
Performance and Genetic Parameters of Poplar Hybrids and Clones in a Field Trial Are Modified by Contrasting Environmental Conditions during the Vegetative Propagation Phase

Abstract
:1. Introduction
2. Results
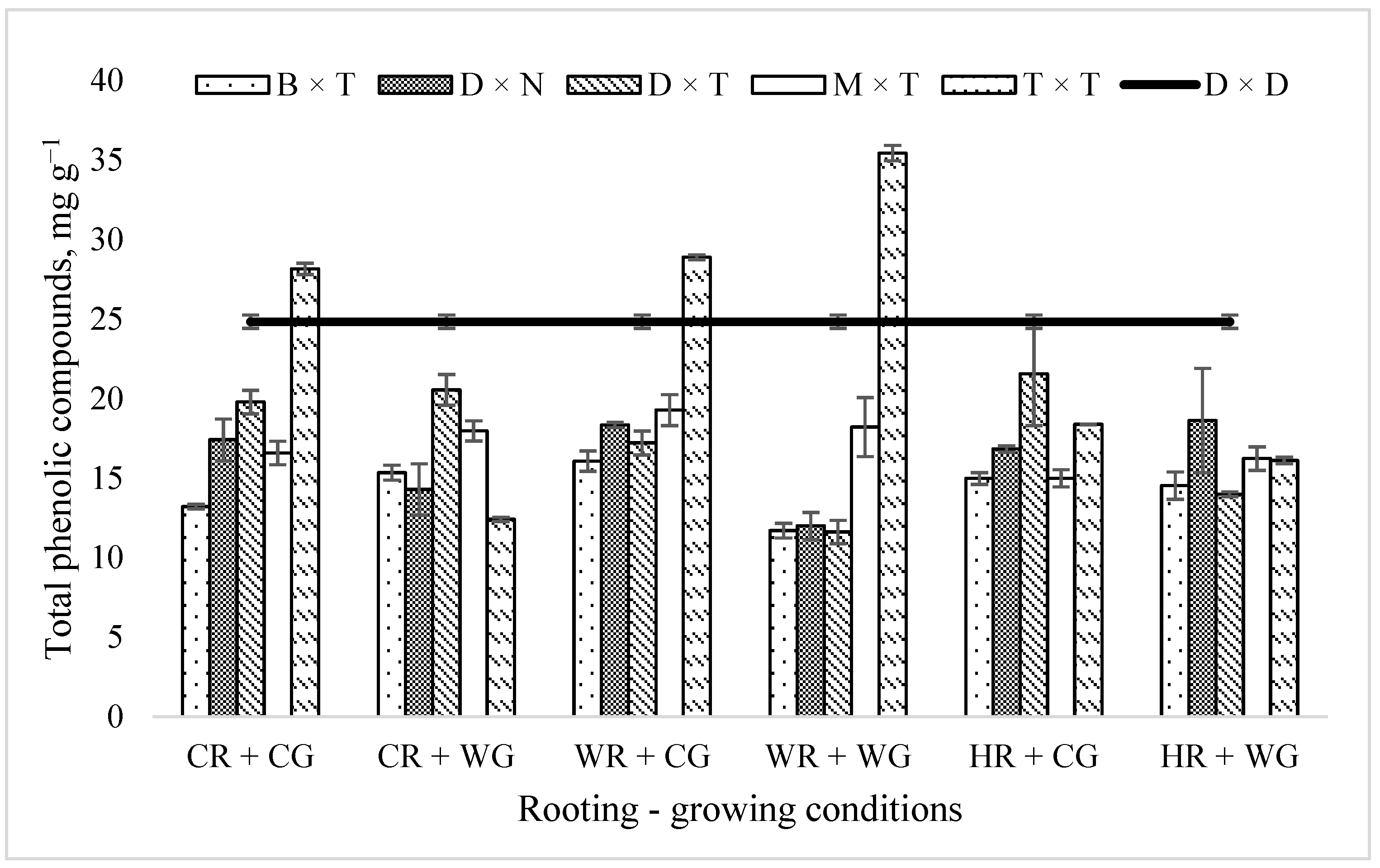
2.1. Impact of Hybrids and Treatments during Vegetative Propagation on the Total Phenolic Compound Content in Hybrid Populus Leaves in a Clonal Field Trial
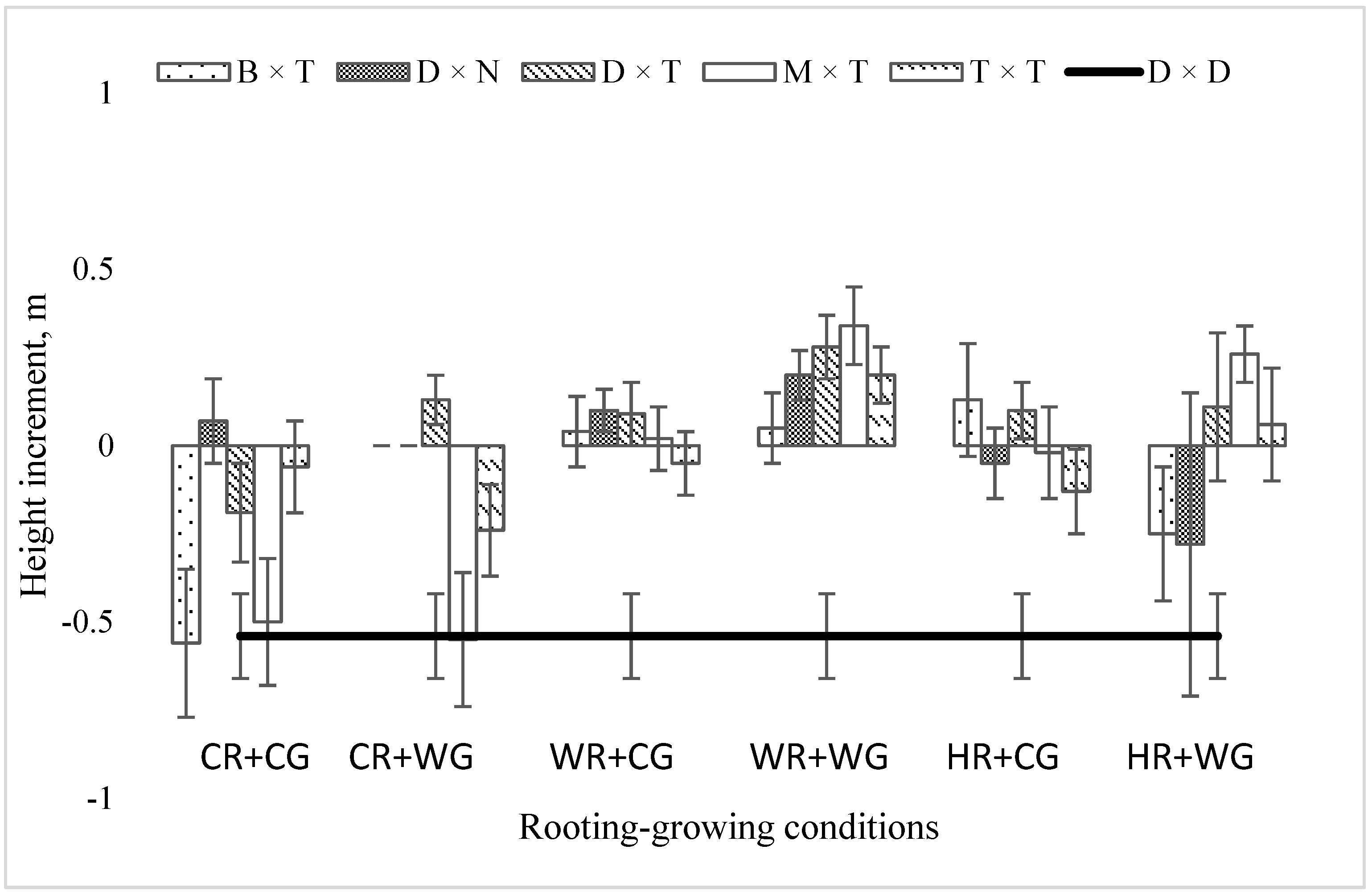
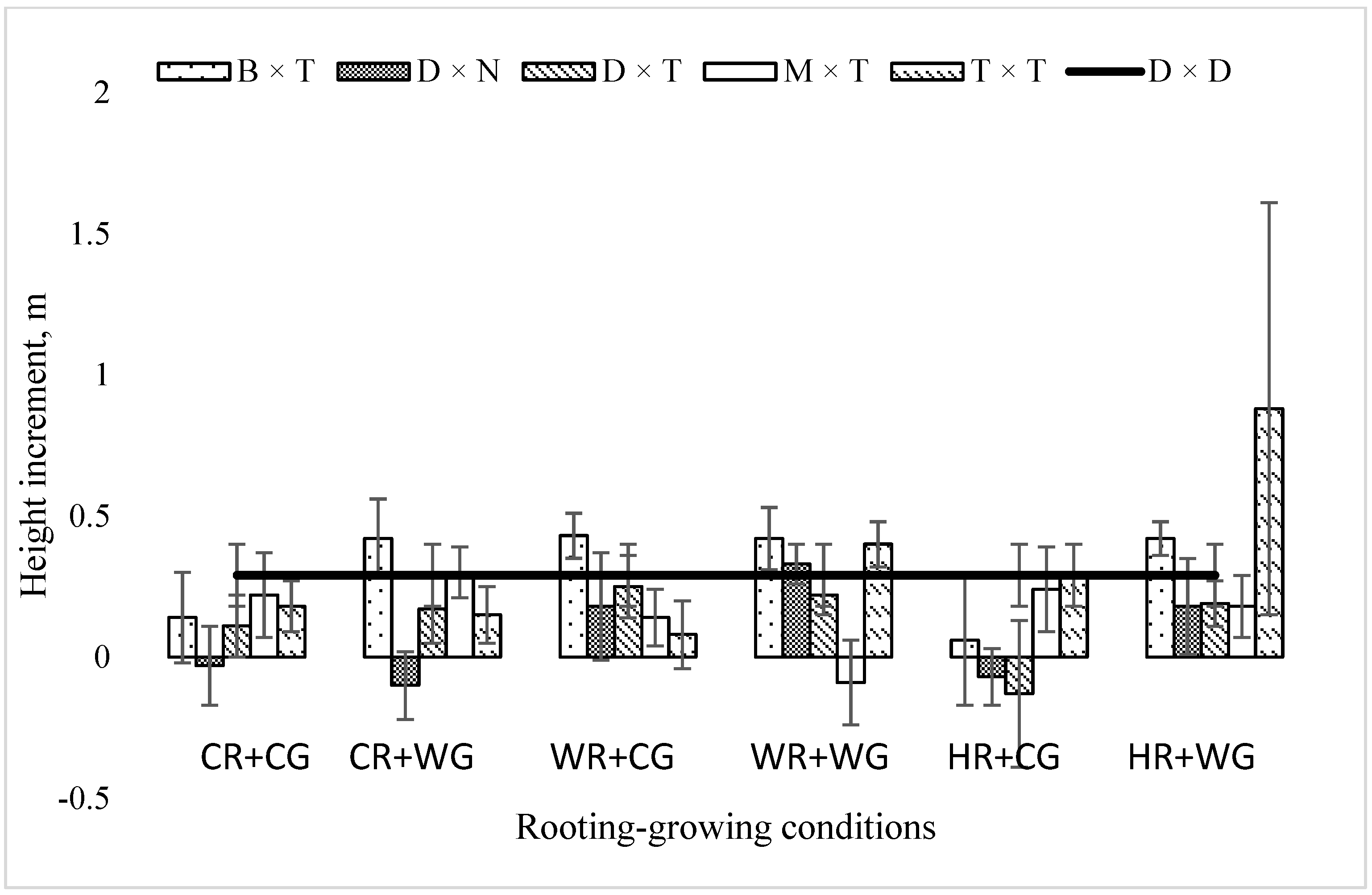
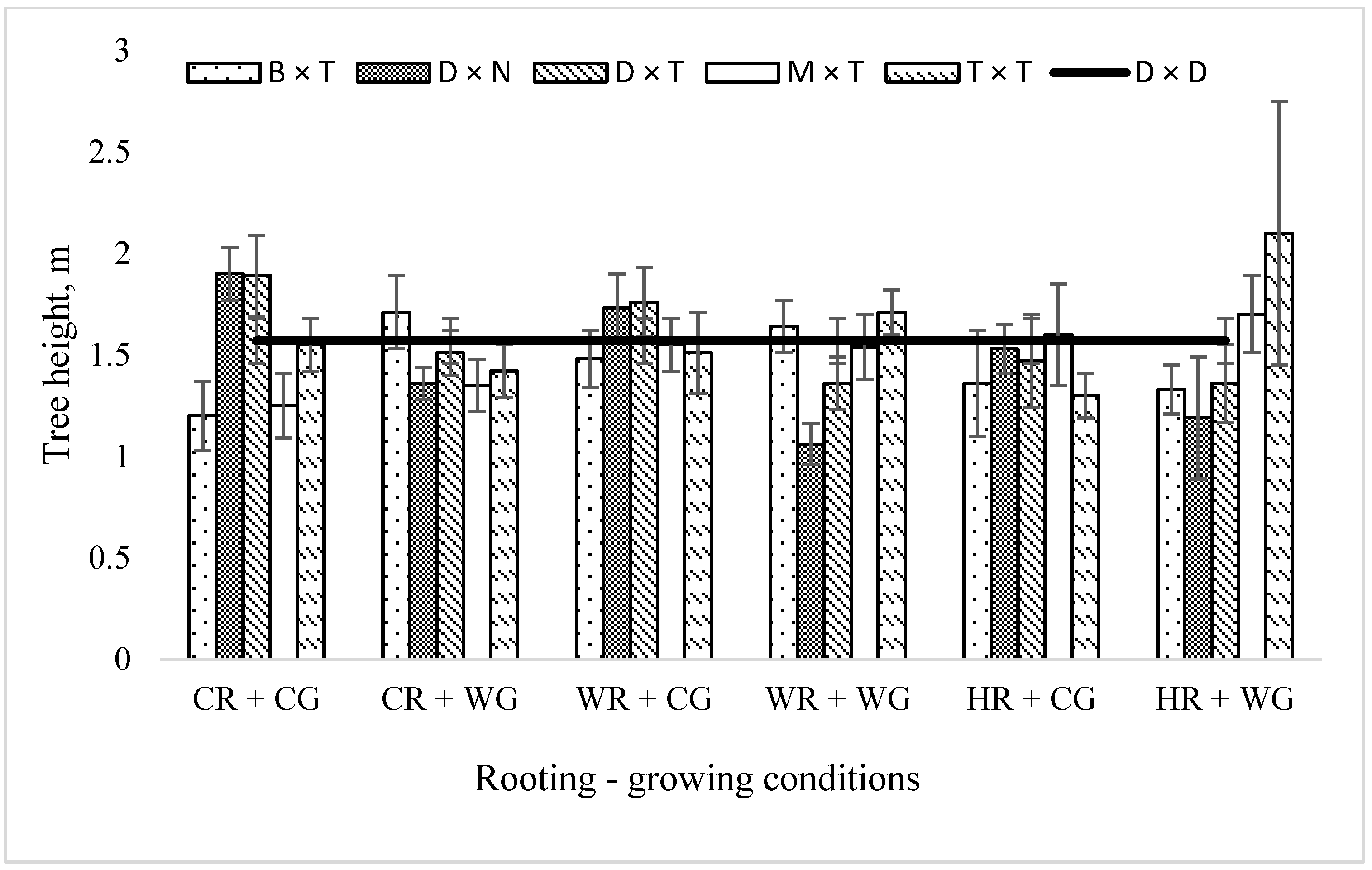
2.2. Dependence of the Growth Rate of Poplar Hybrids in a Clonal Field Trial on the Vegetative Propagation (Rooting–Growing) Conditions
2.3. Changes in Genetic Parameters of Growth Traits and Total Phenolic Compounds of Trees in the Clonal Field Trial Depending on Rooting–Growing Treatments during Vegetative Propagation in the Phytotron Greenhouse
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Design of Experiment
4.3. Measurements and Total Phenolic Compounds Extraction
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sicard, P.; Augustaitis, A.; Belyazid, S.; Calfapietra, C.; De Marco, A.; Fenn, M.E.; Bytnerowicz, A.; Grulke, N.E.; He, S.; Matyssek, R.; et al. Global topics and novel approaches in the study of air pollution, climate change and forest ecosystems. Environ. Pollut. 2016, 213, 977–987. [Google Scholar] [CrossRef] [PubMed]
- Pollastrini, M.; Puletti, N.; Selvi, F.; Iacopetti, G.; Bussotti, F. Widespread crown defoliation after a drought and heat wave in the forests of Tuscany (Central Italy) and their recovery—a case study from summer 2017. Front. For. Glob. Chang. 2019, 2, 74. [Google Scholar] [CrossRef]
- Pliura, A.; Jankauskiene, J.; Lygis, V.; Suchockas, V.; Bajerkevičiene, G.; Verbylaite, R. Response of juvenile progeny of seven forest tree species and their populations to simulated climate change-related stressors, heat, elevated humidity and drought. iFor.—Biogeosci. For. 2018, 11, 374–388. [Google Scholar] [CrossRef]
- Gudynaitė-Franckevičienė, V.; Pliūra, A.; Suchockas, V. Ecogenetic plasticity and genetic variation in Populus hybrids under the impact of simulated climate change related stressors. Balt. For. 2020, 26, 462. [Google Scholar] [CrossRef]
- Diffenbaugh, N.S.; Field, C.B. Changes in ecologically critical terrestrial climate conditions. Science 2013, 341, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Giesecke, T.; Brewer, S.; Finsinger, W.; Leydet, M.; Bradshaw, R.H. Patterns and dynamics of European vegetation change over the last 15,000 years. J. Biogeogr. 2017, 44, 1441–1456. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Matthes, M.C.; Napier, J.A.; Pickett, J.A. Stressful “memories” of plants: Evidence and possible mechanisms. Plant Sci. 2007, 173, 603–608. [Google Scholar] [CrossRef]
- Feeley, K.J.; Rehm, E.M.; Machovina, B. Perspective: The responses of tropical forest species to global climate change: Acclimate, adapt, migrate, or go extinct? Front. Biogeogr. 2012, 4. [Google Scholar] [CrossRef]
- Amaral, J.; Ribeyre, Z.; Vigneaud, J.; Sow, M.D.; Fichot, R.; Messier, C.; Pinto, G.; Nolet, P.; Maury, S. Advances and promises of epigenetics for forest trees. Forests 2020, 11, 976. [Google Scholar] [CrossRef]
- Kvaalen, H.; Johnsen, Ø. Timing of bud set in Picea abies is regulated by a memory of temperature during zygotic and somatic embryogenesis. New Phytol. 2008, 177, 49–59. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Zhu, J.-K. Epigenetic regulation of stress responses in plants. Curr. Opin. Plant Biol. 2009, 12, 133–139. [Google Scholar] [CrossRef]
- Bräutigam, K.; Vining, K.J.; Lafon-Placette, C.; Fossdal, C.G.; Mirouze, M.; Marcos, J.G.; Fluch, S.; Fraga, M.F.; Guevara, M.Á.; Abarca, D.; et al. Epigenetic regulation of adaptive responses of forest tree species to the environment. Ecol. Evol. 2013, 3, 399–415. [Google Scholar] [CrossRef]
- Eriksson, G.; Namkoong, G.; Roberds, J.H. Dynamic gene conservation for uncertain futures. For. Ecol. Manag. 1993, 62, 15–37. [Google Scholar] [CrossRef]
- Matesanz, S.; Gianoli, E.; Valladares, F. Global change and the evolution of phenotypic plasticity in plants. Ann. N. Y. Acad. Sci. 2010, 1206, 35–55. [Google Scholar] [CrossRef]
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef]
- Boyko, A.; Kovalchuk, I. Epigenetic control of plant stress response. Environ. Mol. Mutagen. 2008, 49, 61–72. [Google Scholar] [CrossRef]
- Groszmann, M.; Greaves, I.K.; Albertyn, Z.I.; Scofield, G.N.; Peacock, W.J.; Dennis, E.S. Changes in 24-nt siRNA levels in Arabidopsis hybrids suggest an epigenetic contribution to hybrid vigor. Proc. Natl. Acad. Sci. USA 2011, 108, 2617–2622. [Google Scholar] [CrossRef]
- Mirouze, M.; Paszkowski, J. Epigenetic contribution to stress adaptation in plants. Curr. Opin. Plant Biol. 2011, 14, 267–274. [Google Scholar] [CrossRef]
- Bonasio, R.; Tu, S.; Reinberg, D. Molecular signals of epigenetic states. Science 2010, 330, 612–616. [Google Scholar] [CrossRef]
- Baulcombe, D.; Dean, C. Epigenetic regulation in plant responses to the environment. Cold Spring Harb. Perspect. Biol. 2014, 6, a019471. [Google Scholar] [CrossRef]
- Crisp, P.A.; Ganguly, D.; Eichten, S.R.; Borevitz, J.O.; Pogson, B.J. Reconsidering plant memory: Intersections between stress recovery, RNA turnover, and epigenetics. Sci. Adv. 2016, 2, e1501340. [Google Scholar] [CrossRef] [PubMed]
- Mauch-Mani, B.; Baccelli, I.; Luna, E.; Flors, V. Defense priming: An adaptive part of induced resistance. Annu. Rev. Plant Biol. 2017, 68, 485–512. [Google Scholar] [CrossRef] [PubMed]
- Lämke, J.; Bäurle, I. Epigenetic and chromatin-based mechanisms in environmental stress adaptation and stress memory in plants. Genome Biol. 2017, 18, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bäurle, I.; Trindade, I. Chromatin regulation of somatic abiotic stress memory. J. Exp. Bot. 2020, 71, 5269–5279. [Google Scholar] [CrossRef]
- Johnsen, O.; Daehlen, O.G.; Østreng, G.; Skrøppa, T.; Dæhlen, O.G. Daylength and temperature during seed production interactively affect adaptive performance of Picea abies progenies. New Phytol. 2005, 168, 589–596. [Google Scholar] [CrossRef]
- Gömöry, D.; Foffová, E.; Longauer, R.; Krajmerová, D. Memory effects associated with early-growth environment in Norway spruce and European larch. Forstwiss. Centralblatt 2014, 134, 89–97. [Google Scholar] [CrossRef]
- Grant-Downton, R.; Kourmpetli, S.; Hafidh, S.; Khatab, H.; Le Trionnaire, G.; Dickinson, H.; Twell, D. Artificial microRNAs reveal cell-specific differences in small RNA activity in pollen. Curr. Biol. 2013, 23, R599–R601. [Google Scholar] [CrossRef]
- Martinez, G.; Panda, K.; Köhler, C.; Slotkin, R.K. Silencing in sperm cells is directed by RNA movement from the surrounding nurse cell. Nat. Plants 2016, 2, 16030. [Google Scholar] [CrossRef]
- Raihan, T.; Geneve, R.L.; Perry, S.E.; Lopez, C.M.R. The regulation of plant vegetative phase transition and rejuvenation: miRNAs, a key regulator. Epigenomes 2021, 5, 24. [Google Scholar] [CrossRef]
- Álvarez-Álvarez, P.; Pizarro, C.; Barrio-Anta, M.; Cámara-Obregón, A.; Bueno, J.L.M.; Álvarez, A.; Gutiérrez, I.; Burslem, D.F.R.P. Evaluation of tree species for biomass energy production in northwest Spain. Forests 2018, 9, 160. [Google Scholar] [CrossRef] [Green Version]
- Blonskaya, L.; Sultanova, R.; Muftakhova, S.; Martynova, M.; Konashova, S.; Sabirzyanov, I.; Odintsov, G. Biological indices of Bashkir Lombardy poplar (Populus nigra L. × Populus nigra var. italica Du Roi) in urban landscapes. Bulg. J. Agric. Sci. 2019, 25 (Suppl. S2), 30–36. [Google Scholar]
- Chong, G.W.; Simonson, S.E.; Stohlgren, T.J.; Kalkhan, M.A. Biodiversity: Aspen stands have the lead, but will nonnative species take over. In Sustaining Aspen in Western Landscapes: Symposium; US Department of Agriculture, Forest Service Rocky Mountain Research Station: Fort Collins, CO, USA, 2001; p. 261. [Google Scholar]
- Nilsson, S.G.; Hedin, J.; Niklasson, M. Biodiversity and its assessment in boreal and nemoral forests. Scand. J. For. Res. 2001, 16, 10–26. [Google Scholar] [CrossRef]
- Latva-Karjanmaa, T.; Penttilä, R.; Siitonen, J. The demographic structure of European aspen (Populus tremula) populations in managed and old-growth boreal forests in eastern Finland. Can. J. For. Res. 2007, 37, 1070–1081. [Google Scholar] [CrossRef]
- Boruszewski, P.; Laskowska, A.; Jankowska, A.; Klisz, M.; Mionskowski, M. Potential areas in Poland for forestry plantation. Forests 2021, 12, 1360. [Google Scholar] [CrossRef]
- Dickmann, D.I. An overview of the genus Populus. In Poplar Culture in North America, (Part A); NRC Research Press: Ottawa, ON, Canada, 2001; 397p. [Google Scholar]
- Heimann, L.; Horst, I.; Perduns, R.; Dreesen, B.; Offermann, S.; Peterhansel, C. A common histone modification code on c4 genes in maize and its conservation in sorghum and Setaria italica. Plant Physiol. 2013, 162, 456–469. [Google Scholar] [CrossRef]
- De Rigo, D.; Enescu, C.M.; Houston Durrant, T.; Caudullo, G. Populus nigra in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; Publ. Off. EU: Luxembourg, 2016. [Google Scholar]
- Blake, T.J.; Tschaplinski, T.; Eastham, A. Stomatal control of water use efficiency in poplar clones and hybrids. Can. J. Bot. 1984, 62, 1344–1351. [Google Scholar] [CrossRef]
- Isebrands, J.G.; Richardson, J. (Eds.) Poplars and Willows: Trees for Society and the Environment; FAO: Rome, Italy, 2014. [Google Scholar]
- Gilbert, O. The Ecology of Urban Habitats; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Robinson, N. The Planting Design Handbook; Routledge: London, UK, 2016. [Google Scholar]
- Eroğlu, E.; Acar, C. A Visual assessment of roadside poplar plantings in Turkey. J. Agric. Sci. 2018, 24, 185–198. [Google Scholar] [CrossRef]
- Lin, W.; Yu, Z.; Luo, Y.; He, W.; Yan, G.; Peng, C. Photoprotection differences between dominant tree species at mid- and late-successional stages in subtropical forests in different seasonal environments. Int. J. Mol. Sci. 2022, 23, 5417. [Google Scholar] [CrossRef]
- Alasalvar, C.; Grigor, J.M.; Zhang, D.; Quantick, P.C.; Shahidi, F. Comparison of volatiles, phenolics, sugars, antioxidant vitamins, and sensory quality of different colored carrot varieties. J. Agric. Food Chem. 2001, 49, 1410–1416. [Google Scholar] [CrossRef]
- Acamovic, T.; Brooker, J.D. Biochemistry of plant secondary metabolites and their effects in animals. Proc. Nutr. Soc. 2005, 64, 403–412. [Google Scholar] [CrossRef]
- Edreva, A.; Velikova, V.; Tsonev, T.; Dagnon, S.; Gürel, A.L.; Aktas, L. Stress-protective role of secondary metabolites: Diversity of functions and mechanisms. Gen. Appl. Plant. Physiol. 2008, 34, 67–78. [Google Scholar]
- Cheynier, V.; Comte, G.; Davies, K.M.; Lattanzio, V.; Martens, S. Plant phenolics: Recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Champagne, E.; Turgeon, R.; Munson, A.D.; Raymond, P. Seedling response to simulated browsing and reduced water availability: Insights for assisted migration plantations. Forests 2021, 12, 1396. [Google Scholar] [CrossRef]
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [PubMed]
- Oubida, R.W.; Gantulga, D.; Zhang, M.; Zhou, L.; Bawa, R.; Holliday, J.A. Partitioning of multivariate phenotypes using regression trees reveals complex patterns of adaptation to climate across the range of black cottonwood (Populus trichocarpa). Front. Plant Sci. 2015, 6, 181. [Google Scholar] [CrossRef] [PubMed]
- Anttonen, M.J.; Karjalainen, R.O. Environmental and genetic variation of phenolic compounds in red raspberry. J. Food Compos. Anal. 2005, 18, 759–769. [Google Scholar] [CrossRef]
- Andreotti, C.; Costa, G.; Treutter, D. Composition of phenolic compounds in pear leaves as affected by genetics, ontogenesis and the environment. Sci. Hortic. 2006, 109, 130–137. [Google Scholar] [CrossRef]
- Damestoy, T.; Brachi, B.; Moreira, X.; Jactel, H.; Plomion, C.; Castagneyrol, B. Oak genotype and phenolic compounds differently affect the performance of two insect herbivores with contrasting diet breadth. Tree Physiol. 2019, 39, 615–627. [Google Scholar] [CrossRef]
- Movahedi, A.; Yaghuti, A.A.Z.; Wei, H.; Rutland, P.; Sun, W.; Mousavi, M.; Li, D.; Zhuge, Q. Plant secondary metabolites with an overview of Populus. Int. J. Mol. Sci. 2021, 22, 6890. [Google Scholar] [CrossRef]
- Hale, B.K.; Herms, D.A.; Hansen, R.C.; Clausen, T.P.; Arnold, D. Effects of drought stress and nutrient availability on dry matter allocation, phenolic glycosides, and rapid induced resistance of poplar to two Lymantriid defoliators. J. Chem. Ecol. 2005, 31, 2601–2620. [Google Scholar] [CrossRef]
- Madritch, M.D.; Lindroth, R.L. Condensed tannins increase nitrogen recovery by trees following insect defoliation. New Phytol. 2015, 208, 410–420. [Google Scholar] [CrossRef]
- Lindroth, R.L.; Clair, S.B.S. Adaptations of quaking aspen (Populus tremuloides Michx.) for defense against herbivores. For. Ecol. Manag. 2013, 299, 14–21. [Google Scholar] [CrossRef]
- Bandau, F.; Decker, V.H.G.; Gundale, M.J.; Albrectsen, B.R. Genotypic tannin levels in Populus tremula impact the way nitrogen enrichment affects growth and allocation responses for some traits and not for others. PLoS ONE 2015, 10, e0140971. [Google Scholar] [CrossRef]
- Decker, V.H.; Bandau, F.; Gundale, M.J.; Cole, C.T.; Albrectsen, B.R. Aspen phenylpropanoid genes’ expression levels correlate with genets’ tannin richness and vary both in responses to soil nitrogen and associations with phenolic profiles. Tree Physiol. 2016, 37, 270–279. [Google Scholar] [CrossRef]
- Sobuj, N.; Virjamo, V.; Zhang, Y.; Nybakken, L.; Julkunen-Tiitto, R. Impacts of elevated temperature and CO2 concentration on growth and phenolics in the sexually dimorphic Populus tremula (L.). Environ. Exp. Bot. 2018, 146, 34–44. [Google Scholar] [CrossRef]
- Pliūra, A.; Baliuckas, V. Genetic variation in adaptive traits of progenies of Lithuanian and western European populations of Fraxinus excelsior L. Balt. For. 2007, 13, 28–38. [Google Scholar]
- Gani, M.A.; Shama, M. Phenolic Compounds. In Bioactive Compounds-Biosynthesis, Characterization and Applications; IntechOpen: London, UK, 2021. [Google Scholar]
- Onwuka, B.; Mang, B. Effects of soil temperature on some soil properties and plant growth. Adv. Plants Agric. Res. 2018, 8, 34–37. [Google Scholar] [CrossRef]
- Lipiec, J.; Doussan, C.; Nosalewicz, A.; Kondracka, K. Effect of drought and heat stresses on plant growth and yield: A review. Int. Agrophys. 2013, 27, 463–477. [Google Scholar] [CrossRef]
- Raj, S.; Bräutigam, K.; Hamanishi, E.T.; Wilkins, O.; Thomas, B.R.; Schroeder, W.; Campbell, M.M. Clone history shapes Populus drought responses. Proc. Natl. Acad. Sci. USA 2011, 108, 12521–12526. [Google Scholar] [CrossRef]
- Gudynaitė-Franckevičienė, V. Plantacinei Miškininkystei Tinkamų Tuopų Hibridų ir Klonų Ekogenetinis Plastiškumas ir Adaptacija Lietuvos Gamtinėmis Sąlygomis. Doctoral Dissertation, Aleksandro Stulginskio Universitetas, Akademija, Lithuania, 2017; 199p. [Google Scholar]
- Liu, Y.; El-Kassaby, Y.A. Phenotypic plasticity of natural Populus trichocarpa populations in response to temporally environmental change in a common garden. BMC Evol. Biol. 2019, 19, 231. [Google Scholar] [CrossRef]
- Apuli, R.-P.; Richards, T.; Rendón-Anaya, M.; Karacic, A.; Rönnberg-Wästljung, A.-C.; Ingvarsson, P.K. The genetic basis of adaptation in phenology in an introduced population of Black Cottonwood (Populus trichocarpa, Torr. & Gray). BMC Plant Biol. 2021, 21, 317. [Google Scholar] [CrossRef]
- Loehle, C. Height growth rate tradeoffs determine northern and southern range limits for trees. J. Biogeogr. 1998, 25, 735–742. [Google Scholar] [CrossRef]
- Robinson, K.M.; Ingvarsson, P.K.; Jansson, S.; Albrectsen, B.R. Genetic variation in functional traits influences arthropod community composition in aspen (Populus tremula L.). PLoS ONE 2012, 7, e37679. [Google Scholar] [CrossRef] [Green Version]
- Stevens, M.; Lindroth, R. Induced resistance in the indeterminate growth of aspen (Populus tremuloides). Oecologia 2005, 145, 297–305. [Google Scholar] [CrossRef]
- Pandey, N.; Goswami, N.; Tripathi, D.; Rai, K.K.; Rai, S.K.; Singh, S.; Pandey-Rai, S. Epigenetic control of UV-B-induced flavonoid accumulation in Artemisia annua L. Planta 2018, 249, 497–514. [Google Scholar] [CrossRef]
- D’Amelia, V.; Villano, C.; Batelli, G.; Çobanoğlu, Ö.; Carucci, F.; Melito, S.; Carputo, D. Genetic and epigenetic dynamics affecting anthocyanin biosynthesis in potato cell culture. Plant Sci. J. 2020, 298, 110597. [Google Scholar] [CrossRef]
- Pease, C.M.; Lande, R.; Bull, J.J. A model of population growth, dispersal and evolution in a changing environment. Ecology 1989, 70, 1657–1664. [Google Scholar] [CrossRef]
- Pelé, A.; Rousseau-Gueutin, M.; Chèvre, A.-M. Speciation success of polyploid plants closely relates to the regulation of meiotic recombination. Front. Plant Sci. 2018, 9, 907. [Google Scholar] [CrossRef] [PubMed]
- Uecker, H.; Hermisson, J. The role of recombination in evolutionary rescue. Genetics 2015, 202, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Hoban, S.; Bruford, M.; Jackson, J.D.; Lopes-Fernandes, M.; Heuertz, M.; Hohenlohe, P.A.; Paz-Vinas, I.; Sjögren-Gulve, P.; Segelbacher, G.; Vernesi, C.; et al. Genetic diversity targets and indicators in the CBD post-2020 Global Biodiversity Framework must be improved. Biol. Conserv. 2020, 248, 108654. [Google Scholar] [CrossRef]
- Ali, A.; Johnson, D.L. Heritability estimates for winter hardiness in lentil under natural and controlled conditions. Plant Breed. 2000, 119, 283–285. [Google Scholar] [CrossRef]
- Dillen, S.Y.; Marron, N.; Bastien, C.; Ricciotti, L.; Salani, F.; Sabatti, M.; Pinel, M.P.; Rae, A.M.; Taylor, G.; Ceulemans, R. Effects of environment and progeny on biomass estimations of five hybrid poplar families grown at three contrasting sites across Europe. For. Ecol. Manag. 2007, 252, 12–23. [Google Scholar] [CrossRef]
- Adler, A.; Karacic, A.; Wästljung, A.-C.R.; Johansson, U.; Liepins, K.; Gradeckas, A.; Christersson, L. Variation of growth and phenology traits in poplars planted in clonal trials in Northern Europe—implications for breeding. BioEnergy Res. 2021, 14, 426–444. [Google Scholar] [CrossRef]
- Johnsen, Ø. Phenotypic changes in progenies of northern clones of Picea abies (L) Karst. grown in a southern seed orchard. Scand. J. For. Res. 1989, 4, 317–330. [Google Scholar] [CrossRef]
- Fujimoto, R.; Sasaki, T.; Ishikawa, R.; Osabe, K.; Kawanabe, T.; Dennis, E.S. Molecular mechanisms of epigenetic variation in plants. Int. J. Mol. Sci. 2012, 13, 9900–9922. [Google Scholar] [CrossRef]
- Zeng, X.; Durka, W.; Welk, E.; Fischer, M. Heritability of early growth traits and their plasticity in 14 woody species of Chinese subtropical forest. J. Plant Ecol. 2017, 10, 222–231. [Google Scholar] [CrossRef]
- Burggren, W. Epigenetic inheritance and its role in evolutionary biology: Re-evaluation and new perspectives. Biology 2016, 5, 24. [Google Scholar] [CrossRef]
- Kumar, S.; Singh, A.K.; Mohapatra, T. Epigenetics: History, present status and future perspective. Indian J. Genet. Plant Breed 2017, 77, 445–463. [Google Scholar] [CrossRef]
- Latzel, V.; González, A.P.R.; Rosenthal, J. Epigenetic memory as a basis for intelligent behavior in clonal plants. Front. Plant Sci. 2016, 7, 1354. [Google Scholar] [CrossRef]
- Ragaee, S.; Abdel-Aal, E.M.; Maher, N. Antioxidant activity and nutrient composition of selected cereals for food use. Food Chem. 2006, 95, 32–38. [Google Scholar] [CrossRef]
- SAS Institute. SAS®9.4 TS Level 1M4, X64_7PRO Platform; SAS Institute Inc.: Cary, NC, USA, 2020. [Google Scholar]
- Swiger, L.A.; Harvey, W.R.; Everson, D.O.; Gregory, K.E. The variance of intraclass correlation involving groups with one observation. Biometrics 1964, 20, 818. [Google Scholar] [CrossRef]
- Becker, W.A. Manual of Procedures in Quantitative Genetics, 4th ed.; Academic Enterprises: Pullman, WA, USA, 1984; 190p. [Google Scholar]
- Falconer, D.S.; Mackay, T.F.; Frankham, R. Introduction to quantitative genetics. 4th ed. Trends Genet. 1996, 12, 280. [Google Scholar]
- Falconer, D.S. Introduction to Quantitative Genetics, 3rd ed.; Burnt Mill: Harlow, UK; Wiley: New York, NY, USA, 1989; p. 448. [Google Scholar]
- Wricke, G. Über eine Methode zur Erfassung der okologischen Streubreite in Feldverzuchen. Z. Pflanzenzuchtg 1962, 47, 92–96. [Google Scholar]
- Shukla, G.K. Some statistical aspects of partitioning genotype-environmental components of variability. Heredity 1972, 29, 237–245. [Google Scholar] [CrossRef]
- Danell, Ö. Arbetsgång vid bearbetning av contortaförsök; Inst. For For. Improve. Arbetrapport: Uppsala, Sweden, 1988; Volume 219. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect | Num. DF | Den. DF | F Value | Prob F | p * |
|---|---|---|---|---|---|
| Growing conditions in Phytotron | 1 | 114 | 241.49 | <0.0001 | *** |
| Rooting conditions in Phytotron | 2 | 114 | 2.35 | 0.1003 | . |
| Interaction rooting × growing conditions | 2 | 114 | 88.09 | <0.0001 | *** |
| Interaction rooting conditions × clone | 18 | 114 | 194.87 | <0.0001 | *** |
| Interaction growing conditions × clone | 9 | 114 | 93.43 | <0.0001 | *** |
| Interaction rooting × growing conditions × clone | 15 | 114 | 125.53 | <0.0001 | *** |
| Effect | Num. DF | Den. DF | F Value | Prob F | p * |
|---|---|---|---|---|---|
| Height 2021 | |||||
| Rooting–growing conditions in Phytotron | 5 | 583 | 0.74 | 0.5941 | . |
| Hybrid | 4 | 583 | 0.62 | 0.6458 | . |
| Interaction hybrid × rooting–growing conditions | 20 | 583 | 2.23 | 0.0017 | ** |
| Height increment 2020 | |||||
| Rooting–growing conditions in Phytotron | 5 | 546 | 9.2 | <0.0001 | *** |
| Hybrid | 4 | 546 | 0.92 | 0.4537 | . |
| Interaction hybrid × rooting–growing conditions | 20 | 546 | 2.08 | 0.0041 | ** |
| Height increment 2021 | |||||
| Rooting–growing conditions in Phytotron | 5 | 550 | 1.56 | 0.1702 | . |
| Hybrid | 4 | 550 | 2.79 | 0.0257 | * |
| Interaction hybrid × rooting–growing conditions | 20 | 550 | 1.16 | 0.2843 | . |
| Diameter 2021 | |||||
| Rooting–growing conditions in Phytotron | 5 | 583 | 1.21 | 0.3042 | . |
| Hybrid | 4 | 583 | 2.41 | 0.0483 | * |
| Interaction hybrid × rooting–growing conditions | 20 | 583 | 2.41 | 0.0006 | *** |
| Total phenolic compounds | |||||
| Rooting–growing conditions in Phytotron | 2 | 141 | 0.52 | 0.5968 | . |
| Hybrid | 4 | 141 | 19.01 | <0.0001 | *** |
| Interaction hybrid × rooting–growing conditions | 8 | 141 | 6.29 | <0.0001 | *** |
| Poplar Hybrid * | Hybrid Lsmean Deviation at Different Conditions | Phenotypic Plasticity | Wricke Ecovalence | Shukla Stability Variance | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CR + CG * | CR + WG | WR + CG | WR + WG | HR + CG | HR + WG | CR + CG/CR + WG | CR + CG/WR + CG | CR + CG/WR + WG | CR + CG/HR + CG | CR + CG/HR + WG | CR + WG/WR + CG | CR + WG/WR + CG | CR + WG/ WR + WG | CR + WG/HR + CG | WR + CG/ WR + WG | WR + CG/HR + CG | WR + CG/HR + WG | WR + WG/HR + CG | WR + WG/ HR + WG | HR + CG/HR + WG | Variance | F | p | ||
| Total phenolic compounds | |||||||||||||||||||||||||
| B × T | −6.0 | −1.3 | −3.7 | −2.5 | −2.7 | −1.6 | −4.7 | −2.3 | −3.5 | −3.3 | −4.4 | 2.4 | 1.2 | 1.4 | 1.4 | −1.2 | −1.0 | −2.1 | 0.2 | −0.9 | −1.1 | 14.6 | −2.5 | −3.0 | . |
| D × N | −1.2 | −2.8 | −0.8 | −2.3 | −0.4 | 2.0 | 1.5 | −0.4 | 1.1 | −0.8 | −3.2 | −1.9 | −0.4 | −2.3 | −2.3 | 1.5 | −0.4 | −2.8 | −1.9 | −4.3 | −2.4 | 14.2 | −2.6 | −4.9 | . |
| D × T | 1.5 | 5.8 | −2.3 | −2.6 | 5.4 | −2.1 | −4.3 | 3.7 | 4.1 | −3.9 | 3.6 | 8.1 | 8.4 | 0.4 | 0.4 | 0.3 | −7.7 | −0.1 | −8.0 | −0.5 | 7.5 | 76.2 | 18.1 | 21.6 | 0 |
| M × T | −2.2 | 2.3 | 0.4 | 1.9 | −2.7 | −0.1 | −4.5 | −2.6 | −4.1 | 0.6 | −2.0 | 1.9 | 0.4 | 5.0 | 5.0 | −1.5 | 3.1 | 0.5 | 4.6 | 2.0 | −2.6 | 21.1 | −0.3 | −0.4 | . |
| T × T | 11.0 | −5.3 | 12.8 | 13.6 | 1.5 | −0.2 | 16.3 | −1.8 | −2.5 | 9.6 | 11.3 | −18.1 | −18.9 | −6.8 | −6.8 | −0.8 | 11.3 | 13.0 | 12.1 | 13.8 | 1.7 | 314.8 | 97.6 | 63.7 | 0 |
| Tree height in 2021 | |||||||||||||||||||||||||
| B × T | −0.4 | 0.5 | −0.3 | 0.1 | −0.1 | −0.2 | −0.9 | −0.1 | −0.5 | −0.3 | −0.2 | 0.8 | 0.4 | 0.6 | 0.6 | −0.4 | −0.2 | −0.1 | 0.2 | 0.3 | 0.1 | 0.5 | 0.6 | 37.3 | 0 |
| D × N | 0.2 | −0.1 | 0.2 | −0.3 | 0.1 | −0.3 | 0.4 | 0.0 | 0.5 | 0.2 | 0.5 | −0.4 | 0.1 | −0.2 | −0.2 | 0.5 | 0.2 | 0.5 | −0.3 | 0.0 | 0.3 | 0.3 | 0.4 | 20.6 | 0 |
| D × T | 0.2 | 0.1 | 0.3 | −0.1 | 0.0 | −0.1 | 0.1 | −0.1 | 0.3 | 0.2 | 0.4 | −0.2 | 0.2 | 0.1 | 0.1 | 0.4 | 0.3 | 0.4 | −0.1 | 0.1 | 0.1 | 0.1 | 0.2 | 10.9 | 0.009 |
| M × T | −0.3 | −0.2 | −0.1 | 0.0 | 0.1 | 0.1 | −0.2 | −0.2 | −0.4 | −0.5 | −0.4 | 0.0 | −0.2 | −0.3 | −0.3 | −0.2 | −0.3 | −0.2 | −0.1 | −0.1 | 0.0 | 0.2 | 0.3 | 12.4 | 0.001 |
| T × T | −0.1 | 0.0 | −0.2 | 0.2 | −0.2 | 0.4 | 0.0 | 0.1 | −0.2 | 0.1 | −0.5 | 0.2 | −0.2 | 0.1 | 0.1 | −0.4 | 0.0 | −0.6 | 0.3 | −0.2 | −0.5 | 0.3 | 0.3 | 18.8 | 0 |
| Trait | Treatment * | Trait Mean ± se | Clonal Variance Component, ± se | p * | Clonal Variance Component, % ± se |
|---|---|---|---|---|---|
| Total phenolic compounds | CR + CG | 18.48 ± 0.89 | 22.29 ± 11.32 | * | 95.33 ± 48.44 |
| CR + WG | 16.29 ± 0.71 | 15.51 ± 7.46 | * | 94.04 ± 45.27 | |
| WR + CG | 18.96 ± 0.77 | 17.44 ± 8.76 | * | 98.63 ± 49.54 | |
| WR + WG | 15.44 ± 1.39 | 62.76 ± 29.63 | * | 99.55 ± 46.99 | |
| HR + CG | 17.19 ± 0.76 | 18.80 ± 8.88 | * | 99.38 ± 46.94 | |
| HR + WG | 16.38 ± 1.13 | 36.91 ± 18.52 | * | 98.93 ± 49.64 |
| Trait | Treatment * | CVG, % | Hi2 ± se | Hm2 ± se | CVF, % |
|---|---|---|---|---|---|
| Height | CR + CG | 15.1 | 0.11 ± 0.09 | 0.38 ± 0.11 | 45.35 |
| CR + WG | 8.2 | 0.04 ± 0.07 | 0.20 ± 0.09 | 41.01 | |
| WR + CG | 6.6 | 0.02 ± 0.07 | 0.10 ± 0.09 | 47.01 | |
| WR + WG | 21.3 | 0.22 ± 0.10 | 0.61 ± 0.09 | 44.17 | |
| HR + CG | 15.8 | 0.11 + 0.12 | 0.32 ± 0.13 | 47.14 | |
| HR + WG | 19.6 | 0.11 ± 0.14 | 0.29 ± 0.15 | 59.95 | |
| Height increment | CR + CG | 46.8 | 0.01 ± 0.08 | 0.03 ± 0.08 | 566.77 |
| CR + WG | 212.5 | 0.09 ± 0.08 | 0.37 ± 0.00 | 691.42 | |
| WR + CG | 0.0 | 0.00 ± 0.07 | 0.00 ± 0.07 | 305.95 | |
| WR + WG | 81.1 | 0.17 ± 0.10 | 0.50 ± 0.07 | 195.98 | |
| HR + CG | 227.3 | 0.05 ± 0.11 | 0.17 ± 0.13 | 974.54 | |
| HR + WG | 107.9 | 0.19 ± 0.15 | 0.43 ± 0.14 | 245.45 | |
| Diameter | CR + CG | 17.6 | 0.15 ± 0.10 | 0.46 ± 0.11 | 45.67 |
| CR + WG | 13.2 | 0.09 ± 0.07 | 0.37 ± 0.10 | 45.26 | |
| WR + CG | 5.1 | 0.01 ± 0.07 | 0.05 ± 0.08 | 49.04 | |
| WR + WG | 33.0 | 0.42 ± 0.11 | 0.80 ± 0.06 | 48.9 | |
| HR + CG | 17.0 | 0.14 ± 0.12 | 0.36 ± 0.13 | 46.17 | |
| HR + WG | 21.7 | 0.16 ± 0.14 | 0.40 ± 0.14 | 53.02 | |
| Total phenolic compounds | CR + CG | 25.5 | 0.95 ± 0.03 | 0.98 ± 0.01 | 25.19 |
| CR + WG | 24.1 | 0.94 ± 0.03 | 0.98 ± 0.01 | 24.1 | |
| WR + CG | 22 | 0.99 ± 0.01 | 0.99 ± 0.00 | 21.32 | |
| WR + WG | 51.3 | 1.00 ± 0.00 | 1.00 ± 0.00 | 49.62 | |
| HR + CG | 25.2 | 0.99 ± 0.00 | 1.00 ± 0.00 | 24.42 | |
| HR + WG | 37 | 0.99 ± 0.01 | 1.00 ± 0.00 | 35.82 |
| Crossing Type | Hybrid Abbreviation | Crossing Combination | Clone Number or Cultivar Name Abbreviation |
|---|---|---|---|
| Inter-specific | D × N | P. deltoides × P. nigra | Gr-Comp, Gr-Xe-3, Nyd-Elle, UK-AgatF, UK-Robus, UK-Spitk |
| D × T | P. deltoides × P. trichocarpa | Isl-15, UK-Boela, UK-Donk | |
| M × T | P. maximowiczii × P. trichocarpa | SvSFPo2, SvSFPo6, SvSFPo7, UK-Andro | |
| B × T | P. balsamifera × P. trichocarpa | SvSFPo1, SvSFPo4, SvSFPo13 | |
| Intra-specific | T × T | P. trichocarpa × P. trichocarpa | SvSFPo14, SvSFPo15, UK-FrPau, SvSFPo9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gudynaitė-Franckevičienė, V.; Pliūra, A. Performance and Genetic Parameters of Poplar Hybrids and Clones in a Field Trial Are Modified by Contrasting Environmental Conditions during the Vegetative Propagation Phase. Plants 2022, 11, 2401. https://doi.org/10.3390/plants11182401
Gudynaitė-Franckevičienė V, Pliūra A. Performance and Genetic Parameters of Poplar Hybrids and Clones in a Field Trial Are Modified by Contrasting Environmental Conditions during the Vegetative Propagation Phase. Plants. 2022; 11(18):2401. https://doi.org/10.3390/plants11182401
Chicago/Turabian StyleGudynaitė-Franckevičienė, Valda, and Alfas Pliūra. 2022. "Performance and Genetic Parameters of Poplar Hybrids and Clones in a Field Trial Are Modified by Contrasting Environmental Conditions during the Vegetative Propagation Phase" Plants 11, no. 18: 2401. https://doi.org/10.3390/plants11182401