
The Role of Alternative Electron Pathways for Effectiveness of Photosynthetic Performance of Arabidopsis thaliana, Wt and Lut2, under Low Temperature and High Light Intensity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
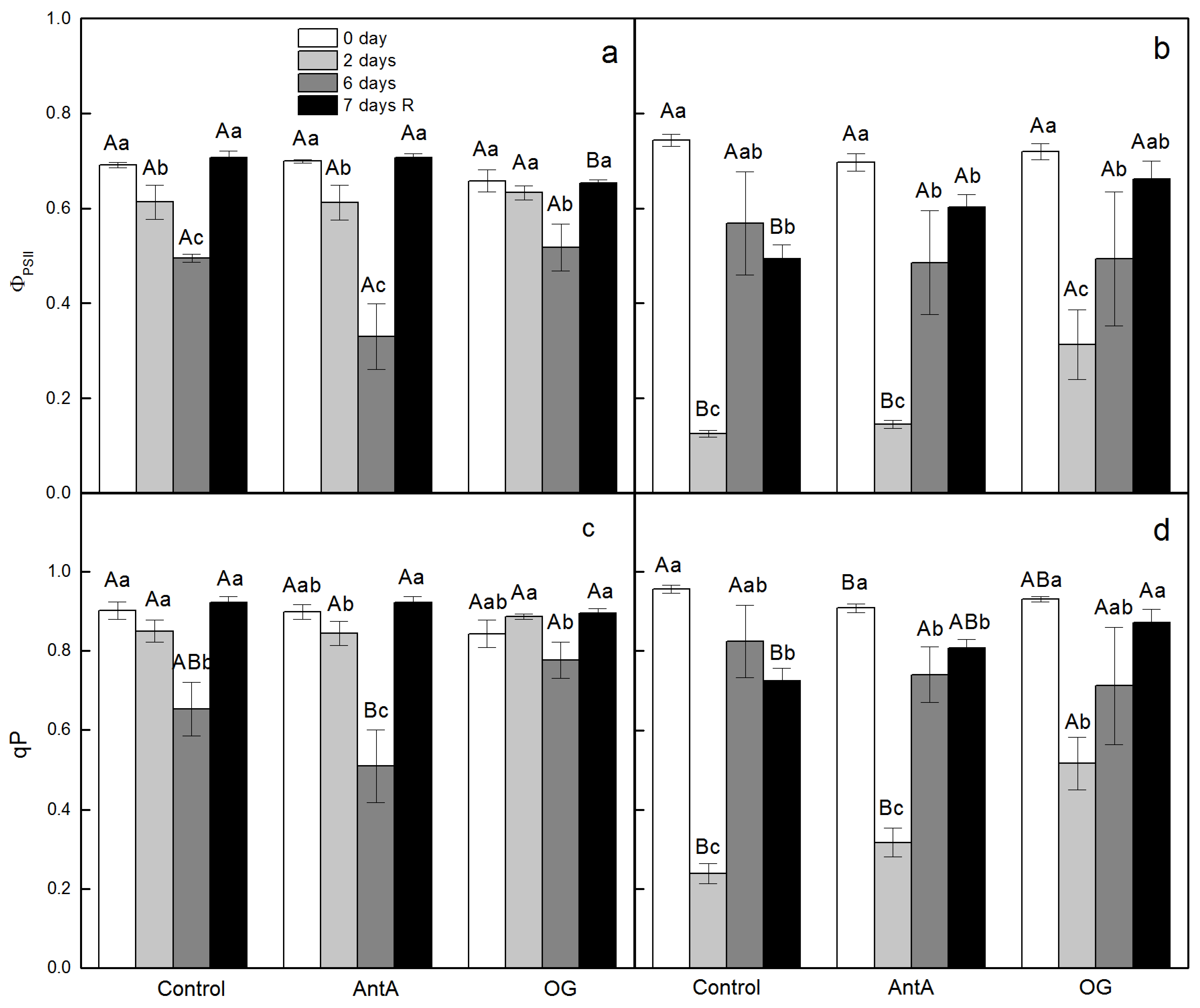
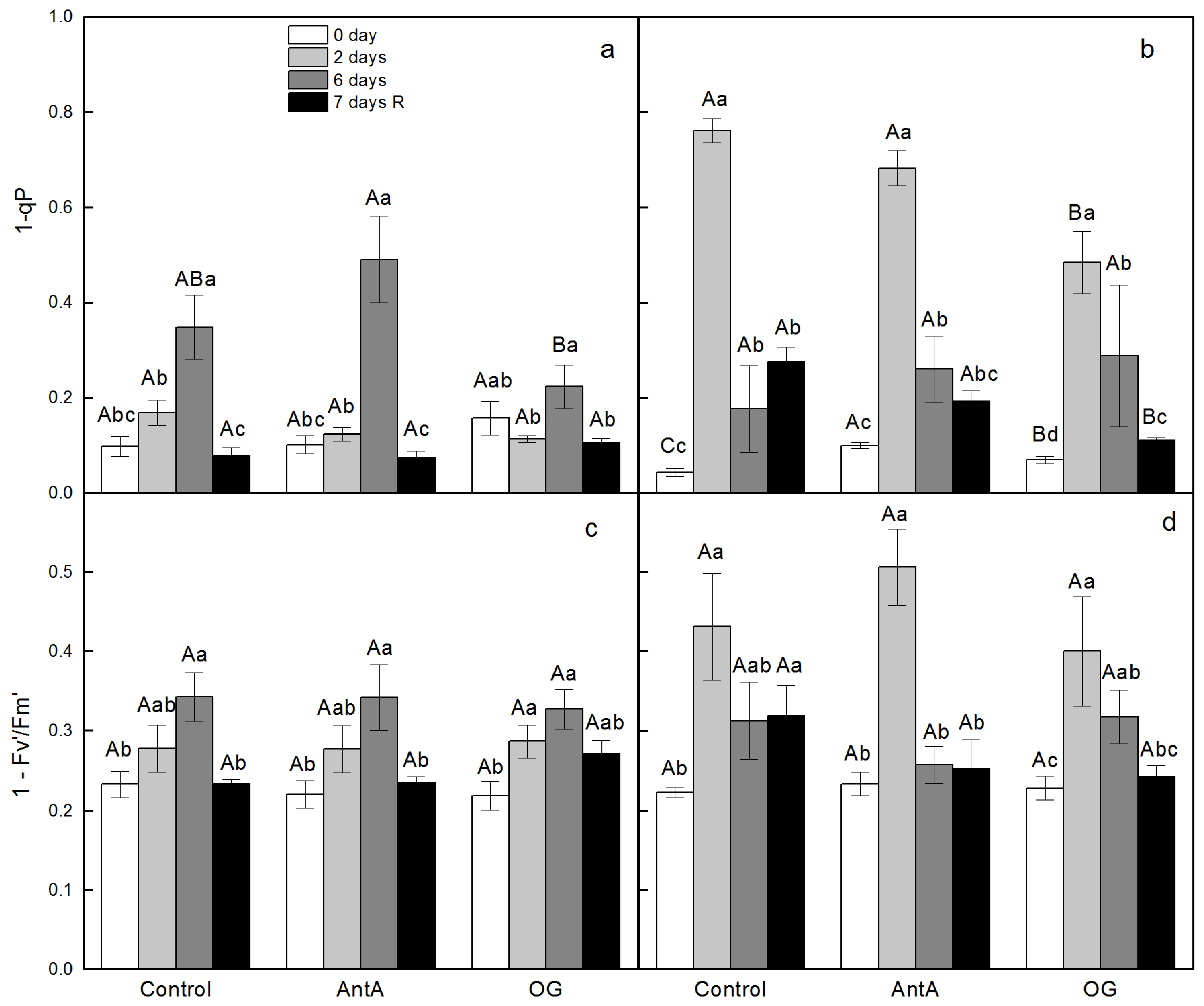
2.1. Effect of Specific Electron Transport Inhibitors on PSII Activity in Leaf Discs of wt and lut2 Plants That Were Treated with Two Stress Factors and Recovered
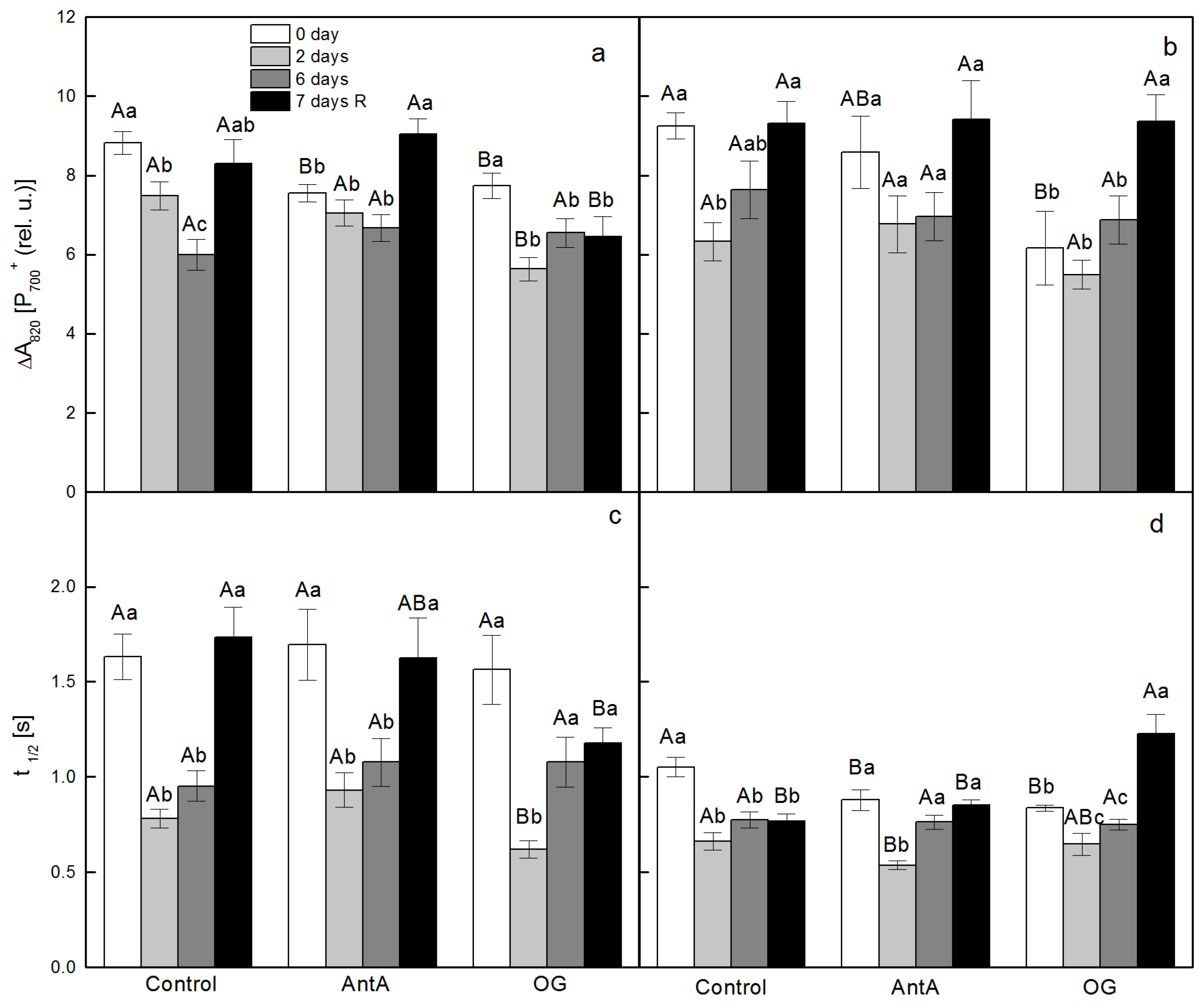
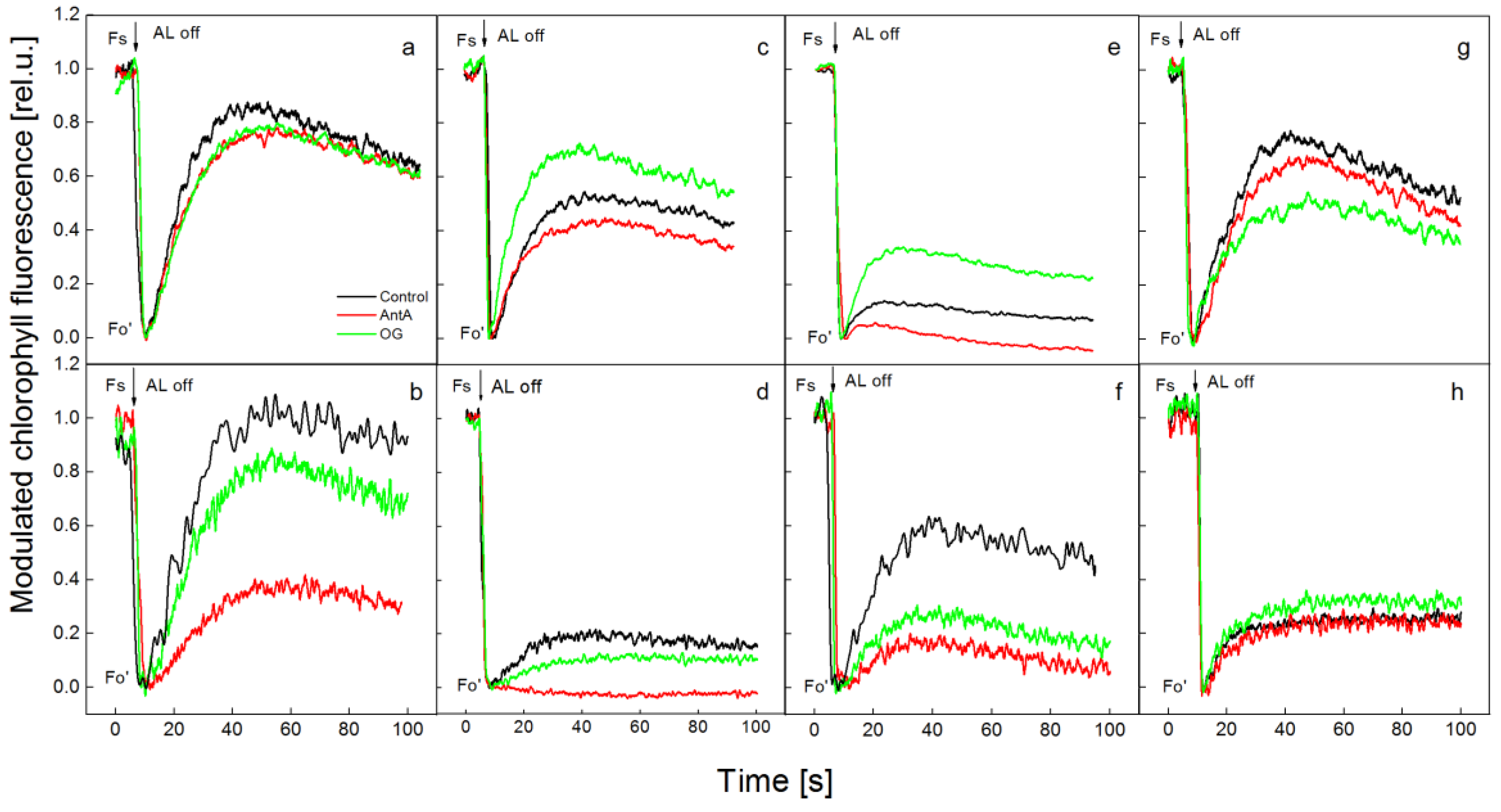
2.2. Effect of Specific Electron Transport Inhibitors on the Redox State of PSI after Treatment of wt and lut2 Plants with Two Stress Factors
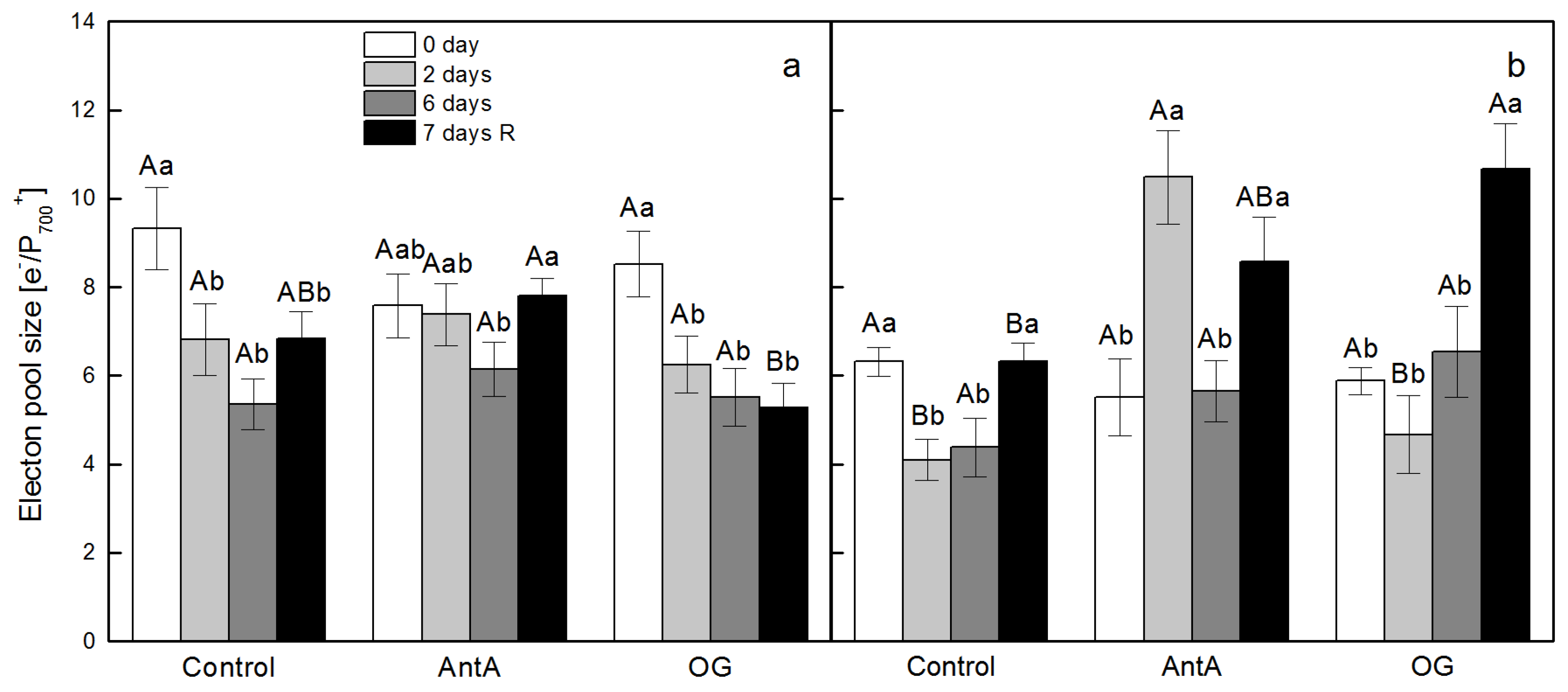
2.3. Intersystem Electron Pool Size and PQ Pool Reduction
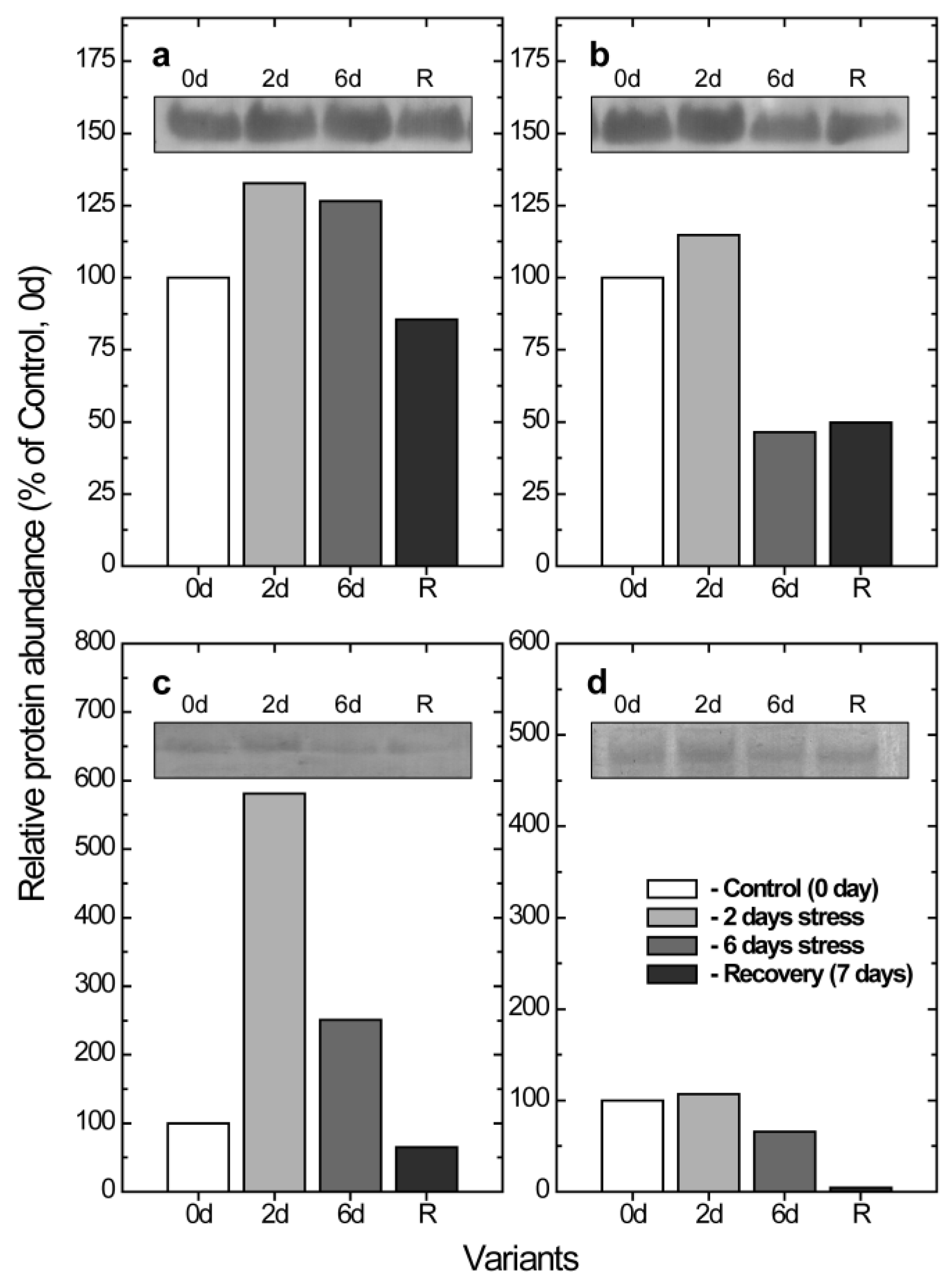
2.4. Alterations of the Abundance of PGR5 and PTOX as a Result of a Low Temperature and High Light Intensity Treatment for Different Periods of Time
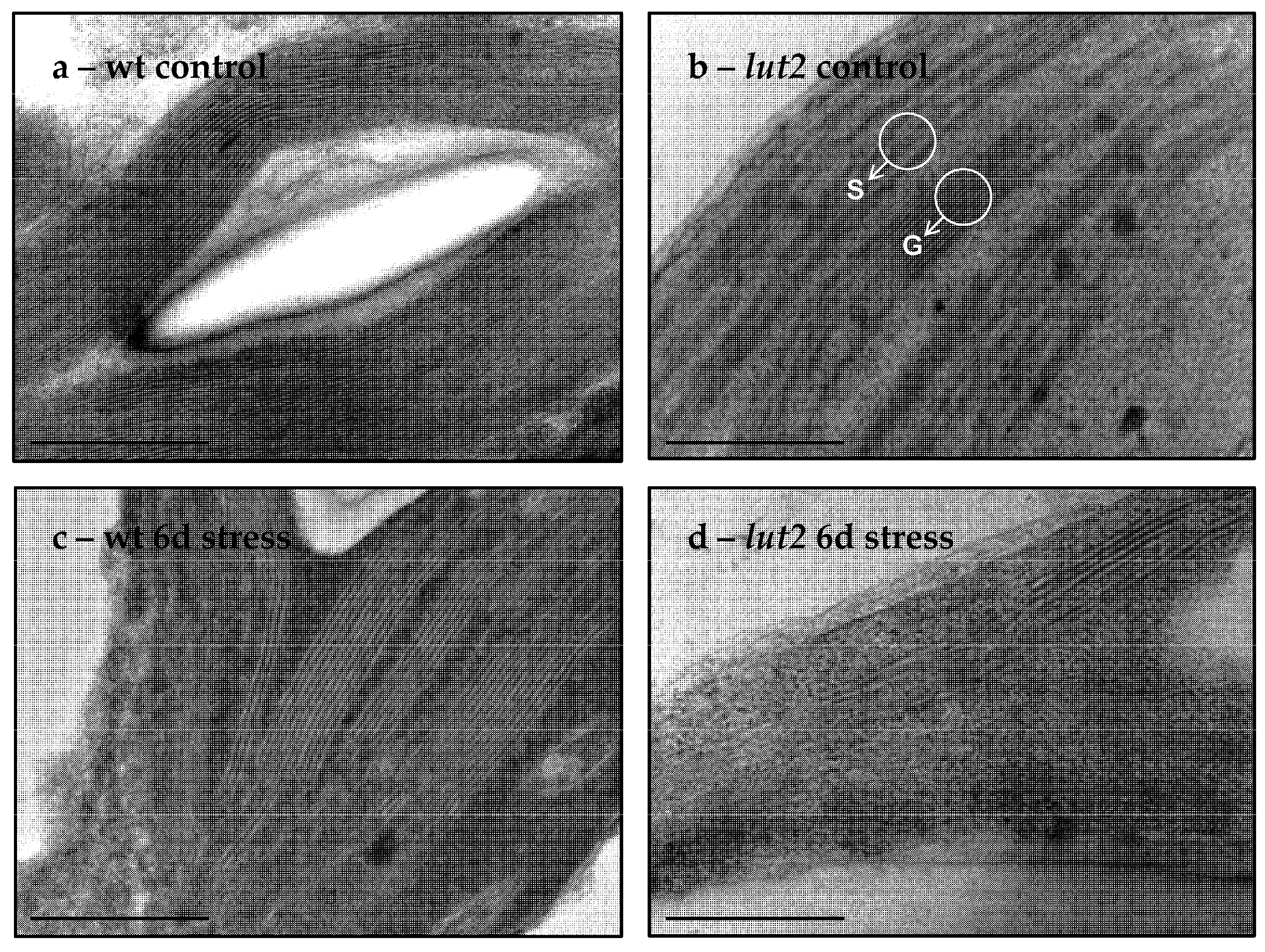
2.5. Chloroplast Ultrastructure in wt and lut2 Mutants
3. Discussion
4. Materials and Methods
4.1. Plant Growth Conditions
4.2. Application of Specific Inhibitors
4.3. Pulse-Amplitude-Modulated Chlorophyll Fluorescence Levels
4.4. Redox State of P700
4.5. Electron Microscopy
4.6. SDS-PAGE Electrophoresis and Western Blot
4.7. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huner, N.P.A.; Maxwell, D.P.; Gray, G.R.; Savitch, L.V.; Krol, M.; Ivanov, A.G.; Falk, S. Sensing environmental change: PSII excitation pressure and redox signaling. Physiol. Plant. 1996, 98, 358–364. [Google Scholar] [CrossRef]
- Huner, N.P.A.; Öquist, G.; Sarhan, F. Energy balance and acclimation to light and cold. Trends Plant Sci. 1998, 3, 224–230. [Google Scholar] [CrossRef]
- Aro, E.M.; Virgin, I.; Andersson, B. Photoinhibition of photosystem II. Inactivation, protein damage and turnover. Biochim. Biophys. Acta-Bioenerg. 1993, 1143, 113–134. [Google Scholar] [CrossRef]
- Long, S.P.; Humphries, S.; Falkowski, P.G. Photoinhibition of photosynthesis in nature. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1994, 45, 633–662. [Google Scholar] [CrossRef]
- Sonoike, K.; Kamo, M.; Hihara, Y.; Hiyama, T.; Enami, I. The mechanism of the degradation of psaB gene product, one of the photosynthetic reaction center subunits of photosystem I, upon photoinhibition. Photosynth. Res. 1997, 53, 55–63. [Google Scholar] [CrossRef]
- Ivanov, A.G.; Morgan, R.M.; Gray, G.R.; Velitchkova, M.Y.; Huner, N.P.A. Temperature/ light dependence development of selective resistance to photoinhibition of photosystem I. FEBS Lett. 1998, 430, 288–292. [Google Scholar] [CrossRef]
- Horton, P.; Ruban, A.V.; Walters, R.G. Regulation of light harvesting in green plants. Annu. Rev. Plant Phys. 1996, 47, 655–684. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W. Photoprotection and other responses of plants to high light stress. Annu. Rev. Plant Phys. 1992, 43, 599–626. [Google Scholar] [CrossRef]
- Ensminger, I.; Busch, F.; Huner, N.P.A. Photostasis and cold acclimation: Sensing low temperature through photosynthesis. Physiol. Plant. 2006, 126, 28–44. [Google Scholar] [CrossRef]
- Ivanov, A.G.; Sane, P.V.; Hurry, V.; Öquist, G.; Huner, N.P.A. Photosystem II reaction center quenching: Mechanisms and physiological role. Photosynth. Res. 2008, 98, 565–574. [Google Scholar] [CrossRef]
- Ivanov, A.G.; Rosso, D.; Savitch, L.V.; Stachula, P.; Rosembert, M.; Öquist, G.; Hurry, V.; Huner, N.P.A. Implications of alternative electron sinks in increased resistance of PSII and PSI photochemistry to high light stress in cold acclimated Arabidopsis thaliana. Photosynth. Res. 2012, 113, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Miyake, C.; Horiguchi, S.; Makino, A.; Shinzaki, Y.; Yamamoto, H.; Tomizawa, K.-i. Effects of light intensity on cyclic electron flow around PSI and its relationship to non-photochemical quenching of Chl fluorescence in tobacco leaves. Plant Cell Physiol. 2005, 46, 1819–1830. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Yang, Y.J.; Zhang, S.B. Specific roles of cyclic electron flow around photosystem I in photosynthetic regulation in immature and mature leaves. J. Plant Physiol. 2017, 209, 76–83. [Google Scholar]
- Peltier, G.; Cournac, L. Chlororespiration. Annu. Rev. Plant Biol. 2002, 53, 523–550. [Google Scholar] [CrossRef]
- Bukhov, N.; Carpentier, R. Alternative Photosystem I-driven electron transport routes: Mechanisms and functions. Photosynth. Res. 2004, 82, 17–33. [Google Scholar] [CrossRef]
- Munekage, Y.; Hojo, M.; Meurer, J.; Endo, T.; Tasaka, M.; Shikanai, T. PGR5 is involved in cyclic electron flow around photosystem I and is essential for photoprotection in Arabidopsis. Cell 2002, 110, 361–371. [Google Scholar] [CrossRef]
- Takahashi, S.; Milward, S.E.; Fan, D.Y.; Chow, W.S.; Badger, M.R. How does cyclic electron flow alleviate photoinhibition in Arabidopsis? Plant Physiol. 2009, 149, 1560–1567. [Google Scholar] [CrossRef]
- Miyake, C. Alternative electron flows (water-water cycle and cyclic electron flow around PSI) in photosynthesis: Molecular mechanisms and physiological functions. Plant Cell Physiol. 2010, 51, 951–963. [Google Scholar] [CrossRef]
- Peltier, G.; Aro, E.-M.; Shikanai, T. NDH-1 and NDH-2 plastoquinone reductases in oxygenic photosynthesis. Annu. Rev. Plant Biol. 2016, 67, 55–80. [Google Scholar] [CrossRef]
- Yamori, W.; Sakata, N.; Suzuki, Y.; Shikanai, T.; Makino, A. Cyclic electron flow around photosystem I via chloroplast NAD(P)H dehydrogenase (NDH) complex performs a significant physiological role during photosynthesis and plant growth at low temperature in rice. Plant J. 2011, 68, 966–976. [Google Scholar] [CrossRef]
- Nishikawa, Y.; Yamamoto, H.; Okegawa, Y.; Wada, S.; Sato, N.; Taira, Y.; Sugimoto, K.; Makino, A.; Shikanai, T. PGR5-dependent cyclic electron transport around PSI contributes to the redox homeostasis in chloroplasts rather than CO2 fixation and biomass production in rice. Plant Cell Physiol. 2012, 53, 2117–2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munekage, Y.; Hashimoto, M.; Miyake, C.; Tomizawa, K.-I.; Endo, T.; Tasaka, M.; Shikanai, T. Cyclic electron flow around photosystem I is essential for photosynthesis. Nature 2004, 429, 579–582. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zhang, S.B.; Cao, K.F. Cyclic Electron Flow Plays an Important Role in Photoprotection of Tropical Trees Illuminated at Temporal Chilling Temperature. Plant Cell Physiol. 2011, 52, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Suorsa, M.; Järvi, S.; Grieco, M.; Nurmi, M. PROTON GRADIENT REGULATION5 is essential for proper acclimation of Arabidopsis photosystem I to naturally and artificially fluctuating light conditions. Plant Cell 2012, 24, 2934–2948. [Google Scholar] [CrossRef]
- Tikkanen, M.; Rantala, S.; Aro, E.-M. Electron flow from PSII to PSI under high light is controlled by PGR5 but not by PSBS. Front. Plant Sci. 2015, 6, 521. [Google Scholar] [CrossRef]
- Chaux, F.; Peltier, G.; Johnson, X. A security network in PSI photoprotection: Regulation of photosynthetic control, NPQ and O2 photoreduction by cyclic electron flow. Front. Plant Sci. 2015, 6, 875. [Google Scholar] [CrossRef]
- Lu, J.; Yin, Z.; Yang, L.X.; Wang, F.; Qi, M.; Li, T.; Liu, T. Cyclic electron flow modulate the linear electron flow and reactive oxygen species in tomato leaves under high temperature. Plant Sci. 2020, 292, 110387. [Google Scholar] [CrossRef]
- Yamori, W.; Shikanai, T.; Makino, A. Corrigendum: Photosystem I cyclic electron flow via chloroplast NADH dehydrogenase-like complex performs a physiological role for photosynthesis at low light. Sci. Rep. 2015, 5, 15593. [Google Scholar] [CrossRef]
- Shikanai, T.; Yamamoto, H. Contribution of cyclic and pseudo-cyclic electron transport to the formation of proton motive force in chloroplasts. Mol. Plant 2017, 10, 20–29. [Google Scholar] [CrossRef]
- Toyoshima, M.; Sakata, M.; Ohnishi, K.; Tokumaru, Y.; Kato, Y.; Tokutsu, R.; Sakamoto, W.; Minagawa, J.; Matsuda, F.; Shimizu, H. Targeted proteome analysis of microalgae under high-light conditions by optimized protein extraction of photosynthetic organisms. J. Biosci. Bioeng. 2019, 127, 394–402. [Google Scholar] [CrossRef]
- Burrows, P.A.; Sazanov, L.A.; Svab, Z.; Maliga, P.; Nixon, P.J. Identification of a functional respiratory complex in chloroplasts through analysis of tobacco mutants containing disrupted plastid NDH genes. EMBO J. 2014, 17, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Shikanai, T.; Endo, T.; Hashimoto, T.; Yamada, Y.; Asada, K.; Yokota, A. Directed disruption of the tobacco ndhB gene impairs cyclic electron flow around photosystem I. Proc. Natl. Acad. Sci. USA 1998, 95, 9705–9709. [Google Scholar] [CrossRef]
- Hertle, A.P.; Blunder, T.; Wunder, T.; Pesaresi, P.; Pribil, M.; Armbruster, U.; Leister, D. PGRL1 is the elusive ferredoxin-plastoquinone reductase in photosynthetic cyclic electron flow. Mol. Cell 2013, 49, 511–523. [Google Scholar] [CrossRef] [PubMed]
- DalCorso, G.; Pesaresi, P.; Masiero, S.; Aseeva, E.; Schünemann, D.; Finazzi, G.; Joliot, P.; Barbato, R.; Leister, D.A. complex containing PGRL1 and PGR5 is involved in the switch between linear and cyclic electron flow in Arabidopsis. Cell 2008, 132, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Ueda, M.; Kuniyoshi, T.; Yamamoto, H.; Sugimoto, K.; Ishizaki, K.; Kogchi, T.; Nishimura, Y.; Shikanai, T. Composition and physiological function of the chloroplast NADH dehydrogenase-like complex in Marchantia polymorpha. Plant J. 2012, 72, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Horváth, E.M.; Peter, S.O.; Joët, T.; Rumeau, D.; Cournac, L.; Horváth, G.V.; Kavanagh, T.A.; Schäfer, C.; Peltier, G.; Medgyesy, P. Targeted inactivation of the plastid ndhB gene in tobacco results in an enhanced sensitivity of photosynthesis to moderate stomatal closure. Plant Physiol. 2000, 123, 1337–1350. [Google Scholar] [CrossRef]
- Joët, T.; Genty, B.; Josse, E.-M.; Kuntz, M.; Cournac, L.; Peltier, G. Involvement of a plastid terminal oxidase in plastoquinone oxidation as evidenced by expression of the Arabidopsis thaliana enzyme in tobacco. J Biol. Chem. 2002, 277, 31623–31630. [Google Scholar] [CrossRef]
- Rosso, D.; Ivanov, A.G.; Fu, A.; Geisler-Lee, J.; Hendrickson, L.; Geisler, M.; Stewart, G.; Krol, M.; Hurry, V.; Rodermel, S.R.; et al. IMMUTANS does not act as a stress-induced safety valve in the protection of the photosynthetic apparatus of Arabidopsis during steady-state photosynthesis. Plant Physiol. 2006, 142, 574–585. [Google Scholar] [CrossRef]
- Stepien, P.; Johnson, G.N. Contrasting responses of photosynthesis to salt stress in the glycophyte Arabidopsis thaliana and the halophyte Thellungiella halophila. Role of the plastid terminal oxidase as an alternative electron sink. Plant Physiol. 2009, 149, 1154–1165. [Google Scholar] [CrossRef]
- Savitch, L.V.; Ivanov, A.G.; Krol, M.; Sprott, D.P.; Öquist, G.; Huner, N.P.A. Regulation of energy partitioning and alternative electron transport pathways during cold acclimation of lodgepole pine is oxygen dependent. Plant Cell Physiol. 2010, 51, 1555–1570. [Google Scholar] [CrossRef]
- Krieger-Liszkay, A.; Feilke, K. The dual role of the plastid terminal oxidase PTOX: Between a protective and a pro-oxidant function. Front. Plant Sci. 2016, 6, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niyogi, K.K. Safety valves for photosynthesis. Curr. Opin. Plant Biol. 2000, 3, 455–460. [Google Scholar] [CrossRef]
- McDonald, E.; Ivanov, A.G.; Bode, R.; Maxwell, D.P.; Rodermel, S.R.; Huner, N.P.A. Flexibility in photosynthetic electron transport: The physiological role of plastoquinol terminal oxidase (PTOX). Biochim. Biophys. Acta 2011, 1807, 954–967. [Google Scholar] [CrossRef] [PubMed]
- Kambakam, S.; Bhattacharjee, U.; Petrich, J.; Rodermel, S. PTOX mediates novel pathways of electron transport in etioplasts of Arabidopsis. Mol. Plant 2016, 9, 1240–1259. [Google Scholar] [CrossRef] [PubMed]
- Bassi, R.; Pineau, B.; Dainese, P.; Marquardt, J. Carotenoidbinding proteins of photosystem I. Eur J. Biochem. 1993, 212, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Havaux, M.; Niyogi, K.K. The violaxanthin cycle protects plants from photooxidative damage by more than one mechanism. Proc. Natl. Acad. Sci. USA 1999, 96, 8762–8767. [Google Scholar] [CrossRef]
- Pogson, B.; McDonald, K.A.; Truong, M.; Britton, G.; DellaPenna, D. Arabidopsis carotenoid mutants demonstrate that lutein is not essential for photosynthesis in higher plants. Plant Cell 1996, 8, 1627–1639. [Google Scholar]
- Dall’Osto, L.; Cazzaniga, S.; North, E.; Marion-Poll, A.; Bassi, R. The Arabidopsis aba4-1 mutant reveals a specific function for neoxanthin in protection against photooxidative stress. Plant Cell 2007, 19, 1048–1064. [Google Scholar] [CrossRef]
- Jahns, P.; Holzwarth, A.R. The role of the xanthophyll cycle and of lutein in photoprotection of photosystem II. Biochim. Biophys. Acta 2012, 1817, 182–193. [Google Scholar] [CrossRef]
- Dall’Osto, L.; Lico, C.; Alric, J.; Giuliano, G.; Havaux, M.; Bassi, R. Lutein is needed for efficient chlorophyll triplet quenching in the major LHCII antenna complex of higher plants and effective photoprotection in vivo under strong light. BMC Plant Biol. 2006, 6, 32. [Google Scholar] [CrossRef]
- Popova, A.V.; Dobrev, K.; ·Velitchkova, M.; Ivanov, A.G. Differential temperature effects on dissipation of excess light energy and energy partitioning in lut2 mutant of Arabidopsis thaliana under photoinhibitory conditions. Photosynth. Res. 2019, 139, 367–385. [Google Scholar] [CrossRef] [PubMed]
- Popova, A.V.; Vladkova, R.; Borisova, P.; Georgieva, K.; Mihailova, G.; Velikova, V.; Tsonev, T.; Ivanov, A.G. Photosynthetic response of lutein-deficient mutant lut2 of Arabidopsis thaliana to low-temperature at high-light. Photosynthetica 2022, 60, 110–120. [Google Scholar] [CrossRef]
- Niyogi, K.K.; Bjorkman, O.; Grossman, A.R. The roles of specific xanthophylls in photoprotection. Proc. Natl. Acad. Sci. USA 1997, 94, 14162–14167. [Google Scholar] [CrossRef] [PubMed]
- Pogson, B.J.; Niyogi, K.K.; Björkman, O.; DellaPenna, D. Altered xanthophyll composition adversely affect chlorophyll accumulation and non-photochemical quenching in Arabidopsis mutants. Proc. Natl. Acad. Sci. USA 1998, 95, 13324–13329. [Google Scholar] [CrossRef]
- Lokstein, H.; Tian, L.; Polle, J.E.W.; DellaPenna, D. Xanthophyll biosynthetic mutants of Arabidopsis thaliana: Altered nonphotochemical quenching of chlorophyll fluorescence is due to changes in Photosystem II antenna size and stability. Biochim. Biophys. Acta-Bioenerg. 2002, 1553, 309–319. [Google Scholar] [CrossRef]
- Asada, K.; Heber, U.; Schreiber, U. Electron flow to the intersystem chain from stromal components and cyclic electron flow in maize chloroplasts, as determined in intact leaves by monitoring redox change of P700 and chlorophyll fluorescence. Plant Cell Physiol. 1993, 34, 39–50. [Google Scholar]
- Shikanai, T. Cyclic electron transport around photosystem I: Genetic approaches. Annu. Rev. Plant Biol. 2007, 58, 199–217. [Google Scholar] [CrossRef]
- Sun, X.; Wen, T. Physiological roles of plastid terminal oxidase in plant stress responses. J. Biosci. 2011, 36, 951–956. [Google Scholar] [CrossRef]
- Cournac, L.; Josse, E.-M.; Joët, T.; Rumeau, D.; Redding, K.; Kuntz, M.; Peltier, G. Flexibility in photosynthetic electron transport: A newly identified chloroplast oxidase involved in chlororespiration. Philos. Trans. R. Soc. Lond. B 2000, 355, 1447–1454. [Google Scholar] [CrossRef]
- Velitchkova, M.; Borisova, P.; Vasilev, D.; Popova, A.V. Different impact of high light on the response and recovery of wild type and lut2 mutant of Arabidopsis thaliana at low temperature. Theor. Exp. Plant Physiol. 2021, 33, 95–111. [Google Scholar] [CrossRef]
- Savitch, L.V.; Ivanov, A.G.; Gudynaite-Savitch, L.; Huner, N.P.A.; Simmonds, J. Cold stress effects on PSI photochemistry in Zea mays: Differential increase of FQR-dependent cyclic electron transport flow and functional implications. Plant Cell Physiol. 2011, 52, 1042–1054. [Google Scholar] [CrossRef] [PubMed]
- Rochaix, J.-D. 2011, Regulation of photosynthetic electron transport. Biochim. Biophys. Acta 2011, 1807, 375–383. [Google Scholar]
- Nellaepalli, S.; Kodru, S.; Raghavendra, A.S.; Subramanyam, R. Antimycin A sensitive pathway independent from PGR5 cyclic electron transfer triggers non-photochemical reduction of PQ pool and state transitions in Arabidopsis thaliana. J. Photochem. Photobiol. B Biol. 2015, 146, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Aluru, M.R.; Rodermel, S.R. Control of chloroplast redox by the IMMUTANS terminal oxidase. Physiol. Plant 2004, 120, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Aluru, M.R.; Yu, F.; Fu, A.; Rodermel, S. Arabidopsis variegation mutants: New insights into chloroplast biogenesis. J. Exp. Bot. 2006, 57, 1871–1881. [Google Scholar] [CrossRef] [PubMed]
- van Kooten, O.; Snel, J.F.H. The use of chlorophyll fluorescence nomenclature in plant stress physiology. Photosynth. Res. 1990, 25, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Hendrickson, L.; Furbank, R.T.; Chow, W.S. A simple alternative approach to assessing the fate of absorbed light energy using chlorophyll fluorescence. Photosynth. Res. 2004, 82, 73–81. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams III, W.W.; Barker, D.H.; Logan, B.A.; Bowling, D.R.; Verhoeven, A.S. Using chlorophyll fluorescence to assess the fraction of absorbed light allocated to thermal dissipation of excess excitation. Physiol. Plant. 1996, 98, 253–264. [Google Scholar] [CrossRef]
- Klughammer, C.; Schreiber, U. Analysis of light-induced absorbency changes in the near-infrared spectral region. 1. Characterization of various components in isolated chloroplasts. Z. Naturforsch. C 1991, 46, 233–244. [Google Scholar] [CrossRef]
- Losciale, P.; Oguchi, R.; Hendrickson, L.; Hope, A.B.; Corelli-Grappadelli, L.; Chow, W.S. A rapid, whole-tissue determination of the functional fraction of PSII after photoinhibition of leaves based on flash-induced P700 redox kinetics. Physiol. Plant. 2008, 132, 23–32. [Google Scholar] [CrossRef]
- Asada, K.; Heber, U.; Schreiber, U. Pool size of electrons that can be donated to P700+, as determined in intact leaves: Donation to P700+ from stromal components via the intersystem chain. Plant Cell Physiol. 1992, 33, 927–932. [Google Scholar]
- Maxwell, P.C.; Biggins, J. Role of cyclic electron transport in photosynthesis as measured by the photoinduced turnover of P700 in vivo. Biochemistry 1976, 15, 3975–3981. [Google Scholar] [CrossRef] [PubMed]
- Ravenel, J.; Peltier, G.; Havaux, M. The cyclic electron pathways around photosystem I in Chlamydomonas reinhardtii as determined in vivo by photoacoustic measurements of energy storage. Planta 1994, 193, 251–259. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Velitchkova, M.; Popova, A.V.; Faik, A.; Gerganova, M.; Ivanov, A.G. Low temperature and high light dependent dynamic photoprotective strategies in Arabidopsis thaliana. Physiol. Plant. 2020, 170, 93–108. [Google Scholar] [CrossRef]
- Ivanov, A.G.; Morgan-Kiss, R.M.; Krol, M.; Allakhverdiev, S.I.; Zanev, Y.; Sane, P.V.; Huner, N.P.A. Photoinhibition of photosystem I in a pea mutant with altered LHCII organization. J. Photochem. Photobiol. B Biol. 2015, 152, 335–346. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popova, A.V.; Stefanov, M.; Ivanov, A.G.; Velitchkova, M. The Role of Alternative Electron Pathways for Effectiveness of Photosynthetic Performance of Arabidopsis thaliana, Wt and Lut2, under Low Temperature and High Light Intensity. Plants 2022, 11, 2318. https://doi.org/10.3390/plants11172318
Popova AV, Stefanov M, Ivanov AG, Velitchkova M. The Role of Alternative Electron Pathways for Effectiveness of Photosynthetic Performance of Arabidopsis thaliana, Wt and Lut2, under Low Temperature and High Light Intensity. Plants. 2022; 11(17):2318. https://doi.org/10.3390/plants11172318
Chicago/Turabian StylePopova, Antoaneta V., Martin Stefanov, Alexander G. Ivanov, and Maya Velitchkova. 2022. "The Role of Alternative Electron Pathways for Effectiveness of Photosynthetic Performance of Arabidopsis thaliana, Wt and Lut2, under Low Temperature and High Light Intensity" Plants 11, no. 17: 2318. https://doi.org/10.3390/plants11172318