

Salt Pretreatment-Mediated Alleviation of Boron Toxicity in Safflower Cultivars: Growth, Boron Accumulation, Photochemical Activities, Antioxidant Defense Response

Abstract
:1. Introduction
2. Results and Discussion
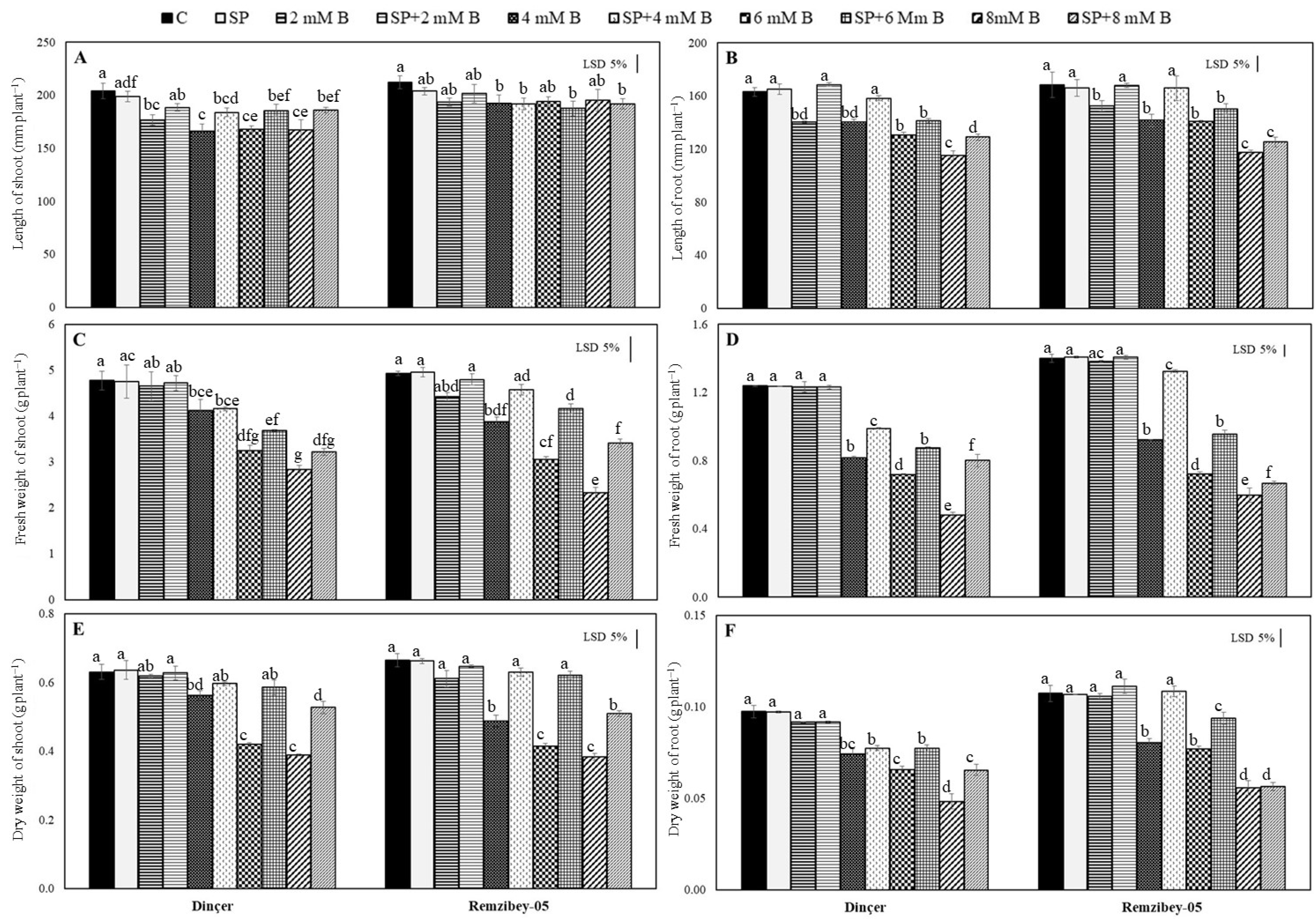
2.1. Plant Growth Response to Toxic B with or without SP
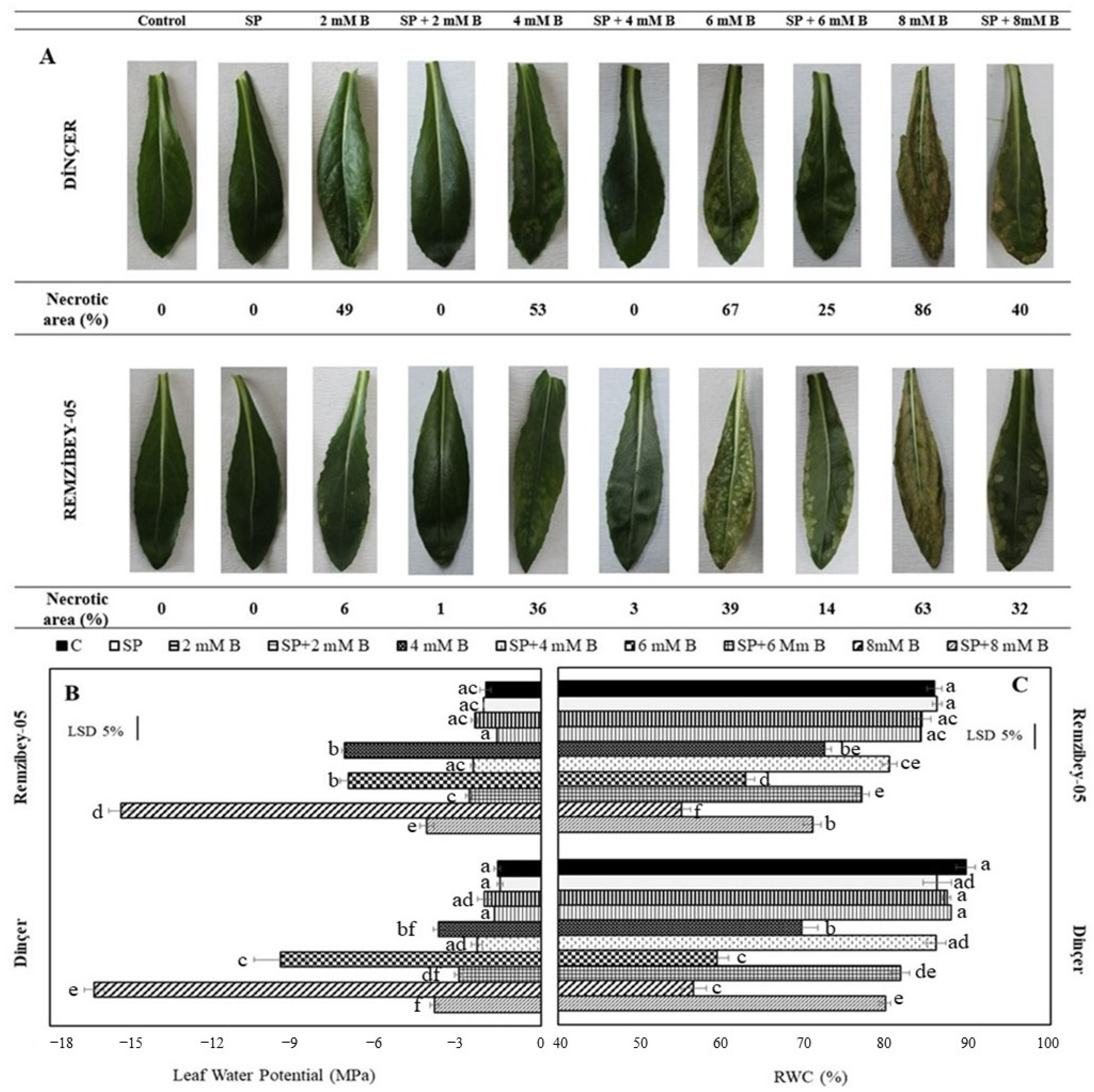
2.2. Allocation of B in Organs and Phytoremediation Potential of Safflower under B Toxicity with or without SP
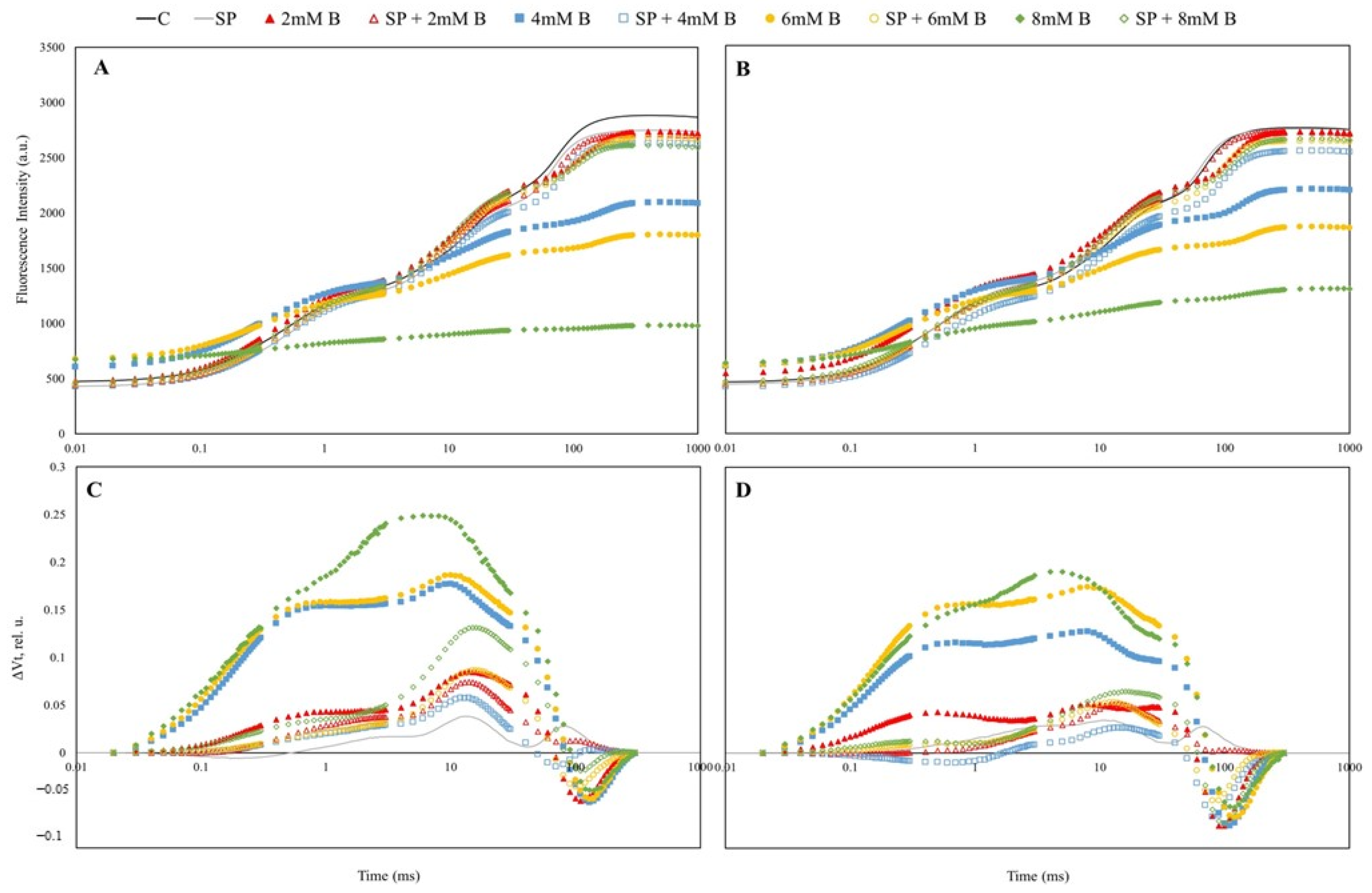
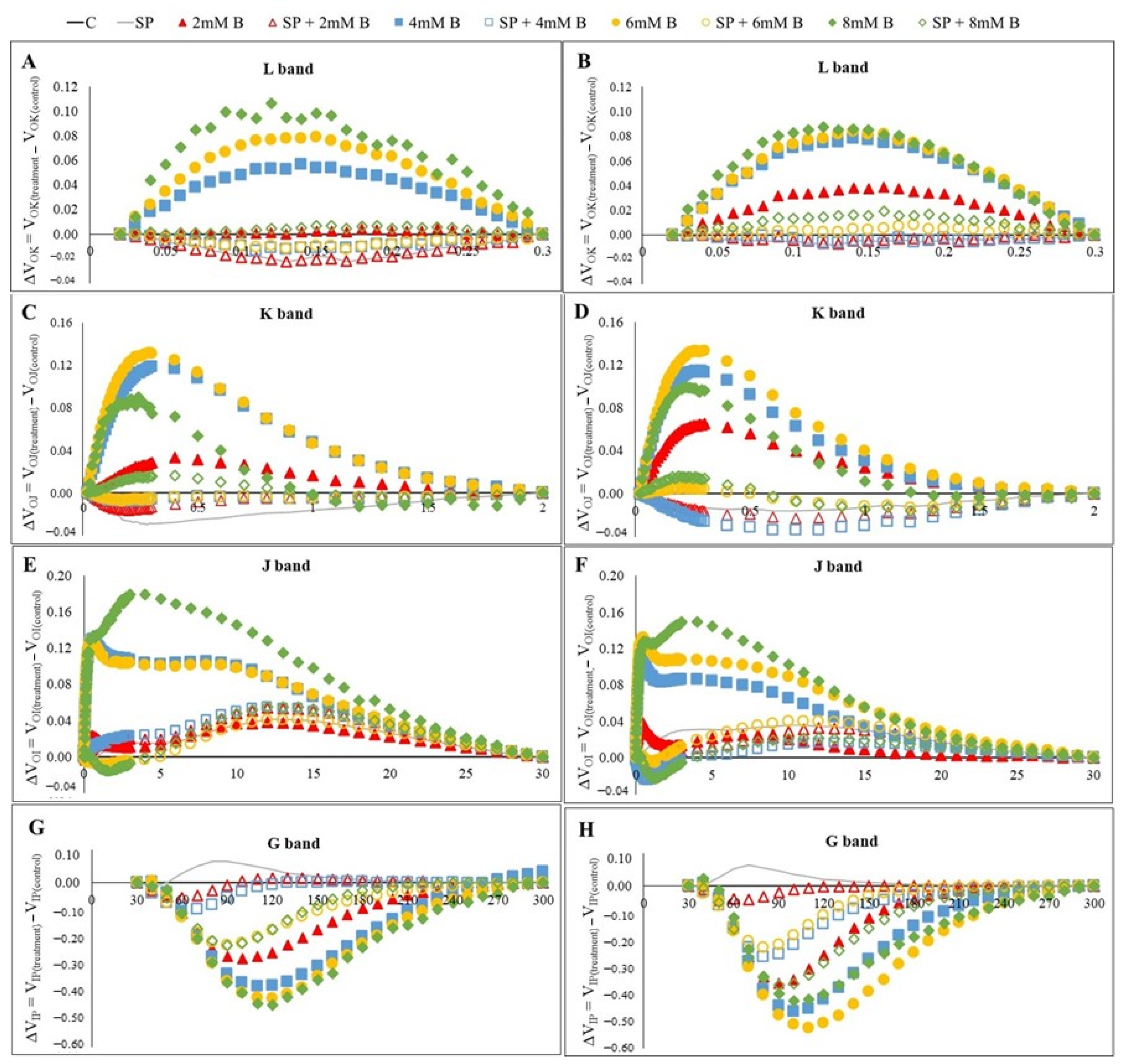
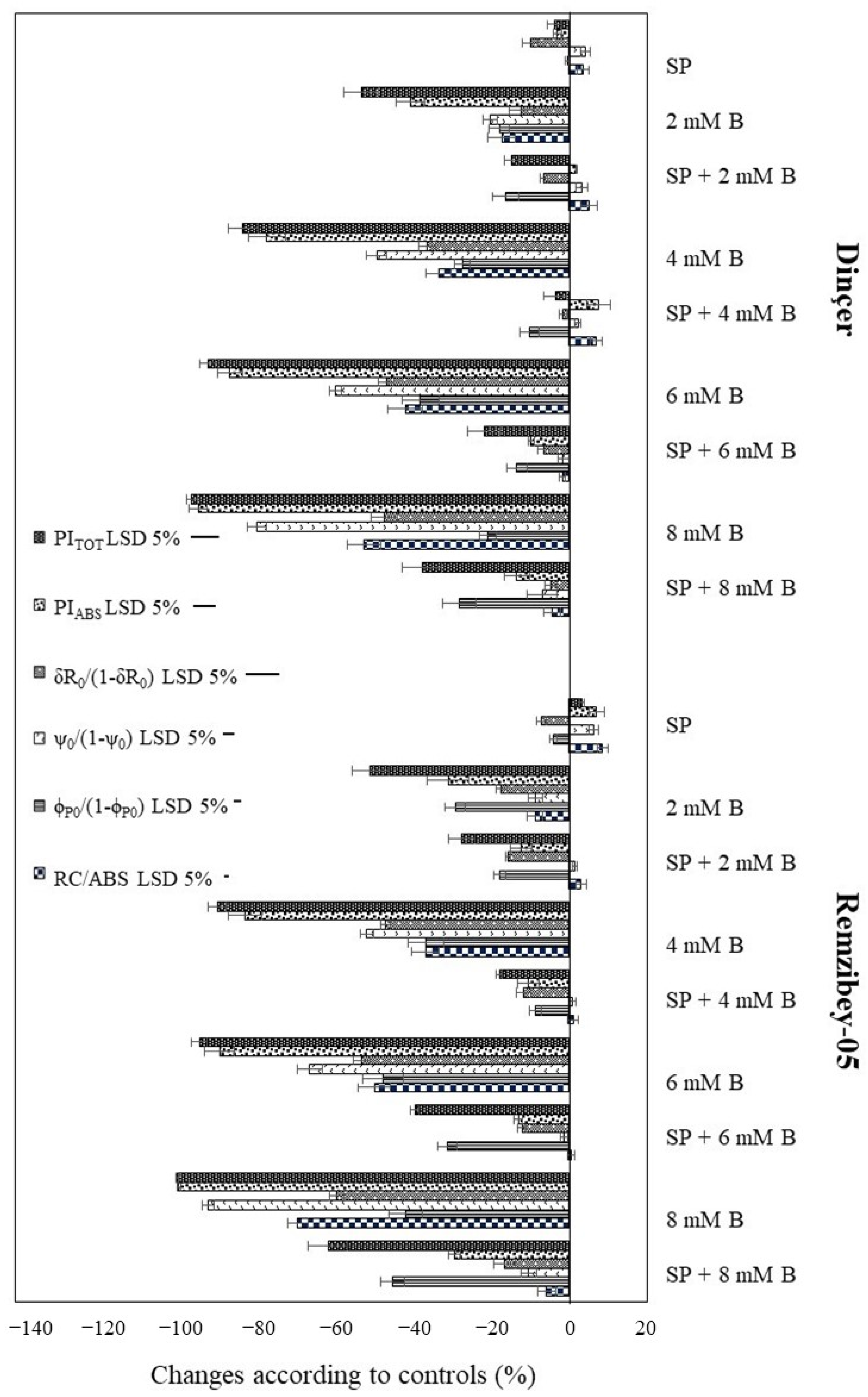
2.3. Chlorophyll a Fluorescence (ChlF) Transient Analysis in Safflower Leaves under B Toxicity with or without SP
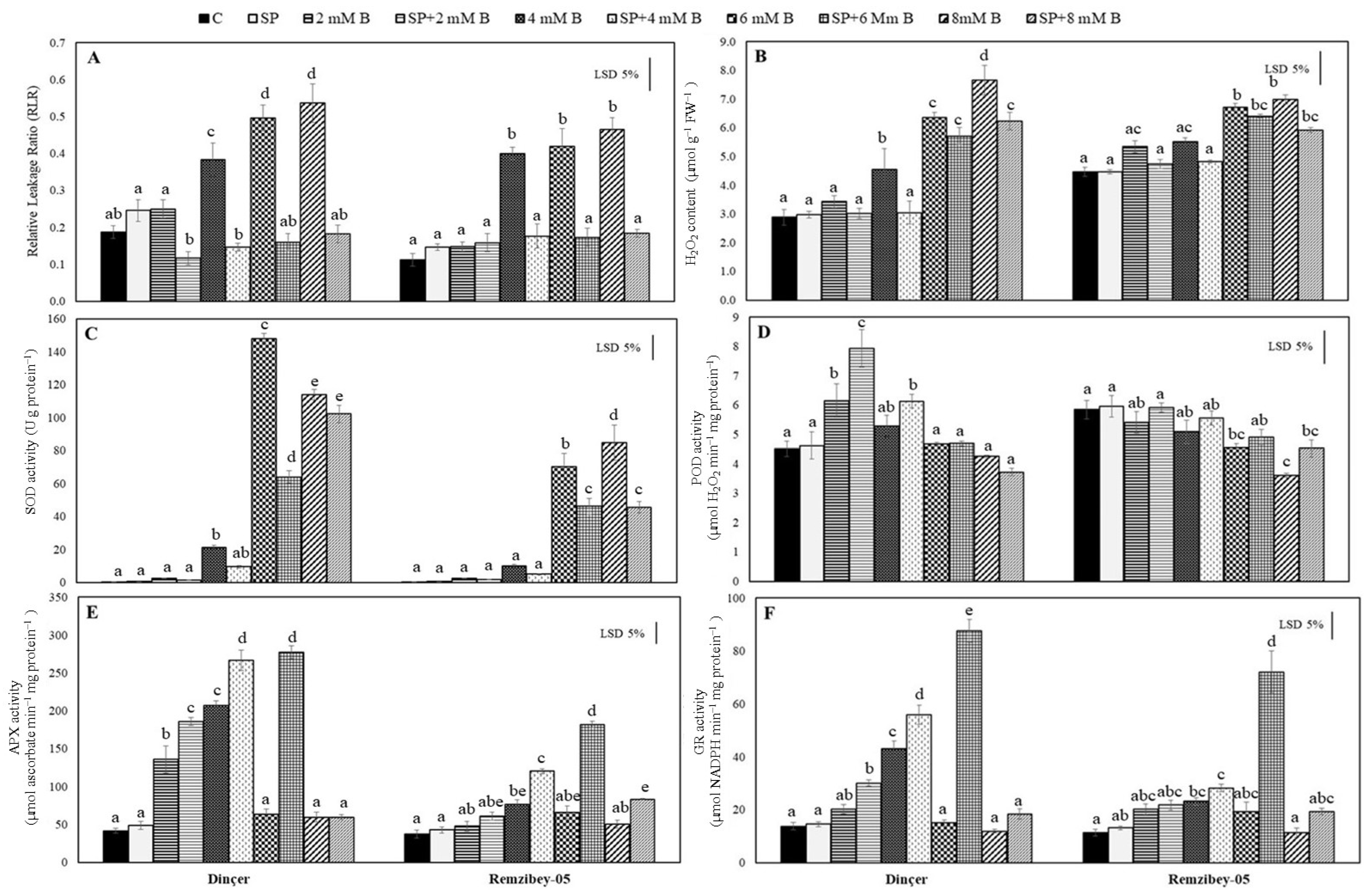
2.4. Alteration of Biochemical Stress Indicators under Toxic B with or without SP
3. Materials and Methods
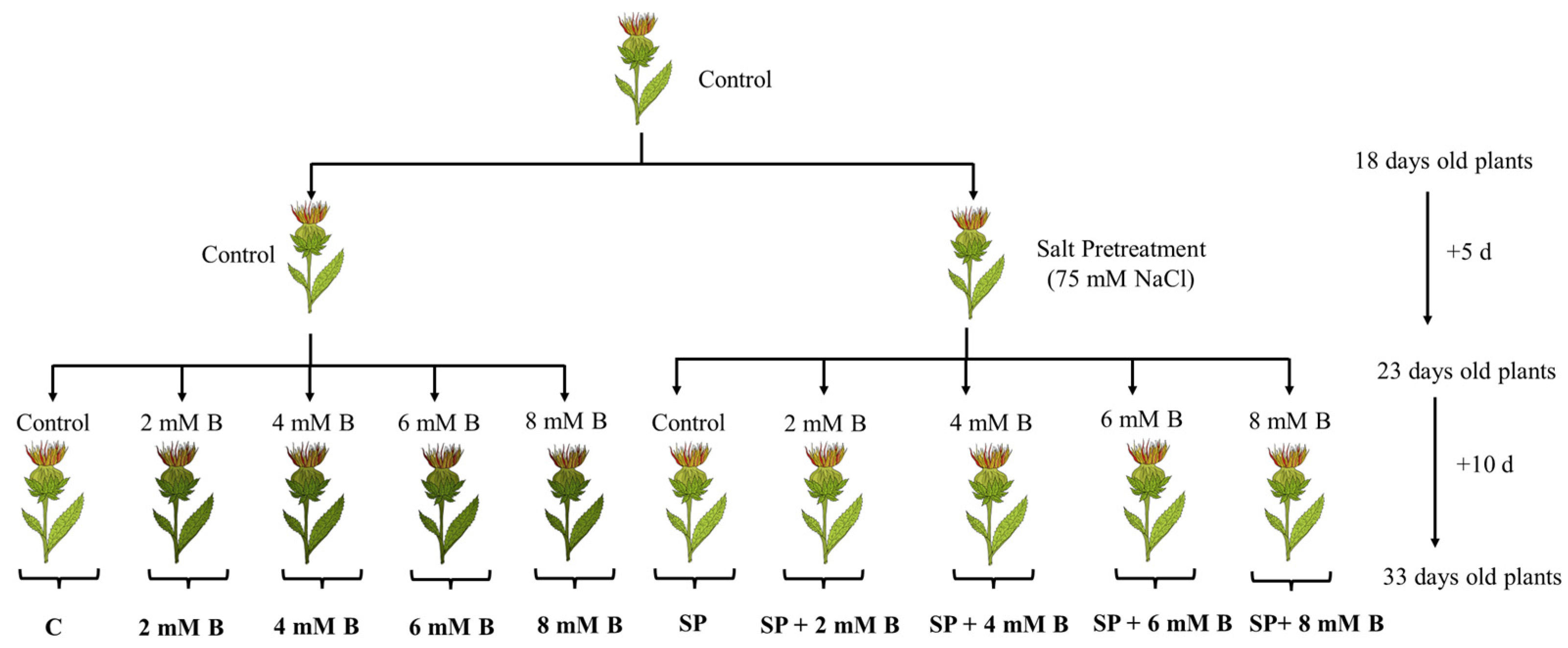
3.1. Plant Materials, Growth, and Treatment Conditions
3.2. Growth Parameters, Necrosis Area, and Water Content of Leaves
3.3. B Contents and Pigment Analysis
3.4. Polyphasic ChlF Measurement
3.5. Relative Leakage Ratio, H2O2 Content, and Antioxidant Enzyme Activities
3.6. Statistical Data Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siddiqui, M.H.; Al-Whaibi, M.H.; Sakran, A.M.; Ali, H.M.; Basalah, M.O.; Faisal, M.; Alatar, A.; Al-Amri, A.A. Calcium-induced amelioration of boron toxicity in radish. J. Plant Growth Regul. 2013, 32, 61–71. [Google Scholar] [CrossRef]
- Farag, M.; Najeeb, U.; Yang, J.; Hu, Z.; Fang, Z.M. Nitric oxide protects carbon assimilation process of watermelon from boron-induced oxidative injury. Plant Physiol. Biochem. 2017, 111, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Brdar-Jokanović, M. Boron toxicity and deficiency in agricultural plants. Int. J. Mol. Sci. 2020, 21, 1424. [Google Scholar] [CrossRef]
- Nable, R.O.; Bañuelos, G.S.; Paull, J.G. Boron toxicity. Plant Soil 1997, 193, 181–198. [Google Scholar] [CrossRef]
- Landi, M.; Margaritopoulou, T.; Papadakis, I.E.; Araniti, F. Boron toxicity in higher plants: An update. Planta 2019, 250, 1011–1032. [Google Scholar] [CrossRef]
- García-Sánchez, F.; Simón-Grao, S.; Martínez-Nicolás, J.J.; Alfosea-Simón, M.; Liu, C.; Chatzissavvidis, C.; Pérez-Pérez, J.G.; Cámara-Zapata, J.M. Multiple stresses occurring with boron toxicity and deficiency in plants. J. Hazard. Mater. 2020, 397, 122713. [Google Scholar] [CrossRef]
- Aydin, M.; Tombuloglu, G.; Sakcali, M.S.; Hakeem, K.R.; Tombuloglu, H. Boron alleviates drought stress by enhancing gene expression and antioxidant enzyme activity. J. Plant Nutr. Soil Sci. 2019, 19, 545–555. [Google Scholar] [CrossRef]
- Hua, T.; Zhang, R.; Sun, H.; Liu, C. Alleviation of boron toxicity in plants: Mechanisms and approaches. Crit. Rev. Environ. Sci. Technol. 2021, 51, 2975–3015. [Google Scholar] [CrossRef]
- Princi, M.P.; Lupini, A.; Araniti, F.; Longo, C.; Mauceri, A.; Sunseri, F.; Abenavoli, M.R. Boron Toxicity and Tolerance in Plants: Recent Advances and Future Perspectives. In Plant Metal Interaction; Ahmad, P., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 115–147. [Google Scholar] [CrossRef]
- Pandey, A.N.; Verma, P. Boron deficiency and toxicity and their tolerance in plants: A review. J. Global Biosci. 2017, 6, 4958–4965. [Google Scholar]
- Choudhary, S.; Zehra, A.; Mukarram, M.; Wani, K.I.; Naeem, M.; Khan, M.M.A.; Aftab, T. Salicylic acid-mediated alleviation of soil boron toxicity in Mentha arvensis and Cymbopogon flexuosus: Growth, antioxidant responses, essential oil contents and components. Chemosphere 2021, 276, 130153. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef]
- Sabagh, A.E.L.; Hossain, A.; Barutçular, C.; Iqbal, M.A.; Islam, M.S.; Fahad, S.; Sytar, O.; Çiğ, F.; Meena, R.S.; Erman, M. Consequences of Salinity Stress on the Quality of Crops and Its Mitigation Strategies for Sustainable Crop Production: An Outlook of Arid and Semi-Arid Regions. In Environment, Climate, Plant and Vegetation Growth; Fahad, S., Hasanuzzaman, M., Alam, M., Ullah, H., Saeed, M., Ali Khan, I., Adnan, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 503–533. [Google Scholar] [CrossRef]
- Kayıhan, D.S.; Kayıhan, C.; Çiftçi, Y.Ö. Excess boron responsive regulations of antioxidative mechanism at physio-biochemical and molecular levels in Arabidopsis thaliana. Plant Physiol. Biochem. 2016, 109, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Dai, Z.; Xia, J.; Chang, C.; Sun, H. Combined effect of salt and drought on boron toxicity in Puccinellia tenuiflora. Ecotoxicol. Environ. Saf. 2018, 157, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Khan, M.K.; Hakki, E.E.; Gezgin, S.; Hamurcu, M. Combined boron toxicity and salinity stress—An insight into its interaction in plants. Plants 2019, 8, 364. [Google Scholar] [CrossRef] [PubMed]
- Yermiyahu, U.; Ben-Gal, A.; Keren, R.; Reid, R.J. Combined effect of salinity and excess boron on plant growth and yield. Plant Soil 2008, 304, 73–87. [Google Scholar] [CrossRef]
- Bañuelos, G.S.; LeDuc, D.; Johnson, J. Evaluating the tolerance of young hybrid poplar trees to recycled waters high in salinity and boron. Int. J. Phytoremediation 2010, 12, 419–439. [Google Scholar] [CrossRef]
- Bañón, S.; Miralles, J.; Ochoa, J.; Sánchez-Blanco, M.J. The effect of salinity and high boron on growth, photosynthetic activity and mineral contents of two ornamental shrubs. Hortic. Sci. 2012, 39, 188–194. [Google Scholar] [CrossRef]
- Karimi, S.; Tavallali, V. Interactive effects of soil salinity and boron on growth, mineral composition and CO2 assimilation of pistachio seedlings. Acta Physiol. Plant. 2017, 39, 242. [Google Scholar] [CrossRef]
- Barua, D.; Mishra, A.; Kirti, P.B.; Barah, P. Identifying signal-crosstalk mechanism in maize plants during combined salinity and boron stress using integrative systems biology approaches. Biomed Res. Int. 2022, 2022, 1027288. [Google Scholar] [CrossRef]
- Ekmekçi, Y.; Çulha Erdal, Ş.; Balkan Nalçaiyi, A.S.; Çiçek, N. The acquisition of boron tolerance by salt pretreatment in two sunflower cultivars. Turk. J. Bot. 2020, 44, 153–166. [Google Scholar] [CrossRef]
- Gecgel, U.; Demirci, M.; Esendal, E.; Tasan, M. Fatty Acid composition of the oil from developing seeds of different varieties of safflower (Carthamus tinctorius L.). J. Am. Oil Chem. Soc. 2007, 84, 47–54. [Google Scholar] [CrossRef]
- Mihaela, P.; Josef, R.; Monica, N.; Rudolf, Z. Perspectives of safflower oil as biodiesel source for south eastern europe (comparative study: Safflower, soybean and rapeseed). Fuel 2013, 111, 114–119. [Google Scholar] [CrossRef]
- Erdal, Ş.Ç.; Çakırlar, H. Impact of salt stress on photosystem II efficiency and antioxidant enzyme activities of safflower (Carthamus tinctorius L.) cultivars. Turk. J. Biol. 2014, 38, 549–560. [Google Scholar] [CrossRef]
- Çulha Erdal, Ş.; Eyidoğan, F.; Ekmekci, Y. Comparative physiological and proteomic analyses of cultivated and wild safflower response to drought stress and rewatering. Physiol. Mol. Biol. Plants 2021, 27, 281–295. [Google Scholar] [CrossRef]
- Day, S.; Çıkılı, Y.; Aasim, M. Screening of three safflower (Carthamus tinctorius L.) cultivars under boron stress. Acta Sci. Pol. Hortorum Cultus. 2017, 16, 109–116. [Google Scholar] [CrossRef]
- Ashagre, H.; Hamza, I.A.; Fita, U.; Estifanos, E. Boron toxicity on seed germination and seedling growth of safflower (Car-thamus tinctorius L.). Herald J. Agric. Food Sci. Res. 2014, 3, 001–006. [Google Scholar]
- Sulus, S.; Leblebici, S. The effect of boric acid application on ecophysiological characteristics of safflower varieties (Carthamus tinctorius L.). Fresenius Environ. Bull. 2020, 29, 8177–8185. [Google Scholar]
- Choi, E.Y.; Kolesik, P.; McNeill, A.N.N.; Collins, H.; Zhang, Q.; Huynh, B.L.; Graham, R.; Stangoulis, J. The mechanism of boron tolerance for maintenance of root growth in barley (Hordeum vulgare L.). Plant Cell Environ. 2007, 30, 984–993. [Google Scholar] [CrossRef]
- Aquea, F.; Federici, F.; Moscoso, C.; Vega, A.; Jullian, P.; Haseloff, J.; Arce-Johnson, P. A molecular framework for the inhibition of Arabidopsis root growth in response to boron toxicity. Plant Cell Environ. 2012, 35, 719–734. [Google Scholar] [CrossRef]
- Çelik, H.; Turan, M.A.; Aşık, B.B.; Öztüfekçi, S.; Katkat, V.A. Effects of soil-applied materials on the dry weight and boron uptake of maize shoots (Zea mays L.) under high boron conditions. Commun. Soil Sci. Plant Anal. 2019, 50, 811–826. [Google Scholar] [CrossRef]
- Papadakis, I.E.; Tsiantas, P.I.; Tsaniklidis, G.; Landi, M.; Psychoyou, M.; Fasseas, C. Changes in sugar metabolism associated to stem bark thickening partially assist young tissues of Eriobotrya japonica seedlings under boron stress. J. Plant Physiol. 2018, 231, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Lu, X.; Riaz, M.; Yan, L.; Jiang, C. Boron toxicity induced specific changes of cell ultrastructure and architecture of components in leaf center and tip of trifoliate orange [Poncirus trifoliata (L.) Raf.]. J. Environ. Manag. 2019, 246, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Macho-Rivero, M.A.; Herrera-Rodríguez, M.B.; Brejcha, R.; Schäffner, A.R.; Tanaka, N.; Fujiwara, T.; Gonzalez-Fontes, A.; Camacho-Cristobal, J.J. Boron toxicity reduces water transport from root to shoot in Arabidopsis plants. Evidence for a reduced transpiration rate and expression of major PIP aquaporin genes. Plant Cell Physiol. 2018, 59, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Metwally, A.M.; Radi, A.A.; El-Shazoly, R.M.; Hamada, A.M. The role of calcium, silicon and salicylic acid treatment in protection of canola plants against boron toxicity stress. J. Plant Res. 2018, 131, 1015–1028. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Akram, N.A.; Ashraf, M. Kinetin and indole acetic acid promote antioxidant defense system and reduce oxidative stress in maize (Zea mays L.) plants grown at boron toxicity. J. Plant Growth Regul. 2018, 37, 1258–1266. [Google Scholar] [CrossRef]
- Kaya, C.; Sarıoğlu, A.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Gibberellic acid-induced generation of hydrogen sulfide alleviates boron toxicity in tomato (Solanum lycopersicum L.) plants. Plant Physiol. Biochem. 2020, 153, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Reid, R. Understanding the boron transport network in plants. Plant Soil 2014, 385, 1–13. [Google Scholar] [CrossRef]
- Pereira, G.L.; Siqueira, J.A.; Batista-Silva, W.; Cardoso, F.B.; Nunes-Nesi, A.; Araújo, W.L. Boron: More than an essential element for land plants? Front. Plant Sci. 2021, 11, 610307. [Google Scholar] [CrossRef]
- Öz, M.T.; Turan, Ö.; Kayıhan, C.; Eyidoğan, F.; Ekmekçi, Y.; Yücel, M.; Öktem, H.A. Evaluation of photosynthetic performance of wheat cultivars exposed to boron toxicity by the JIP fluorescence test. Photosynthetica 2014, 52, 555–563. [Google Scholar] [CrossRef]
- Çatav, Ş.S.; Genç, T.O.; Kesik Oktay, M.; Küçükakyüz, K. Effect of boron toxicity on oxidative stress and genotoxicity in wheat (Triticum aestivum L.). Bull. Environ. Contam. Toxicol. 2018, 100, 502–508. [Google Scholar] [CrossRef]
- Stiles, A.R.; Bautista, D.; Atalay, E.; Babaoglu, M.; Terry, N. Mechanisms of boron tolerance and accumulation in plants: A physiological comparison of the extremely boron-tolerant plant species, Puccinellia distans, with the moderately boron-tolerant Gypsophila arrostil. Environ. Sci. Technol. 2010, 44, 7089–7095. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Sun, Q.; Dong, P.; Ma, C.; Sun, H.; Liu, C. Jasmonic acid alleviates boron toxicity in Puccinellia tenuiflora, a promising species for boron phytoremediation. Plant Soil 2019, 445, 397–407. [Google Scholar] [CrossRef]
- Chen, Z.; Taylor, A.A.; Astor, S.R.; Xin, J.; Terry, N. Removal of boron from wastewater: Evaluation of seven poplar clones for B accumulation and tolerance. Chemosphere 2017, 167, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Usman, K.; Al-Ghouti, M.A.; Abu-Dieyeh, M.H. The assessment of cadmium, chromium, copper, and nickel tolerance and bioaccumulation by shrub plant Tetraena qataranse. Sci. Rep. 2019, 9, 5658. [Google Scholar] [CrossRef]
- Miwa, K.; Takano, J.; Omori, H.; Seki, M.; Shinozaki, K.; Fujiwara, T. Plants tolerant of high boron levels. Science 2007, 318, 1417. [Google Scholar] [CrossRef]
- Sutton, T.; Baumann, U.; Hayes, J.; Collins, N.C.; Shi, B.J.; Schnurbusch, T.; Hay, A.; Mayo, G.; Pallotta, M.; Tester, M.; et al. Boron-toxicity tolerance in barley arising from efflux transporter amplification. Science 2007, 318, 1446–1449. [Google Scholar] [CrossRef]
- Oikonomou, A.; Ladikou, E.V.; Chatziperou, G.; Margaritopoulou, T.; Landi, M.; Sotiropoulos, T.; Araniti, F.; Papadakis, I.E. Boron excess imbalances root/shoot allometry, photosynthetic and chlorophyll fluorescence parameters and sugar metabolism in apple plants. Agronomy 2019, 9, 731. [Google Scholar] [CrossRef]
- Karimi, S.; Tavallali, V.; Wirthensohn, M. Boron amendment improves water relations and performance of Pistacia vera under salt stress. Sci. Hortic. 2018, 241, 252–259. [Google Scholar] [CrossRef]
- Kaya, C.; Sarioğlu, A.; Akram, N.A.; Ashraf, M. Thiourea-mediated nitric oxide production enhances tolerance to boron toxicity by reducing oxidative stress in bread wheat (Triticum aestivum L.) and durum wheat (Triticum durum Desf.) plants. J. Plant Growth. Regul. 2019, 38, 1094–1109. [Google Scholar] [CrossRef]
- Arslan, Ö. Boron toxicity impacts on photosystem II photochemical efficiency of sage (Salvia officinalis). Bot. Serb. 2022, 46, 113–119. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the Chlorophyll a Fluorescence Transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Govindjee, G., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar]
- Stirbet, A.; Lazár, D.; Papageorgiou, G.C. Chlorophyll a Fluorescence in Cyanobacteria: Relation to Photosynthesis. In Cyanobacteria; Mishra, A.K., Tiwari, D.N., Rai, A.N., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 79–130. [Google Scholar]
- Padhi, B.; Chauhan, G.; Kandoi, D.; Stirbet, A.; Tripathy, B.C.; Govindjee, G. A comparison of chlorophyll fluorescence transient measurements, using Handy PEA and FluorPen fluorometers. Photosynthetica 2021, 59, 399–408. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestič, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Lukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plant under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef]
- Han, S.; Tang, N.; Jiang, H.X.; Yang, L.T.; Yan, L.; Chen, L.S. CO2 assimilation, photosystem II photochemistry, carbohydrate metabolism and antioxidant system of citrus leaves in response to boron stress. Plant Sci. 2009, 176, 143–153. [Google Scholar] [CrossRef]
- Goltsev, V.N.; Kalaji, H.M.; Paunov, M.; Bąba, W.; Horaczek, T.; Mojski, J.; Kociel, H.; Allakhverdiev, S.I. Variable chlorophyll fluorescence and its use for assessing physiological condition of plant photosynthetic apparatus. Russ. J. Plant Physiol. 2016, 63, 869–893. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Dangre, D.; Rai, M. Biophysical Phenomics Reveals Functional Building Blocks of Plants Systems Biology: A Case Study for the Evaluation of the Impact of Mycorrhization with Piriformospora Indica. In Advanced Techniques in Soil Microbiology; Varma, A., Oelmüller, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 319–341. [Google Scholar]
- Yusuf, M.D.; Kumar, D.; Rajwanshi, R.; Strasser, R.J.; Tsimilli-Michael, M.; Govindjee; Sarin, N.B. Overexpression of γ-tocopherol methyl transferase gene in transgenic Brassica juncea plants alleviates abiotic stress: Physiological and chlorophyll a fluorescence measurements. Biochim. Biophys. Acta Bioenerg. 2010, 1797, 1428–1438. [Google Scholar] [CrossRef] [PubMed]
- Albert, K.R.; Mikkelsen, T.N.; Michelsen, A.; Ro-Poulsen, H.; van der Linden, L. Interactive effects of drought, elevated CO2 and warming on photosynthetic capacity and photosystem performance in temperate heath plants. J. Plant Physiol. 2011, 168, 1550–1561. [Google Scholar] [CrossRef]
- Koutra, E.; Chondrogiannis, C.; Grammatikopoulos, G. Variability of the photosynthetic machinery tolerance when imposed to rapidly or slowly imposed dehydration in native Mediterranean plants. Photosynthetica 2022, 60, 88–101. [Google Scholar] [CrossRef]
- Moustafa-Farag, M.; Mohamed, H.I.; Mahmoud, A.; Elkelish, A.; Misra, A.N.; Guy, K.M.; Kamran, M.; Ai, S.; Zhang, M. Salicylic acid stimulates antioxidant defense and osmolyte metabolism to alleviate oxidative stress in watermelons under excess boron. Plants 2020, 9, 724. [Google Scholar] [CrossRef]
- Oliveira, K.R.; Junior, J.P.S.; Bennett, S.J.; Checchio, M.V.; de Cássia Alves, R.; Felisberto, G.; de Mello Prado, R.; Gratao, P.L. Exogenous silicon and salicylic acid applications improve tolerance to boron toxicity in field pea cultivars by intensifying antioxidant defence systems. Ecotoxicol. Environ. Saf. 2020, 201, 110778. [Google Scholar] [CrossRef]
- Cervilla, L.M.; Blasco, B.; Rios, J.J.; Rosales, M.A.; Sánchez-Rodríguez, E.; Rubio-Wilhelmi, M.M.; Romero, L.; Ruiz, J.M. Parameters symptomatic for boron toxicity in leaves of tomato plants. J. Bot. 2012, 2012, 726206. [Google Scholar] [CrossRef]
- Nawaz, M.; Ishaq, S.; Ishaq, H.; Khan, N.; Iqbal, N.; Ali, S.; Rizwan, M.; Alsahli, A.A.; Alyemeni, M.N. Salicylic acid improves boron toxicity tolerance by modulating the physio-biochemical characteristics of maize (Zea mays L.) at an early growth stage. Agronomy 2020, 10, 2013. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, L.; Li, Y.; Chen, Q.; Ye, Y.; Zhang, Y.; Luo, Y.; Sun, B.; Wang, X.; Tang, H. Effect of red and blue light on anthocyanin accumulation and differential gene expression in strawberry (Fragaria × ananassa). Molecules 2018, 23, 820. [Google Scholar] [CrossRef] [PubMed]
- Landi, M.; Guidi, L.; Pardossi, A.; Tattini, M.; Gould, K.S. Photoprotection by foliar anthocyanins mitigates effects of boron toxicity in sweet basil (Ocimum basilicum). Planta 2014, 240, 941–953. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Rothstein, S.J. ROS-Induced anthocyanin production provides feedback protection by scavenging ROS and maintaining photosynthetic capacity in Arabidopsis. Plant Signal Behav. 2018, 13, 1364–1377. [Google Scholar] [CrossRef]
- Landi, M.; Degl’Innocenti, E.; Pardossi, A.; Guidi, L. Antioxidant and photosynthetic responses in plants under boron toxicity: A review. Am. J. Agric. Biol. Sci. 2012, 7, 255–270. [Google Scholar] [CrossRef]
- Yousefi, H.; Dalir, N.; Rahnemaie, R.; Babaei, A. The alleviation of salinity-induced stress by using boron in soilless grown rose. J. Plant Nutr. 2020, 43, 526–537. [Google Scholar] [CrossRef]
- Surgun, Y.; Çöl, B.; Bürün, B. 24-Epibrassinolide ameliorates the effects of boron toxicity on Arabidopsis thaliana (L.) Heynh by activating an antioxidant system and decreasing boron accumulation. Acta Physiol. Plant. 2016, 38, 71. [Google Scholar] [CrossRef]
- Song, B.; Hao, X.; Wang, X.; Yang, S.; Dong, Y.; Ding, Y. Boron stress inhibits beet (Beta vulgaris L.) growth through influencing endogenous hormones and oxidative stress response. Soil Sci. Plant Nutr. 2019, 65, 346–352. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water culture method for growing plants without soil. Circ. Calif. Agr. Exp. Sta. 1950, 347, 32. [Google Scholar]
- Farrant, J.M. A comparison of mechanisms of desiccation tolerance among three angiosperm resurrection plant species. Plant Ecol. 2000, 151, 29–39. [Google Scholar] [CrossRef]
- Yoon, J.; Cao, X.; Zhou, Q.; Ma, L.Q. Accumulation of Pb, Cu, and Zn in native plants growing on a contaminated Florida site. Sci. Total Environ. 2006, 368, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Roccotiello, E.; Manfredi, A.; Drava, G.; Minganti, V.; Mariotti, M.G.; Berta, G.; Cornara, L. Zinc tolerance and accumulation in the ferns Polypodium cambricum L. and Pteris vittata L. Ecotox. Environ. Safe. 2010, 73, 1264–1271. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Mancinelli, A.L.; Yang, C.P.H.; Lindquist, P.; Anderson, O.; Rabino, I. Photocontrol of anthocyanin synthesis: The action of streptomycin on the synthesis of chlorophyll and anthocyanin. Plant Physiol. 1975, 55, 251–257. [Google Scholar] [CrossRef]
- Mirecki, R.M.; Teramura, A.H. Effects of ultraviolet-b irradiance on soybean 1: V. The dependence of plant sensitivity on the photosynthetic photon flux density during and after leaf expansion. Plant Physiol. 1984, 74, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.J.; Tsimilli-Michael, M.; Qiang, S.; Goltsev, V. Simultaneous in vivo recording of prompt and delayed fluorescence and 820-nm reflection changes during drying and after rehydration of the resurrection plant Haberlea rhodopensis. Biochim. Biophys. Acta Bioenerg. 2010, 1797, 1313–1326. [Google Scholar] [CrossRef]
- Redmann, R.E.; Haraldson, J.; Gusta, L.V. Leakage of UV-absorbing substances as a measure of salt injury in leaf tissue of woody species. Physiol. Plant. 1986, 67, 87–91. [Google Scholar] [CrossRef]
- Esterbauer, H.; Cheeseman, K.H. Determination of aldehydic lipid peroxidation products: Malonaldehyde and 4-hydroxynonenal. Methods Enzymol. 1990, 186, 407–421. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgramquantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Wang, S.Y.; Jiao, H.J.; Faust, M. Changes in ascorbate, glutathione, and related enzyme activities during thidiazuron-induced bud break of apple. Physiol. Plant. 1991, 82, 231–236. [Google Scholar] [CrossRef]
- Rao, M.V.; Hale, B.A.; Ormrod, D.P. Amelioration of ozone-induced oxidative damage in wheat plants grown under high carbon dioxide (role of antioxidant enzymes). Plant Physiol. 1995, 109, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Pütter, J. Peroxidases. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Academic Press: New York, NY, USA, 1974; Volume II, pp. 685–690. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivars | Treatment | B Content of Leaves | B Content of Roots | TF | BF | BCF |
|---|---|---|---|---|---|---|
| Dinçer | C | 150.0 * ± 1.7 a | 39.5 ± 2.8 a | 3.88 ± 0.24 a | 277.72 ± 3.20 a | 73.14 ± 5.25 a |
| SP | 98.3 ± 3.2 a | 26.5 ± 0.5 a | 3.72 ± 0.19 a | 182.10 ± 5.87 b | 49.13 ± 0.96 b | |
| 2 mM B | 1585.4 ± 12.4 b | 118.5 ± 5.7 b | 13.51 ± 0.55 bc | 73.37 ± 0.57 c | 5.48 ± 0.27 c | |
| SP + 2 mM B | 403.2 ± 14.1 c | 81.7 ± 0.8 c | 4.93 ± 0.15 a | 18.66 ± 0.65 d | 3.78 ± 0.04 df | |
| 4 mM B | 2283.0 ± 16.8 d | 149.7 ± 2.3 d | 15.26 ± 0.13 b | 52.80 ± 0.39 e | 3.46 ± 0.05 dg | |
| SP + 4 mM B | 577.4 ± 12.0 e | 99.4 ± 2.8 bc | 5.82 ± 0.05 a | 13.35 ± 0.28 f | 2.30 ± 0.07 e | |
| 6 mM B | 2666.8 ± 120.8 f | 290.8 ± 5.9 e | 9.16 ± 0.32 ac | 41.12 ± 1.86 g | 4.48 ± 0.09 f | |
| SP + 6 mM B | 1168.6 ± 66.8 g | 176.9 ± 7.2 f | 6.73 ± 0.65 a | 18.02 ± 1.03 d | 2.73 ± 0.11 g | |
| 8 mM B | 2769.2 ± 25.7 f | 312.0 ± 2.3 g | 8.88 ± 0.10 ac | 32.02 ± 0.30 h | 3.61 ± 0.03 dg | |
| SP + 8 mM B | 1475.2 ± 28.5 b | 248.4 ± 2.3 h | 5.95 ± 0.17 a | 17.06 ± 0.33 d | 2.87 ± 0.03 g | |
| Remzibey-05 | C | 137.4 ± 1.8 a | 30.8 ± 0.9 a | 4.48 ± 0.08 a | 254.52 ± 3.41 a | 56.94 ± 1.67 a |
| SP | 74.5 ± 1 a | 17.7 ± 1.0 a | 4.29 ± 0.29 a | 138.02 ± 1.94 b | 32.76 ± 1.78 b | |
| 2 mM B | 1272.5 ± 23.1 b | 151.4 ± 1.0 be | 8.41 ± 0.11 ab | 58.88 ± 1.07 c | 7.00 ± 0.05 c | |
| SP + 2 mM B | 332.0 ± 2.3 c | 74.4 ± 1.6 c | 4.48 ± 0.13 a | 15.36 ± 0.11 d | 3.44 ± 0.08 df | |
| 4 mM B | 2315.5 ± 14.5 d | 171.6 ± 1.9 de | 13.5 ± 0.07 b | 53.55 ± 0.34 e | 3.97 ± 0.04 d | |
| SP + 4 mM B | 639.1 ± 10.2 e | 157.5 ± 7.4 e | 4.10 ± 0.18 a | 14.78 ± 0.24 d | 3.64 ± 0.17 df | |
| 6 mM B | 2554.3 ± 58.8 f | 333.3 ± 1.6 f | 7.67 ± 0.19 ab | 39.38 ± 0.91 f | 5.14 ± 0.02 e | |
| SP + 6 mM B | 1048.1 ± 2.9 g | 184.0 ± 9.0 d | 5.77 ± 0.29 a | 16.16 ± 0.05 d | 2.84 ± 0.14 fg | |
| 8 mM B | 3316.6 ± 53.4 h | 350.6 ± 20.8 f | 9.58 ± 0.42 ab | 38.35 ± 0.62 f | 4.05 ± 0.24 d | |
| SP + 8 mM B | 1427.2 ± 22.2 ı | 210.6 ± 8.5 g | 6.85 ± 0.38 a | 16.50 ± 0.26 d | 2.44 ± 0.10 g | |
| LSD 5% | 120 | 20.0 | 5.83 | 4.16 | 0.89 |
| Cultivars | Treatment | Chl a+b Content | Carotenoid Content | Anthocyanin Content | Flavonoid Content |
|---|---|---|---|---|---|
| Dinçer | C | 534 × 10−4 * ± 0.0 a | 87 × 10−4 ± 0.0 ab | 49 × 10−6 ± 0.0 a | 100 ± 0 a |
| SP | 551 × 10−4 ± 0.0 a | 86 × 10−4 ± 0.0 ab | 52 × 10−6 ± 0.0 a | 98 ± 3 a | |
| 2 mM B | 547 × 10−4 ± 0.0 a | 96 × 10−4 ± 0.0 a | 50 × 10−6 ± 0.0 a | 105 ± 8 a | |
| SP + 2 mM B | 535 × 10−4 ± 0.0 a | 88 × 10−4 ± 0.0 ab | 55 × 10−6 ± 0.0 a | 111 ± 3 a | |
| 4 mM B | 436 × 10−4 ± 0.0 bd | 66 × 10−4 ± 0.0 bc | 441 × 10−6 ± 0.0 bd | 320 ± 17 b | |
| SP + 4 mM B | 467 × 10−4 ± 0.0 b | 72 × 10−4 ± 0.0 bc | 539 × 10−6 ± 0.0 ce | 376 ± 8 c | |
| 6 mM B | 248 × 10−4 ± 0.0 c | 55 × 10−4 ± 0.0 cd | 382 × 10−6 ± 0.0 df | 264 ± 4 d | |
| SP + 6 mM B | 383 × 10−4 ± 0.0 d | 62 × 10−4 ± 0.0 c | 607 × 10−6 ± 0.0 e | 472 ± 3 e | |
| 8 mM B | 148 × 10−4 ± 0.0 e | 32 × 10−4 ± 0.0 d | 350 × 10−6 ± 0.0 f | 237 ± 7 d | |
| SP + 8 mM B | 252 × 10−4 ± 0.0 e | 33 × 10−4 ± 0.0 d | 581 × 10−6 ± 0.0 e | 499 ± 6 e | |
| Remzibey-05 | C | 454 × 10−4 ± 0.0 a | 66 × 10−4 ± 0.0 ab | 124 × 10−6 ± 0.0 a | 100 ± 0 a |
| SP | 453 × 10−4 ± 0.0 a | 68 × 10−4 ± 0.0 ab | 124 × 10−6 ± 0.0 a | 119 ± 7 a | |
| 2 mM B | 439 × 10−4 ± 0.0 ab | 75 × 10−4 ± 0.0 ab | 258 × 10−6 ± 0.0 b | 225 ± 5 b | |
| SP + 2 mM B | 465 × 10−4 ± 0.0 a | 89 × 10−4 ± 0.0 a | 468 × 10−6 ± 0.0 c | 412 ± 7 c | |
| 4 mM B | 379 × 10−4 ± 0.0 bd | 68 × 10−4 ± 0.0 ab | 326 × 10−6 ± 0.0 b | 271 ± 17 d | |
| SP + 4 mM B | 459 × 10−4 ± 0.0 a | 75 × 10−4 ± 0.0 ab | 450 × 10−6 ± 0.0 c | 424 ± 8 c | |
| 6 mM B | 298 × 10−4 ± 0.0 cd | 54 × 10−4 ± 0.0 b | 160 × 10−6 ± 0.0 a | 209 ± 8 b | |
| SP + 6 mM B | 455 × 10−4 ± 0.0 a | 63 × 10−4 ± 0.0 b | 456 × 10−6 ± 0.0 c | 500 ± 16 e | |
| 8 mM B | 235 × 10−4 ± 0.0 c | 13 × 10−4 ± 0.0 c | 127 × 10−6 ± 0.0 a | 171 ± 1 f | |
| SP + 8 mM B | 335 × 10−4 ± 0.0 d | 25 × 10−4 ± 0.0 a | 542 × 10−6 ± 0.0 d | 527 ± 1 e | |
| LSD 5% | 64 × 10−4 | 23 × 10−4 | 72 × 10−6 | 33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arslan, Ö.; Çulha Erdal, Ş.; Ekmekçi, Y. Salt Pretreatment-Mediated Alleviation of Boron Toxicity in Safflower Cultivars: Growth, Boron Accumulation, Photochemical Activities, Antioxidant Defense Response. Plants 2022, 11, 2316. https://doi.org/10.3390/plants11172316
Arslan Ö, Çulha Erdal Ş, Ekmekçi Y. Salt Pretreatment-Mediated Alleviation of Boron Toxicity in Safflower Cultivars: Growth, Boron Accumulation, Photochemical Activities, Antioxidant Defense Response. Plants. 2022; 11(17):2316. https://doi.org/10.3390/plants11172316
Chicago/Turabian StyleArslan, Özlem, Şeküre Çulha Erdal, and Yasemin Ekmekçi. 2022. "Salt Pretreatment-Mediated Alleviation of Boron Toxicity in Safflower Cultivars: Growth, Boron Accumulation, Photochemical Activities, Antioxidant Defense Response" Plants 11, no. 17: 2316. https://doi.org/10.3390/plants11172316