Bardat, J. Plant diversity in a managed temperate deciduous forest: Understorey response to two silvicultural systems. J. Appl. Ecol. 2004, 41, 1065–1079. [Google Scholar] [CrossRef]
- Miškų kirtimo taisyklės (Regulations of Forest Cutting); Nr. D1–79; Ministry of Environment of the Republic of Lithuania: Vilnius, Lithuania, 2010; updated in 2020.
- Keenan, R.J.; Kimmins, J.P. The ecological effects of clear-cutting. Environ. Rev. 1993, 1, 121–144. [Google Scholar] [CrossRef]
- Bradshaw, C.J.A.; Warkentin, I.G.; Sodhi, N.S. Urgent preservation of boreal carbon stocks and biodiversity. Trends Ecol. Evol. 2009, 24, 541–548. [Google Scholar] [CrossRef]
- McCune, B.; Antos, J.A. Correlations between forest layers in the Swan Valley, Montana. Ecology 1981, 62, 1196–1204. [Google Scholar] [CrossRef]
- Stefańska-Krzaczek, E.; Staniaszek-Kik, M.; Fałtynowicz, W. Positive Aspects of Clear-Cut Logging? Ground Bryophyte Diversity Along the Age Gradient of Managed Pinus sylvestris Stands. Cryptog. Bryol. 2016, 37, 181–197. [Google Scholar] [CrossRef]
- Karazija, S. Age-related dynamics of pine forest communities in Lithuania. Balt. For. 2002, 9, 50–62. [Google Scholar]
- Widenfalk, O.; Weslien, J. Forest ecology and management plant species richness in managed boreal forests—Effects of stand succession and thinning. For. Ecol. Manag. 2009, 257, 1386–1394. [Google Scholar] [CrossRef]
- Stefańska-Krzaczek, E.; Szymura, T.H. Species diversity of forest floor vegetation in age gradient of managed Scots pine stands. Balt. For. 2015, 21, 233–243. [Google Scholar]
- Corns, I.G.W. Forest site classification in Alberta: Its evolution and present status. For. Chron. 1992, 68, 85–93. [Google Scholar] [CrossRef]
- Busby, J.R.; Bliss, L.C.; Hamilton, C.D. Microclimatic control of growth rates and habitats of the boreal forest mosses, Tomenthypnum nitens and Hylocomium splendens. Ecol. Monogr. 1978, 48, 95–110. [Google Scholar] [CrossRef]
- Légaré, S.; Bergeron, Y.; Paré, D. Influence of forest composition on understory cover in boreal mixedwood forests of western Quebec. Silva Fenn. 2002, 36, 353–366. [Google Scholar] [CrossRef] [Green Version]
- Strengbom, J.; Nasholm, T.; Ericson, L. Light, not nitrogen, limits growth of the grass Deschampsia flexuosa in boreal forests. Can. J. Bot. 2004, 82, 430–435. [Google Scholar] [CrossRef]
- Hart, S.A.; Chen, H.Y.H. Understory Vegetation Dynamics of North American Boreal Forests. CRC Crit. Rev. Plant Sci. 2006, 25, 381–397. [Google Scholar] [CrossRef]
- Bainbridge, E.L.; Strong, W.L. Pinus contorta understory vegetation dynamics following clearcutting in west-central Alberta, Canada. For. Ecol. Manag. 2005, 213, 133–150. [Google Scholar] [CrossRef]
- Sheil, D.; Jennings, S.; Savill, P. Long-term permanent plot observations of vegetation dynamics in Budongo, a Ugandan rainforest. J. Trop. Ecol. 2000, 16, 765–800. [Google Scholar] [CrossRef] [Green Version]
- Whittaker, R.; Bush, M.; Richards, K. Plant recolonization and vegetation succession on the Krakatau islands, Indonesia. Ecol. Monogr. 1989, 59, 59–123. [Google Scholar] [CrossRef]
- Lichter, J. Primary succession and forest development on coastal Lake Michigan sand dunes. Ecol. Monogr. 1998, 68, 487–510. [Google Scholar] [CrossRef]
- Anderson, K.J. Temporal patterns in rates of community change during succession. Am Nat. 2007, 169, 780–793. [Google Scholar] [CrossRef]
- Mátyás, C.; Ackzell, L.; Samuel, C.J.A. EUFORGEN Technical Guidelines for Genetic Conservation and Use for Scots Pine (Pinus sylvestris); International Plant Genetic Resources Institute: Rome, Italy, 2004; p. 6. [Google Scholar]
- Petrokas, R.; Baliuckas, V.; Manton, M. Successional Categorization of European Hemi-boreal Forest Tree Species. Plants 2020, 9, 1381. [Google Scholar] [CrossRef]
- Česonienė, L.; Daubaras, R.; Kaškonas, P.; Kaškonienė, V.; Maruška, A.S.; Tiso, N.; Zych, M. Initial impact of clear-cut logging on dynamics of understory vascular plants and pollinators in Scots pine-dominated forests in Lithuania. Turk. J. Agric. For 2018, 42, 6. [Google Scholar] [CrossRef]
- Česonienė, L.; Daubaras, R.; Tamutis, V.; Kaškonienė, V.; Kaškonas, P.; Stakėnas, V.; Zych, M. Effect of clear-cutting on the understory vegetation, soil and diversity of litter beetles in Scots pine-dominated forest. J. Sustain. For. 2019, 38, 791–808. [Google Scholar] [CrossRef]
- Karazija, S. Lietuvos Miškų Tipai. (Forest Types of Lithuania); Mokslas: Vilnius, Lithuania, 1988; p. 210. (In Lithuanian) [Google Scholar]
- Navasaitis, M.; Ozolinčius, R.; Smaliukas, D.; Balevičienė, J. Lietuvos Dendroflora (Dendroflora of Lithuania); Lututė: Kaunas, Lithuania, 2003; p. 576. (In Lithuanian) [Google Scholar]
- Chávez, V.; Macdonald, S.E. The influence of canopy patch mosaics on understory plant community composition in boreal mixedwood forest. For. Ecol. Manag. 2010, 259, 1067–1075. [Google Scholar] [CrossRef]
- Kumar, P.; Chen, H.Y.H.; Thomas, S.C.; Shahi, C. Linking resource availability and heterogeneity to understorey species diversity through succession in boreal forests of Canada. J. Ecol. 2018, 106, 1266–1276. [Google Scholar] [CrossRef]
- Bråkenhielm, S.; Liu, Q. Long-term effects of clear-felling on vegetation dynamics and species diversity in a boreal pine forest. Biodivers. Conserv. 1998, 7, 207–220. [Google Scholar] [CrossRef]
- Nygaard, P.H.; Odegaard, T. Sixty years of vegetation dynamics in a south boreal coniferous forest in southern Norway. J. Veg. Sci. 1999, 10, 5–16. [Google Scholar] [CrossRef]
- Pykälä, J. Immediate increase in plant species richness after clear-cutting of boreal herb-rich forests. Appl. Veg. Sci. 2004, 7, 29–34. [Google Scholar] [CrossRef]
- Bradbury, S. Understorey plant communities in boreal cutblocks with different sizes and numbers of residual tree patches. Can. J. For. Res. 2004, 34, 1220–1227. [Google Scholar] [CrossRef]
- Tellier, R.; Duchesne, L.C.; Ruel, J.C.; Mcalpine, R.S. Effects of Directed Burning Intensity on Diversity of Plant-Species in A Jack Pine Stand (Pinus-Banksiana Lamb). Ecoscience 1995, 2, 159–167. [Google Scholar] [CrossRef]
- During, H.J.; Tooren, B.F.V. Bryophyte interactions with other plants. Bot. J. Lin. 1990, 104, 79–98. [Google Scholar] [CrossRef]
- Soudzilovskaia, N.A.; Graae, B.J.; Douma, J.C.; Grau, O.; Milbau, A.; Shevtsova, A.; Wolters, L.; Cornelissen, J.H.C. How do bryophytes govern generative recruitment of vascular plants? New Phytol. 2011, 190, 1019–1031. [Google Scholar] [CrossRef] [PubMed]
- Saetre, P. Spatial patterns of ground vegetation, soil microbial biomass and activity in a mixed spruce-birch stand. Ecography 1999, 22, 183–192. [Google Scholar] [CrossRef]
- Grime, J.P.; Rincon, E.R.; Wickerson, B.E. Bryophytes and plant strategy theory. Bot. J. Linn. Soc. 1990, 104, 175–186. [Google Scholar] [CrossRef]
- Hajek, M.; Rolecek, J.; Cottenie, K.; Kintrova, K.; Horsak, M.; Poulickova, A.; Hajkova, P.; Frankova, M.; Dite, D. Environmental and spatial controls of biotic assemblages in a discrete semi-terrestrial habitat: Comparison of organisms with different dispersal abilities sampled in the same plots. J. Biogeogr. 2011, 38, 1683–1693. [Google Scholar] [CrossRef]
- Peltzer, D.A.; Bast, M.L.; Wilson, S.D.; Gerry, A.K. Plant diversity and tree responses following contrasting disturbances in boreal forest. For. Ecol. Manag. 2000, 127, 191–203. [Google Scholar] [CrossRef]
- Haeussler, S.; Bedford, L.; Leduc, A.; Bergeron, Y.; Kranabetter, J.M. Silvicultural disturbance severity and plant communities of the southern Canadian boreal forest. Silva Fenn. 2002, 76, 307–327. [Google Scholar] [CrossRef] [Green Version]
- Rees, D.C.; Juday, G.P. Plant species diversity on logged versus burned sites in central Alaska. For. Ecol. Manag. 2002, 155, 291–302. [Google Scholar] [CrossRef]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W. Zeigerwerte von Pflanzen in Mitteleuropa. 3rd ed. Scr. Geobot. 2001, 18, 1–18, (In German with English summary). [Google Scholar]
- Nordin, A.; Strengbom, J.; Forsum, Å.; Ericson, L. Complex biotic interactions drive long-term vegetation change in a nitrogen enriched boreal forest. Ecosystems 2009, 12, 1204–1211. [Google Scholar] [CrossRef]
- Hannerz, M.; Hånell, B. Effects on the flora of Norway spruce forests following clearcutting and shelterwood cutting. For. Ecol. Manag. 1997, 90, 29–49. [Google Scholar] [CrossRef]
- Bergstedt, J.; Milberg, P. The impact of logging intensity on field-layer vegetation in Swedish boreal forests. For. Ecol. Manag. 2001, 154, 105–115. [Google Scholar] [CrossRef]
- Bergstedt, J.; Hagner, M.; Milberg, P. Effects on vegetation composition of a modified forest harvesting and propagation method compared with clear-cutting, scarification and planting. Appl. Veg. Sci. 2008, 11, 159–168. [Google Scholar] [CrossRef]
- Vitt, D.H.; Li, Y.; Belland, R. Patterns of bryophyte diversity in peatlands of continental western Canada. Bryologist 1995, 98, 218–227. [Google Scholar] [CrossRef]
- Lõhmus, A.; Remm, L. Disentangling the effects of seminatural forestry on an ecosystem good: Bilberry (Vaccinium myrtillus) in Estonia. For. Ecol. Manag. 2017, 404, 75–83. [Google Scholar] [CrossRef]
- Atlegrim, O.; Sjöberg, K. Response of bilberry (Vaccinium myrtillus) to clear-cutting and single-tree selection harvests in uneven-aged boreal Picea abies forests. For. Ecol. Manag. 1996, 86, 39–50. [Google Scholar] [CrossRef]
- Mäkipää, R.; Heikkinen, J. Large-scale changes in abundance of terricolous bryophytes and macrolichens in Finland. J. Veg. Sci. 2003, 14, 497–508. [Google Scholar] [CrossRef]
- EEA. Europe’s Biodiversity—Biogeographical Regions and Seas. EEA Report 2002 No 1. 2002. Available online: https://www.eea.europa.eu/publications/report_2002_0524_154909 (accessed on 2 September 2022).
- Lietuvos Hidrometeorologijos Tarnyba [Lithuanian Hydrometeorological Service]. Standartine Klimato Norma 1991–2020 m; Lietuvos Hidrometeorologijos Tarnyba: Vilnius, Lithuania, 2021. [Google Scholar]
- Vaičys, M.; Karazija, S.; Kuliešis, A.; Rutkauskas, A. Miškų Augavietės (Forest Sites); Lutute: Kaunas, Lithuania, 2006; p. 95. (In Lithuanian) [Google Scholar]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014; International Soil Classification System for Naming 615 Soils and Creating Legends for Soil Maps; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015; p. 192. [Google Scholar]
- Bastrup-Birk, A.; Neville, P.; Chirici, G.; Houston, T. The BioSoil—Forest Biodiversity; Field Manual, Ver. 1.0/1.1/1.1a; for the field assessment 2006–2007, Forest Focus Demonstration Project; BioSoil, Forest Research: Farnham, UK, 2007; 51p. [Google Scholar]
- Ehlers, T.; Berch, S.M.; MacKinnon, A. Inventory of non-timber forest product plant and fungal species in the Robson Valley. BC J. Ecosyst. Manag. 2003, 4, 2. [Google Scholar]
- Gustienė, D.; Varnagirytė-Kabašinskienė, I.; Stakėnas, V. Ground vegetation, forest floor and mineral topsoil at clear-cuttings and reforested Scots pine stands of different age: A pilot study. J. For. Res. 2022, 33, 1247–1257. [Google Scholar] [CrossRef]
- Canullo, R.; Starlinger, F.; Granke, O.; Fischer, R.; Aamlid, D.; Neville, P. Assessment of Ground Vegetation. Manual Part VII.1. In ICP Forests Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; UNECE ICP Forests Programme Co-ordinating Centre: Hamburg, Germany, 2011; p. 9. [Google Scholar]
- Egnér, H.; Riehm, H.; Domingo, W. Untersuchungen über die chemische Bodenanalyse als Grundlage für die Beurteilung des Nährstoffzustandes der Böden. II. Chemische Extraktionsmethoden zur Phosphor- und Kaliumbestimmung. K. Lantbr. Ann. 1960, 26, 199–215. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Publishing: Oxford, UK, 2004; p. 256. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clear-Cuts (Years) | Age of Pinus sylvestris Stand (Years) | |||||||||||||||||||||||
| 1 | 2 | 6 | 10 | 15 | 30 | 70 | 130 | |||||||||||||||||
| Taxa names | MC (%) | F (%) | PV | MC (%) | F (%) | PV | MC (%) | F (%) | PV | MC (%) | F (%) | PV | MC (%) | F (%) | PV | MC (%) | F (%) | PV | MC (%) | F (%) | PV | MC (%) | F (%) | PV |
| Herbs and dwarf shrubs lower than 50 cm | ||||||||||||||||||||||||
| Agrostis tenuis Sibith. (syn. A. vulgaris) | 0.03 ± 0.02 | 15.00 | 0.12 | 0.34 ± 0.20 | 26.32 | 1.75 | ||||||||||||||||||
| Anthoxanthum odoratum L. | 0.07 ± 0.04 | 12.82 | 0.24 | |||||||||||||||||||||
| Arnica montana L. | * | |||||||||||||||||||||||
| Betula pendula Roth. | 0.06 ± 0.05 | 7.50 | 0.17 | 0.14 ± 0.05 | 30.77 | 0.77 | 0.18 ± 0.11 | 20.00 | 0.80 | 1.65 ± 0.55 | 55.00 | 12.20 | 0.13 ± 0.07 | 15.00 | 0.48 | 0.04 ± 0.03 | 10.53 | 0.14 | ||||||
| Calamagrostis arundinacea (L.) Roth | 0.04 ± 0.03 | 7.50 | 0.10 | 0.17 ± 0.15 | 15.00 | 0.67 | 0.01 ± 0.01 | 5.00 | 0.01 | |||||||||||||||
| Calluna vulgaris (L.) Hull | 0.07 ± 0.02 | 35.90 | 0.43 | 25.94 ± 3.40 | 100.0 | 259.4 | 23.38 ± 5.69 | 85.00 | 215.5 | 2.27 ± 1.25 | 25.00 | 11.33 | 0.05 ± 0.03 | 15.79 | 0.20 | |||||||||
| Carex sp. | 0.02 ± 0.02 | 2.56 | 0.04 | 0.03 ± 0.03 | 5.00 | 0.06 | * | 0.01 ± 0.01 | 5.26 | 0.01 | 0.02 ± 0.01 | 10.00 | 0.05 | |||||||||||
| Chamaenerion angustifolium (L.) Holub (syn. Epilobium angustifolium L.) | 0.02 ± 0.01 | 5.13 | 0.04 | 0.01 ± 0.01 | 5.00 | 0.03 | ||||||||||||||||||
| Chimaphila umbellata (L.) W. P. C. Barton | 0.04 ± 0.03 | 7.50 | 0.10 | 0.10 ± 0.10 | 5.00 | 0.22 | 0.04 ± 0.03 | 10.00 | 0.12 | |||||||||||||||
| Conyza canadensis (L.) Cronquist | 0.01 ± 0.01 | 2.56 | 0.01 | |||||||||||||||||||||
| Convallaria majalis L. | 0.01 ± 0.01 | 2.50 | 0.02 | 0.14 ± 0.14 | 2.56 | 0.23 | * | |||||||||||||||||
| Convolvulus arvensis L. | 0.02 ± 0.02 | 2.56 | 0.03 | |||||||||||||||||||||
| Epilobium parviflorum Schreb. | 0.03 ± 0.02 | 5.13 | 0.06 | 0.02 ± 0.02 | 5.00 | 0.04 | ||||||||||||||||||
| Festuca ovino L. | 0.31 ± 0.23 | 5.13 | 0.71 | 2.10 ± 1.12 | 20.00 | 9.38 | 0.38 ± 0.31 | 10.00 | 1.19 | 1.11 ± 0.42 | 95.00 | 10.82 | 0.03 ± 0.03 | 5.26 | 0.06 | 0.056 ± 0.04 | 10.00 | 0.17 | ||||||
| Festucapsammophila (Hack. ex Čelak.) Fritsch | 0.50 ± 0.29 | 12.50 | 1.76 | 0.26 ± 0.15 | 7.69 | 0.71 | 1.38 ± 0.66 | 30.00 | 7.53 | 4.73 ± 2.25 | 52.63 | 34.29 | 0.09 ± 0.06 | 10.00 | 0.27 | |||||||||
| Fragaria vesca L. | 0.06 ± 0.04 | 7.69 | 0.17 | 0.10 ± 0.07 | 10.00 | 0.32 | 0.03 ± 0.03 | 5.00 | 0.07 | 0.02 ± 0.02 | 5.26 | 0.04 | ||||||||||||
| Frangula alnus Mill. | 0.03 ± 0.02 | 7.50 | 0.08 | 0.05 ± 0.02 | 20.51 | 0.24 | 0.16 ± 0.08 | 10.00 | 0.51 | * | 0.07 ± 0.03 | 21.05 | 0.31 | |||||||||||
| Galeopsis tetrahit L. | 0.03 ± 0.03 | 2.50 | 0.04 | 0.26 ± 0.14 | 10.26 | 0.84 | 0.01 ± 0.01 | 5.00 | 0.01 | 0.04 ± 0.03 | 10.53 | 0.12 | ||||||||||||
| Goodyera repens (L.) R. Br. | 0.05 ± 0.05 | 5.26 | 0.11 | 0.07 ± 0.03 | 25.00 | 0.33 | 0.10 ± 0.05 | 20.00 | 0.45 | |||||||||||||||
| Hieracium murorum L. | 0.12 ± 0.09 | 10.00 | 0.36 | |||||||||||||||||||||
| Hieracium pilosella L. | 0.04 ± 0.04 | 2.50 | 0.06 | |||||||||||||||||||||
| Hypericum perforatum L. | 0.03 ± 0.03 | 7.69 | 0.08 | 0.01 ± 0.01 | 5.00 | 0.02 | ||||||||||||||||||
| Hypochaeris radicata L. | 0.04 ± 0.03 | 5.13 | 0.10 | |||||||||||||||||||||
| Juniperus communis L. | * | * | * | 0.30 ± 0.30 | 5.00 | 0.67 | 0.01 ± 0.01 | 5.00 | 0.02 | * | 0.01 ± 0.01 | * | ||||||||||||
| Knautia arvensis (L.) Coult | 0.10 ± 0.10 | 2.56 | 0.16 | 0.03 ± 0.03 | 5.00 | 0.06 | ||||||||||||||||||
| Leontodon hispidus L. | 0.06 ± 0.06 | 2.56 | 0.10 | |||||||||||||||||||||
| Lycopodium clavatum L. | 0.43 ± 0.23 | 15.00 | 1.66 | 0.14 ± 0.10 | 7.69 | 0.38 | * | * | ||||||||||||||||
| Luzula pilosa (L.) Willd | 0.35 ± 0.15 | 27.50 | 1.84 | 1.07 ± 0.21 | 76.92 | 9.38 | 0.02 ± 0.01 | 10.00 | 0.05 | 2.10 ± 1.20 | 45.00 | 14.07 | 0.22 ± 0.10 | 25.00 | 1.08 | 0.38 ± 0.09 | 63.16 | 2.99 | 0.18 ± 0.09 | 25.00 | 0.90 | 0.10 ± 0.10 | 5.00 | 0.22 |
| Maianthemum bifolium (L.) F.W.Schmidt | * | |||||||||||||||||||||||
| Melampyrum pratense L. | 0.01 ± 0.01 | 2.50 | 0.02 | 0.10 ± 0.04 | 15.38 | 0.38 | 1.53 ± 0.47 | 55.00 | 11.31 | 0.43 ± 0.16 | 70.00 | 3.56 | 0.16 ± 0.16 | 5.26 | 0.36 | 0.56 ± 0.20 | 40.00 | 3.54 | * | |||||
| Picea abies (L.) H. Karst | 0.04 ± 0.04 | 5.00 | 0.08 | 0.01 ± 0.01 | 5.26 | 0.02 | ||||||||||||||||||
| Pinus sylvestris L. | 0.11 ± 0.04 | 27.50 | 0.59 | 0.64 ± 0.11 | 76.92 | 5.60 | 0.44 ± 0.24 | 30.00 | 2.41 | 0.02 ± 0.01 | 15.79 | 0.06 | ||||||||||||
| Plantago lanceolata L. | 0.01 ± 0.01 | 2.56 | 0.01 | |||||||||||||||||||||
| Polygonum maculosa L. | 0.25 ± 0.25 | 2.56 | 0.40 | |||||||||||||||||||||
| Populus tremula L. | 0.04 ± 0.04 | 2.50 | 0.06 | 0.07 ± 0.07 | 5.00 | 0.16 | 0.44 ± 0.17 | 35.00 | 2.60 | 0.02 ± 0.02 | 5.26 | 0.04 | ||||||||||||
| Potentilla arenaria Borkh. | * | * | ||||||||||||||||||||||
| Pteridium aquilinum (L.) Kuhn | 2.26 ± 0.95 | 15.00 | 8.73 | 2.37 ± 1.28 | 20.00 | 10.60 | 2.4 ± 1.70 | 30.00 | 13.15 | 0.15 ± 0.15 | 5.00 | 0.34 | ||||||||||||
| Rubus caesius L. | 0.41 ± 0.26 | 17.95 | 1.75 | 0.49 ± 0.30 | 20.00 | 2.19 | ||||||||||||||||||
| Rubus idaeus L. | 0.13 ± 0.06 | 15.00 | 0.51 | 0.80 ± 0.30 | 30.77 | 4.43 | 1.79 ± 0.87 | 55.00 | 13.26 | 5.65 ± 1.65 | 65.00 | 45.53 | 0.18 ± 0.09 | 20.00 | 0.78 | 0.10 ± 0.07 | 10.53 | 0.33 | ||||||
| Rumex acetosella L. | 0.16 ± 0.13 | 5.13 | 0.37 | 0.09 ± 0.06 | 15.00 | 0.34 | 0.08 ± 0.08 | 5.00 | 0.18 | * | ||||||||||||||
| Salix caprea L. | 0.21 ± 0.12 | 7.50 | 0.57 | 0.31 ± 0.08 | 43.59 | 2.05 | ||||||||||||||||||
| Senecio vulgaris L. | 0.19 ± 0.18 | 5.00 | 0.42 | 4.83 ± 0.85 | 76.92 | 42.36 | 0.19 ± 0.11 | 25.00 | 0.93 | |||||||||||||||
| Sorbus aucuparia L. | 0.07 ± 0.07 | 2.50 | 0.11 | 0.14 ± 0.14 | 5.00 | 0.30 | ||||||||||||||||||
| Succisa pratensis Moench | 0.01 ± 0.01 | 2.56 | 0.01 | |||||||||||||||||||||
| Trientalis europaea L. | 0.68 ± 0.22 | 45.00 | 4.53 | 1.12 ± 0.32 | 46.15 | 7.61 | 0.11 ± 0.05 | 25.00 | 0.55 | 0.04 ± 0.04 | 5.00 | 0.08 | ||||||||||||
| Trifolium repens L. | 0.01 ± 0.00 | 10.26 | 0.03 | |||||||||||||||||||||
| Vaccinium myrtillus L. | 6.76 ± 0.83 | 95.00 | 65.90 | 6.64 ± 0.98 | 97.44 | 65.51 | 0.50 ± 0.28 | 30.00 | 2.72 | 7.83 ± 2.11 | 75.00 | 67.83 | 4.56 ± 1.23 | 65.00 | 36.74 | 13.0 ± 4.42 | 68.42 | 107.5 | 22.47 ± 2.25 | 100.0 | 224.7 | 10.58 ± 1.92 | 100.0 | 105.8 |
| Vaccinium vitis-idaea L. | 2.20 ± 0.34 | 80.00 | 19.63 | 1.89 ± 0.45 | 71.79 | 16.01 | 5.08 ± 2.14 | 65.00 | 40.96 | 13.42 ± 3.20 | 80.00 | 120.0 | 24.82 ± 2.44 | 100.0 | 248.2 | 6.30 ± 1.66 | 100.0 | 63.02 | 15.87 ± 1.96 | 100.0 | 158.7 | 5.81 ± 2.03 | 65.00 | 46.80 |
| Veronica officinalis L. | 0.31 ± 0.15 | 15.38 | 1.21 | 0.11 ± 0.07 | 20.00 | 0.50 | 0.02 ± 0.02 | 5.26 | 0.04 | |||||||||||||||
| Verbascum thapsus L. | * | 0.02 ± 0.02 | 5.00 | 0.04 | ||||||||||||||||||||
| Viola canina L. | 0.02 ± 0.02 | 5.13 | 0.05 | 0.01 ± 0.01 | 5.00 | 0.03 | ||||||||||||||||||
| Herbs and dwarf shrubs higher than 50 cm | ||||||||||||||||||||||||
| Betula pendula Roth. | 0.04 ± 0.04 | 6.00 | 0.10 | |||||||||||||||||||||
| Calamagrostis arundinacea (L.) Roth | 0.01 ± 0.01 | 5.00 | 0.02 | 0.35 ± 0.19 | 25.00 | 1.73 | 0.06 ± 0.06 | 5.00 | 0.12 | |||||||||||||||
| Pinus sylvestris L. | 0.76 ± 0.76 | 1.00 | 0.76 | |||||||||||||||||||||
| Pteridium aquilinum (L.) Kuhn | 0.62 ± 0.36 | 8.00 | 1.75 | 0.65 ± 0.41 | 9.00 | 1.95 | ||||||||||||||||||
| Mosses and lichens | ||||||||||||||||||||||||
| Cirriphyllum piliferum (Hed.) Grout. | 28.98 ± 6.44 | 75.00 | 250.9 | |||||||||||||||||||||
| Cladonia fimbriata (L.) Fr. | 0.09 ± 0.03 | 40.00 | 0.60 | 0.48 ± 0.45 | 5.00 | 1.06 | 0.40 ± 0.40 | 5.00 | 0.89 | |||||||||||||||
| Dicranum polysetum Sw. | 0.22 ± 0.13 | 12.50 | 0.78 | 0.18 ± 0.11 | 20.51 | 0.80 | 2.31 ± 1.95 | 25.00 | 11.55 | 4.75 ± 2.18 | 35.00 | 28.10 | 10.49 ± 4.55 | 100.0 | 104.9 | 3.74 ± 2.04 | 63.16 | 29.73 | 1.19 ± 0.68 | 30.00 | 6.49 | 0.07 ± 0.05 | 10.00 | 0.23 |
| Hylocium splendens (Hedw.) Schimp. | 13.57 ± 2.91 | 40.00 | 85.82 | 0.48 ± 0.24 | 17.95 | 2.03 | 1.61 ± 0.92 | 15.00 | 6.23 | 1.40 ± 0.59 | 40.0 | 8.85 | 0.83 ± 0.35 | 30.0 | 4.52 | 30.32 ± 6.41 | 78.95 | 269.4 | 13.71 ± 4.21 | 70.00 | 114.7 | 53.73 ± 8.00 | 100.0 | 537.3 |
| Pleurozium schreberi (Brid.) Mitt. | 37.18 ± 5.16 | 80.00 | 332.5 | 13.34 ± 2.82 | 84.62 | 122.7 | 5.19 ± 1.56 | 85.00 | 47.82 | 33.58 ± 5.09 | 90.0 | 318.5 | 17.50 ± 7.45 | 35.0 | 103.5 | 50.68 ± 7.18 | 100.0 | 506.8 | 67.78 ± 6.30 | 100.0 | 677.8 | 20.94 ± 6.21 | 75.00 | 181.4 |
| Pohlia nutans (Hedw.) Lindb. | 0.45 ± 0.24 | 25.64 | 2.27 | 13.89 ± 4.11 | 80.00 | 124.3 | 0.75 ± 0.34 | 25.00 | 3.75 | |||||||||||||||
| Polytrichum commune L. | 0.01 ± 0.01 | 2.56 | 0.02 | 1.68 ± 0.62 | 60.00 | 13.02 | 6.55 ± 1.80 | 65.00 | 52.81 | 0.50 ± 0.26 | 20.00 | 2.24 | 2.87 ± 2.87 | 5.26 | 6.59 | 2.80 ± 1.07 | 55.00 | 20.73 | ||||||
| Polytrichum juniperinum Hedw. | 0.01 ± 0.02 | 5.13 | 0.02 | |||||||||||||||||||||
| Ptilium crista-castrensis (Hedw.) De Not | 5.13 ± 3.02 | 7.50 | 14.05 | 3.36 ± 1.54 | 33.33 | 19.39 | 4.13 ± 1.45 | 55.00 | 30.59 | 1.15 ± 1.10 | 10.00 | 3.64 | 5.56 ± 2.99 | 36.84 | 33.72 | 10.20 ± 3.88 | 60.00 | 79.01 | 14.11 ± 6.09 | 40.00 | 89.21 | |||
| Rhacomitrium canescens (Hedw.) Brid. | 0.52 ± 0.52 | 5.00 | 1.16 | 1.35 ± 0.70 | 25.00 | 6.75 | 0.30 ± 0.30 | 5.00 | 0.67 | |||||||||||||||
| Clear-Cut (Years) | Pinus sylvestris Stand (Years) | |||||||||||||||
| 1 | 2 | 6 | 10 | 15 | 30 | 70 | 130 | |||||||||
| MC (%) | MR | MC (%) | MR | MC (%) | MR | MC (%) | MR | MC (%) | MR | MC (%) | MR | MC (%) | MR | MC (%) | MR | |
| Mosses | 56.2 ± 4.2 | 2.5 ± 0.2 | 18.1 ± 3.3 | 4.3 ± 0.3 | 25.1 ± 5.4 | 5.0 ± 0.7 | 53.0 ± 4.4 | 6.5 ± 0.5 | 59.8 ± 6.3 | 5.3 ± 0.3 | 92.5 ± 1.3 | 4.3 ± 0.3 | 96.0 ± 2.4 | 4.8 ± 0.6 | 88.9 ± 3.5 | 3.5 ± 0.3 |
| Herbs, dwarf shrubs | 14.5 ± 2.1 | 9.3 ± 1.3 | 23.2 ± 2.3 | 17.4 ± 0.7 | 39.0 ± 1.8 | 10.0 ± 1.6 | 59.0 ± 4.4 | 12.8 ± 1.6 | 34.6 ± 1.4 | 9.8 ± 0.8 | 24.1 ± 7.1 | 10.3 ± 1.0 | 39.6 ± 3.1 | 6.5 ± 0.3 | 16.6 ± 3.1 | 3.5 ± 0.7 |
| Forest floor cover | 95.8 ± 1.7 | - | 39.6 ± 4.7 | - | 86.6 ± 2.8 | - | 80.1 ± 4.8 | - | 99.8 ± 0.1 | - | 100.0 ± 0.0 | - | 100.0 ± 0.0 | - | 100.0 ± 0.0 | - |
| Clear-Cut (Years) | Age of Pinus sylvestris Stand (Years) | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 6 | 10 | 15 | 30 | 70 | 130 | |
| Shannon diversity index (H′) | 1.58 ± 0.10 b | 2.04 ± 0.10 c | 1.52 ± 0.13 b | 1.85 ± 0.06 bc | 1.89 ± 0.06 bc | 1.67 ± 0.14 bc | 0.95 ± 0.09a | 1.34 ± 0.06 b |
| Equitability (E) | 0.48 ± 0.04 a | 0.59 ± 0.03 ab | 0.55 ± 0.04 ab | 0.68 ± 0.01b | 0.44 ± 0.08 a | 0.51 ± 0.03 a | 0.44 ± 0.05 a | 0.58 ± 0.03 ab |
| Ellenberg Indicator Values | Clear-Cut (Years) | Age of Pinus sylvestris Stand (Years) | ||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 6 | 10 | 15 | 30 | 70 | 130 | |
| Herbs and dwarf shrubs | ||||||||
| Light (L) | 5.4 | 6.0 | 7.3 | 6.5 | 5.3 | 5.0 | 5.0 | 5.0 |
| Temperature (T) | 4.9 | 5.0 | 5.0 | 5.1 | 4.9 | 5.5 | 5.4 | 5.9 |
| Moisture (F) | 4.6 | 4.7 | 4.3 | 4.2 | 4.0 | 4.1 | 4.0 | 4.0 |
| pH | 2.4 | 2.8 | 1.4 | 1.8 | 2.0 | 2.1 | 2.0 | 2.0 |
| Nitrogen (N) | 2.7 | 3.2 | 1.4 | 2.0 | 1.4 | 2.4 | 2.2 | 2.3 |
| Mosses | ||||||||
| Light (L) | 5.8 | 5.6 | 5.5 | 6.4 | 6.5 | 5.9 | 5.8 | 5.7 |
| Temperature (T) | 2.9 | 2.8 | 1.3 | 2.9 | 3.0 | 2.9 | 2.9 | 2.8 |
| Moisture (F) | 4.2 | 4.4 | 4.2 | 5.1 | 4.6 | 4.2 | 4.3 | 4.3 |
| pH | 2.8 | 2.3 | 2.5 | 3.1 | 3.5 | 3.2 | 2.6 | 4.0 |
| pHCaCl2 | Concentrations in Forest Floor (FF) and Mineral Soil at a Depth of 0–10 cm (MS) | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Organic C (g kg–1) | Total N (g kg–1) | Total P (mg kg–1) | Total K (mg kg–1) | Total Ca (mg kg–1) | Total Mg (mg kg–1) | P2O5 (mg kg–1) | K2O (mg kg–1) | Mobile Ca (mg kg–1) | Mobile Mg (mg kg–1) | Mineral N (mg kg–1) | |||||||||||||||
| r | p | r | p | r | p | r | p | r | p | r | p | r | p | r | p | r | p | r | p | r | p | r | p | ||
| Vaccinium myrtillus | FF | 0.687 | 0.000 | −0.581 | 0.000 | 0.697 | 0.000 | −0.704 | 0.000 | −0.701 | 0.000 | −0.700 | 0.000 | −0.532 | 0.002 | n.d.* | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| MS | 0.389 | 0.033 | n.s.** | n.s. | n.s. | n.s. | −0.698 | 0.000 | −0.751 | 0.000 | −0.675 | 0.000 | −0.731 | 0.000 | −0.580 | 0.000 | −0.721 | 0.000 | −0.724 | 0.000 | n.s. | n.s. | 0.469 | 0.009 | |
| Vaccinium vitis-idaea | FF | 0.560 | 0.001 | −0.497 | 0.004 | 0.447 | 0.010 | −0.643 | 0.000 | −0.644 | 0.000 | −0.628 | 0.000 | −0.513 | 0.003 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| MS | 0.685 | 0.000 | 0.607 | 0.000 | 0.666 | 0.000 | −0.665 | 0.000 | −0.688 | 0.000 | −0.657 | 0.000 | −0.697 | 0.000 | −0.597 | 0.001 | −0.693 | 0.000 | −0.611 | 0.000 | −0.486 | 0.006 | 0.453 | 0.012 | |
| Luzula pilosa | FF | 0.504 | 0.004 | −0.485 | 0.007 | 0.494 | 0.006 | −0.577 | 0.000 | −0.580 | 0.001 | −0.551 | 0.002 | −0.420 | 0.021 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| MS | 0.413 | 0.023 | 0.581 | 0.001 | 0.652 | 0.000 | −0.568 | 0.001 | −0.590 | 0.001 | −0.529 | 0.003 | −0.587 | 0.001 | −0.507 | 0.004 | - | −0.565 | −0.553 | 0.002 | n.s. | n.s. | 0.416 | 0.023 | |
| Melampyrum pratense | FF | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | −0.442 | 0.015 | −0.420 | 0.021 | n.s. | n.s. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| MS | 0.468 | 0.009 | n.s. | n.s. | 0.510 | 0.004 | −0.443 | 0.014 | 0.457 | 0.011 | −0.403 | 0.028 | −0.453 | 0.012 | −0.393 | 0.032 | −0.433 | 0.017 | −0.444 | 0.001 | n.s. | n.s. | n.s. | n.s. | |
| Rubus idaeus | FF | 0.564 | 0.001 | −0.390 | 0.035 | n.s. | n.s. | −0.439 | 0.015 | −0.446 | 0.014 | −0.414 | 0.023 | n.s. | n.s. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| MS | n.s. | n.s. | n.s. | n.s. | 0.510 | 0.004 | −0.443 | 0.014 | −0.453 | 0.012 | −0.409 | 0.025 | −0.409 | 0.252 | −0.401 | 0.029 | −0.430 | 0.018 | −0.425 | 0.019 | n.s. | n.s. | 0.421 | 0.020 | |
| Pleurozium schreberi | FF | 0.384 | 0.036 | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| MS | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | −0.365 | 0.047 | n.s. | n.s. | |
| Dicranum polysetum | FF | 0.391 | 0.033 | n.s. | n.s. | 0.471 | 0.007 | n.s. | n.s. | −0.406 | 0.026 | 0.386 | 0.035 | n.s. | n.s. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| MS | n.s. | n.s. | n.s. | n.s. | 0.406 | 0.026 | −0.410 | 0.029 | −0.424 | 0.020 | −0.391 | 0.033 | −0.423 | 0.020 | n.s. | n.s. | −0.413 | 0.023 | −0.401 | 0.029 | −0.417 | 0.022 | n.s. | n.s. | |
| Hylocium splendens | FF | 0.509 | 0.004 | 0.568 | 0.001 | n.s. | n.s. | −0.556 | 0.001 | −0.572 | 0.001 | −0.509 | 0.004 | 0.465 | 0.010 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| MS | 0.395 | 0.031 | n.s. | n.s. | 0.508 | 0.004 | −0.563 | 0.001 | −0.577 | 0.001 | −0.436 | 0.016 | −0.555 | 0.002 | −0.480 | 0.007 | −0.578 | 0.001 | −0.579 | 0.001 | −0.531 | 0.003 | 0.441 | 0.015 | |
| Ptilium crista- castrensis | FF | n.s. | n.s. | 0.436 | 0.016 | 0.465 | 0.010 | −0.496 | 0.005 | −0.500 | 0.005 | −0.486 | 0.006 | −0.386 | 0.035 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| MS | n.s. | n.s. | 0.502 | 0.005 | 0.590 | 0.001 | −0.491 | 0.006 | −0.523 | 0.003 | −0.456 | 0.011 | −0.512 | 0.004 | −0.419 | 0.022 | −0.500 | 0.005 | −0.515 | 0.004 | n.s. | n.s. | n.s. | n.s. | |
| Clear-Cut (Years) | Age of Pinus sylvestris Stand (Years) | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 6 | 10 | 15 | 30 | 70 | 130 | |
| Mosses (kg ha–1) | 5269 ± 634 c | 1063 ± 302 a | 1153 ± 248 a | 3790 ± 54 b | 2374 ± 973 ab | 3136 ± 830 b | 1965 ± 931 ab | 5353 ± 964 c |
| Herbs and dwarf shrubs (kg ha–1) | 1207 ± 352 b | 7120 ± 502 c | 309 ± 214 a | 10,650 ± 353 d | 12993 ± 823 d | 1148 ± 420 b | 7094 ± 1248 c | 1848 ± 191 b |
| Total (kg ha–1) | 6476 ± 725 c | 8183 ± 586 d | 1462 ± 328 a | 14440 ± 357 e | 15368 ± 1275 e | 4284 ± 930 b | 9059 ± 1557 d | 7201 ± 983 cd |
| Study Site (Years) | Tree Species Composition* (%) | Mean Tree H (m) | Mean Tree DBH (cm) | Stand Volume (m3 ha– 1) | Basal Area (m2 ha– 1) | Underbrush | ||
|---|---|---|---|---|---|---|---|---|
| Species Composition (%) * | Density (%) | Mean H (m) | ||||||
| Clear-cuts | ||||||||
| 1 | - | - | - | - | - | 43Gw+23P+14Rw+12B+8A | 50 | 0.1 |
| 2 | 90 P + 10 S | 0.2 | - | - | - | 39Gw+38P+18B+5Bc | 40 | 0.2 |
| Pinus sylvestrisstands | ||||||||
| 6 | 100 P | 1.6 | 0.4 | 10 | - | 90 B + 10 P | 46 | 1.0 |
| 10 | 100 P | 4.5 | 5.0 | 58 | 16 | 55B+39Bc+3S+3A | 30 | 1.3 |
| 15 | 100 P | 7.0 | 9.0 | 56 | 14 | 56A+17Bc+11J+11P+5B | 25 | 1.3 |
| 30 | 90 P + 10 S | 15.0 | 15.0 | 165 | 28 | 55S+36J+9Bc | 20 | 1.5 |
| 70 | 100 P | 25.0 | 25.0 | 294 | 32 | 60Bc+40J | 10 | 2.0 |
| 130 | 100 P | 29.0 | 38.0 | 465 | 36 | 50S+30J+20Bc | 20 | 2.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gustienė, D.; Varnagirytė-Kabašinskienė, I.; Stakėnas, V. Ground Vegetation in Pinus sylvestris Forests at Different Successional Stages following Clear Cuttings: A Case Study. Plants 2022, 11, 2651. https://doi.org/10.3390/plants11192651
Gustienė D, Varnagirytė-Kabašinskienė I, Stakėnas V. Ground Vegetation in Pinus sylvestris Forests at Different Successional Stages following Clear Cuttings: A Case Study. Plants. 2022; 11(19):2651. https://doi.org/10.3390/plants11192651
Chicago/Turabian StyleGustienė, Dovilė, Iveta Varnagirytė-Kabašinskienė, and Vidas Stakėnas. 2022. "Ground Vegetation in Pinus sylvestris Forests at Different Successional Stages following Clear Cuttings: A Case Study" Plants 11, no. 19: 2651. https://doi.org/10.3390/plants11192651